Abstract
Climate warming can directly affect biological processes in marine environments. Here, we investigated if warming (+2 °C) can change dynamics in viral and prokaryotic populations in the cold seasons in natural seawaters. We monitored the changes in viral production and prokaryotic growth rate. The prokaryotic average gross growth rates were 0.08 and 0.34 h−1 in November and 0.06 and 0.41 h−1 in December in the in situ and warming experiments, respectively. We found that warming water temperature resulted in a significant increase in prokaryotic growth rates. In warming experiments, the overall viral production rate was about 0.77–14.4 × 105 viruses mL−1 h−1, and a rough estimate of prokaryotic mortality was about 5.6–6.8 × 104 cells mL−1 h−1. Based on our estimation, burst sizes of about 21 and 14 viruses prokaryotes−1 were measured under the experimental warming period. Moreover, the results found that an increased water temperature in the subtropical western Pacific coastal waters increases prokaryotic growth rates, enhances viral production, and changes the carbon fluxes in the trophic interactions of microbes.
1. Introduction
Viruses are an integral part of the microbial community, substantially causing mortality among marine prokaryotes. According to the principle of the “Killing The Winner” model, changes in the prokaryotic community’s structure manifest the ecological impacts of viral lysis [1]. They also play a crucial function in the operations of marine food webs and nutrients cycling [1], a vital process in sustaining microbial food webs, particularly in oligotrophic ecosystems.
Temperature is an important environmental factor that influences prokaryotic growth efficiency [2], and growth rate [3,4]. Changes in prokaryotic growth were expected to affect microbial processes (e.g., viral lysis and prokaryotic mortality rates). Several field studies have further evidenced an increase in the rates of viral lysis among phytoplanktons over the North Atlantic Ocean from high to low latitudes, which, interestingly, is positively correlated with temperature [5]. Changes in host physiology caused by temperature were indeed found altering the mechanisms of viral lysis, which possibly engenders the development of viral resistance [6]. The rise of sea surface temperature caused by global warming may significantly impact the viral population and their interaction with the marine microbial community [7]. The mechanisms of how global warming affects microbial communities and viruses remain in question. There is still limited knowledge on the effect of warming on prokaryotic mortality by viruses and even less information about the prokaryotes-viruses interaction.
We have performed several studies about prokaryotic growth on the subtropical western Pacific coastal waters [8,9]. These waters have been described in our previous reports derived from data collected from 1999 to 2001 [8]. A previous study found surface water temperatures in March reaching about 15–16 °C and gradually rising to 29 °C by the month of July in the subtropical western Pacific coastal waters. The monthly average concentration of nitrate is the lowest between June and October (>1 μM), when it may reach 12 μM from November to May [8]. Few studies have reported the prokaryotic effects of nanoflagellates and viruses in such a marine environment [9]. Recently, we found a significantly higher occurrence of viral lysis than prokaryotic mortality because of nanoflagellates grazing during cold seasons [9]. Viruses have a crucial role in the aquatic microbial food web by recycling large amounts of carbon and nutrients in winter and preventing prokaryotic production in higher trophic levels [9]. Tsai et al. [8] also reported a seasonal cycle with two phases, namely warm season (>25 °C) and cold season (<25 °C), having a 10-fold variation of prokaryotic growth. The prokaryotic growth rate and temperatures exceeding 25 °C had no significant correlation. Hence, the possibility of changes at the food web’s base (due to increased water temperature) significantly affecting different aspects of marine ecosystems’ operations and structure. To better understand the global nutrient and carbon cycle in seawaters, it is particularly important to examine the impact of increased temperatures on microbial communities.
