
Diversity and Physiological Tolerance of Native and Invasive Jellyfish/Ctenophores along the Extreme Salinity Gradient of the Baltic Sea


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
1.1. The Baltic Sea and Its General Gelatinous Macro-Zooplankton Community
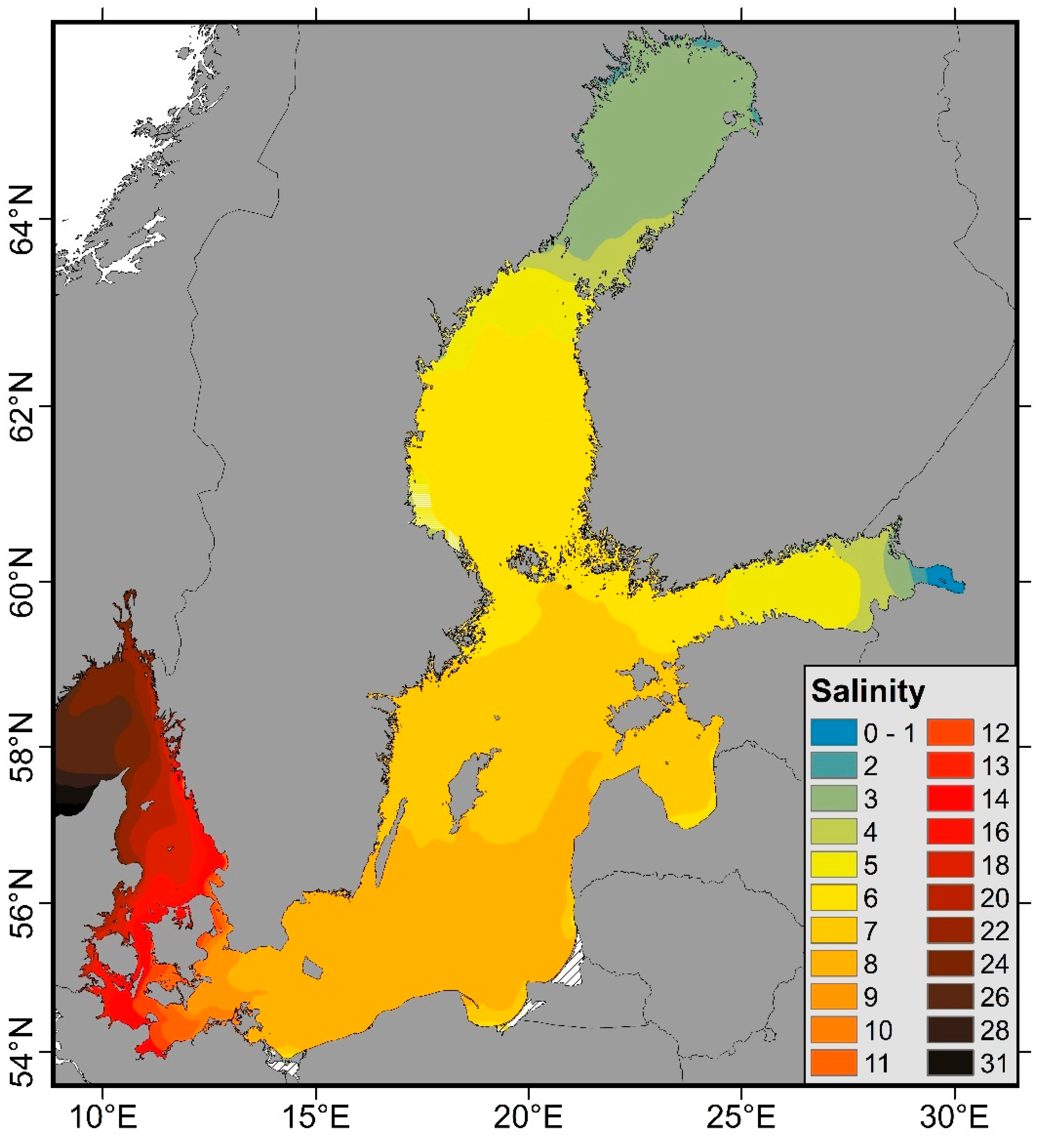
1.2. Physical Environment of the Baltic Sea
1.3. Global Change and Jellyfish Blooms Potential Consequences for the Baltic Sea
2. Jellyfish (Cnidaria)
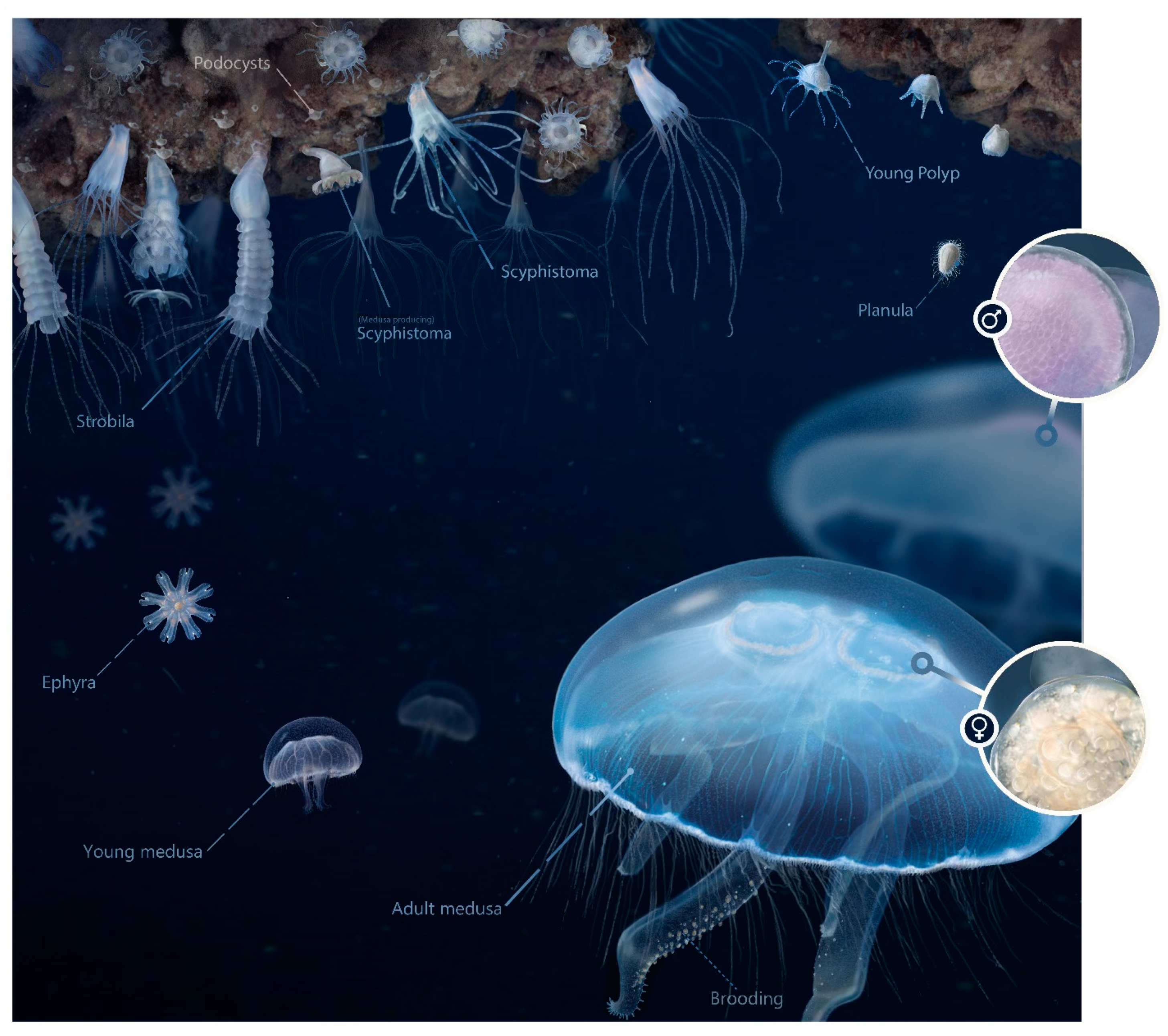
2.1. The Moon Jelly Aurelia aurita (Linnaeus, 1758)
2.1.1. Importance in the Baltic Sea
2.1.2. Distribution in the Baltic
2.1.3. Environmental Tolerances and Preferences
