
Diversity and Phylogenetics of Freshwater Mussels (Unionidae) from Southern Thailand with the Description of One New Genus and Five New Species-Group Taxa

 , , , , and
, , , , and
Abstract

1. Introduction
2. Materials and Methods
2.1. Data Sampling
2.2. Morphological Studies
2.3. Molecular and Phylogenetic Analyses, Species Delimitation
2.4. Nomenclatural Acts
3. Results
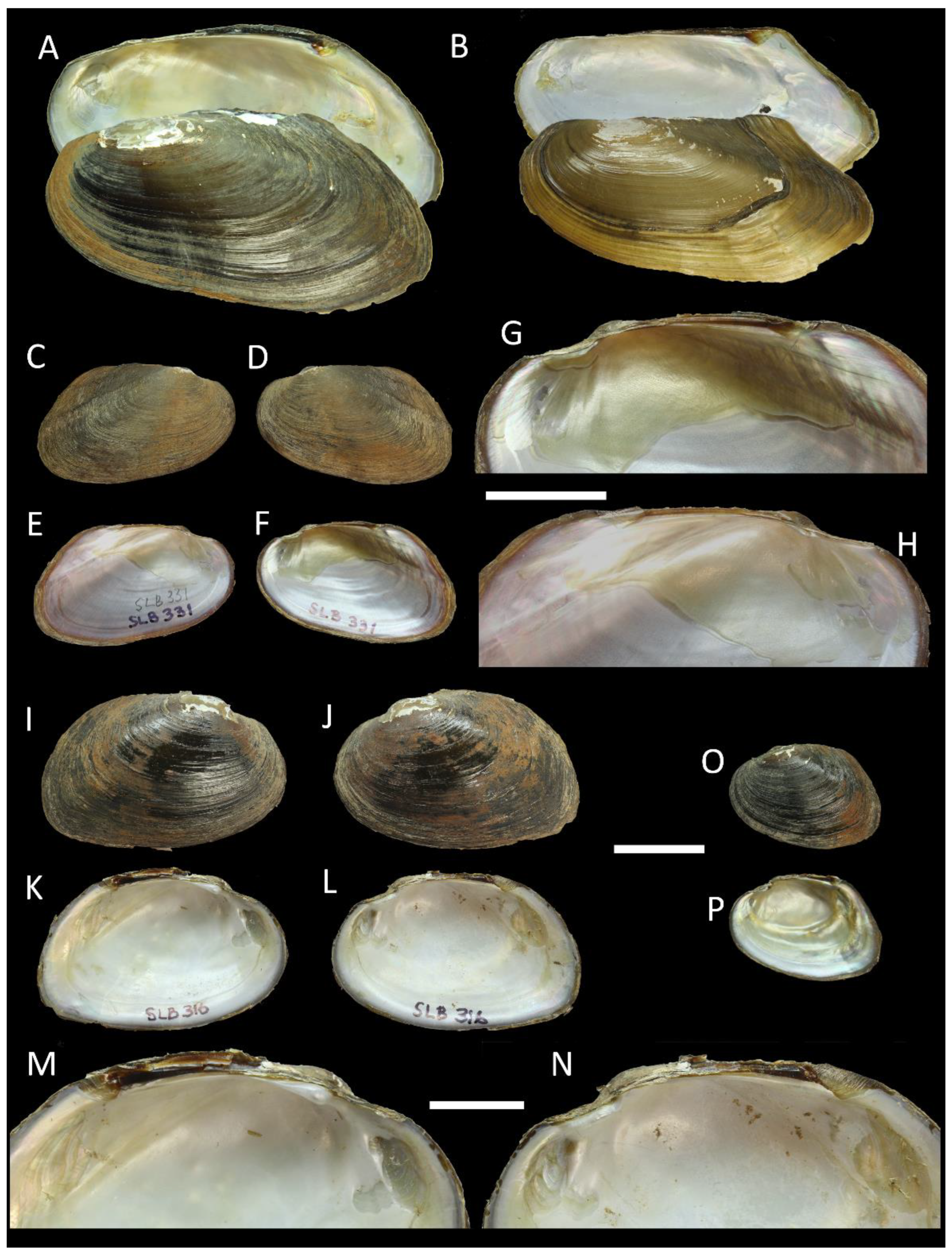
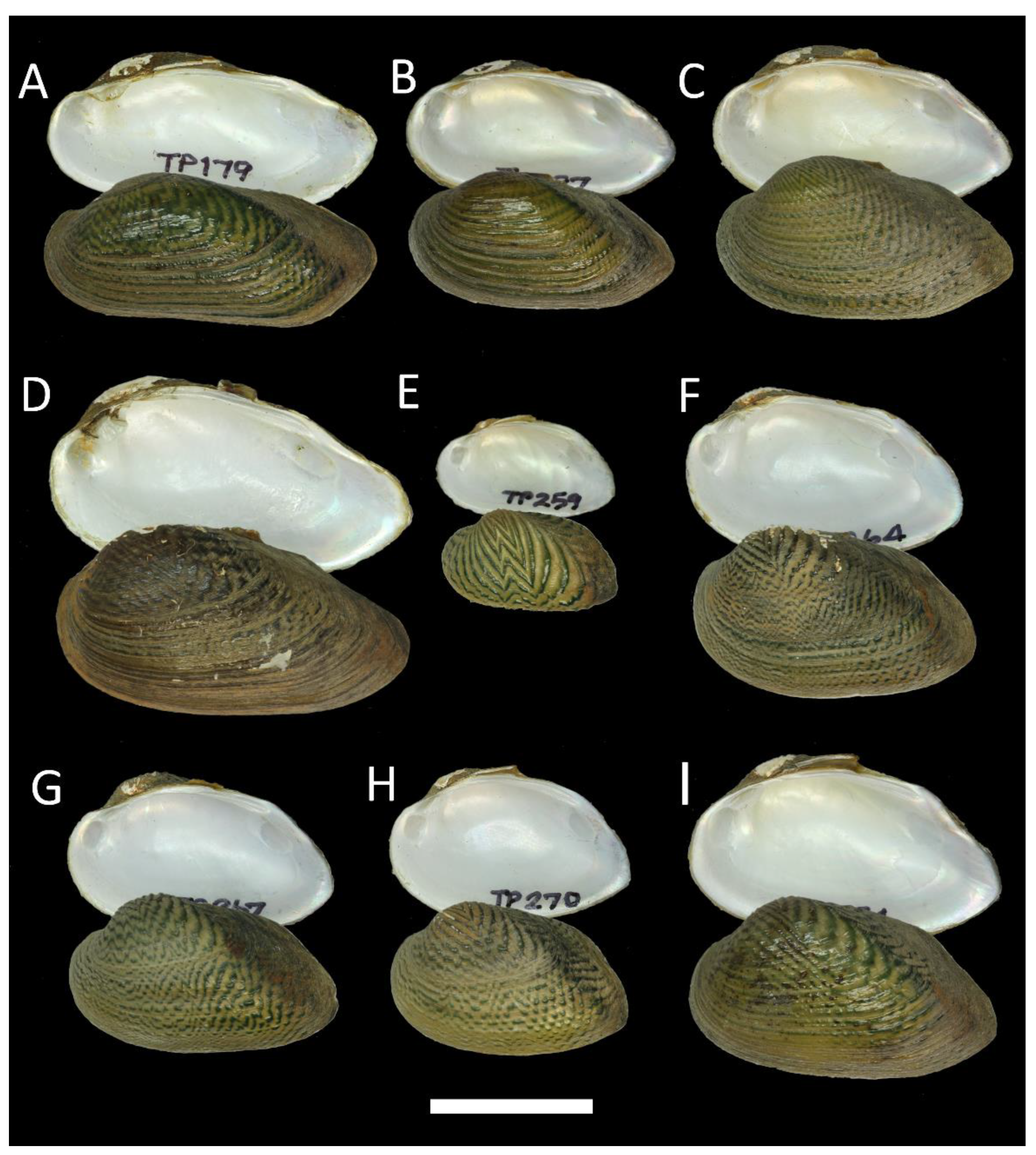
3.1. Morphological Data
3.2. Molecular Phylogeny and Species Delimitation
3.3. Distribution Patterns
3.4. Taxonomy
4. Discussion
4.1. Taxonomic Issues
4.2. Distributional Patterns and Conservation Priorities
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brandt, R.A.M. The non-marine aquatic Mollusca of Thailand. Arch. Molluskenkd. 1974, 105, 1–423. [Google Scholar]
- Zieritz, A.; Bogan, A.E.; Froufe, E.; Klishko, O.; Kondo, T.; Kovitvadhi, U.; Kovitvadhi, S.; Lee, J.H.; Lopes-Lima, M.; Pfeiffer, J.M.; et al. Diversity, biogeography and conservation of freshwater mussels (Bivalvia: Unionida) in East and Southeast Asia. Hydrobiologia 2018, 810, 29–44. [Google Scholar] [CrossRef]