After a century of global warming, large portions of ocean surface waters would incrementally increase to an average temperature of 2 °C in approximation [10], most likely leading to structural and functional changes of marine ecosystems. In this study, we examined the impact of a temperature increase of 2 °C on viral production during colder seasons. We hypothesized that a temperature-driven increase in growth rates of prokaryotes would enhance viral production, indicating that temperature rise has a different effect on the viral lysis of prokaryotes in the colder months (November and December 2020). In this study, Wilhelm et al.’s virus dilution technique [11] was used to determine the rates of viral production, through which we could estimate the prokaryotic mortality by viruses and burst size in this study.
2. Materials and Methods
2.1. Sampling
Samples were collected in November and December 2020 from the surface waters at an established station located in Taiwan’s northeastern coastal waters (25°09.4′ N, 121°46.3′ E). For each sampling, a bucket was used to collect seawater from 07:00 to 08:00 h in the morning (local time). Then, the seawater was gently poured into a clean 5-L Niskin bottle for dilution experiments, and the water’s temperature was measured at the time of casting. Finally, all samples were delivered to the lab immediately within 30 min from the time of sampling.
2.2. Viral Production and Prokaryotic Growth Rate Experiments
First, the grazing-free whole water was prepared, gently vacuum-filtering 2 L of surface seawater through a 47-mm diameter and a 2 μm pore-size polycarbonate track-etched filter membrane (Whatman). 500 mL of virus-free water was produced for viral dilution by filtering grazer-free seawater through a Minimate TFF Capsule (Pall), with a 30-kDa molecular weight cut-off. Removal of viruses through 30 kDa TFF does not fundamentally change the carbon and nutrient composition and prokaryotic assemblage in water. Thus, with slight manipulations, this method is suitable for virus dilution studies [12]. Dilution was performed by adding 400 mL of the virus-free water to 100 mL of grazer-free water, decreasing the prokaryotic and viral abundance to approximately 20% to that of the original seawater [11]. The diluted incubation water was thoroughly mixed and filled in 50 mL plastic incubation tubes. All treatments were incubated in a water bath set at the original temperature where the seawater was during sampling. Further, the experimental warming temperature was set at 2 °C above in situ values (Table 1). Then, the bottles were immediately moved outside the laboratory after preparation to a location near the sampling site. They were then incubated for 12 h under natural light in a thermo-controlled incubator. Particularly, the treatments were performed in triplicates. To determine prokaryotic and viral abundance, 1-mL subsamples were taken every 1 h for a 12 h period at the onset of the experiment. Further, the linear regression between viral abundance and incubation time was used to calculate viral production (VP) (viruses mL−1 h−1). Notably, there was a significant linear relationship between viral abundance and incubation time, while VP was defined as the regression slope [13]. In addition, the growth rate of prokaryotes was estimated in these 20% diluted samples in the exponential growth phase, where the prokaryotic abundance was monitored over time, and the prokaryotic growth rate changes were compared with the in situ and experimental warming temperature samples. The prokaryotic growth rate was calculated as follows:
where μ is growth rate (h−1), N0 and Nt are the prokaryotic abundance at the beginning and the peak of the exponential growth phase, and t is the incubation time that arrival the peak of the exponential growth phase (h).
μ = ln (Nt/N0) t

Table 1.
Water temperature for the incubation experiments. Prokaryotic growth rate, viral production, number of lysed prokaryotes, and burst size in in situ and warming incubation. The VPR (virus-to-prokaryotes ratio) in diluted waters was calculated after the experiments. * The significance of the difference between the value of in situ and warming incubation. ± represents the SD estimated from triplicate measurements. nd means not detected.
2.3. Determination of Prokaryotic Mortality by Viruses and Burst Size
Prokaryotic mortality by viruses was measured following the same viral production analysis, where the prokaryotic abundance was monitored over time. Particularly, this was only estimated when prokaryotic abundance significantly decreased during the incubation period (i.e., generally 8 h after the onset of the experiment). Then, the linear regression between prokaryotic abundance and time observed during the time-course experiments was used to calculate prokaryotic mortality by viruses; it was expressed as the number of cells mL−1 h−1. Subsamples were collected at each hour during the incubation time. Furthermore, the burst size was estimated using the results from the time courses; the increase in viral abundance with time was divided by the number of killed prokaryotic cells [14].
