
Elevational Patterns of Blowfly Parasitism in Two Hole Nesting Avian Species


Abstract
1. Introduction
2. Materials and Methods
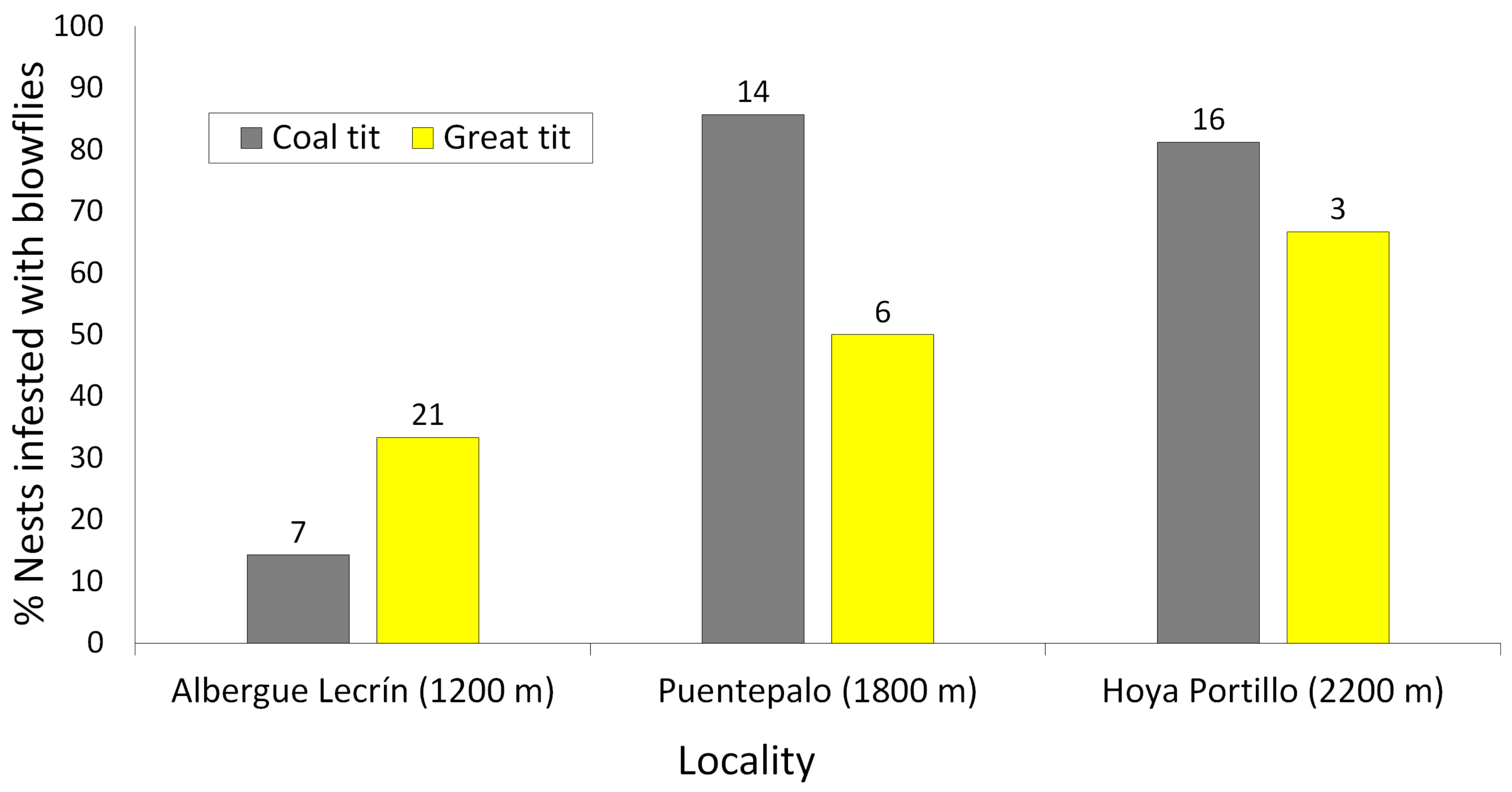
3. Results
4. Discussion
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Ethics Statement
Appendix A

{kind=link}
{kind=link}
| Nest ID | Locality | P. azurea Pupae | Host Species |
|---|---|---|---|
| A02 | Albergue de Lecrín | 0 | Parus major |
| A03 | Albergue de Lecrín | 22 | Parus major |
| A06 | Albergue de Lecrín | 0 | Parus major |
| A07 | Albergue de Lecrín | 23 | Parus major |
| A09 | Albergue de Lecrín | 0 | Periparus ater |
| A09 | Albergue de Lecrín | 0 | Parus major |
| A10 | Albergue de Lecrín | 0 | Periparus ater |
| A12 | Albergue de Lecrín | 0 | Parus major |
| A13 | Albergue de Lecrín | 0 | Parus major |
| A15 | Albergue de Lecrín | 0 | Periparus ater |
| A16 | Albergue de Lecrín | 17 | Parus major |
| A18 | Albergue de Lecrín | 0 | Parus major |
| A19 | Albergue de Lecrín | 0 | Parus major |
| A20 | Albergue de Lecrín | 0 | Parus major |
| A27 | Albergue de Lecrín | 0 | Parus major |
| A28 | Albergue de Lecrín | 0 | Parus major |
| A33 | Albergue de Lecrín | 0 | Periparus ater |
| A38 | Albergue de Lecrín | 0 | Parus major |
| A42 | Albergue de Lecrín | 0 | Parus major |
| A43 | Albergue de Lecrín | 10 | Parus major |
| A45 | Albergue de Lecrín | 0 | Periparus ater |
| A46 | Albergue de Lecrín | 2 | Parus major |
| A48 | Albergue de Lecrín | 0 | Periparus ater |
| A51 | Albergue de Lecrín | 0 | Parus major |
| A52 | Albergue de Lecrín | 2 | Periparus ater |
| A54 | Albergue de Lecrín | 5 | Parus major |
| A55 | Albergue de Lecrín | 28 | Parus major |
| A60 | Albergue de Lecrín | 0 | Parus major |
| H01 | Hoya del Portillo | 22 | Periparus ater |
| H03 | Hoya del Portillo | 16 | Periparus ater |
| H04 | Hoya del Portillo | 4 | Periparus ater |
| H08 | Hoya del Portillo | 40 | Periparus ater |
| H12 | Hoya del Portillo | 0 | Parus major |
| H13 | Hoya del Portillo | 4 | Periparus ater |
| H14 | Hoya del Portillo | 39 | Periparus ater |
| H19 | Hoya del Portillo | 6 | Periparus ater |
| H26 | Hoya del Portillo | 10 | Periparus ater |
| H34 | Hoya del Portillo | 0 | Periparus ater |
| H36 | Hoya del Portillo | 3 | Periparus ater |
| H37 | Hoya del Portillo | 1 | Parus major |
| H38 | Hoya del Portillo | 5 | Periparus ater |
| H43 | Hoya del Portillo | 0 | Periparus ater |
| H46 | Hoya del Portillo | 0 | Periparus ater |
| H50 | Hoya del Portillo | 2 | Periparus ater |
| H51 | Hoya del Portillo | 20 | Periparus ater |
| H55 | Hoya del Portillo | 24 | Periparus ater |
| H60 | Hoya del Portillo | 4 | Parus major |
| P01 | Puentepalo | 10 | Periparus ater |