- Bolotov, I.N.; Konopleva, E.S.; Vikhrev, I.V.; Gofarov, M.Y.; Lopes-Lima, M.; Bogan, A.E.; Lunn, Z.; Chan, N.; Win, T.; Aksenova, O.V.; et al. New freshwater mussel taxa discoveries clarify biogeographic division of Southeast Asia. Sci. Rep. 2020, 10, 6616. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffer, J.M.; Graf, D.L.; Cummings, K.S.; Page, L.M. Taxonomic revision of a radiation of Southeast Asian freshwater mussels (Unionidae: Gonideinae: Contradentini+Rectidentini). Invertebr. Syst. 2021, 35, 394–470. [Google Scholar] [CrossRef]
- Liao, P.-C.; Chiang, Y.-C.; Huang, S.; Wang, J.-C. Gene flow of Ceriops tagal (Rhizophoraceae) across the Kra Isthmus in the Thai Malay Peninsula. Bot. Stud. 2009, 50, 193–204. [Google Scholar]
- Parnell, J. The biogeography of the Isthmus of Kra region: A review. Nord. J. Bot. 2013, 31, 1–15. [Google Scholar] [CrossRef]
- Lheknim, V. Annotated Checklist for a Collection of Fishes from Tapi River Basin, south Thailand. Nat. Hist. J. Chulalongkorn Univ. 2004, 4, 83–98. [Google Scholar]
- Iwasaki, S.; Shaw, R. Integrated Lagoon Fisheries Management: Resource Dynamics and Adaptation; Emerald Publisher: Bingley, UK, 2010. [Google Scholar]
- Pfeiffer, J.M.; Graf, D.L. Evolution of bilaterally asymmetrical larvae in freshwater mussels (Bivalvia: Unionoida: Unionidae). Zool. J. Linn. Soc. 2015, 175, 307–318. [Google Scholar] [CrossRef]