2.4. Enumeration of Viral and Prokaryotic Abundance by Flow Cytometry (FCM)
Viral subsamples of 0.5 mL were collected every 1 h from each incubation set up and fixed in glutaraldehyde (0.5% final concentration) at 4 °C for at least 15 min. Subsequently, the collected subsamples were deep-frozen in liquid nitrogen. On the other hand, 1-mL of prokaryotic subsamples were also collected every 1 h from each incubation and fixed in paraformaldehyde (1% final concentration). The collected viral and prokaryotic samples were preserved at a temperature of −80 °C until the flow cytometry (FCM) analysis.
Viral and prokaryotic samples were analyzed using a CytoFLEX S flow cytometer (Beckman Coulter, Indianapolis, IN, USA) with a 488 nm air-cooled argon-ion laser, a standard 525 nm filter, and an SYBR signal trigger. Prior to staining, viral samples were diluted at 1:10 in TE buffer (pH 8.0, EM grade) to minimize the interference from high particle abundance. The diluted samples were stained with SYBR Green I (final concentration 1:50,000 of commercial stock) and incubated for 10 min at 80 °C without light. After staining, these were then cooled to 25 °C in an ice bath and processed through FCM according to Brussaard’s method [15]. TE buffer’s blank controls stained with the same concentration of SYBR Green I were run to detect and eliminate any noise from the buffer. The quantification of viruses by FCM is comparable to epifluorescence microscopy and is suitable for fast quantification [16]. Furthermore, the prokaryotic samples were pigmented with an SYBR Green I (final concentration 1:10,000) in the dark for 15 min and processed through FCM according to the protocol of Hammes and Egli [17].
2.5. Statistical Analysis
The relationship between viral abundance and the time for triplicate incubations was analyzed using linear regression analysis. Meanwhile, an analysis of variance (ANOVA) was used to test the significance of the slope. Warmer conditions and ambient were compared using the nonparametric Mann-Whitney test, while The STATISTICA 7.0 software was used for all statistical operations. Notably, a probability value of <0.05 was considered significant.
3. Results
In the study, in situ prokaryotic and viral abundance were 4.0 ± 0.6 × 105 cells mL−1 and 5.9 ± 0.9 × 106 viruses mL−1 in November, and 1.6 ± 0.4 × 105 cells mL−1 and 4.3 ± 0.3 × 106 viruses mL−1 in December, respectively. The time-course experiments in this study revealed an increased prokaryotic abundance in the first 6 or 7 h, maintained the higher values for 3–4 h, which subsequently decreased (Figure 1A,B). Furthermore, during the sampling period in the exponential growth phase, the prokaryotic growth rate was 0.08 and 0.34 h−1 in November and 0.06 and 0.41 h−1 in December in the in situ and warming experiments during the winter seasons, respectively (Table 1). Significantly increased prokaryotic growth rates resulted from the warming conditions (Mann-Whitney test, p < 0.05) (Table 1).
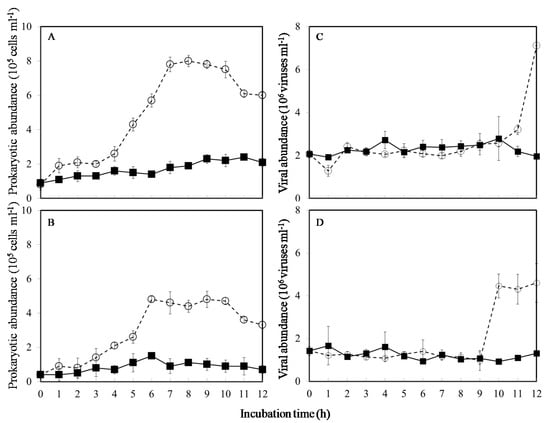
Figure 1.