2.2. Lion’s Mane Jellyfish Cyanea capillata (Linnaeus, 1758)
2.2.1. Importance for the Baltic Sea
2.2.2. Distribution in the Baltic
2.2.3. Environmental Tolerances and Preferences
2.3. Hydromedusa Blackfordia virginica (Mayer, 1910)
2.3.1. Importance for the Baltic Sea
2.3.2. Distribution in the Baltic Sea
2.3.3. Environmental Tolerances and Preferences
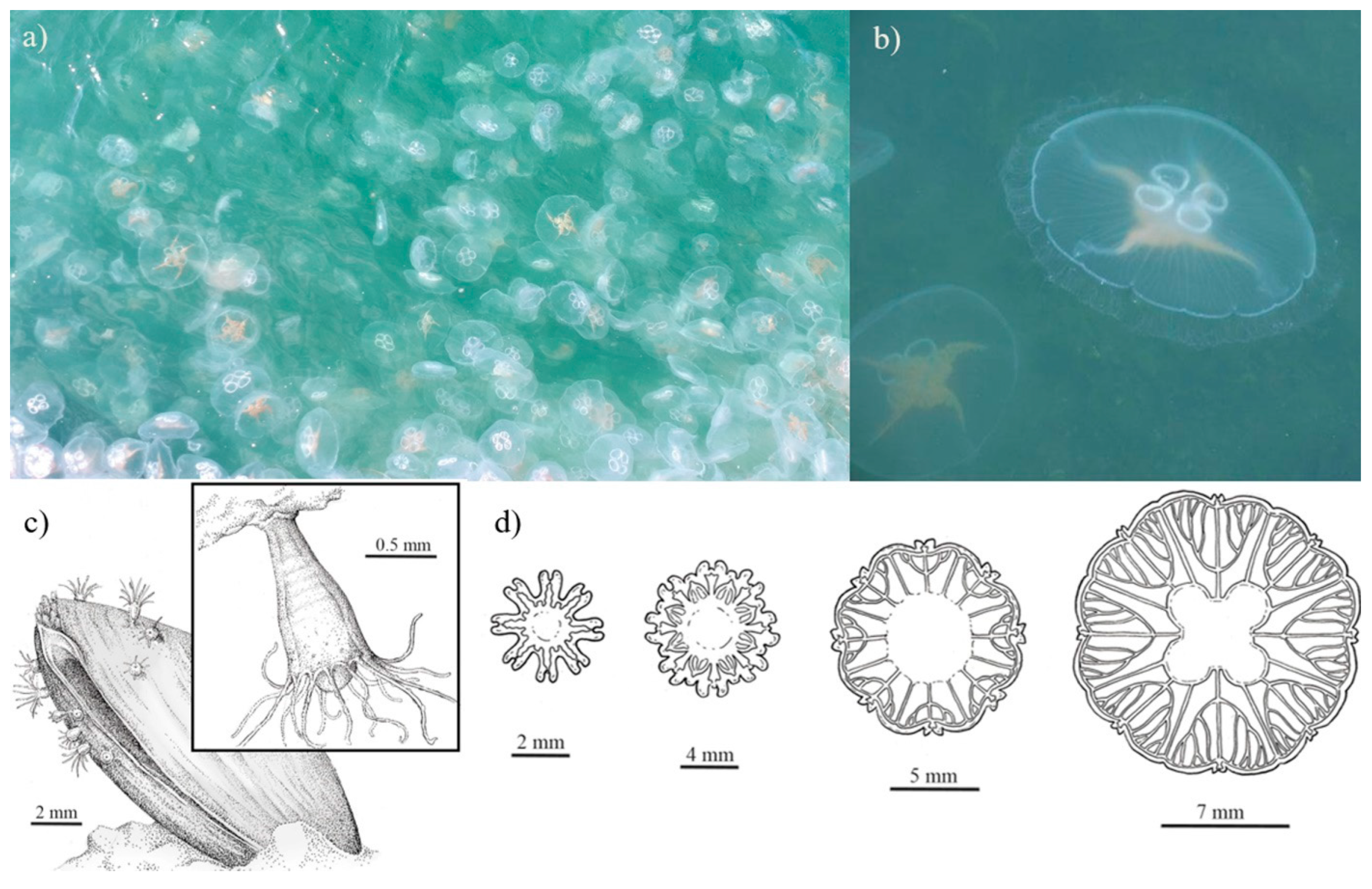
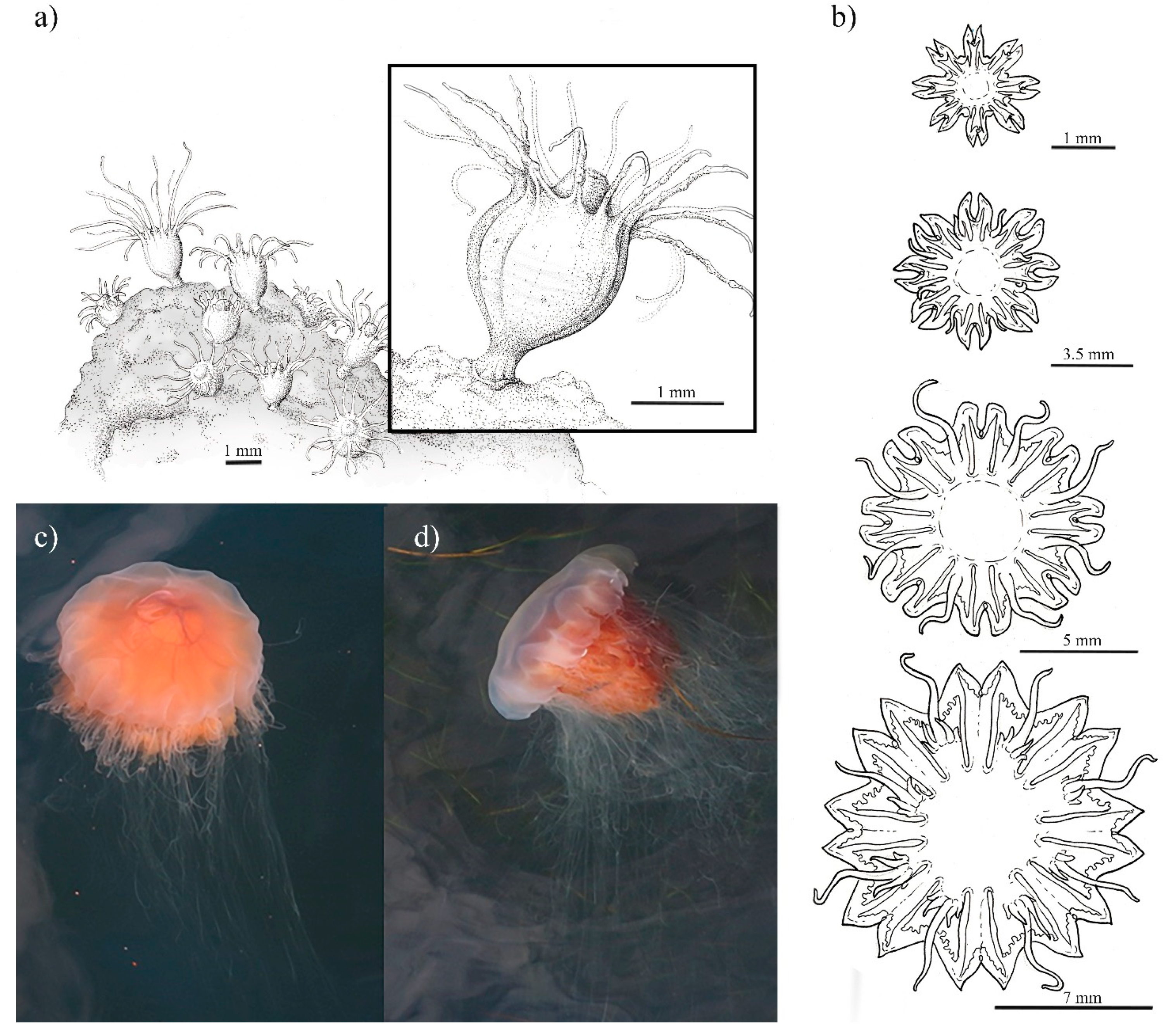
2.4. Morphological Characteristics of Polyps, Ephyrae and Medusa
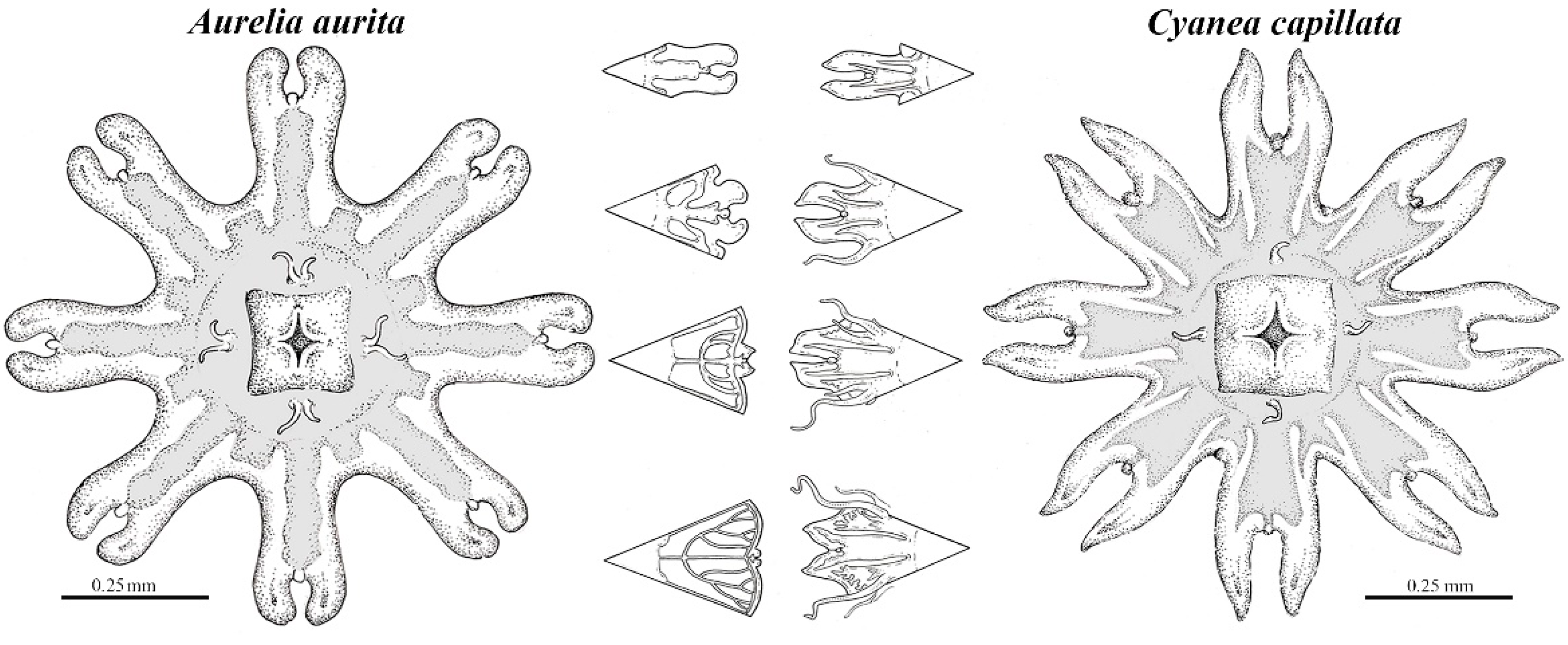
2.4.1. Aurelia aurita
2.4.2. Cyanea capillata
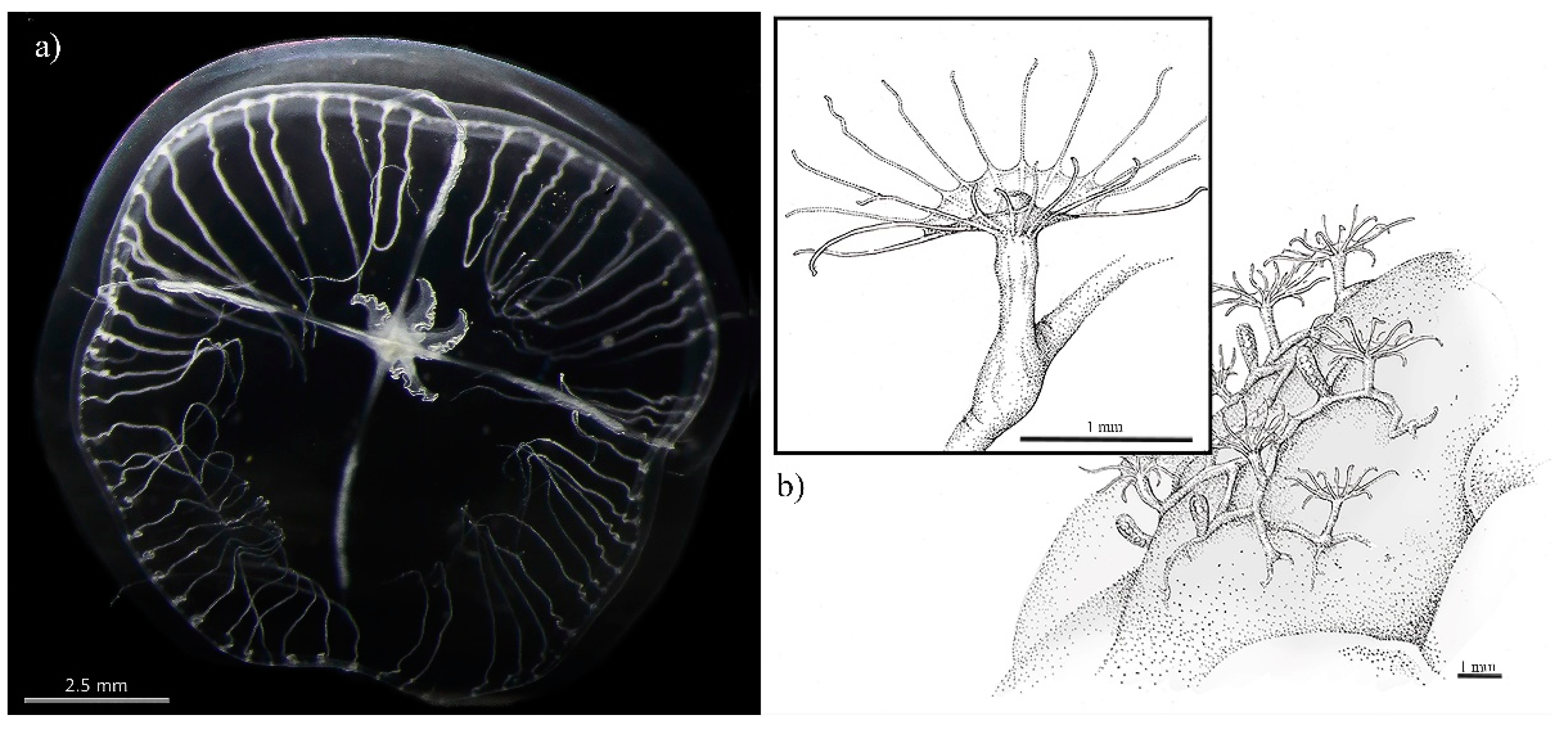
2.4.3. Blackfordia virginica