- Lopes-Lima, M.; Froufe, E.; Ghamizi, M.; Mock, K.E.; Kebapçı, Ü.; Klishko, O.; Kovitvadhi, S.; Kovitvadhi, U.; Paulo, O.S.; Pfeiffer, J.M.; et al. Phylogeny of the most species-rich freshwater bivalve family (Bivalvia: Unionida: Unionidae): Defining modern subfamilies and tribes. Mol. Phylogenet. Evol. 2017, 10, 174–191. [Google Scholar] [CrossRef]
- Pfeiffer, J.M.; Graf, D.L.; Cummings, K.S.; Page, L.M. Molecular phylogeny and taxonomic revision of two enigmatic freshwater mussel genera (Bivalvia: Unionidae incertae sedis: Harmandia and Unionetta) reveals a diverse clade of Southeast Asian Parreysiinae. J. Molluscan Stud. 2018, 84, 404–416. [Google Scholar] [CrossRef]
- Jeratthitikul, E.; Phuangphong, S.; Sutcharit, C.; Prasankok, P.; Kongim, B.; Panha, S. Integrative taxonomy reveals phenotypic plasticity in the freshwater mussel Contradens contradens (Bivalvia: Unionidae) in Thailand, with a description of a new species. Syst. Biodivers. 2019, 17, 134–147. [Google Scholar] [CrossRef]
- Jeratthitikul, E.; Sucharit, C.; Prasankok, P. Molecular phylogeny of the Indochinese freshwater mussel genus Scabies Haas, 1911 (Bivalvia: Unionidae). Trop. Nat. Hist. 2019, 19, 21–36. Available online: https://li01.tci-thaijo.org/index.php/tnh/article/view/181195 (accessed on 21 July 2022).
