
Tomato Landraces Are Competitive with Commercial Varieties in Terms of Tolerance to Plant Pathogens—A Case Study of Hungarian Gene Bank Accessions on Organic Farms

 ,
, 
 ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
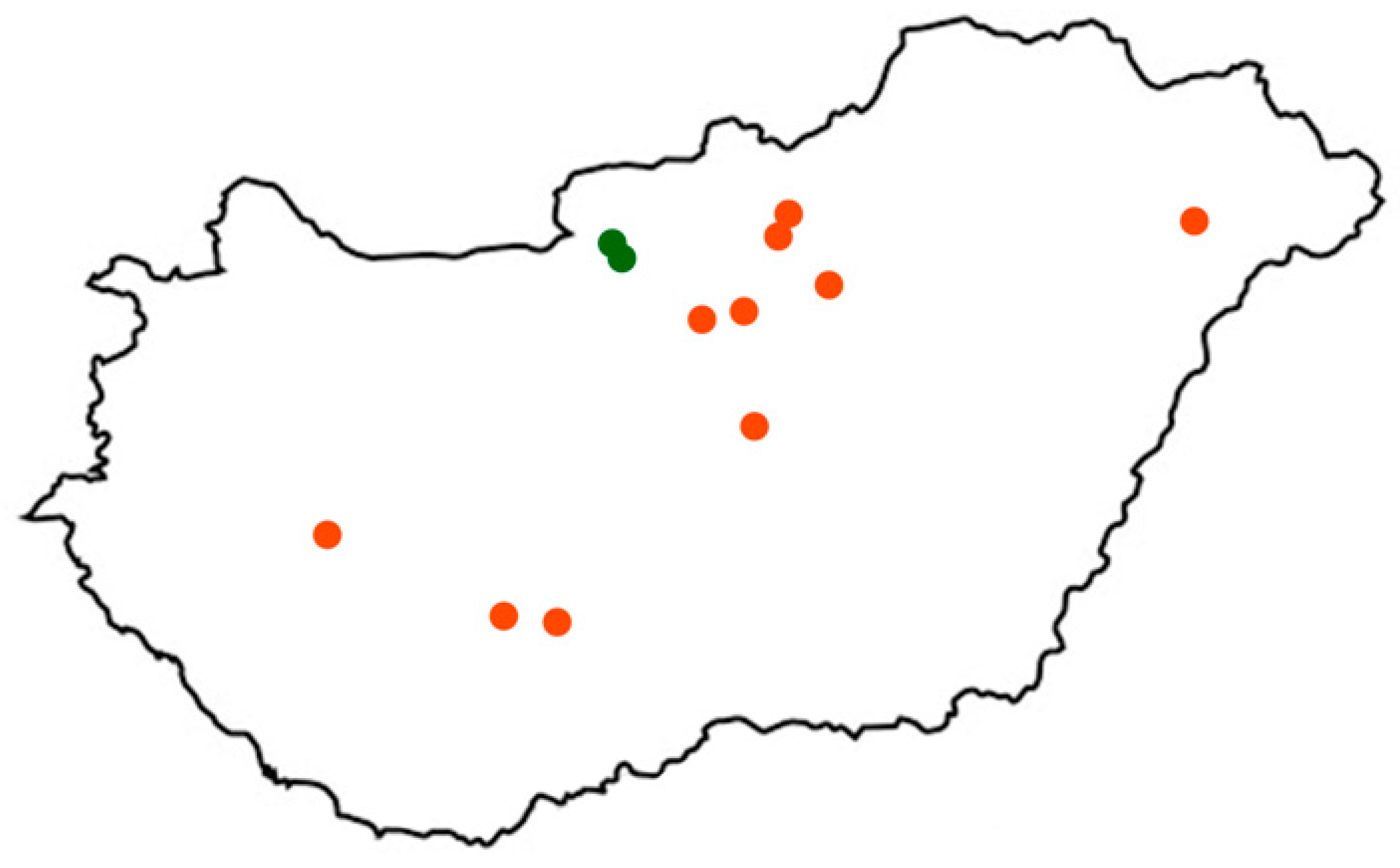
2.1. Location
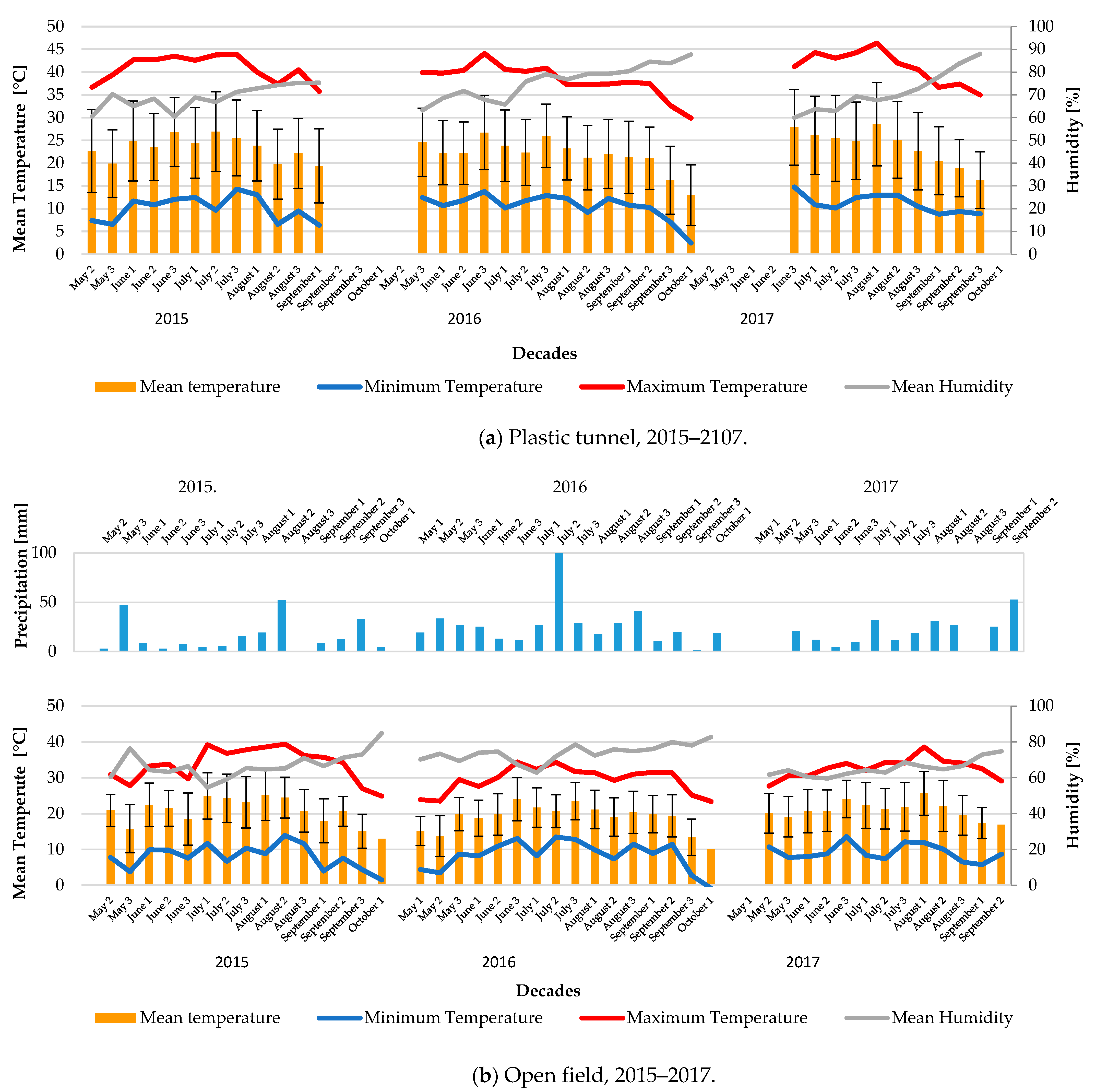
2.2. Weather and Soil
2.3. Landrace Accessions
2.4. Plant Material and Planting