3. Comb Jellies (Ctenophora)
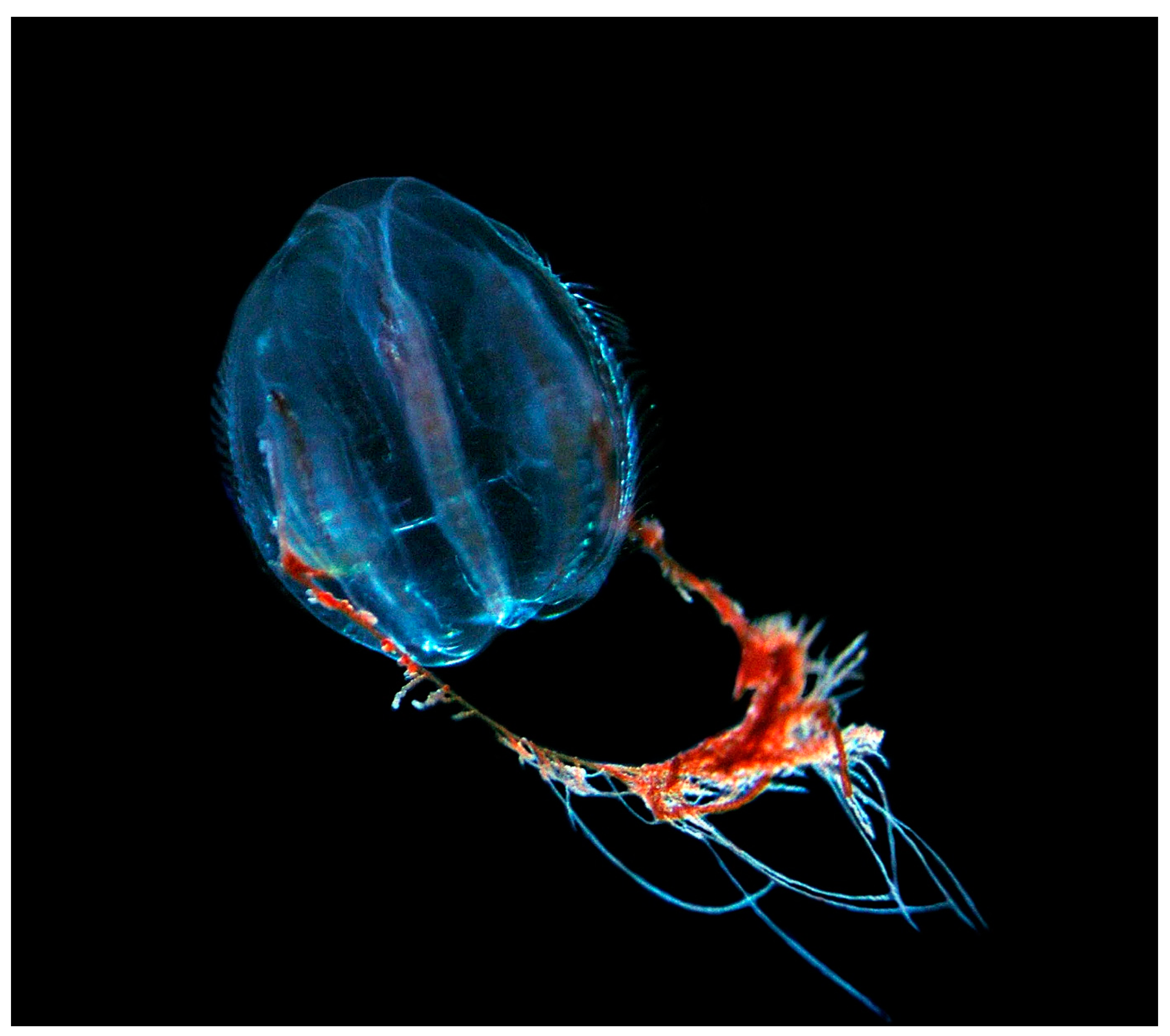
3.1. Sea Walnut Mnemiopsis leidyi (Agassiz, 1860)
3.1.1. Importance for the Baltic Sea
3.1.2. Distribution in the Baltic
3.1.3. Environmental Tolerances and Preferences
3.2. Sea Nut Mertensia ovum (Fabricius, 1780)
3.2.1. Distribution in the Baltic
3.2.2. Importance for the Baltic Sea
3.2.3. Distribution in the Baltic
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ojaveer, H.; Jaanus, A.; Mackenzie, B.R.; Martin, G.; Olenin, S.; Radziejewska, T.; Telesh, I.; Zettler, M.L.; Zaiko, A. Status of biodiversity in the Baltic Sea. PLoS ONE 2010, 5, e12467. [Google Scholar] [CrossRef] [PubMed]
- Möller, H. Significance of coelenterates in relation to other plankton organisms. Meeresforsch. Rep. Mar. Res. 1979, 27, 1–18. [Google Scholar]
- Möller, H. A summer survey of large zooplankton, particularly Scyphomedusae in North Sea and Baltic. Meeresforsch. 1980, 28, 61–68. [Google Scholar]
- Lucas, C.; Gelcich, S.; Uye, S.-i. Living with Jellyfish: Management and adaptation strategies. In Jellyfish Blooms; Pitt, K., Lucas, C., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 129–150. [Google Scholar]
- Hinrichsen, H.-H.; Piatkowski, U.; Jaspers, C. Sightings of extraordinary marine species in the SW Baltic Sea linked to saline water inflows. J. Sea Res. under review.