| P02 | Puentepalo | 0 | Parus major |
| P03 | Puentepalo | 0 | Parus major |
| P04 | Puentepalo | 7 | Periparus ater |
| P14 | Puentepalo | 1 | Periparus ater |
| P21 | Puentepalo | 0 | Periparus ater |
| P23 | Puentepalo | 2 | Periparus ater |
| P25 | Puentepalo | 9 | Periparus ater |
| P28 | Puentepalo | 0 | Parus major |
| P29 | Puentepalo | 6 | Periparus ater |
| P32 | Puentepalo | 9 | Parus major |
| P33 | Puentepalo | 53 | Periparus ater |
| P36 | Puentepalo | 18 | Periparus ater |
| P43 | Puentepalo | 1 | Periparus ater |
| P46 | Puentepalo | 6 | Periparus ater |
| P53 | Puentepalo | 3 | Periparus ater |
| P54 | Puentepalo | 0 | Periparus ater |
| P57 | Puentepalo | 22 | Periparus ater |
| P59 | Puentepalo | 37 | Parus major |
| P60 | Puentepalo | 13 | Parus major |
References
- Schmid-Hempel, P. Evolutionary Parasitology: The Integrated Study of Infections, Immunology, Ecology, and Genetics; Oxford University Press: Oxford, UK, 2011. [Google Scholar]
- Møller, A.P. Parasitism and the evolution of host life history. In Host-Parasite Evolution: General Principles and Avian Models; Clayton, D.H., Moore, J., Eds.; Oxford University Press: Oxford, UK, 1997; pp. 105–127. [Google Scholar]
- Møller, A.P.; Arriero, E.; Lobato, E.; Merino, S. A meta-analysis of parasite virulence in nestling birds. Biol. Rev. 2009, 84, 567–588. [Google Scholar] [CrossRef]
- Brown, C.R.; Brown, M.B. Ectoparasitism as a cost of coloniality in cliff swallows (Hirundo pyrrhonota). Ecology 1986, 67, 1206–1218. [Google Scholar] [CrossRef]
- Fitze, P.S.; Clobert, J.; Richner, H. Long-term life-history consequences of ectoparasite-modulated growth and development. Ecology 2004, 85, 2018–2026. [Google Scholar] [CrossRef]
- Thomas, D.W.; Shipley, B.; Blondel, J.; Perret, P.; Simon, A.; Lambrechts, M.M. Common paths link food abundance and ectoparasite loads to physiological performance and recruitment in nestling blue tits. Funct. Ecol. 2007, 21, 947–955. [Google Scholar] [CrossRef]
- Richner, H.; Tripet, F. Ectoparasitism and the trade-off between current and future reproduction. Oikos 1999, 86, 535–538. [Google Scholar] [CrossRef]
- Bennett, G.F.; Whitworth, T.L. Studies on the life history of some species of Protocalliphora (Diptera: Calliphoridae). Can. J. Zool. 1991, 69, 2048–2058. [Google Scholar] [CrossRef]
- Veiga, J.; Moreno, E.; Benzal, J.; Valera, F. Off-host longevity of the winged dispersal stage of Carnus hemapterus (Insecta: Diptera) modulated by gender, body size and food provisioning. Parasitology 2019, 146, 241–245. [Google Scholar] [CrossRef]
- Heeb, P.; Kölliker, M.; Richner, H. Bird ectoparasite interactions, nest humidity, and ectoparasite community structure. Ecology 2000, 81, 958–968. [Google Scholar]