2.5. Farming Methods and Agrotechnical Work
2.6. Experimental Design
2.7. Disease and Yield Evaluation
2.8. Statistical Methods
3. Results
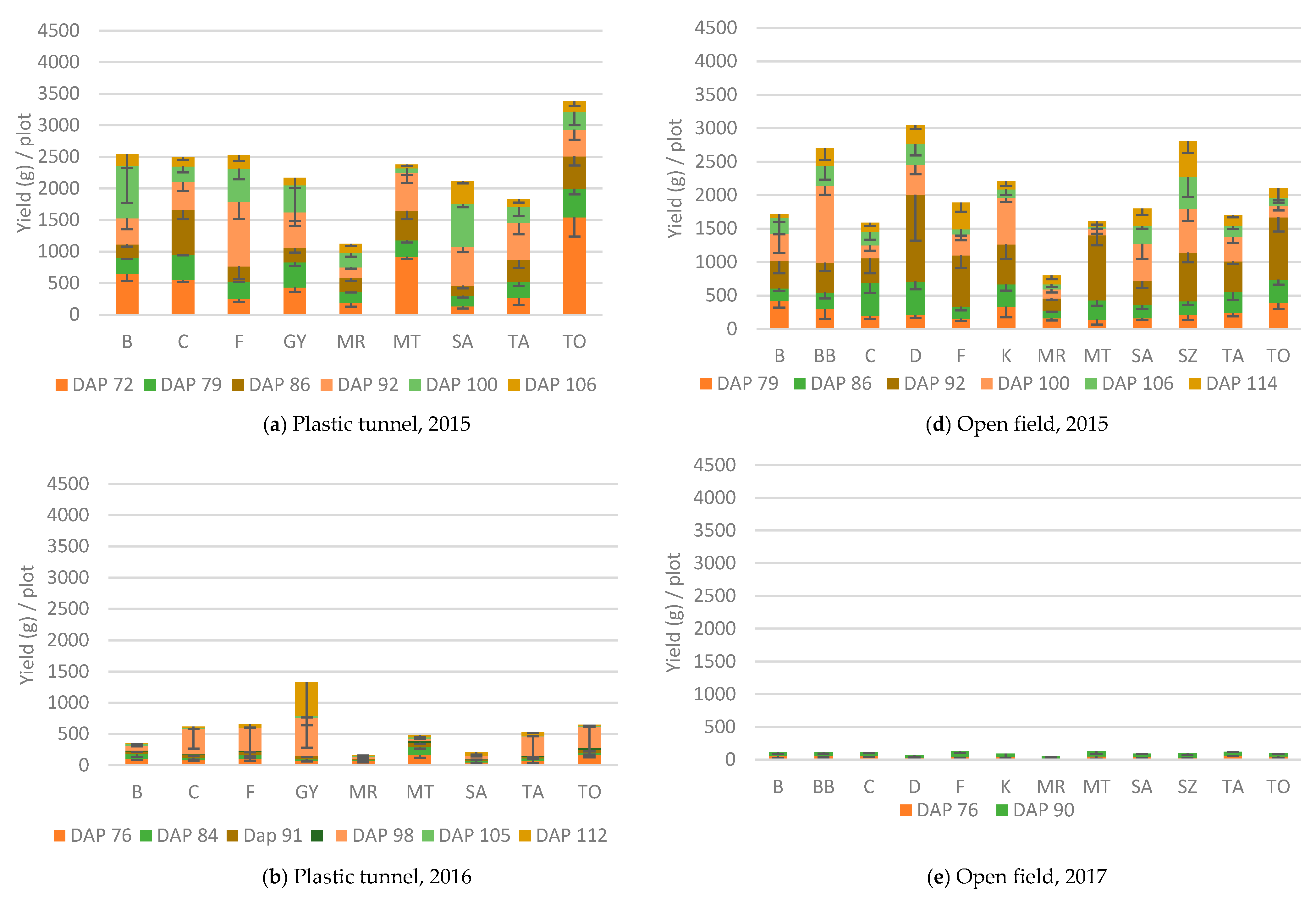
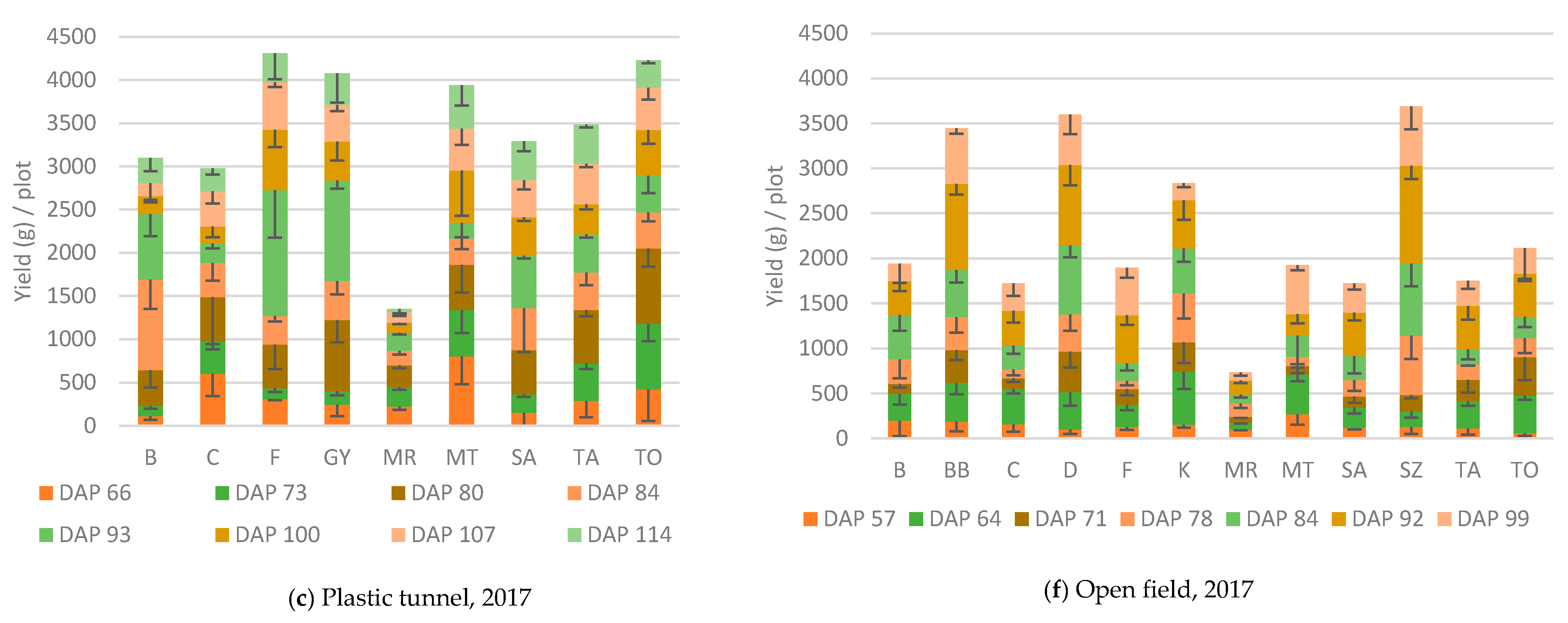
3.1. Yield
3.1.1. Plastic Tunnel
3.1.2. Open field
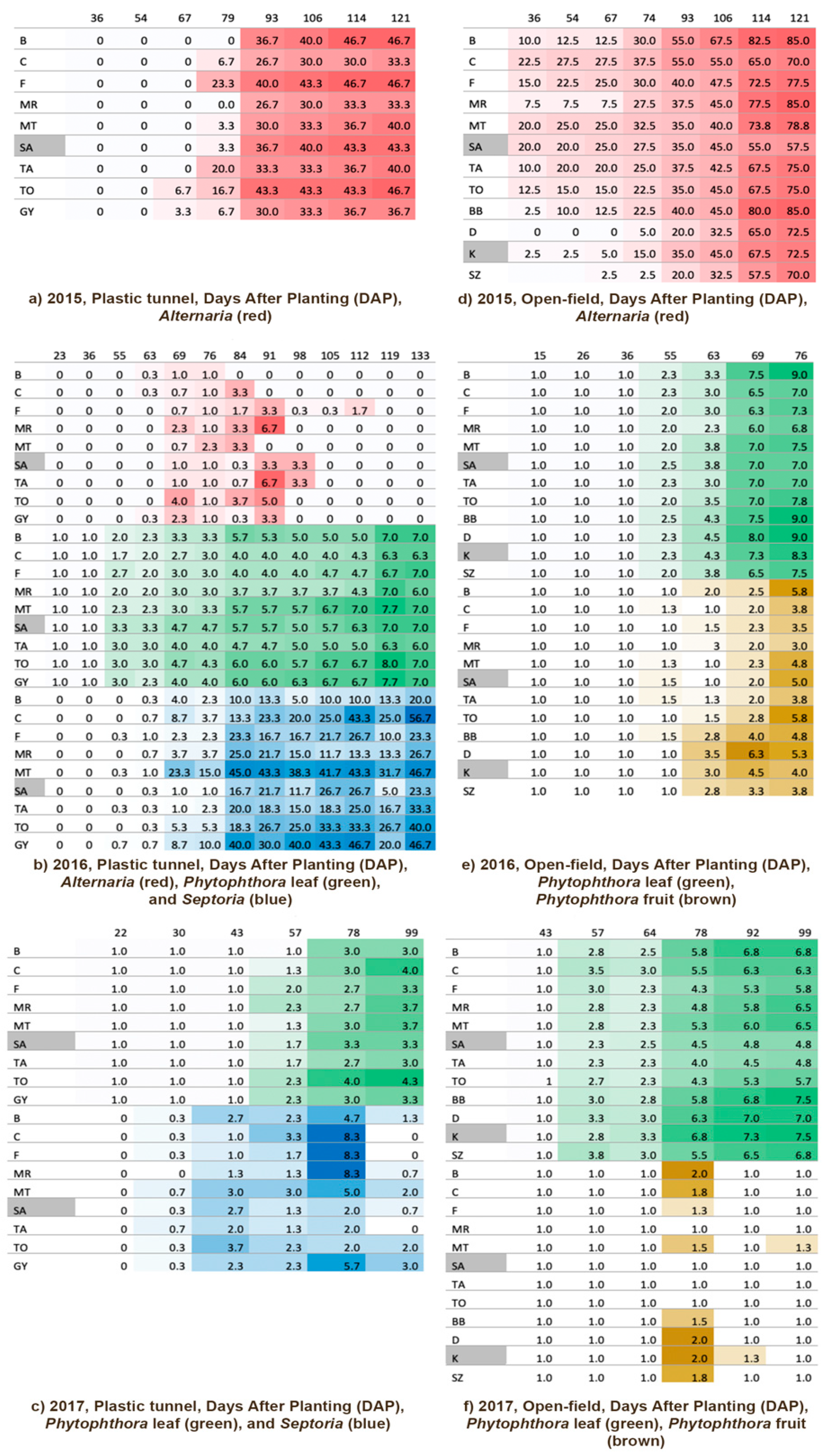
3.2. Diseases
3.2.1. Plastic Tunnel
3.2.2. Open Field
3.3. Subjective Observations
3.3.1. Yield and Fruit Quality
3.3.2. Vegetative Characteristics
4. Discussion
- (1)
- Constitute a viable option for tomato assortment diversification even under current environmental and climatic conditions;
- (2)
- Have sufficient resistance against plant diseases;
- (3)
- Can thus substitute commercial varieties under organic farming conditions, even if they are cultivated outside of their place of origin.