Temporal variations in prokaryotic and viral abundance at ambient (■), and warming temperature treatments (○) in November (A,C) and December (B,D), respectively. Vertical bars show ± standard deviations (n = 3).
In this study, temporal patterns exhibited a change in viral abundance with incubation time between in situ and warmer conditions in the cold seasons (t-test, p < 0.05) (Figure 1C,D). In the in-situ temperature incubation, the viral abundance fluctuated between 1.9 and 2.7 × 106 viruses mL−1 in November and 0.9 and 1.6 × 106 viruses mL−1 in December (Figure 1C,D). In contrast to in situ temperature incubation, viral abundance increased abruptly at 12 h and 10 h during the warming incubation in November and December, respectively (Figure 1C,D).
In the current study, a significantly increased viral abundance with incubation time after 9 h was observed in warmer conditions (Figure 1C,D). Thus, the VP rates were estimated from the viral abundance’s slope of the incubation experiment between 9 h and 12 h (Figure 2A) to compare the difference between in situ and warmer conditions in the colder months. Obviously, in the colder months, the overall value of VP was about 0.77–1.44 × 106 viruses mL−1 h−1 under warming experiments (Table 1). However, no significant slope of the regression line was observed in ambient temperature for 12 h incubation, and we did not estimate the VP under ambient treatments in this study (ANOVA, p > 0.05) (Table 1).
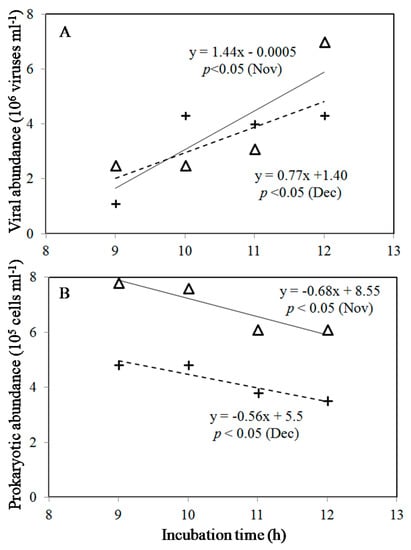
Figure 2.
Relationships between viral abundance (A), prokaryotic abundance (B), and incubation time between 9 h and 12 h in November (△) and December (+). (—) and (– –) are regression lines for November and December.
The prokaryotic mortality by viruses and burst size are reported in Table 1. In this study, we estimated prokaryotic mortality using the linear regression between prokaryotic abundance and time between 9 h and 12 h (Figure 2B). The prokaryotic effects of viral infection increased along with an increase in temperature, accounting for 0.68 and 0.56 × 105 cells mL−1 h−1 in November and December in warmer conditions, respectively (Table 1). Furthermore, based on our estimation, the burst size of 21 and 14 gives a rough estimate in this study (Table 1).
4. Discussion
Some models on global climate change predicted a +2 °C increase in temperature increase over the next century [10]. Thus, our experimental scales on warming treatment were reasonable with this study’s temperature change rates. The favorable impact of the small and incremental increases in temperature on prokaryotic growth rates observed in this study is consistent with those in prior studies [4,18]. For one, Li [19] suggested that temperature dominantly affects prokaryotic growth rates in colder waters for a year. Meanwhile, our findings demonstrate that possible temperature changes in the Pacific coastal waters may result in increased prokaryotic activity in the future, which will promote other microbial activities. However, previous studies reported that due to counteracting effects (e.g., elevated predation), an increase in abundance might not necessarily follow after a warming stimulation of prokaryotic growth [20]. Further, a few studies have investigated the temperature’s effects on carbon transfer between prokaryotes and protists in natural microbial communities. Notably, they reported positive effects of temperature on the nanoflagellates’ grazing rates on prokaryotes [4]. In the present study, the size fractionation used for the grazers (<2 μm) was employed to eliminate nanoflagellates. While we did not directly assess prokaryotic grazing rates in our experiments, nanoflagellate grazing rates seemed higher in the warming experiments during winter than ambient temperature treatment in our study environment [21].