- Dawson, R.D.; Hillen, K.K.; Whitworth, T.L. Effects of experimental variation in temperature on larval densities of parasitic Protocalliphora (Diptera: Calliphoridae) in nests of tree swallows (Passeriformes: Hirundinidae). Environ. Entomol. 2005, 34, 563–568. [Google Scholar] [CrossRef]
- Castaño-Vázquez, F.; Martínez, J.; Merino, S.; Lozano, M. Experimental manipulation of temperature reduce ectoparasites in nests of blue tits Cyanistes caeruleus. J. Avian Biol. 2018, 49, e01695. [Google Scholar] [CrossRef]
- Merino, S.; Potti, J. Weather dependent effects of nest ectoparasites on their bird hosts. Ecography 1996, 19, 107–113. [Google Scholar] [CrossRef]
- Stephenson, S.; Hannon, S.; Proctor, H. The function of feathers in tree swallow nests: Insulation or ectoparasite barrier? Condor 2009, 111, 479–487. [Google Scholar] [CrossRef]
- Clayton, D.H.; Koop, J.A.H.; Harbison, C.W.; Moyer, B.R.; Bush, S.E. How birds combat ectoparasites. Open Ornithol. J. 2010, 3, 41–71. [Google Scholar] [CrossRef]
- Bush, S.E.; Clayton, D.H. Anti-parasite behaviour of birds. Philos. Trans. R. Soc. B 2018, 373, 20170196. [Google Scholar] [CrossRef] [PubMed]
- Merino, S. Immunocompetence and parasitism in nestlings from wild populations. Open Ornithol. J. 2010, 3, 27–32. [Google Scholar] [CrossRef][Green Version]
- Owen, J.P.; Nelson, A.C.; Clayton, D.H. Ecological immunology of bird-ectoparasite systems. Trends Parasitol. 2010, 26, 530–539. [Google Scholar] [CrossRef]
- Tschirren, B.; Bischoff, L.L.; Saladin, V.; Richner, H. Host condition and host immunity affect parasite fitness in a bird-ectoparasite system. Funct. Ecol. 2007, 21, 372–378. [Google Scholar] [CrossRef]
- Bize, P.; Jeanneret, C.; Klopfenstein, A.; Roulin, A. What makes a host profitable? Parasites balance host nutritive resources against immunity. Am. Nat. 2008, 171, 107–118. [Google Scholar] [CrossRef]
- DeSimone, J.G.; Clotfelter, E.D.; Black, E.C.; Knutie, S.A. Avoidance, tolerance, and resistance to ectoparasites in nestling and adult tree swallows. J. Avian Biol. 2018, 49, e01641. [Google Scholar] [CrossRef]
- Grab, K.M.; Hiller, B.J.; Hurlbert, J.H.; Ingram, M.E.; Parker, A.B.; Pokutnaya, D.Y.; Knutie, S.A. Host tolerance and resistance to parasitic nest flies differs between two wild bird species. Ecol. Evol. 2019, 9, 12144–12155. [Google Scholar] [CrossRef] [PubMed]
- Knutie, S.A. Food supplementation affects gut microbiota and immunological resistance to parasites in a wild bird species. J. Appl. Ecol. 2020, 57, 536–547. [Google Scholar] [CrossRef]
- Møller, A.P.; Martín-Vivaldi, M.; Merino, S.; Soler, J.J. Density-dependent and geographical variation in bird immune response. Oikos 2006, 115, 463–474. [Google Scholar] [CrossRef]