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bergougnoux, V. The history of tomato: From domestication to biopharming. Biotech. Adv. 2014, 32, 170–189. [Google Scholar] [CrossRef] [PubMed]
- FAOSTAT. 2019. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 10 March 2021).
- Yi, S.S.; Jatoi, S.A.; Fujimura, T.; Yamanaka, S.; Watanabe, J.; Watanabe, K.N. Potential loss of unique genetic diversity in tomato landraces by genetic colonization of modern cultivars at a non-center of origin. Plant Breed. 2008, 127, 189–196. [Google Scholar] [CrossRef]
- Mäder, P.; Fliessbach, A.; Dubois, D.; Gunst, L.; Fried, P.; Niggli, U. Soil fertility and biodiversity in organic farming. Science 2002, 296, 1694–1697. [Google Scholar] [CrossRef] [PubMed]
- Østergård, H.; Finckh, M.R.; Fontaine, L.; Goldringer, I.; Hoad, S.P.; Kristensen, K.; Lammerts van Bueren, E.T.; Mascher, F.; Munki, L.; Wolfe, M.S. Time for a shift in crop production: Embracing complexity through diversity at all levels. J. Sci. Food Agric. 2009, 89, 1439–1445. [Google Scholar] [CrossRef]
- Willer, H.; Lernoud, J. The World of Organic Agriculture Statistics and Emerging Trends; FiBL-IFOAM Report; Research Institute of Organic Agriculture (FiBL) Frick, and IFOAM—Organics International: Bonn, Germany, 2015; pp. 1–333. [Google Scholar]
- Lammerts van Bueren, E.T.; Struik, P.C.; Jacobsen, E. Ecological concepts in organic farming and their consequences for an organic crop ideotype. Neth. J. Agric. Sci. 2002, 50, 1–26. [Google Scholar] [CrossRef]
- Murphy, K.M.; Campbell, K.G.; Lyon, S.R.; Jones, S.S. Evidence of varietal adaptation to organic farming systems. Field Crops Res. 2007, 102, 172–177. [Google Scholar] [CrossRef]
- Wolfe, M.S.; Baresel, J.P.; Desclaux, D.; Goldringer, I.; Hoad, S.; Kovacs, G.; Löschenberger, F.; Miedaner, T.; Østergård, H.; Lammerts van Bueren, E.T. Developments in breeding cereals for organic agriculture. Euphytica 2008, 163, 323–346. [Google Scholar] [CrossRef]
- Tembe, K.O.; Chemining, G.; Ambuko, J. Evaluation of African tomato landraces (Solanum lycopersicum) based on morphological and horticultural traits. Agric. Nat. Resour. 2018, 52, 536–542. [Google Scholar] [CrossRef]
- Lammerts van Bueren, E.T.; Jonesc, S.S.; Tammd, L.; Murphy, K.M.; Myerse, J.R.; Leifertf, C.; Messmerd, M.M. The need to breed crop varieties suitable for organic farming, using wheat, tomato and broccoli as examples: A review. J. Life Sci. 2011, 58, 193–205. [Google Scholar] [CrossRef]
- Camacho Villa, T.C.; Maxted, N.; Scholten, M.; Ford-Lloyd, B. Defining and identifying crop landraces. Plant Genet. Resour. 2005, 3, 373–384. [Google Scholar] [CrossRef]
- Harlan, J.R. Our vanishing genetic resources. Science 1975, 188, 618–621. [Google Scholar] [CrossRef]
- Hawkes, J.G. The Diversity of Crop Plants; Harvard University Press: Cambridge, MA, USA, 2013; p. 102. [Google Scholar] [CrossRef]
- Gupta, C.; Salgotra, R.K.; Mahajan, G. Future threats and opportunities facing crop wild relatives and landrace diversity. In Rediscovery of Genetic and Genomic Resources for Future Food Security; Springer: Singapore, 2020; pp. 351–364. [Google Scholar] [CrossRef]
- Perrino, E.V.; Perrino, P. Crop wild relatives: Know how past and present to improve future research, conservation and utilization strategies, especially in Italy: A review. Resour. Crop Evol. 2020, 67, 1067–1105. [Google Scholar] [CrossRef]
- Harlan, J.R.; de Wet, J.M.J. Towards a rational classification of cultivated plants. Taxon 1971, 20, 509–517. [Google Scholar] [CrossRef]
- Perrino, E.V.; Wagensommer, R.P. Crop wild relatives (CWR) priority in Italy: Distribution, ecology, in situ and ex situ conservation and expected actions. Sustainability 2021, 13, 1682. [Google Scholar] [CrossRef]
- Fullana-Pericàs, M.; Conesa, M.; Douthe, C.; El Aou-ouad, H.; Ribas-Carbó, M.; Galmés, J. Tomato landraces as a source to minimize yield losses and improve fruit quality under water deficit conditions. Agric. Water Manag. 2019, 223, 105722. [Google Scholar] [CrossRef]
- Tieman, D.; Zhu, G.; Resende, M.F.R.; Lin, T.; Nguyen, C.; Bies, D.; Rambla, J.L.; Beltran, K.S.O.; Taylor, M.; Zhang, B.; et al. A chemical genetic roadmap to improved tomato flavor. Science 2017, 355, 391–394. [Google Scholar] [CrossRef]
- Akino, S.; Takemoto, D.; Hosaka, K. Phytophthora infestans: A review of past and current studies on potato late blight. J. Gen. Plant Pathol. 2014, 80, 24–37. [Google Scholar] [CrossRef]
- Chaerani, R.; Voorrips, R.E. Tomato early blight (Alternaria solani): The pathogen, genetics, and breeding for resistance. J. Gen. Plant Pathol. Pathol. 2006, 72, 335–347. [Google Scholar] [CrossRef]
- Fry, W.E. Late Blight of Potatoes and Tomatoes; Fact Sheet 726.20, Vegetable Crops, Integrated Pest Management; New York State IPM Program, Cornell University: New York, NY, USA, 1998. [Google Scholar]
