Abstract
Aging profoundly reshapes the immune system, leading to increased susceptibility to infection, impaired vaccine responses, chronic inflammation, and age-associated inflammatory diseases. While immune aging has traditionally been attributed to defects in immune cell development, signaling, and metabolism, emerging evidence highlights dysregulation of programmed cell death as a central and unifying mechanism. Apoptosis, necroptosis, pyroptosis, and ferroptosis are increasingly recognized not only as terminal cellular events but also as active regulators of immune homeostasis and inflammatory signaling. In aged immune cells, coordination among these death pathways is disrupted, weakening apoptotic resolution and favoring inflammatory forms of cell death that amplify tissue damage and sustain inflammaging. In this review, we summarize current evidence on how aging remodels programmed cell death pathways in the immune system, discuss the molecular mechanisms underlying this network-level shift, and consider potential strategies for restoring immune function by modulating cell death decisions.
1. Introduction
Immune aging, or immunosenescence, is characterized by a progressive decline in protective immunity, including impaired pathogen clearance, weakened vaccine responsiveness, and compromised immune memory [1,2,3,4,5]. This functional deterioration is frequently accompanied by inflammaging, a chronic, low-grade inflammatory state implicated in a wide range of age-related diseases such as cardiovascular disorders, neurodegeneration, metabolic syndrome, and cancer [6,7,8,9]. Although these two phenomena are tightly linked, immune aging and inflammaging are conceptually distinct. Inflammaging describes a systemic inflammatory condition, whereas immune aging refers to intrinsic, cell-autonomous and population-level changes within the immune system that precede, shape, and sustain this inflammatory milieu [10].
Most studies on immune aging have focused on altered immune cell composition, stem cell exhaustion, replicative senescence, and defective signaling pathways [11]. In contrast, inflammaging has largely been attributed to persistent innate immune activation, accumulation of senescent cells, and chronic exposure to endogenous danger signals [12,13,14,15,16]. However, the mechanistic bridge connecting immune-intrinsic aging processes to sustained inflammation remains incompletely understood.
Programmed cell death provides a critical conceptual and mechanistic link between immune aging and inflammaging. Under homeostatic conditions, apoptosis enables immunologically silent removal of damaged or excess immune cells, thereby limiting inflammation [17,18]. Aging, however, alters cellular stress responses, mitochondrial integrity, redox balance, and proteostasis, reshaping the decision-making landscape of cell death [19,20,21,22]. Rather than simply increasing cell loss, aging fundamentally disrupts the coordinated regulation of programmed cell death, weakening apoptosis-mediated silent resolution and favoring inflammatory death pathways such as necroptosis, pyroptosis, and ferroptosis. This network-level rewiring transforms immune cell turnover into a persistent source of inflammation, positioning immune aging as a causal driver of inflammaging rather than a passive consequence [23,24,25,26].
In this review, we discuss immune aging through the framework of programmed cell death, highlighting how age-driven changes in death pathway selection within immune cells provide a mechanistic basis for inflammaging.
2. Hallmarks of Immune Aging Relevant to Cell Death
Several hallmarks of immune aging directly predispose immune cells to altered death decisions. These changes operate across multiple levels, ranging from stem cell dysfunction to cellular metabolic stress and tissue-level inflammatory cues, collectively reshaping the threshold and modality of programmed cell death.
At the level of hematopoiesis, aging is associated with profound alterations in hematopoietic stem cell (HSC) function. Aged HSCs exhibit clonal skewing, reduced self-renewal capacity, and a bias toward myeloid differentiation, resulting in immune cell populations that are functionally compromised and prone to stress [27,28,29]. Importantly, these progenitor-derived immune cells inherit mitochondrial dysfunction, impaired stress responses, and epigenetic alterations that influence survival and death signaling [27,28,29,30,31,32,33,34]. Such intrinsic defects prime aged immune cells to respond aberrantly to inflammatory or metabolic stress, increasing their susceptibility to dysregulated cell death [35].