- Saino, N.; Calza, S.; Møller, A.P. Immunocompetence of nestling barn swallows in relation to brood size and parental effort. J. Anim. Ecol. 1997, 66, 827–836. [Google Scholar] [CrossRef]
- Hoi-Leitner, M.; Romero-Pujante, M.; Hoi, H.; Pavlova, A. Food availability and immune capacity in serin (Serinus serinus) nestlings. Behav. Ecol. Sociobiol. 2001, 49, 333–339. [Google Scholar] [CrossRef]
- Ardia, D.R. Cross-fostering reveals an effect of spleen size and nest temperatures on immune responses in nestling European starlings. Oecologia 2005, 145, 327–334. [Google Scholar] [CrossRef]
- Garvin, J.C.; Abroe, B.; Pedersen, M.C.; Dunn, P.O.; Whittingham, L.A. Immune response of nestling warblers varies with extra-pair paternity and temperature. Mol. Ecol. 2006, 15, 3833–3840. [Google Scholar] [CrossRef]
- Merino, S.; Møller, A.P. Host-parasite interactions and climate change. In Effects of Climate Change on Birds; Møller, A.P., Fiedler, W., Berthold, P., Eds.; Oxford University Press: Oxford, UK, 2010; pp. 213–226. [Google Scholar]
- Møller, A.P.; Merino, S.; Soler, J.J.; Antonov, A.; Badás, E.P.; Calero-Torralbo, M.A.; de Lope, F.; Eeva, T.; Figuerola, J.; Flensted-Jensen, E.; et al. Assessing the effects of climate on host-parasite interactions: A comparative study of European birds and their parasites. PLoS ONE 2013, 8, e82886. [Google Scholar] [CrossRef]
- Pounds, J.A.; Bustamante, M.R.; Coloma, L.A.; Consuegra, J.A.; Fogden, M.P.L.; Foster, P.N.; La Marca, E.; Masters, K.L.; Merino-Viteri, A.; Puschendorf, R.; et al. Widespread amphibian extinctions from epidemic disease driven by global warming. Nature 2006, 439, 161–167. [Google Scholar] [CrossRef]
- Badyaev, A.V. Altitudinal variation in sexual dimorphism: A new pattern and alternative hypotheses. Behav. Ecol. 1997, 8, 675–690. [Google Scholar] [CrossRef]
- Baur, A.; Baur, B. Interpopulation variation in the prevalence and intensity of parasitic mite infection in the land snail Arianta arbustorum. Invertebr. Biol. 2005, 124, 194–201. [Google Scholar] [CrossRef]
- Meléndez, L.; Laiolo, P.; Mironov, S.; García, M.; Magaña, O.; Jovani, R. Climate-driven variation in the intensity of a host-symbiont animal interaction along a broad elevation gradient. PLoS ONE 2014, 9, e101942. [Google Scholar] [CrossRef]
- Álvarez-Ruiz, L.; Megía-Palma, R.; Reguera, S.; Ruiz, S.; Zamora-Camacho, F.J.; Figuerola, J.; Moreno-Rueda, G. Opposed elevational variation in prevalence and intensity of endoparasites and their vectors in a lizard. Curr. Zool. 2018, 64, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Carbayo, J.; Martín, J.; Civantos, E. Habitat type influences parasite load in Algerian Psammodromus algirus. Can. J. Zool. 2019, 97, 172–180. [Google Scholar] [CrossRef]