- Fry, W. Phytophthora infestans: The plant (and R gene) destroyer. Mol. Plant. Pathol. 2008, 9, 385–402. [Google Scholar] [CrossRef]
- Grigolli, J.F.J.; Kubota, M.M.; Alves, D.P.; Rodrigues, G.B.; Cardoso, C.R.; Henriques da Silva, D.J.; Mizubuti, E.S.G. Characterization of tomato accessions for resistance to early blight. Crop Breed. Appl. Biotechnol. 2011, 11, 174–180. [Google Scholar] [CrossRef]
- Jones, J.B.; Jones, J.P.; Stall, R.E.; Zitter, T.A. Compendium of Tomato Diseases; American Phytopathological Society: St. Paul, MN, USA, 1991; p. 73. [Google Scholar]
- Nowicki, M.; Foolad, M.R.; Nowakowska, M.; Kozik, E.U. Potato and tomato late blight caused by Phytophthora infestans: An overview of pathology and resistance breeding. Plant Dis. 2012, 96, 4–17. [Google Scholar] [CrossRef]
- Nowicki, M.; Kozik, E.U.; Foolad, M.R. Late blight of tomato. In Translational Genomics for Crop Breeding; Varshney, R.K., Tuberosa, R., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2013; pp. 241–265. [Google Scholar]
- Poysa, V.; Brammallz, R.A.; Pitblados, R.E. Effects of foliar fungicide sprays on disease and yield of processing tomatoes in Ontario. Can. J. Plant Sci. 1993, 73, 1209–1215. [Google Scholar] [CrossRef][Green Version]
- Peralta, I.E.; Knap, S.; Spooner, D.M. New species of wild tomatoes (Solanum section Lycopersicon: Solanaceae) from northern Peru. Syst. Bot. 2005, 30, 424–434. [Google Scholar] [CrossRef]
- Sanoubar, R.; Barbanti, L. Fungal diseases on tomato plant under greenhouse condition. Eur. J. Biol. Res. 2017, 7, 299–308. [Google Scholar]
- Madden, L.V. Measuring and modeling crop losses at the field level. Phytopathology 1983, 73, 1591–1596. [Google Scholar] [CrossRef]
- Monteiro1, F.P.; Ogoshi, C.; Maindra, L.C.; Becker, W.F. Culture medium based on tomato leaves for abundant production of conidia from Septoria lycopersici. Asian J. Agric. Res. 2018, 3, 1–6. [Google Scholar] [CrossRef]
- Acciarri, N.; Sabatini, E.; Ciriaci, T.; Rotino, L.G.; Valentino, D.; Tamietti, G. The presence of genes for resistance against Verticillium dahliae in Italian tomato landraces. Eur. J. Hortic. Sci. 2010, 75, 8–14. [Google Scholar]
- Salim, A.P.; Saminaidu, K.; Marimuthu, M.; Perumal, Y.; Rethinasamy, V.; Palanisami, J.R.; Vadivel, K. Defense responses in tomato landrace and wild genotypes to early blight pathogen Alternaria solani infection and accumulation of pathogenesis-related proteins. Arch. Phytopathol. Plant Prot. 2011, 44, 1147–1164. [Google Scholar] [CrossRef]
- Rogers, M.A.; Wszelaki, A.L. Influence of high tunnel production and planting date on yield, growth, and early blight development on organically grown heirloom and hybrid tomato. HortTechnology 2012, 22, 452–462. [Google Scholar] [CrossRef]
- Panthee, D.R.; Gardner, R.G. “Mountain Merit”: A late blight-resistant large-fruited tomato hybrid. HortScience 2010, 45, 1547–1548. [Google Scholar] [CrossRef]
- Seidl Johnson, A.C.; Jordan, S.A.; Gevens, A.J. Novel resistance in heirloom tomatoes and effectiveness of resistance in hybrids to Phytophthora infestans US-22, US-23, and US-24 clonal lineages. Plant Dis. 2014, 98, 761–765. [Google Scholar] [CrossRef]
- Cseperkálóné-Mirek, B.; Reiter, D.; Divéky-Ertsey, A.; Drexler, D. On-farm assessment of landrace of tomato (Lycopersicon esculentum L.) under organic conditions in Hungary. Acta Fytotechn. Zootechn. 2015, 18, 134–137. [Google Scholar] [CrossRef]
- Csambalik, L. Paradicsom Tájfajták Szerepe az Ökológiai Gazdálkodásban (The Role of Tomato Landraces in Organic Farming, in Hungarian). Ph.D. Thesis, Hungarian University of Agriculture and Life Sciences (formerly Szent István University), Budapest, Hungary, 2016; p. 199. [Google Scholar]
- Csambalik, L.; Divéky-Ertsey, A.; Pusztai, P.; Boros, F.; Orbán, C.; Kovács, S.; Gere, A.; Sipos, L. Multi-perspective evaluation of phytonutrients—Case study on tomato landraces for fresh consumption. J. Funct. Foods 2017, 33, 211–216. [Google Scholar] [CrossRef]
- Csambalik, L.; Gál, I.; Sipos, L.; Gere, A.; Koren, D.; Bíró, B.; Divéky-Ertsey, A. Evaluation of processing type tomato plant genetic resources (Solanum lycopersicum L.) for their nutritional properties in different environments. Plant Genet. Resour. 2019, 17, 488–498. [Google Scholar] [CrossRef]
- Horneburg, B.; Becker, H.C. Selection for Phytophthora field resistance in the F2 generation of organic outdoor tomatoes. Euphytica 2011, 180, 357–367. [Google Scholar] [CrossRef]
- Hoagland, L.; Navazio, J.; Zystro, J.; Kaplan, I.; Vargas, J.G.; Gibson, K. Key traits and promising germplasm for an organic participatory tomato breeding program in the U.S. HortScience 2015, 50, 1301–1308. [Google Scholar] [CrossRef]