The present study mainly aimed to observe the effect of the increase in temperature on the prokaryotic loss caused by viruses and the viral ecology in the Pacific coastal waters. The results of the warming experiments confirmed the hypothesis that prokaryotic and viral abundance increase with rising temperatures. The virus dilution technique is preferred to determine marine viral production [10]. Particularly, a dilution of viral abundance “ensures the low occurrence of new phage infections and reduces possible viral losses due to new infection” [22]. Our results observed that the rise in sea surface temperature caused by global warming could significantly affect the viral population and their interaction with the marine microbial community. At the same time, studies reported that warming stimulates prokaryotic growth rates and viral production [23,24,25,26]. Noteworthily, although these studies focused mainly on single-specie host cultures and viral isolates, there is little knowledge on the effects of warming on the viral dynamics of more complex marine environments.
One limitation of this study is the inability of prokaryotic communities in the short experimental time to properly simulate the effect of long-term adaptation likely occurring in natural systems. Thus, the results may be biased. However, in order to interpret the microbial plankton’s response to environmental stressors, the warming experiment was used. The data of the experiment show that warming (+2 °C) enhanced VP in this study. Indeed, we think that variations of prokaryotic growth and VP are subjected to increasing temperature. Previous studies reported that while the temperature can promote the enhancement of prokaryotic production and growth rates [27], it can “also shorten virus latent period, and increase burst size”, which increases viral production [23,24]. The shortened virus latent period may induce higher VP in the warming experiments, which could explain the overall increased VP in the warmer experimental treatments compared to the in situ condition. Interestingly, there was no significant difference in VPR (virus-to-prokaryotes ratio) in our study between in situ and warmer conditions at the end of incubation experiments (Table 1). Recent findings demonstrated how phages infect their prokaryotic hosts differently, “causing lysis at high temperatures and lysogeny at lower temperatures” [28]. As a result, variations in daily temperature would lead to “switches in phage-prokaryotes interactions” [28].
5. Conclusions
In conclusion, based on these short-term experimental results, and allow us to hypothesize that climate-change-induced warming can be expected to affect prokaryotic and viral activities significantly. Our results provide evidence that the effects of rising temperatures resulted in increased viral production and found that it can exert significant effects on prokaryotic mortality under the global warming scenario. Moreover, this study also implied that warming water temperature reduces the energy transfer efficiency from prokaryotes to higher trophic levels in coastal waters during cold seasons.
Author Contributions
Conceptualization: A.-Y.T.; methodology: A.-Y.T.; validation: A.-Y.T.; formal analysis: A.-Y.T. and G.-C.G.; investigation: A.-Y.T.; resources: A.-Y.T. and G.-C.G.; data curation: A.-Y.T.; writing–original draft preparation: A.-Y.T. and V.M.; writing–review and editing: A.-Y.T. and V.M.; funding acquisition: A.-Y.T. and V.M. All authors have read and agreed to the published version of the manuscript.