- Llanos-Garrido, A.; Díaz, J.A.; Pérez-Rodríguez, A.; Arriero, E. Variation in male ornaments in two lizard populations with contrasting parasite loads. J. Zool. 2017, 303, 218–225. [Google Scholar] [CrossRef]
- Seddon, R.J.; Hews, D.K. Correlates of melanization in multiple high- and low-elevation populations of the lizard, Sceloporus occidentalis: Behavior, hormones, and parasites. J. Exp. Zool. Part A 2017, 327, 481–492. [Google Scholar] [CrossRef]
- Comas, M. Body condition, sex and elevation in relation to mite parasitism in a high mountain gecko. J. Zool. 2020, 310, 298–305. [Google Scholar] [CrossRef]
- Wiedenfeld, D.A.; Jiménez U., G.A.; Fessl, B.; Kleindorfer, S.; Valarezo, J.C. Distribution of the introduced parasitic fly Philornis downsi (Diptera, Muscidae) in the Galapagos Islands. Pac. Conserv. Biol. 2007, 13, 14. [Google Scholar] [CrossRef]
- O’Connor, J.A.; Dudaniec, R.Y.; Kleindorfer, S. Parasite infestation and predation in Darwin’s small ground finch: Contrasting two elevational habitats between islands. J. Trop. Ecol. 2010, 26, 285–292. [Google Scholar] [CrossRef]
- Young, B.E. Effects of the parasitic botfly Philornis carinatus on nestling house wrens, Troglodytes aedon, in Costa Rica. Oecologia 1993, 93, 256–262. [Google Scholar] [CrossRef]
- Hurtrez-Boussès, S.; Blondel, J.; Perret, P.; Renaud, F. Relationship between intensity of blowfly infestation and reproductive success in a Corsican population of Blue Tits. J. Avian Biol. 1997, 28, 267–270. [Google Scholar] [CrossRef]
- Hurtrez-Boussès, S.; Perret, P.; Renaud, F.; Blondel, J. High blowfly parasitic loads affect breeding success in a Mediterranean population of blue tits. Oecologia 1997, 112, 514–517. [Google Scholar] [CrossRef] [PubMed]
- Merino, S.; Potti, J. Growth, nutrition, and blow fly parasitism in nestling Pied Flycatchers. Can. J. Zool. 1998, 76, 936–941. [Google Scholar] [CrossRef]
- Bańbura, J.; Perret, P.; Blondel, J.; Thomas, D.W.; Cartan-Son, M.; Lambrechts, M.M. Effects of Protocalliphora parasites on nestling food composition in Corsican Blue Tits Parus caeruleus: Consequences for nestling performance. Acta Ornithol. 2004, 39, 93–103. [Google Scholar] [CrossRef]
- Puchala, P. Detrimental effects of larval blow flies (Protocalliphora azurea) on nestlings and breeding success of Tree Sparrows (Passer montanus). Can. J. Zool. 2004, 82, 1285–1290. [Google Scholar] [CrossRef]
- Simon, A.; Thomas, D.; Blondel, J.; Perret, P.; Lambrechts, M.M. Physiological ecology of Mediterranean blue tits (Parus caeruleus L.): Effects of ectoparasites (Protocalliphora spp.) and food abundance on metabolic capacity of nestlings. Physiol. Biochem. Zool. 2004, 77, 492–501. [Google Scholar] [CrossRef]