- Lischka, M. Abundanz, Verteilung und Nahrungsökologie von Scyphomedusen in der zentralen Ostsee. Master’s Thesis, Institut für Meereskunde, University of Kiel, Kiel, Germany, 31 August 1999; p. 91. [Google Scholar]
- Barz, K.; Hinrichsen, H.-H.; Hirche, H.-J. Scyphozoa in the Bornholm Basin (central Baltic Sea)—The role of advection. J. Mar. Syst. 2006, 60, 167–176. [Google Scholar] [CrossRef]
- Barz, K.; Hirche, H.J. Seasonal development of scyphozoan medusae and the predatory impact of Aurelia aurita on the zooplankton community in the Bornholm Basin (central Baltic Sea). Mar. Biol. 2005, 147, 465–476. [Google Scholar] [CrossRef]
- Majaneva, S.; Setala, O.; Gorokhova, E.; Lehtiniemi, M. Feeding of the Arctic ctenophore Mertensia ovum in the Baltic Sea: Evidence of the use of microbial prey. J. Plankton Res. 2014, 36, 91–103. [Google Scholar] [CrossRef]
- Gorokhova, E.; Lehtiniemi, M.; Viitasalo-Frösen, S.; Haddock, S.H.D. Molecular evidence for the occurrence of ctenophore Mertensia ovum in the northern Baltic Sea and implications for the status of the Mnemiopsis leidyi invasion. Limnol. Oceanogr. 2009, 54, 2025–2033. [Google Scholar] [CrossRef]
- Lehtiniemi, M.; Gorokhova, E.; Bolte, S.; Haslob, H.; Huwer, B.; Katajisto, T.; Lennuk, L.; Majaneva, S.; Pollumaee, A.; Schaber, M.; et al. Distribution and reproduction of the Arctic ctenophore Mertensia ovum in the Baltic Sea. Mar. Ecol. Prog. Ser. 2013, 491, 111–124. [Google Scholar] [CrossRef]
- Jaspers, C.; Haraldsson, M.; Bolte, S.; Reusch, T.B.H.; Thygesen, U.H.; Kiørboe, T. Ctenophore population recruits entirely through larval reproduction in the central Baltic Sea. Biol. Lett. 2012, 8, 809–812. [Google Scholar] [CrossRef]
- Jaspers, C.; Haraldsson, M.; Lombard, F.; Bolte, S.; Kiørboe, T. Seasonal dynamics of early life stages of invasive and native ctenophores give clues to invasion and bloom potential in the Baltic Sea. J. Plankton Res. 2013, 35, 582–594. [Google Scholar]
- Kube, S.; Postel, L.; Honnef, C.; Augustin, C.B. Mnemiopsis leidyi in the Baltic Sea—Distribution and overwintering between autumn 2006 and spring 2007. Aquat. Invasions 2007, 2, 137–145. [Google Scholar] [CrossRef]
- Jaspers, C.; Huwer, B.; Weiland-Bräuer, N.; Clemmesen, C. First record of the non-indigenous jellyfish Blackfordia virginica (Mayer, 1910) in the Baltic Sea. Helgol. Mar. Res. 2018, 72, 13. [Google Scholar] [CrossRef]
- Shiganova, T.A.; Riisgård, H.U.; Ghabooli, S.; Tendal, O.S. First report on Beroe ovata in an unusual mixture of ctenophores in the Great Belt (Denmark). Aquat. Invasions 2014, 9, 111–116. [Google Scholar] [CrossRef]
- Fennel, W.; Sturm, M. Dynamics of the Western Baltic. J. Mar. Syst. 1992, 3, 183–205. [Google Scholar] [CrossRef]
- Lehmann, A.; Krauss, W.; Hinrichsen, H.H. Effects of remote and local atmospheric forcing on circulation and upwelling in the Baltic Sea. Tellus A Dyn. Meteorol. Oceanol. 2002, 54, 299–316. [Google Scholar] [CrossRef]
- Christensen, K.H.; Sperrevik, A.K.; Brostrom, G. On the variability in the onset of the Norwegian Coastal Current. J. Phys. Oceanogr. 2018, 48, 723–738. [Google Scholar] [CrossRef]
- Hinrichsen, H.-H.; Huwer, B.; Makarchouk, A.; Petereit, C.; Schaber, M.; Voss, R. Climate-driven long-term trends in Baltic Sea oxygen concentrations and the potential consequences for eastern Baltic cod (Gadus morhua). ICES J. Mar. Sci. 2011, 68, 2019–2028. [Google Scholar] [CrossRef]
- Schinke, H.; Matthäus, W. On the causes of major Baltic inflows—An analysis of long time series. Cont. Shelf Res. 1998, 18, 67–97. [Google Scholar] [CrossRef]
- Saderne, V.; Fietzek, P.; Herman, P.M.J. Extreme variations of pCO2 and pH in a macrophyte meadow of the Baltic Sea in summer: Evidence of the effect of photosynthesis and local upwelling. PLoS ONE 2013, 8, e62689. [Google Scholar]
- Lehmann, A.; Myrberg, K. Upwelling in the Baltic Sea—A review. J. Mar. Syst. 2008, 74, 3–12. [Google Scholar] [CrossRef]
- Köster, F.W.; Möllmann, C.; Hinrichsen, H.-H.; Wieland, K.; Tomkiewicz, J.; Kraus, G.; Voss, R.; Makarchouk, A.; MacKenzie, B.R.; St. John, M.A.; et al. Baltic cod recruitment—The impact of climate variability on key processes. ICES J. Mar. Sci. 2005, 62, 1408–1425. [Google Scholar] [CrossRef]
- Köster, F.W.; Huwer, B.; Hinrichsen, H.H.; Neumann, V.; Makarchouk, A.; Eero, M.; Dewitz, B.v.; Hüssy, K.; Tomkiewicz, J.; Margonski, P.; et al. Eastern Baltic cod recruitment revisited—Dynamics and impacting factors. ICES J. Mar. Sci. 2017, 74, 3–19. [Google Scholar] [CrossRef]
- MacKenzie, B.R.; Hinrichsen, H.H.; Plikshs, M.; Wieland, K.; Zezera, A.S. Quantifying environmental heterogeneity: Habitat size necessary for successful development of cod Gadus morhua eggs in the Baltic Sea. Mar. Ecol. Prog. Ser. 2000, 193, 143–156. [Google Scholar] [CrossRef]
- Breitburg, D.L.; Adamack, A.; Rose, K.A.; Kolesar, S.E.; Decker, M.B.; Purcell, J.E.; Keister, J.E.; Cowan, J.H. The pattern and influence of low dissolved oxygen in the Patuxent River, a seasonally hypoxic estuary. Estuaries 2003, 26, 280–297. [Google Scholar] [CrossRef]
- BACC. Assessment of climate change for the Baltic Sea basin. In Regional Climate Studies, 1st ed.; Springer: Berlin, Germany, 2008; Volume 1, 474p. [Google Scholar]
- Leppäranta, M.; Myrberg, K. Physical Oceanography of the Baltic Sea, 1st ed.; Springer: Berlin, Germany, 2009; 400p. [Google Scholar]
- BACC. Second assessment of climate change for the Baltic Sea basins. In Regional Climate Studies, 2nd ed.; Bolle, H.J., Menenti, M., Ichtiaque Rasool, S., Eds.; Springer Verlag: Geesthacht, Germany, 2015; Volume 2, 515p. [Google Scholar]
- Lehmann, A.; Getzlaff, K.; Harlass, J. Detailed assessment of climate variability of the Baltic Sea area for the period 1958–2009. Clim. Res. 2011, 46, 185–196. [Google Scholar] [CrossRef]
- Carstensen, J.; Andersen, J.H.; Gustafsson, B.G.; Conley, D.J. Deoxygenation of the Baltic Sea during the last century. Proc. Natl. Acad. Sci. USA 2014, 111, 5628–5633. [Google Scholar] [CrossRef]
- Brotz, L.; Cheung, W.W.L.; Kleisner, K.; Pakhomov, E.; Pauly, D. Increasing jellyfish populations: Trends in large marine ecosystems. Hydrobiologia 2012, 690, 3–20. [Google Scholar] [CrossRef]
- Flynn, B.; Richardson, A.; Brierley, A.; Boyer, D.C.; Axelsen, B.E.; Scott, L.; Moroff, N.; Kainge, P.; Tjizoo, B.M.; Gibbons, M. Temporal and spatial patterns in the abundance of jellyfish in the northern Benguela upwelling ecosystem and their link to thwarted pelagic fishery recovery. Afr. J. Mar. Sci. 2012, 34, 131–146. [Google Scholar] [CrossRef]