Funding
The research was conducted in the frame of the Russian state assignment No. 121040600178-6 and supported by RFBR projects 18-44-920026 (works on the GAF phenomenon) and 21-55-52001, and the Ministry of Science and Technology, ROC (Taiwan), grant number NSC 109-2611-M-019-013.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We appreciate the language editing and helpful comments related to this manuscript from Choice Language Service.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wommack, K.E.; Colwell, R.R. Virioplankton: Viruses in aquatic ecosystems. Microbiol. Mol. Biol. Rev. 2000, 64, 69–114. [Google Scholar] [CrossRef]
- Rivkin, R.B.; Legendre, L. Biogenic carbon cycling in the upper ocean: Effects of microbial respiration. Science 2001, 291, 2398–2400. [Google Scholar] [CrossRef]
- Rivkin, R.B.; Anderson, M.R.; Lajzerowicz, C. Microbial processes in cold oceans. I. Relationship between temperature and bacterial growth rate. Aquat. Microb. Ecol. 1996, 10, 243–254. [Google Scholar] [CrossRef][Green Version]
- Vázquez-Domínguez, E.; Vaqué, D.; Gasol, J.M. Temperature effects on the heterotrophic bacteria, heterotrophic nanoflagellates and microbial top predators of the NW Mediterranean. Aquat. Microb. Ecol. 2012, 67, 107–121. [Google Scholar] [CrossRef]
- Mojica, K.D.A.; Huisman, J.; Wilhelm, S.W.; Brussaard, C.P.D. Latitudinal variation in virus-induced mortality of phytoplankton across the North Atlantic Ocean. ISME J. 2015, 10, 500–513. [Google Scholar] [CrossRef] [PubMed]
- Kendrick, B.J.; DiTullio, G.R.; Cyronak, T.J.; Fulton, J.M.; Van Mooy, B.A.S.; Bidle, K.D. Temperature induced viral resistance in Emiliania huxleyi (Prymnesiophyceae). PLoS ONE 2014, 9, e112134-14. [Google Scholar] [CrossRef] [PubMed]
- Pradeep Ram, A.S.; Boucher, D.; Sime-Ngando, T.; Debroas, D.; Romagoux, J.C. Phage bacteriolysis, protistan bacterivory potential, and bacterial production in a freshwater reservoir: Coupling with temperature. Microb. Ecol. 2005, 50, 64–72. [Google Scholar] [PubMed]
- Tsai, A.Y.; Chiang, K.P.; Chang, J.; Gong, G.C. Seasonal diel variations of picoplankton and nanoplankton in a subtropical Western Pacific coastal ecosystem. Limnol. Oceanogr. 2005, 50, 1221–1231. [Google Scholar] [CrossRef]
- Tsai, A.Y.; Gong, G.C.; Hung, J. Seasonal variations of virus- and nanoflagellate-mediated mortality of heterotrophic bacteria in the coastal ecosystem of subtropical western Pacific. Biogeosciences 2013, 10, 3055–3065. [Google Scholar] [CrossRef]
- Timmermann, A.; Oberhuber, J.; Bacher, A.; Esch, M.; Latif, M.; Roeckner, E. Increased El Niño frequency in a climate model forced by future greenhouse warming. Nature 1999, 398, 694–697. [Google Scholar] [CrossRef]
- Wilhelm, S.W.; Brigden, S.M.; Suttle, C.A. A dilution technique for the direct measurement of viral production: A comparison in stratified and tidally mixed coastal waters. Microb. Ecol. 2002, 43, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Shelford, E.J.; Middelboe, M.; Møller, E.F.; Suttle, C.A. Virus-driven nitrogen cycling enhances phytoplankton growth. Aquat. Microb. Ecol. 2012, 66, 41–46. [Google Scholar] [CrossRef]
- Mei, M.L.; Danovaro, R. Virus production and life strategies in aquatic sediments. Limnol. Oceanogr. 2004, 49, 459–470. [Google Scholar] [CrossRef]