- Healy, G.K.; Emerson, B.J.; Dawson, J.C. Tomato variety trials for productivity and quality in organic hoop house versus open field management. Renew. Agric. Food. Syst. 2017, 32, 562–572. [Google Scholar] [CrossRef]
- Powell, M.; Gundersen, B.; Cowan, J.; Miles, C.A.; Inglis, D.A. The effect of open-ended high tunnels in western Washington on late blight and physiological leaf roll among five tomato cultivars. Plant Dis. 2014, 98, 1639–1647. [Google Scholar] [CrossRef]
- Ara, N.; Bashar, M.K.; Begum, S.; Kakon, S.S. Effect of spacing and stem pruning on the growth and yield of tomato. International. J. Sustain. Crop Prod. 2007, 2, 35–39. [Google Scholar]
- Kanyomeka, L.; Shivute, B. Influence of pruning on tomato production under controlled environments. Agric. Trop. Subtrop. 2005, 38, 79–83. [Google Scholar]
- Conesa, M.; Fullana-Pericàs, M.; Granell, A.; Galmés, J. Mediterranean long shelf-life landraces: An untapped genetic resource for tomato improvement. Front. Plant Sci. 2020, 10, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Guida, G.; Houssemeddine, M.; Mistretta, C.; Oliva, M.; Buonomo, R.; Mascellis, R.D.; Patanè, C.; Rouphael, Y.; Albrizio, R.; Giorio, P. Agronomical, physiological and fruit quality responses of two Italian long-storage tomato landraces under rain-fed and full irrigation conditions. Agric. Water Manag. 2017, 180, 126–135. [Google Scholar] [CrossRef]
- Sharma, R.L.; Ahir, R.R. Physiological studies of Alternaria causing Alternaria blight of tomato. J. Entomol. Zool. Stud. 2018, 6, 844–847. [Google Scholar]
- Gul, Z.; Ahmed, M.; Ullah-Khan, Z.; Khan, B.; Iqbal, M. Evaluation of tomato lines against septoria leaf spot under field conditions and its effect on fruit yield. Agric. Sci. 2016, 7, 181–186. [Google Scholar] [CrossRef]
- Joslin, K.; Taber, H.; Helland, S.; Gleason, M. Control of the foliar disease, Septoria lycopersici, in organic tomato production. HortScience 2019, 39, 831. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH | SOM (%) | N (ppm) | P (ppm) | K (ppm) | ||
|---|---|---|---|---|---|---|
| 2015 | Plastic tunnel | 7.47 | 2.31 | 20.6 | 250 | 372 |
| Open field | 7.27 | 2.58 | 18 | 146 | 224 | |
| 2016 | Plastic tunnel | 7.42 | 2.53 | 176 | 643 | 562 |
| Open field | 7.32 | 2.8 | 24.9 | 120 | 439 | |
| 2017 | Plastic tunnel | 7.26 | 2.74 | 42.8 | 703 | 326 |
| Open field | 7.24 | 2.45 | 13.5 | 296 | 466 |
| Gene Bank Accession Code | Name | Main Use | Average Fruit Weight (g) | Fruit Shape, Size | Growing Type | Location |
|---|---|---|---|---|---|---|
| RCAT030566 | ‘Balatonboglár’ | fresh consumption, processing | 150–190 | circular, medium | Semi-determinate, determinate | P,O |
| RCAT030275 | ‘Cegléd’ | fresh consumption | 160–180 | circular, medium | indeterminate | P,O |
| RCAT030373 | ‘Fadd’ | fresh consumption | 70–90 | rectangular, medium | indeterminate | P,O |
| RCAT031257 | ‘Gyöngyös’ | salad | 150–200 | cylindrical, medium | indeterminate | P,O |
| RCAT030731 | ‘Máriapócs’ | fresh consumption | 15–20 | circular, small | indeterminate | P,O |
| RCAT057656 | ‘Mátrafüred’ | processing | 300–320 | heart-shaped, large | indeterminate | P,O |
| RCAT030370 | ‘Tarnaméra’ | processing | 50–70 | cylindrical, medium | indeterminate | P,O |
| RCAT030184 | ‘Tolna megye’ | processing | 300–350 | slightly flattened, large | indeterminate | P,O |
| RCAT057829 | ‘Dány’ | processing | 110–130 | circular, medium | determinate | O |
| RCAT078726 | ‘Szentlőrinckáta’ | processing | 50–55 | ovate, medium | determinate | O |
| n.a. (control)) | ‘San Marzano’ | processing | 100–110 | cylindrical, medium | indeterminate | P,O |
| n.a. (control) | ‘Kecskeméti 549’ | processing | 50–60 | ovate, medium | determinate | O |
| Year. | Sowing | Planting |
|---|---|---|
| 2015 | 20 March | 18 May |
| 2016 | 16 March | 25 May |
| 2017 | 12 March | 29 May. |
| Experiment | Year | PCA | MANOVA with Factor ‘Varieties’ | ||||
|---|---|---|---|---|---|---|---|
| PCA Total Variance Explained % | Principal Components (Explained Variances %) | Highly Correlated Explaining Variables | Referred to as | Wilk’s λ | Between-Subject Effects F(8;18) | ||
| Plastic tunnel | 2015 | 74.64% | F1(39.11%) | Y72, Y79, Y86 | ‘Yield_2015_early’ | 0.01 *** | 35.24 *** |
| F2(18.73%) | Y100, Y106 | ‘Yield_2015_late’ | 3.48 * | ||||
| F3(16.79%) | Y92 | ‘Yield_2015_DAP92’ | 3.16 ns | ||||
| 2016 | 78.65% | F1(33.04%) | Y76, Y84, Y91 | ‘Yield_2016_early’ | <0.001 *** | 11.19 *** | |
| F2(25.62%) | Y98, Y112 | ‘Yield_2016_late’ | 1.84 ns | ||||
| F3(19.99%) | Y105 | ‘Yield_2016_DAP105’ | 0.83 ns | ||||
| 2017 | 67.07% | F1(32.25%) | Y80, Y100, Y107, Y114 | ‘Yield_2017_late’ | 0.07 *** | 6.41 *** | |
| F2(21.60%) | Y73, Y93 | ‘Yield_2017_early2’ | 17.24 *** | ||||