- Lynam, C.P.; Gibbons, M.J.; Axelsen, B.E.; Sparks, C.A.; Coetzee, J.; Heywood, B.G.; Brierley, A.S. Jellyfish overtake fish in a heavily fished ecosystem. Curr. Biol. 2006, 16, R492–R493. [Google Scholar] [CrossRef]
- Condon, R.H.; Duarte, C.M.; Pitt, K.A.; Robinson, K.L.; Lucas, C.H.; Sutherland, K.R.; Mianzan, H.W.; Bogeberg, M.; Purcell, J.E.; Decker, M.B.; et al. Recurrent jellyfish blooms are a consequence of global oscillations. Proc. Natl. Acad. Sci. USA 2013, 2012, 1000–1005. [Google Scholar] [CrossRef] [PubMed]
- Condon, R.H.; Graham, W.; Pitt, K.; Cathy, H. Questioning the rise of gelatinous zooplankton in the world’s oceans. Bioscience 2012, 62, 160–169. [Google Scholar] [CrossRef]
- Richardson, A.J.; Bakun, A.; Hays, G.C.; Gibbons, M.J. The jellyfish joyride: Causes, consequences and management responses to a more gelatinous future. Trends Ecol. Evol. 2009, 24, 312–322. [Google Scholar] [CrossRef] [PubMed]
- West, E.J.; Welsh, D.T.; Pitt, K.A. Influence of decomposing jellyfish on the sediment oxygen demand and nutrient dynamics. Hydrobiologia 2009, 616, 151–160. [Google Scholar] [CrossRef]
- Møller, L.F.; Riisgard, H.U. Population dynamics, growth and predation impact of the common jellyfish Aurelia aurita and two hydromedusae, Sarsia tubulosa, and Aequorea vitrina in Limfjorden (Denmark). Mar. Ecol. Prog. Ser. 2007, 346, 153–165. [Google Scholar] [CrossRef]
- Duarte, C.M.; Pitt, K.A.; Lucas, C.H.; Purcell, J.E.; Uye, S.-i.; Robinson, K.; Brotz, L.; Decker, M.B.; Sutherland, K.R.; Malej, A.; et al. Is global ocean sprawl a cause of jellyfish blooms? Front. Ecol. Environ. 2013, 11, 91–97. [Google Scholar] [CrossRef]
- Janßen, H.; Augustin, C.B.; Hinrichsen, H.H.; Kube, S. Impact of secondary hard substrate on the distribution and abundance of Aurelia aurita in the western Baltic Sea. Mar. Pollut. Bull. 2013, 75, 224–234. [Google Scholar] [CrossRef]
- Jaspers, C.; Huwer, B.; Antajan, E.; Hosia, A.; Hinrichsen, H.-H.; Biastoch, A.; Angel, D.; Asmus, R.; Augustin, C.; Bagheri, S.; et al. Ocean current connectivity propelling the secondary spread of a marine invasive comb jelly across western Eurasia. Global Ecol. Biogeogr. 2018, 27, 814–827. [Google Scholar] [CrossRef]
- Tiselius, P.; Møller, E.F. Community cascades in a marine pelagic food web controlled by the non-visual apex predator Mnemiopsis leidyi. J. Plankton Res. 2017, 39, 271–279. [Google Scholar]
- Holst, S. Morphology and development of benthic and pelagic life stages of North Sea jellyfish (Scyphozoa, Cnidaria) with special emphasis on the identification of ephyra stages. Mar. Biol. 2012, 159, 2707–2722. [Google Scholar] [CrossRef]
- Thein, H.; Ikeda, H.; Uye, S.-I. The potential role of podocysts in perpetuation of the common jellyfish Aurelia aurita s.l. (Cnidaria: Scyphozoa) in anthropogenically perturbed coastal waters. Hydrobiologia 2012, 690, 157–167. [Google Scholar] [CrossRef]
- Schneider, G.; Behrends, G. Top-down control in a neritic plankton system by Aurelia aurita medusae—A summary. Ophelia 1998, 48, 71–82. [Google Scholar] [CrossRef]
- Möller, H. Reduction of a larval Herring population by jellyfish predator. Science 1984, 224, 621–622. [Google Scholar] [CrossRef] [PubMed]
- Hansson, L.J.; Moeslund, O.; Kiørboe, T.; Riisgård, H.U. Clearance rates of jellyfish and their potential predation impact on zooplankton and fish larvae in a neritic ecosystem (Limfjorden, Denmark). Mar. Ecol. Prog. Ser. 2005, 304, 117–131. [Google Scholar] [CrossRef]
- Schneider, G.; Behrends, G. Population dynamics and the trophic role of Aurelia aurita medusae in the Kiel Bight and Western Baltic. ICES J. Mar. Sci. 1994, 51, 359–367. [Google Scholar] [CrossRef]
- Gröndahl, F. A comparative ecological study on the scyphozoans Aurelia aurita, Cyanea capillata and Cyanea lamarckii in the Gullmar Fjord, Western Sweden, 1982–1986. Mar. Biol. 1988, 97, 541–550. [Google Scholar] [CrossRef]
- Hosia, A.; Granhag, L.; Katajisto, T.; Lehtiniemi, M. Experimental feeding rates of gelatinous predators Aurelia aurita and Mnemiopsis leidyi at low northern Baltic Sea salinity. Boreal Environ. Res. 2012, 17, 473–483. [Google Scholar]
- Wikström, D.A. Beobachtungen über die Ohrenqualle (Aurelia aurita L.) in den Schären SW-Finnlands. Mem. Soc. Pro Fauna Flora Fennica 1932, 8, 14–17. [Google Scholar]
- Riisgård, H.U.; Barth-Jensen, C.; Madsen, C.V. High abundance of the jellyfish Aurelia aurita excludes the invasive ctenophore Mnemiopsis leidyi to establish in a shallow cove (Kertinge Nor, Denmark). Aquat. Invasions 2010, 5, 347–356. [Google Scholar] [CrossRef]
- Olesen, N.J.; Frandsen, K.; Riisgard, H.U. Population dynamics, growth and energetics of jellyfish Aurelia aurita in a shallow fjord. Mar. Ecol. Prog. Ser. 1994, 105, 9–18. [Google Scholar] [CrossRef]
- Thiel, H. Untersuchungen über die Strobilisation von Aurelia aurita LAM. an einer Population der Kieler Förde. Kiel Meeresforsch. 1962, 18, 198–230. [Google Scholar]
- Janas, U.; Witek, Z. The occurrence of medusae in the Baltic and their importance in the ecosystem, with special emphasis on Aurelia aurita. Oceanologia 1993, 34, 69–84. [Google Scholar]
- Thill, H. Beiträge zur Kenntnis der Aurelia aurita (L.). Z. Wiss. Zool. 1937, 150, 51–96. [Google Scholar]
- Olesen, N.J. Clearance potential of jellyfish Aurelia aurita and predation impact on zooplankton in a shallow cove. Mar. Ecol. Prog. Ser. 1995, 124, 63–72. [Google Scholar] [CrossRef]
- Möller, H. Population dynamics of Aurelia aurita medusae in Kiel Bight, Germany. Mar. Biol. 1980, 60, 123–128. [Google Scholar] [CrossRef]
- Dam, H.G. Evolutionary adaptation of marine zooplankton to global change. Ann. Rev. Mar. Sci. 2013, 5, 349–370. [Google Scholar] [CrossRef]
- Båmstedt, U.; Ishii, H.; Martinussen, M.B. Is the scyphomedusa Cyanea capillata (L.) dependent on gelatinous prey for its early development? Sarsia 1997, 82, 269–273. [Google Scholar] [CrossRef]
- Gröndahl, F.; Hernroth, L. Release and growth of Cyanea capillata (L) ephyra in the Gullmar Fjord, Western Sweden. J. Exp. Mar. Biol. Ecol. 1987, 106, 91–101. [Google Scholar] [CrossRef]
- Haahtela, H.; Lassig, J. Records of Cyanea capillata (Scyphozoa) and Hyperia galba (Amphipoda) from the Gulf of Finland and the northern Baltic. Ann. Zool. Fenn. 1967, 4, 469. [Google Scholar]