- Jeratthitikul, E.; Paphatmethin, S.; Zieritz, A.; Lopes-Lima, M.; Ngor, P.B. Hyriopsis panhai, a new species of freshwater mussel from Thailand (Bivalvia: Unionidae). Raffles Bull. Zool. 2021, 69, 124–136. [Google Scholar] [CrossRef]
- Muanta, S.; Jeratthitikul, E.; Panha, S.; Prasankok, P. Phylogeography of the freshwater bivalve genus Ensidens (Unionidae) in Thailand. J. Molluscan Stud. 2019, 85, 224–231. [Google Scholar] [CrossRef]
- Konopleva, E.S.; Bolotov, I.N.; Pfeiffer, J.M.; Vikhrev, I.V.; Kondakov, A.V.; Gofarov, M.Y.; Tomilova, A.A.; Tanmuangpak, K.; Tumpeesuwan, S. New freshwater mussels from two Southeast Asian genera Bineurus and Thaiconcha (Pseudodontini, Gonideinae, Unionidae). Sci. Rep. 2021, 11, 8244. [Google Scholar] [CrossRef]
- Konopleva, E.S.; Bolotov, I.N.; Vikhrev, I.V.; Gofarov, M.Y.; Kondakov, A.V. An integrative approach underscores the taxonomic status of Lamellidens exolescens, a freshwater mussel from the Oriental tropics (Bivalvia: Unionidae). Syst. Biodivers. 2017, 15, 204–217. [Google Scholar] [CrossRef]
- Konopleva, E.S.; Pfeiffer, J.M.; Vikhrev, I.V.; Kondakov, A.V.; Gofarov, M.Y.; Aksenova, O.V.; Lunn, Z.; Chan, N.; Bolotov, I.N. A new genus and two new species of freshwater mussels (Unionidae) from western Indochina. Sci. Rep. 2019, 9, 4106. [Google Scholar] [CrossRef]
- Bolotov, I.N.; Kondakov, A.V.; Vikhrev, I.V.; Aksenova, O.V.; Bespalaya, Y.V.; Gofarov, M.Y.; Kolosova, Y.S.; Konopleva, E.S.; Spitsyn, V.M.; Tanmuangpak, K.; et al. Ancient river inference explains exceptional Oriental freshwater mussel radiations. Sci. Rep. 2017, 7, 2135. [Google Scholar] [CrossRef]
- Bolotov, I.N.; Vikhrev, I.V.; Kondakov, A.V.; Konopleva, E.S.; Gofarov, M.Y.; Aksenova, O.V.; Tumpeesuwan, S. New taxa of freshwater mussels (Unionidae) from a species-rich but overlooked evolutionary hotspot in Southeast Asia. Sci. Rep. 2017, 7, 11573. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Chernomor, O.; Von Haeseler, A.; Minh, B.Q. Terrace aware data structure for phylogenomic inference from supermatrices. Syst. Biol. 2016, 65, 997–1008. [Google Scholar] [CrossRef] [PubMed]
- Hoang, D.T.; Chernomor, O.; Von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2017, 35, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; Van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarisation in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef]
- Zhang, J.; Kapli, P.; Pavlidis, P.; Stamatakis, A. A general species delimitation method with applications to phylogenetic placements. Bioinformatics 2013, 29, 2869–2876. [Google Scholar] [CrossRef]
- Kapli, P.; Lutteropp, S.; Zhang, J.; Kobert, K.; Pavlidis, P.; Stamatakis, A.; Flouri, T. Multi-rate Poisson tree processes for single-locus species delimitation under maximum likelihood and Markov chain Monte Carlo. Bioinformatics 2017, 33, 1630–1638. [Google Scholar] [CrossRef]