- Arriero, E.; Moreno, J.; Merino, S.; Martínez, J. Habitat effects on physiological stress response in nestling blue tits are mediated through parasitism. Physiol. Biochem. Zool. 2008, 81, 195–203. [Google Scholar] [CrossRef]
- Merino, S.; Potti, J. Mites and blowflies decrease growth and survival in nestling pied flycatchers. Oikos 1995, 73, 95–103. [Google Scholar] [CrossRef]
- Bouslama, Z.; Chabi, Y.; Lambrechts, M. Chicks resist high parasite intensities in an Algerian population of blue tits. Écoscience 2001, 8, 320–324. [Google Scholar] [CrossRef]
- Potti, J. Blowfly infestation at the nestling stage affects egg size in the Pied Flycatcher Ficedula hypoleuca. Acta Ornithol. 2008, 43, 76–82. [Google Scholar] [CrossRef]
- Eeva, T.; Andersson, T.; Berglund, Å.M.M.; Brommer, J.E.; Hyvönen, R.; Klemola, T.; Laaksonen, T.; Loukola, O.; Morosinotto, C.; Rainio, K.; et al. Species and abundance of ectoparasitic flies (Diptera) in pied flycatcher nests in Fennoscandia. Parasites Vectors 2015, 8, 648. [Google Scholar] [CrossRef]
- Garrido-Bautista, J.; Moreno-Rueda, G.; Baz, A.; Canal, D.; Camacho, C.; Cifrián, B.; Nieves-Aldrey, J.L.; Carles-Tolrá, M.; Potti, J. Variation in parasitoidism of Protocalliphora azurea (Diptera: Calliphoridae) by Nasonia vitripennis (Hymenoptera: Pteromalidae) in Spain. Parasitol. Res. 2020, 119, 559–566. [Google Scholar] [CrossRef] [PubMed]
- Gold, C.S.; Dahlsten, D.L. Prevalence, habitat selection, and biology of Protocalliphora (Diptera: Calliphoridae) found in nests of mountain and chestnut-backed chickadees in California. Hilgardia 1989, 57, 1–19. [Google Scholar] [CrossRef]
- Hori, K.; Iwasa, M.; Ogawa, R. Biology of two species of the Protocalliphora (Diptera: Calliphoridae) in Tokachi, Hokkaido, Japan: Feeding behaviour of larvae, larval and pupal duration, voltinism and host specificity. Appl. Entomol. Zool. 1990, 25, 475–482. [Google Scholar] [CrossRef]
- Polo, V. Carbonero Garrapinos—Periparus ater (Linnaeus, 1758). In Enciclopedia Virtual de los Vertebrados Españoles; Salvador, A., Morales, M.B., Eds.; Museo Nacional de Ciencias Naturales: Madrid, Spain, 2016; Available online: http://www.vertebradosibericos.org (accessed on 13 May 2021).
- Gosler, A. The Great Tit; Hamlyn: London, UK, 1993. [Google Scholar]
- Atiénzar, F.; Álvarez, E.; Barba, E. Carbonero común—Parus major (Linnaeus, 1758). In Enciclopedia Virtual de los Vertebrados Españoles; Salvador, A., Morales, M.B., Eds.; Museo Nacional de Ciencias Naturales: Madrid, Spain, 2016; Available online: http://www.vertebradosibericos.org (accessed on 21 June 2021).