- Holst, S.; Jarms, G. Effects of low salinity on settlement and strobilation of scyphozoa (Cnidaria): Is the lion’s mane Cyanea capillata (L.) able to reproduce in the brackish Baltic Sea? Hydrobiologia 2010, 645, 53–68. [Google Scholar] [CrossRef]
- Verwey, J. Die Periodizität im Auftreten und die aktiven und passiven Bewegungen der Quallen. Arch. Neerland. Zool. 1942, 6, G3–G468. [Google Scholar] [CrossRef]
- Freire, M.; Genzano, G.N.; Neumann-Leitao, S.; Perez, C.D. The non-indigenous medusa Blackfordia virginica (Hydrozoa, Leptothecata) in tropical Brazil: 50 years of unnoticed presence. Biol. Invasions 2014, 16, 1–5. [Google Scholar] [CrossRef]
- Moore, S.J. Redescription of the leptomedusan Blackfordia virginica. J. Mar. Biol. Assoc. 1987, 67, 287–291. [Google Scholar] [CrossRef]
- Baumsteiger, J.; O’Rear, T.A.; Cook, J.D.; Manfree, A.D.; Moyle, P.B. Factors affecting distribution and abundance of jellyfish medusae in a temperate estuary: A multi-decadal study. Biol. Invasions 2018, 20, 105–119. [Google Scholar] [CrossRef]
- Wintzer, A.P.; Meek, M.H.; Moyle, P.B. Abundance, size, and diel feeding ecology of Blackfordia virginica (Mayer, 1910), a non-native hydrozoan in the lower Napa and Petaluma Rivers, California (USA). Aquat. Invasions 2013, 8, 147–156. [Google Scholar] [CrossRef]
- Marques, F.; Angelico, M.M.; Costa, J.L.; Teodosio, M.A.; Presado, P.; Fernandes, A.; Chainho, P.; Domingos, I. Ecological aspects and potential impacts of the non-native hydromedusa Blackfordia virginica in a temperate estuary. Estuar. Coast. Shelf Sci. 2017, 197, 69–79. [Google Scholar] [CrossRef]
- Von Dorrien, C.; Hammer, C.; Zimmermann, C.; Stepputtis, D.; Stuermer, I.W.; Kotterba, P.; Polte, P. A review on the Herring Clupea harengus (Actinopterygii: Clupeiformes: Clupeidae) recruitment and early life stage ecology in the western Baltic Sea. Acta Ichthyol. Pisc. 2013, 43, 169–182. [Google Scholar] [CrossRef]
- Mayer, A.G. Medusae of the world: The hydromedusae, 1st ed.; Carnegie Institution of Washington: Washington, DC, USA, 1910; Volume 109, 230p. [Google Scholar]
- Harrison, G.F.; Kim, K.; Collins, A.G. Low genetic diversity of the putatively introduced, brackish water hydrozoan, Blackfordia virginica (Leptothecata: Blackfordiidae), throughout the United States, with a new record for Lake Pontchartrain, Louisiana. Proc. Biol. Soc. Wash. 2013, 126, 91–102. [Google Scholar] [CrossRef]
- Thiel, M.E. Zur Kenntnis der Hydromeduseanfauna des Schwarzen Meeres. Zool. Anz. 1935, 11, 160–175. [Google Scholar]
- Kramp, P.L. Hydromedusae in the Indian Museum. Rec. Indian Mus. 1958, 53, 339–376. [Google Scholar]
- Vannucci, M.; Santhakumari, V.; Dos Santos, E.P. The ecology of hydromedusae from the Cochin area. Mar. Biol. 1970, 7, 49–58. [Google Scholar] [CrossRef]
- Mills, C.E.; Rees, J.T. New observations and corrections concerning the trio of invasive hydromedusae Maeotias marginata, (= M. inexpectata), Blackfordia virginica, and Moerisia sp in the San Francisco Estuary. Scientia Marina 2000, 64, 151–155. [Google Scholar] [CrossRef]
- Mills, C.E.; Sommer, F. Invertebrate introductions in marine habitats—Two species of hydromedusae (Cnidaria) natibe to the Black Sea, Maeotias inexspectata and Blackfordia virginica invade San Francisco Bay. Mar. Biol. 1995, 122, 279–288. [Google Scholar]
- Bardi, J.; Marques, A.C. The invasive hydromedusae Blackfordia virginica Mayer, 1910 (Cnidaria: Blackfordiidae) in southern Brazil, with comments on taxonomy and distribution of the genus Blackfordia. Zootaxa 2009, 41–50. [Google Scholar] [CrossRef]
- Buecher, E.; Goy, J.; Gibbons, M. Hydromedusae of the Agulhas current. Afr. Invertebr. 2005, 46, 27–69. [Google Scholar]
- Denayer, J.C. 3 new or little known medusae of french coasts—Maeotis inexpectata, Ostrooumov, 1896; Blackfordia virginica, Mayer, 1910; Nemopsis bachei, Agassiz, 1849. Cah. Biol. Mar. 1973, 14, 285–294. [Google Scholar]
- Faasse, M.; Melchers, M. The exotic jellyfish Blackfordia virginica introduced into the Netherlands (Cnidaria: Hydrozoa). Nederlandse Faunist. Mededel. 2014, 43, 103–110. [Google Scholar]
- Nowaczyk, A.; David, V.; Lepage, M.; Goarant, A.; De Oliveira, E.; Sautour, B. Spatial and temporal patterns of occurrence of three alien hydromedusae, Blackfordia virginica (Mayer, 1910), Nemopsis bachei (Agassiz, 1849) and Maeotias marginata (Modeer, 1791), in the Gironde Estuary (France). Aquat. Invasions 2016, 11, 397–409. [Google Scholar] [CrossRef]
- Santhakumari, V.; Ramaiah, N.; Nair, V.R. Ecology of hydromedusae from Bombay Harbour–Thana and Bassein Creek estuarine complex. Indian J. Mar. Sci. 1997, 26, 162–168. [Google Scholar]
- Marques, F.; Chainho, P.; Costa, J.L.; Domingos, I.; Angelico, M.M. Abundance, seasonal patterns and diet of the non-native jellyfish Blackfordia virginica in a Portuguese estuary. Estuar. Coast. Shelf Sci. 2015, 167, 212–219. [Google Scholar] [CrossRef]
- Russel, R. The Medusae of the British Isles. II Pelagic Scyphozoa with a Supplement to the 1st Volume on Hydromedusae, 2nd ed.; Cambridge University Press: Cambridge, UK, 1970; 284p. [Google Scholar]
- Dawson, M.N. Macro-morphological variation among cryptic species of the moon jellyfish, Aurelia (Cnidaria: Scyphozoa). Mar. Biol. 2003, 143, 369–379. [Google Scholar] [CrossRef]
- Brewer, R.H. Larval settling behavior in Cyanea capillata (Cnidaria: Scyphozoa). Biol. Bull. 1976, 150, 183–199. [Google Scholar] [CrossRef]
- Jaspers, C.; Costello, J.H.; Colin, S.P. Carbon content of Mnemiopsis leidyi eggs and specific egg production rates in northern Europe. J. Plankton Res. 2015, 37, 11–15. [Google Scholar] [CrossRef]
- Colin, S.P.; Costello, J.H.; Hansson, L.J.; Titelman, J.; Dabiri, J.O. Stealth predation and the predatory success of the invasive ctenophore Mnemiopsis leidyi. Proc. Natl. Acad. Sci. USA 2010, 107, 17223–17227. [Google Scholar] [CrossRef] [PubMed]
- Colin, S.P.; MacPherson, S.; Gemmell, B.; Costello, J.H.; Sutherland, K.R.; Jaspers, C. Elevating the predatory effect: Sensory-scanning foraging strategy by the lobate ctenophore Mnemiopsis leidyi. Limnol. Oceanogr. 2015, 60, 100–109. [Google Scholar] [CrossRef]
- Riisgård, H.U.; Bøttiger, L.; Madsen, C.; Purcell, J. Invasive ctenophore Mnemiopsis leidyi in Limfjorden (Denmark) in late summer 2007—Assessment of abundance and predation effects. Aquat. Invasions 2007, 2, 395–401. [Google Scholar] [CrossRef]
- Haraldsson, M.; Jaspers, C.; Tiselius, P.; Aksnes, D.L.; Andersen, T.; Titelman, J. Environmental constraints of the invasive Mnemiopsis leidyi in Scandinavian waters. Limnol. Oceanogr. 2013, 58, 37–48. [Google Scholar] [CrossRef]