- Puillandre, N.; Brouillet, S.; Achaz, G. ASAP: Assemble species by automatic partitioning. Mol. Ecol. Resour. 2021, 21, 609–620. [Google Scholar] [CrossRef]
- Jeratthitikul, E.; Paphatmethin, S.; Sutcharit, C.; Ngor, P.B.; Inkhavilay, K.; Prasankok, P. Phylogeny and biogeography of Indochinese freshwater mussels in the genus Pilsbryoconcha Simpson, 1900 (Bivalvia: Unionidae) with descriptions of four new species. Sci. Rep. 2022, 12, 20458. [Google Scholar] [CrossRef]
- Lopes-Lima, M.; Hattori, A.; Kondo, T.; Lee, J.H.; Kim, S.K.; Shirai, A.; Hayashi, H.; Usui, T.; Sakuma, K.; Toriya, T.; et al. Freshwater mussels (Bivalvia: Unionidae) from the rising sun (Far East Asia): Phylogeny, systematics, and distribution. Mol. Phylogenet. Evol. 2020, 146, 1–27. [Google Scholar] [CrossRef]
- Pieri, A.M.; Inoue, K.; Johnson, N.A.; Smith, C.H.; Harris, J.L.; Robertson, C.; Randklev, C.R. Molecular and morphometric analyses reveal cryptic diversity with freshwater mussels (Bivalvia: Unionidae) of the western Gulf coastal drainages of the USA. Biol. J. Linn. Soc. 2018, 124, 261–277. [Google Scholar] [CrossRef]
- Smith, C.H.; Johnson, N.A.; Inoue, K.; Doyle, R.D.; Randklev, C.R. Integrative taxonomy reveals a new species of freshwater mussel, Potamilus streckersoni sp. nov. (Bivalvia: Unionidae): Implications for conservation and management. Syst. Biodivers. 2019, 17, 331–348. [Google Scholar] [CrossRef]
- Klunzinger, M.W.; Whisson, C.; Zieritz, A.; Benson, J.A.; Stewart, B.A.; Kirkendale, L. Integrated taxonomy reveals new threatened freshwater mussels (Bivalvia: Hyriidae: Westralunio) from southwestern Australia. Sci. Rep. 2022, 12, 20385. [Google Scholar] [CrossRef] [PubMed]
- Zieritz, A.; Lopes-Lima, M.; Bogan, A.E.; Sousa, R.; Walton, S.; Rahim, K.A.A.; Wilson, J.J.; Ng, P.Y.; Froufe, E.; McGowan, S. Factors driving changes in freshwater mussel (Bivalvia, Unionida) diversity and distribution in Peninsular Malaysia. Sci. Total Environ. 2016, 571, 1069–1078. [Google Scholar] [CrossRef] [PubMed]
- Zieritz, A.; Lopes-Lima, M. Handbook and National Red-List of the Freshwater Mussels of Malaysia; Zieritz & Lopes-Lima: Kuala Lumpur, Malaysia, 2018. [Google Scholar]
- Suwannapoom, C.; Grismer, L.L.; Pawangkhanant, P.; Poyarkov, N.A. A new species of stream toad of the genus Ansonia Stoliczka, 1870 (Anura: Bufonidae) from Nakhon Si Thammarat Range in southern Thailand. Zootaxa 2022, 5168, 119–136. [Google Scholar] [CrossRef]
- Trivalairat, P.; Sumontha, M.; Kunya, K.; Chiangkul, K. Acanthosaura meridiona sp. nov. (Squamata: Agamidae), a new short-horned lizard from southern Thailand. Herpetol. J. 2022, 32, 34–50. [Google Scholar] [CrossRef]