| F3(13.22%) | Y66, Y84 | ‘Yield_2017_early1’ | 3.45 * | ||||
| Experiment | Year | Principal Component | Referred to as… | Pairwise Comparisons | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Plastic tunnel | 2015 | F1 | ‘Yield_2015_early’ | MR | SA | TA | F | B | GY | MT | C | TO |
| a | a | ab | abc | abc | bcd | bcd | cd | d | ||||
| F2 | ‘Yield_2015_late’ | MT | TA | MR | C | GY | TO | F | B | SA | ||
| a | ab | a | a | ab | ab | ab | b | b | ||||
| 2016 | F1 | ‘Yield_2016_early’ | SA | MR | TA | GY | C | B | F | TO | MT | |
| a | a | ab | ab | ab | b | b | bc | c | ||||
| 2017 | F1 | ‘Yield_2017_late’ | MR | B | C | SA | TA | GY | MT | F | TO | |
| a | ab | ab | b | b | b | b | b | b | ||||
| F2 | ‘Yield_2017_early2’ | F | GY | B | SA | MR | TA | C | MT | TO | ||
| a | ab | bc | bc | cd | cd | cd | d | d | ||||
| F3 | ‘Yield_2017_early1’ | MT | F | MR | C | TO | GY | TA | SA | B | ||
| a | a | a | a | ab | ab | ab | ab | b | ||||
| Experiment | Year | PCA | MANOVA with Factor ‘Varieties’ | ||||
|---|---|---|---|---|---|---|---|
| PCA Total Variance Explained % | Principal Components (Explained Variances %) | Highly Correlated Explaining Variables | Referred to as | Wilk’s λ | Between-Subject Effects F(11;36) | ||
| Open field | 2015 | 74.55% | F1(30.00%) | Y100, Y106, Y114 | ‘Yield_2015_late’ | 0.02 *** | 11.14 *** |
| F2(23.66%) | Y86, Y92 | ‘Yield_2015_early’ | 8.34 *** | ||||
| F3(20.89%) | Y79 | ‘Yield_2015_first’ | 6.50 *** | ||||
| 2016 | Instead of PCA, MANOVA was performed with the original variables Y76, Y90 | ‘Yield_2016_DAP76’ | 0.55 ns | - | |||
| ‘Yield_2016_DAP90’ | - | ||||||
| 2017 | 74.55% | F1(30.00%) | Y71, Y78, Y84, Y92, Y99 | ‘Yield_2017_late’ | 0.05 *** | 16.18 *** | |
| F2(23.66%) | Y57, Y64 | ‘Yield_2017_early’ | 6.29 *** | ||||
| Experiment | Year | Principal Component | Referred as | Pairwise Comparisons | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Open field | 2015 | F1 | ‘Yield_2015_late’ | MT | MR | TO | C | TA | F | B | K | SA | D | BB | SZ |
| a | ab | abc | abc | abc | abc | abc | bc | cd | cde | de | e | ||||
| F2 | ‘Yield_2015_early’ | MR | SA | B | BB | F | TA | SZ | K | C | MT | TO | D | ||
| a | ab | ab | ab | ab | ab | ab | ab | bc | bc | bc | c | ||||
| F3 | ‘Yield_2015_first’ | F | SZ | MT | MR | SA | D | C | TA | TO | BB | K | B | ||
| a | a | ab | abc | abc | abc | abcd | abcd | bcd | bcd | cd | d | ||||
| 2016 | no significant differences were detected | ||||||||||||||
| 2017 | F1 | ‘Yield_2017_late’ | MR | MT | C | SA | TA | B | F | TO | K | BB | D | SZ | |
| a | ab | ab | ab | ab | ab | ab | bc | bcd | cde | de | e | ||||
| F2 | ‘Yield_2017_early’ | MR | SZ | SA | F | TA | TO | D | B | C | BB | MT | K | ||
| a | ab | abc | abc | abcd | abcd | abcd | abcd | bcd | cd | d | d | ||||
| Experiment | Infection Type Detected | Year | PCA | MANOVA with Factor ‘Varieties’ | ||||
|---|---|---|---|---|---|---|---|---|
| PCA Total Variance Explained % | Principal Components (Explained Variances %) | Highly Correlated Explaining Variables | Referred to as | Wilk’s λ | Between-Subject Effects F(8;18) | |||
| Plastic tunnel | Alternaria | 2015 | 79.91% | F1(61.99%) | A106, A114, A125 | ‘Alternaria_2015_late’ | 0.29 ns | - |
| F2(7.92%) | A67, A79 | ‘Alternaria_2015_early’ | - | |||||
| Alternaria, Phytophthora_leaf, Septoria | 2016 | 67.64% | F1(28.36%) | PL (all) | ‘Phytophthora_leaf_2016’ | <0.001 *** | 10.78 *** | |
| F2(19.81%) | S69, S84, S91, S98, S105, S112, S119, S133 | ‘Septoria_2016_late’ | 1.55 ns | |||||
| F3(11.80%) | S55, S63, S76, S84 | ‘Septoria_2016_early’ | 4.57 *** | |||||
| F4(7.67%) | Alternaria (all) | ‘Alternaria_2016’ | 1.19 ns | |||||
| Phytophthora_leaf, Septoria | 2017 | 71.02% | F1(31.56%) | S30, S43, FL57 | ‘Alternaria+Phytophthora_leaf_2017_early’ | 0.07 *** | 0.76 ns | |
| F2(20.73%) | PL78, PL92 | ‘Phytophthora_leaf_2017_late’ | 6.86 *** | |||||
| F3(18.73%) | S57, S78 | ‘Alternaria_2017_late’ | 2.26 ns | |||||
| Experiment | Infection Type Detected | Year | Principal Component | Referred as | Pairwise Comparisons | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Plastic tunnel | Alternaria spp. | 2015 | no significant differences were detected (Figure 4) | ||||||||||
| 2016 | |||||||||||||
| 2017 | |||||||||||||
| Phytophthora infestans (leaf) | 2015 | no significant differences were detected | |||||||||||
| 2016 | F1 | ‘Phytophthora_leaf_2016’ | C | MR | F | TA | B | SA | MT | TO | GY | ||
| a | a | ab | ab | abc | bc | bc | bc | c | |||||
| 2017 | F1 | ‘Phytophthora_leaf_2017_early’ | no significant differences were detected | ||||||||||
| F2 | ‘Phytophthora_leaf_2017_late’ | F | TA | MR | B | C | GY | MT | SA | TO | |||
| a | a | a | a | a | a | a | a | b | |||||
| Septoria lycopersici | 2015 | no significant differences were detected | |||||||||||