- Danovaro, R.; Corinaldesi, C.; Filippini, M.; Fischer, U.R.; Gessner, M.R.; Jaqchet, S.; Magagnini, M.; Velimirov, B. Viriobenthos in freshwater and marine sediments: A review. Freshw. Biol. 2008, 53, 1186–1213. [Google Scholar] [CrossRef]
- Brussaard, C.P.D. Optimization of procedures for counting viruses by flow cytometry. Appl. Environ. Microbiol. 2004, 70, 1506–1513. [Google Scholar] [CrossRef]
- Chen, F.; Lu, J.R.; Binder, B.J.; Liu, Y.C.; Hodson, R.E. Application of digital image analysis and flow cytometry to enumerate marine viruses stained with SYBR Gold. Appl. Environ. Microbiol. 2001, 67, 539–545. [Google Scholar] [CrossRef]
- Hammes, F.; Egli, T. Cytometric methods for measuring bacteria in water: Advantages, pitfalls and applications. Anal. Bioanal. Chem. 2010, 397, 1083–1095. [Google Scholar] [CrossRef]
- Pomeroy, L.R.; Wiebe, W.J. Temperature and substrates as interactive limiting factors for marine heterotrophic bacteria. Aquat. Microb. Ecol. 2001, 23, 187–204. [Google Scholar] [CrossRef]
- Li, W.K.W. Annual average abundance of heterotrophic bacteria and Synechococcus in surface ocean waters. Limnol. Oceanogr. 1998, 43, 1746–1753. [Google Scholar] [CrossRef]
- Christoffersen, K.; Andersen, N.; Søndergaard, M.; Liboriussen, L.; Jeppesen, E. Implications of climateenforced temperature increases on freshwater pico- and nanoplankton populations studied in artificial ponds during 16 months. Hydrobiologia 2006, 560, 259–266. [Google Scholar] [CrossRef]
- Tsai, A.Y.; Gong, G.C.; Shiau, W. Viral lysis and nanoflagellate grazing on prokaryotes: Effects of short-term warming in a coastal subtropical marine system. Hydrobiologia 2015, 751, 43–54. [Google Scholar] [CrossRef]
- Weinbauer, M.G.; Rowe, J.M.; Wilhelm, S.W. Determining rates of virus production in aquatic systems by the virus reduction approach. Manu. Aquat. Viral Ecol. 2010, 1, 1–8. [Google Scholar]
- Steenhauer, L.M.; Wierenga, J.; Carreira, C.; Limpens, R.W.; Koster, A.J.; Pollard, P.C.; Brussaard, C.P. Isolation of cyanophage CrV infecting Cylindrospermopsis raciborskii and the influence of temperature and irradiance on CrV proliferation. Aquat. Microb. Ecol. 2016, 78, 11–23. [Google Scholar] [CrossRef][Green Version]
- Maat, D.S.; Prins, M.A.; Brussaard, C.P.D. Sediments from arctic tide-water glaciers remove coastal marine viruses and delay host infection. Viruses 2019, 11, 123. [Google Scholar] [CrossRef] [PubMed]
- Danovaro, R.; Corinaldesi, C.; Dell’Anno, A.; Fuhrman, J.A.; Middelburg, J.J.; Noble, R.T.; Suttle, C.A. Marine viruses and global climate change. FEMS Microbiol. Rev. 2011, 35, 993–1034. [Google Scholar] [CrossRef]
- Cavicchioli, R.; Ripple, W.J.; Timmis, K.N.; Azam, F.; Bakken, L.R.; Baylis, M.; Behrenfeld, M.J.; Boetius, A.; Boyd, P.W.; Classen, A.T.; et al. Scientists’ warning to humanity: Microorganisms and climate change. Nat. Rev. Microbiol. 2019, 17, 569–586. [Google Scholar] [CrossRef] [PubMed]
- Lymer, D.; Logue, J.B.; Brussaard, C.P.D.; Baudoux, A.C.; Vrede, K.; Lindström, E.S. Temporal variation in freshwater viral and bacterial community composition. Freshw. Biol. 2008, 53, 1163–1175. [Google Scholar] [CrossRef]
- Egilmez, H.I.; Morozov, A.Y.; Clokie, M.R.J.; Shan, J.; Letarov, A.; Galyov, E.E. Temperature-dependent virus lifecycle choices may reveal and predict facets of the biology of opportunistic pathogenic bacteria. Sci. Rep. 2018, 8, 8642. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).