- Yodthong, S.; Rujirawan, A.; Stuart, B.L.; Aowphol, A. A new Limnonectes (Anura:Dicroglossidae) From Southern Thailand. Animals 2021, 11, 566. [Google Scholar] [CrossRef]
- Harvey, M.S. A new species of Garypus (Pseudoscorpiones: Garypidae) from southern Thailand. Rev. Suisse Zool. 2021, 128, 221–225. [Google Scholar] [CrossRef]
- Aratrakorn, S.; Thunhikorn, S.; Donald, P.F. Changes in bird communities following conversion of lowland forest to oil palm and rubber plantations in southern Thailand. Bird Conserv. Int. 2006, 16, 71–82. [Google Scholar] [CrossRef]
- Sousa, R.; Halabowski, D.; Labecka, A.M.; Douda, K.; Aksenova, O.; Bespalaya, Y.; Bolotov, I.; Geist, J.; Jones, H.A.; Konopleva, E.; et al. The role of anthropogenic habitats in freshwater mussel conservation. Glob. Chang. Biol. 2021, 27, 2298–2314. [Google Scholar] [CrossRef]
- Bolotov, I.N.; Konopleva, E.S.; Vikhrev, I.V.; Win, T.; Lunn, Z.; Chan, N.; Gofarov, M.Y.; Kondakov, A.V.; Tomilova, A.A.; Pasupuleti, R.; et al. Follow the Footsteps of Leonardo Fea: An Example of an Integrative Revision of Freshwater Mussel Taxa Described from the Former British Burma (Myanmar). J. Zool. Syst. Evol. Res. 2022, 2022, 6600359. [Google Scholar] [CrossRef]
- Jeratthitikul, E.; Sutcharit, C.; Ngor, P.B.; Prasankok, P. Molecular phylogeny reveals a new genus of freshwater mussels from the Mekong River Basin (Bivalvia: Unionidae). Eur. J. Taxon. 2021, 775, 119–142. [Google Scholar] [CrossRef]
- Pfeiffer, J.M.; Graf, D.L. Re-analysis confirms the polyphyly of Lamprotula Simpson, 1900 (Bivalvia: Unionidae). J. Molluscan Stud. 2013, 79, 249–256. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Taxonomic Status of Specimen | Specimen Voucher (RMBH) | Measurements | GenBank Acc. Numbers | ||||
|---|---|---|---|---|---|---|---|---|
| L | H | W | COI | 16S rRNA | 28S rRNA | |||
| Trapezoideus thachiadensissp. nov. | Holotype | biv 1340 | 37.5 | 24.4 | 12.0 | OP729805 | OP735372 | OP735357 |
| Monodontina vondembuschiana tapienicassp. nov. | Holotype | biv 1272/1 | 58.1 | 36.2 | 20.3 | OP729818 | OP735377 | n/a |
| M. vondembuschiana tapienicassp. nov. | Paratype | biv 1272/2 | 46.9 | 27.3 | 15.3 | OP729819 | OP735378 | OP735361 |
| M. vondembuschiana tapienicassp. nov. | Paratype | biv 1272/3 | 43.9 | 28.6 | 14.5 | OP729820 | n/a | OP735362 |
| M. vondembuschiana tapienicassp. nov. | Paratype | biv 1272/4 | 48.0 | 29.0 | 14.6 | OP729821 | n/a | OP735363 |
| M. vondembuschiana thasaenicassp. nov. | Holotype | biv 1321/1 | 62.8 | 35.4 | 17.7 | OP729822 | OP735379 | OP735364 |
| M. vondembuschiana thasaenicassp. nov. | Paratype | biv 1321/2 | 47.7 | 29.4 | 14.9 | OP729823 | n/a | n/a |
| Sundadontina plugpomenicasp. nov. | Holotype | biv 1324/1 | 54.3 | 35.0 | 21.1 | OP729828 | OP735383 | OP735367 |
| S. plugpomenicasp. nov. | Paratype | biv 1324/2 | 56.8 | 37.6 | 21.0 | OP729829 | n/a | n/a |