- Costello, J.H.; Bayha, K.M.; Mianzan, H.W.; Shiganova, T.A.; Purcell, J.E. Transitions of Mnemiopsis leidyi (Ctenophora: Lobata) from a native to an exotic species: A review. Hydrobiologia 2012, 690, 21–46. [Google Scholar] [CrossRef]
- Schaber, M.; Haslob, H.; Huwer, B.; Harjes, A.; Hinrichsen, H.H.; Köster, F.W.; Storr-Paulsen, M.; Schmidt, J.O.; Voss, R. The invasive ctenophore Mnemiopsis leidyi in the central Baltic Sea: Seasonal phenology and hydrographic influence on spatio-temporal distribution patterns. J. Plankton Res. 2011, 33, 1053–1065. [Google Scholar] [CrossRef]
- Schaber, M.; Haslob, H.; Huwer, B.; Harjes, A.; Hinrichsen, H.-H.; Storr-Paulsen, M.; Schmidt, J.O.; Voss, R.; Neumann, V.; Köster, F.W. Spatio-temporal overlap of the alien invasive ctenophore Mnemiopsis leidyi and ichthyoplankton in the Bornholm Basin (Baltic Sea). Biol. Invasions 2011, 13, 2647–2660. [Google Scholar] [CrossRef]
- Huwer, B.; Storr-Paulsen, M.; Riisgård, H.U.; Haslob, H. Abundance, horizontal and vertical distribution of the invasive ctenophore Mnemiopsis leidyi in the central Baltic Sea, November 2007. Aquat. Invasions 2008, 3, 113–124. [Google Scholar] [CrossRef]
- Haslob, H.; Clemmesen, C.; Schaber, M.; Hinrichsen, H.; Schmidt, J.; Voss, R.; Kraus, G.; Köster, F. Invading Mnemiopsis leidyi as a potential threat to Baltic fish. Mar. Ecol. Prog. Ser. 2007, 349, 303–306. [Google Scholar] [CrossRef]
- Haraldsson, M.; Båmstedt, U.; Tiselius, P.; Titelman, J.; Aksnes, D.L. Evidence of diel vertical migration in Mnemiopsis leidyi. PLoS ONE 2014, 9, 10. [Google Scholar] [CrossRef] [PubMed]
- Jaspers, C.; Costello, J.; Sutherland, K.; Gemmell, B.; Lucas, K.; Tackett, J.; Dodge, K.; Colin, S. Resilience in moving water: Effects of turbulence on the predatory impact of the lobate ctenophore Mnemiopsis leidyi. Limnol. Oceanogr. 2018, 63, 445–458. [Google Scholar] [CrossRef]
- Mianzan, H.W.; Martos, P.; Costello, J.H.; Guerrero, R.A. Avoidance of hydrodynamically mixed environments by Mnemiopsis leidyi (Ctenophora: Lobata) in open-sea populations from Patagonia, Argentina. Hydrobiologia 2010, 645, 113–124. [Google Scholar] [CrossRef]
- Sutherland, K.R.; Costello, J.H.; Colin, S.P.; Dabiri, J.O. Ambient fluid motions influence swimming and feeding by the ctenophore Mnemiopsis leidyi. J. Plankton Res. 2014, 36, 1310–1322. [Google Scholar] [CrossRef]
- Jaspers, C.; Titelman, J.; Hansson, L.J.; Haraldsson, M.; Røllike, C. The invasive ctenophore Mnemiopsis leidyi poses no direct threat to Baltic cod eggs and larvae. Limnol. Oceanogr. 2011, 56, 431–439. [Google Scholar] [CrossRef]
- Riisgård, H.U.; Jaspers, C.; Serre, S.; Lundgreen, K. Occurence, inter-annual variability and zooplankton-predation impact of the invasive ctenophore Mnemiopsis leidyi and the native jellyfish Aurelia aurita in Limfjorden (Denmark) in 2010 and 2011. BioInvasions Rec. 2012, 1, 145–159. [Google Scholar] [CrossRef]
- Javidpour, J.; Molinero, J.C.; Peschutter, J.; Sommer, U. Seasonal changes and population dynamics of the ctenophore Mnemiopsis leidyi after its first year of invasion in the Kiel Fjord, Western Baltic Sea. Biol. Invasions 2009, 11, 873–882. [Google Scholar] [CrossRef]
- Jaspers, C.; Møller, L.F.; Kiørboe, T. Salinity gradient of the Baltic Sea limits the reproduction and population expansion of the newly invaded comb jelly Mnemiopsis leidyi. PLoS ONE 2011, 6, e24065. [Google Scholar] [CrossRef]
- Lehtiniemi, M.; Lehmann, A.; Javidpour, J.; Myrberg, K. Spreading and physico-biological reproduction limitations of the invasive American comb jelly Mnemiopsis leidyi in the Baltic Sea. Biol. Invasions 2011, 14, 341–354. [Google Scholar] [CrossRef]
- Gambill, M.; Møller, L.F.; Peck, M.A. Effects of temperature on the feeding and growth of the larvae of the invasive ctenophore Mnemiopsis leidyi. J. Plankton Res. 2015, 37, 1001–1005. [Google Scholar] [CrossRef]
- Costello, J.H.; Sullivan, B.K.; Gifford, D.J.; Van Keuren, D.; Sullivan, L.J. Seasonal refugia, shoreward thermal amplification, and metapopulation dynamics of the ctenophore Mnemiopsis leidyi in Narragansett Bay, Rhode Island. Limnol. Oceanogr. 2006, 51, 1819–1831. [Google Scholar] [CrossRef]
- Jaspers, C.; Møller, E.F.; Kiørboe, T. Reproduction rates under variable food conditions and starvation in Mnemiopsis leidyi: Significance for the invasion success of a ctenophore. J. Plankton Res. 2015, 37, 1011–1018. [Google Scholar] [CrossRef]
- Jaspers, C.; Marty, L.; Kiørboe, T. Selection for life-history traits to maximize population growth in an invasive marine species. Global Chang. Biol. 2018, 24, 1164–1174. [Google Scholar] [CrossRef] [PubMed]
- Javidpour, J.; Molinero, J.-C.; Ramírez-Romero, E.; Roberts, P.; Larsen, T. Cannibalism makes invasive comb jelly, Mnemiopsis leidyi, resilient to unfavourable conditions. Comm. Biol. 2020, 3, 212. [Google Scholar] [CrossRef]
- Granhag, L.; Hosia, A. Feeding and starvation in the native ctenophore Bolinopsis infundibulum and the introduced Mnemiopsis leidyi in the North Sea—Implications for ctenophore transport in ships’ ballast water. J. Plankton Res. 2015, 37, 1006–1010. [Google Scholar] [CrossRef][Green Version]
- Kolesar, S.E.; Breitburg, D.L.; Purcell, J.E.; Decker, M.B. Effects of hypoxia on Mnemiopsis leidyi, ichthyoplankton and copepods: Clearance rates and vertical habitat overlap. Mar. Ecol. Prog. Ser. 2010, 411, 173–188. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jaspers, C.; Bezio, N.; Hinrichsen, H.-H. Diversity and Physiological Tolerance of Native and Invasive Jellyfish/Ctenophores along the Extreme Salinity Gradient of the Baltic Sea. Diversity 2021, 13, 57. https://doi.org/10.3390/d13020057
Jaspers C, Bezio N, Hinrichsen H-H. Diversity and Physiological Tolerance of Native and Invasive Jellyfish/Ctenophores along the Extreme Salinity Gradient of the Baltic Sea. Diversity. 2021; 13(2):57. https://doi.org/10.3390/d13020057
Chicago/Turabian StyleJaspers, Cornelia, Nicholas Bezio, and Hans-Harald Hinrichsen. 2021. "Diversity and Physiological Tolerance of Native and Invasive Jellyfish/Ctenophores along the Extreme Salinity Gradient of the Baltic Sea" Diversity 13, no. 2: 57. https://doi.org/10.3390/d13020057
APA StyleJaspers, C., Bezio, N., & Hinrichsen, H.-H. (2021). Diversity and Physiological Tolerance of Native and Invasive Jellyfish/Ctenophores along the Extreme Salinity Gradient of the Baltic Sea. Diversity, 13(2), 57. https://doi.org/10.3390/d13020057