| 2016 | F3 | ‘Septoria_2016_early’ | SA | TO | TA | MR | B | C | GY | F | MT | ||
| a | a | a | ab | ab | ab | ab | ab | b | |||||
| 2017 | no significant differences were detected | ||||||||||||
| Experiment | Infection Type Detected | Year | PCA | MANOVA with Factor ‘Varieties’ | ||||
|---|---|---|---|---|---|---|---|---|
| PCA Total Variance Explained % | Principal Components (Explained Variances %) | Highly Correlated Explaining Variables | Referred to as | Wilk’s λ | Between-Subject Effects F(11;36) | |||
| Open field | Alternaria | 2015 | 75.45% | F1(48.07%) | A36, A54, A67, A74, A93 | ‘Alternaria_2015_early’ | 0.20 *** | 7.36 *** |
| F2(27.38%) | A93, A106, A114, A121 | ‘Alternaria_2015_late’ | 1.78 ns | |||||
| Phytophthora leaf, Phytophthora_ fruit | 2016 | 67.64% | F1(44.77%) | PF63, PF69, PF76, PL (all except DAP 55) | ‘Phytophthora_fruit_and_leaf_2016_ _exceptDAP55’ | <0.13 *** | 13.97 *** | |
| F2(20.09%) | PF55, PL55 | ‘Phytophthora_fruit and leaf_ 2016_DAP55’ | 1.49 ns | |||||
| Phytophthora_leaf | 2017 | Instead of PCA, MANOVA was performed with the original variables PL57, PL64, PL78, PL92, PL99 | ‘Phytophthora_leaf_2017 exceptDAP64’ | 0.05 *** | > 3.60 *** | |||
| ‘Phytophthora_leaf_2017_DAP64’ | 1.69 ns | |||||||
| Experiment | Infection Type Detected | Year | Principal Component | Referred as | Pairwise Comparison | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Open field | Alternaria spp. | 2015 | F1 | ‘Alternaria _2015_early’ | D | SZ | K | BB | MR | TO | B | TA | F | MT | SA | C |
| a | a | ab | abc | abc | abcd | abcd | bcd | bcd | bcd | cd | d | |||||
| 2016 | no significant differences were detected | |||||||||||||||
| 2017 | ||||||||||||||||
| Phytophthora infestans (leaf, fruit) | 2015 | no significant differences were detected | ||||||||||||||
| 2016 | F1 | ‘Phytophthora _fruit_and_leaf 2016 exceptDAP55’ | MR | C | TA | F | SA | MT | SZ | TO | B | K | BB | |||
| a | ab | ab | ab | abc | abcd | cde | bcde | cde | def | ef | f | |||||
| Phytophthora infestans (leaf) | 2017 | FL57 | ‘Phytophthora _leaf_2017 exceptDAP64’ | SA | TA | TO | B | K | MR | MT | BB | F | D | C | SZ | |
| a | a | ab | ab | ab | ab | ab | ab | ab | ab | b | b | |||||
| FL78 | TA | F | TO | SA | MR | MT | C | SZ | B | BB | D | K | ||||
| a | ab | ab | ab | abc | abcd | abcd | abcd | bcd | bcd | cd | d | |||||
| FL92 | TA | SA | F | TO | MR | MT | C | SZ | B | BB | D | K | ||||
| a | ab | abc | abcd | abcde | bcdef | cdef | cdef | def | def | ef | f | |||||
| FL99 | SA | TA | TO | F | C | MR | MT | B | SZ | D | BB | K | ||||
| a | a | ab | abc | abc | abc | abc | bc | bc | bc | c | c | |||||
| Landrace/Control Variety Name | Fruit Physiological Disorders | Foliage Density | Handling of Vegetative Growth | Harvesting |
|---|---|---|---|---|
| ‘Cegléd’ | cracking | intermediate | easy | easy |
| ‘Fadd’ | cracking, green shoulder | intermediate | easy | moderate |
| ‘Gyöngyös’ | cracking | sparse | difficult | easy |
| ‘Máriapócs’ | cracking | intermediate | moderate | difficult |
| ‘Mátrafüred’ | cracking, green shoulder, flower abortion | sparse | difficult | moderate |
| ‘Tarnaméra’ | blossom end rot | dense | easy | moderate |
| ‘Tolna megye’ | catface, flower abortion | sparse | difficult | moderate |
| ‘San Marzano’ (control) | blossom end rot | dense | easy | moderate |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boziné-Pullai, K.; Csambalik, L.; Drexler, D.; Reiter, D.; Tóth, F.; Tóthné Bogdányi, F.; Ladányi, M. Tomato Landraces Are Competitive with Commercial Varieties in Terms of Tolerance to Plant Pathogens—A Case Study of Hungarian Gene Bank Accessions on Organic Farms. Diversity 2021, 13, 195. https://doi.org/10.3390/d13050195
Boziné-Pullai K, Csambalik L, Drexler D, Reiter D, Tóth F, Tóthné Bogdányi F, Ladányi M. Tomato Landraces Are Competitive with Commercial Varieties in Terms of Tolerance to Plant Pathogens—A Case Study of Hungarian Gene Bank Accessions on Organic Farms. Diversity. 2021; 13(5):195. https://doi.org/10.3390/d13050195
Chicago/Turabian StyleBoziné-Pullai, Krisztina, László Csambalik, Dóra Drexler, Dániel Reiter, Ferenc Tóth, Franciska Tóthné Bogdányi, and Márta Ladányi. 2021. "Tomato Landraces Are Competitive with Commercial Varieties in Terms of Tolerance to Plant Pathogens—A Case Study of Hungarian Gene Bank Accessions on Organic Farms" Diversity 13, no. 5: 195. https://doi.org/10.3390/d13050195
APA StyleBoziné-Pullai, K., Csambalik, L., Drexler, D., Reiter, D., Tóth, F., Tóthné Bogdányi, F., & Ladányi, M. (2021). Tomato Landraces Are Competitive with Commercial Varieties in Terms of Tolerance to Plant Pathogens—A Case Study of Hungarian Gene Bank Accessions on Organic Farms. Diversity, 13(5), 195. https://doi.org/10.3390/d13050195