- Baucells Colomer, J.; Camprodon i Subirachs, J.; Cerdeira i Ribot, J.; Vila Perdiguero, P. Guía de las Cajas Nido y Comederos Para Aves y Otros Vertebrados; Lynx Edicions: Barcelona, Spain, 2003. [Google Scholar]
- Moreno-Rueda, G. Selección de cajas-nido por aves insectívoras en Sierra Nevada. Zool. Baet. 2003, 13, 131–138. [Google Scholar]
- Rózsa, L.; Reiczigel, J.; Majoros, G. Quantifying parasites in samples of hosts. J. Parasitol. 2000, 86, 228–232. [Google Scholar] [CrossRef]
- Zuur, A.F.; Ieno, E.N.; Elphick, C.S. A protocol for data exploration to avoid common statistical problems. Methods Ecol. Evol. 2010, 1, 3–14. [Google Scholar] [CrossRef]
- Johnson, L.S.; Eastman, M.D.; Kermott, L.H. Effect of ectoparasitism by larvae of the blow fly Protocalliphora parorum (Diptera: Calliphoridae) on nestling House Wrens, Troglodytes aedon. Can. J. Zool. 1991, 69, 1441–1446. [Google Scholar] [CrossRef]
- Musgrave, K.; Bartlow, A.W.; Fair, J.M. Long-term variation in environmental conditions influences host-parasite fitness. Ecol. Evol. 2019, 9, 7688–7703. [Google Scholar] [CrossRef]
- Eeva, T.; Lehikoinen, E.; Nurmi, J. Effects of ectoparasites on breeding success of great tits (Parus major) and pied flycatchers (Ficedula hypoleuca) in an air pollution gradient. Can. J. Zool. 1994, 72, 624–635. [Google Scholar] [CrossRef]
- Eeva, T.; Klemola, T. Variation in prevalence and intensity of two avian ectoparasites in a polluted area. Parasitology 2013, 140, 1384–1393. [Google Scholar] [CrossRef]
- Lafferty, K.D. Environmental parasitology: What can parasites tell us about human impacts on the environment? Parasitol. Today 1997, 13, 251–255. [Google Scholar] [CrossRef]
- Butler, M.W.; Garvin, J.C.; Wheelwright, N.T.; Freeman-Gallant, C.R. Ambient temperature, but not paternity, is associated with immune response in savannah sparrows (Passerculus sandwichensis). Auk 2009, 126, 536–542. [Google Scholar] [CrossRef]
- Arneberg, P.; Skorping, A.; Grenfell, B.; Read, A.F. Host densities as determinants of abundance in parasite communities. Proc. R. Soc. Lond. Ser. B 1998, 265, 1283–1289. [Google Scholar] [CrossRef]
- Bennett, G.F.; Whitworth, T.L. Host, nest, and ecological relationship of species of Protocalliphora (Diptera: Calliphoridae). Can. J. Zool. 1992, 70, 51–61. [Google Scholar] [CrossRef]
- Cantarero, A.; López-Arrabé, J.; Rodríguez-García, V.; González-Braojos, S.; Ruiz-De-Castañeda, R.; Redondo, A.J.; Moreno, J. Factors affecting the presence and abundance of generalist ectoparasites in nests of three sympatric hole-nesting bird species. Acta Ornithol. 2013, 48, 39–54. [Google Scholar] [CrossRef]
- Moreno, J.; Merino, S.; Lobato, E.; Ruiz-De-Castañeda, R.; Martínez-de la Puente, J.; del Cerro, S.; Rivero-de Aguilar, J. Nest-dwelling ectoparasites of two sympatric hole-nesting passerines in relation to nest composition: An experimental study. Écoscience 2009, 16, 418–427. [Google Scholar] [CrossRef]
- Remeš, V.; Krist, M. Nest design and the abundance of parasitic Protocalliphora blow flies in two hole-nesting passerines. Écoscience 2005, 12, 549–553. [Google Scholar] [CrossRef]


| DF | χ2 | p | |
|---|---|---|---|
| Locality | 2 | 11.53 | 0.003 |
| Species | 1 | 0.465 | 0.50 |
| Locality*Species | 2 | 3.69 | 0.16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreno-Rueda, G. Elevational Patterns of Blowfly Parasitism in Two Hole Nesting Avian Species. Diversity 2021, 13, 591. https://doi.org/10.3390/d13110591
Moreno-Rueda G. Elevational Patterns of Blowfly Parasitism in Two Hole Nesting Avian Species. Diversity. 2021; 13(11):591. https://doi.org/10.3390/d13110591
Chicago/Turabian StyleMoreno-Rueda, Gregorio. 2021. "Elevational Patterns of Blowfly Parasitism in Two Hole Nesting Avian Species" Diversity 13, no. 11: 591. https://doi.org/10.3390/d13110591
APA StyleMoreno-Rueda, G. (2021). Elevational Patterns of Blowfly Parasitism in Two Hole Nesting Avian Species. Diversity, 13(11), 591. https://doi.org/10.3390/d13110591