| S. plugpomenicasp. nov. | Paratype | biv 1324/3 | 54.0 | 34.7 | 21.5 | OP729830 | OP735384 | OP735368 |
| S. plugpomenicasp. nov. | Paratype | biv 1331 | 33.7 | 22.9 | 13.5 | OP729831 | OP735385 | n/a |
| Songkhlanaia tamodienicagen. & sp. nov. | Holotype | biv 1332 | 44.2 | 25.3 | 13.4 | OP729832 | OP735386 | OP735369 |
| Species | Mean COI p-Distance from the Nearest Neighbor of New Species, % | Nearest Neighbor of the New Species | Fixed Unique Nucleotide Differences Based on the Sequence Alignment of Congeners | ||
|---|---|---|---|---|---|
| COI | 16S rRNA | 28S rRNA | |||
| Trapezoideus thachiadensissp. nov. | 2.4 ± 0.6 | T. subviridus Pfeiffer, Graf, Cummings & Page, 2021 | 338 A, 380 T, 437 C | 177 C, 208 G, 240 A, 269 G, 315 C, 322 C, 327 C, 379 C | 436 A, 66 4 T * |
| Monodontina vondembuschiana tapienicassp. nov. | 1.9 ± 0.5 | M. v. thasaenicassp. nov. | 233 C, 347 T | 242 T | 763 C |
| M. vondembuschiana thasaenicassp. nov. | 1.9 ± 0.5 | M. v. tapienicassp. nov. | 348 C, 419 T, 494 A | 209 A, 289 A, 332 C | None |
| Sundadontina plugpomenicasp. nov. | 2.8 ± 0.6 | S. cumingii (Lea, 1851) | 182 G, 605 C | 291 G | None |
| Songkhlanaia tamodienicagen. & sp. nov. | 9.7 ± 1.2 | Sundadontina brandti Bolotov et al., 2020 | 317 C, 329 G, 350 T | 244 T, 248 A, 316 C | 597 C |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Konopleva, E.S.; Lheknim, V.; Sriwoon, R.; Kondakov, A.V.; Tomilova, A.A.; Gofarov, M.Y.; Vikhrev, I.V.; Bolotov, I.N. Diversity and Phylogenetics of Freshwater Mussels (Unionidae) from Southern Thailand with the Description of One New Genus and Five New Species-Group Taxa. Diversity 2023, 15, 10. https://doi.org/10.3390/d15010010
Konopleva ES, Lheknim V, Sriwoon R, Kondakov AV, Tomilova AA, Gofarov MY, Vikhrev IV, Bolotov IN. Diversity and Phylogenetics of Freshwater Mussels (Unionidae) from Southern Thailand with the Description of One New Genus and Five New Species-Group Taxa. Diversity. 2023; 15(1):10. https://doi.org/10.3390/d15010010
Chicago/Turabian StyleKonopleva, Ekaterina S., Vachira Lheknim, Rujinard Sriwoon, Alexander V. Kondakov, Alena A. Tomilova, Mikhail Y. Gofarov, Ilya V. Vikhrev, and Ivan N. Bolotov. 2023. "Diversity and Phylogenetics of Freshwater Mussels (Unionidae) from Southern Thailand with the Description of One New Genus and Five New Species-Group Taxa" Diversity 15, no. 1: 10. https://doi.org/10.3390/d15010010
APA StyleKonopleva, E. S., Lheknim, V., Sriwoon, R., Kondakov, A. V., Tomilova, A. A., Gofarov, M. Y., Vikhrev, I. V., & Bolotov, I. N. (2023). Diversity and Phylogenetics of Freshwater Mussels (Unionidae) from Southern Thailand with the Description of One New Genus and Five New Species-Group Taxa. Diversity, 15(1), 10. https://doi.org/10.3390/d15010010