At the cellular level, mitochondrial dysfunction represents a central hallmark linking immune aging to altered death pathway selection. Aged immune cells accumulate mitochondrial DNA damage, exhibit reduced oxidative phosphorylation efficiency and generate excessive reactive oxygen species (ROS) [36,37,38]. These changes not only sensitize cells to death-inducing stimuli but also impair the execution of canonical apoptosis by disrupting mitochondrial outer membrane permeabilization and caspase activation [36,39]. Elevated ROS further promotes lipid peroxidation, iron dysregulation, and inflammasome activation, thereby favoring inflammatory forms of cell death such as ferroptosis and pyroptosis [40,41,42,43,44].
Impaired proteostasis and defective autophagy further exacerbate the death pathway imbalance in aged immune cells. Autophagy plays a critical role in maintaining mitochondrial quality control and limiting inflammasome activation [45,46]. With aging, reduced autophagic flux allows damaged mitochondria and protein aggregates to accumulate, amplifying cytosolic danger signals [47]. This environment lowers the threshold for inflammasome activation and necroptotic signaling, while simultaneously limiting the cell’s ability to resolve stress through adaptive survival pathways [48].
The aging immune system is also affected by chronic exposure to inflammatory cytokines, particularly TNF, IL-1β, and type I interferons. Persistent cytokine signaling enforces a state of sublethal stress that continuously engages death-related signaling pathways. In young immune cells, such signals are typically resolved through apoptosis followed by efficient clearance. In aged cells, however, chronic cytokine exposure intersects with reduced caspase-8 activity, altered NF-κB signaling, and metabolic insufficiency, conditions that favor diversion toward necroptosis or pyroptosis rather than apoptotic resolution [12,49].
Beyond cell-intrinsic defects, aging profoundly impairs efferocytosis—the process by which dying cells are recognized and cleared by phagocytes [50]. Aged macrophages and stromal cells exhibit reduced capacity to engulf apoptotic cells, leading to the accumulation of uncleared corpses within tissues [51,52]. These dying cells frequently undergo secondary necrosis, releasing damage-associated molecular patterns (DAMPs) that further activate innate immune receptors and reinforce inflammatory cell death pathways in neighboring cells [14,53]. Thus, inadequate clearance of apoptotic cells shifts apoptosis from a silent process to a chronic inflammatory stimulus [50,54].
Together, these hallmarks establish an immune environment in which cell death is not only more frequent but also qualitatively altered. With aging, immune cells transition from tightly regulated, immunologically silent apoptosis toward inflammatory and metabolically driven death that triggers systemic inflammation. This rewiring of death pathway selection provides a mechanistic foundation for understanding how immune aging actively promotes inflammaging rather than merely accompanying it. Consistent with these functional alterations, aging is accompanied by a progressive shift in the expression and activity of key cell death regulators, including increased signaling capacity of necroptotic and inflammasome pathways and diminished antioxidant defenses that normally protect against lipid peroxidation (Figure 1) [55,56,57,58,59].
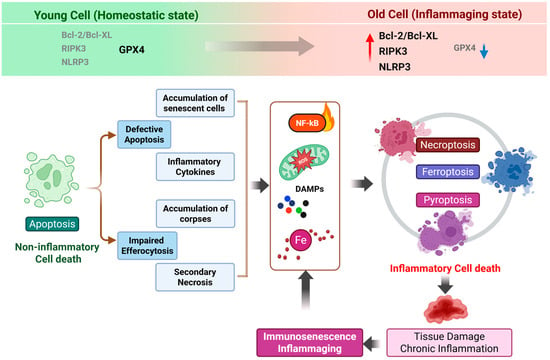
Figure 1.
From apoptosis to inflammatory cell death: a vicious cycle underlying immune aging. This schematic illustrates an age-associated shift in cell death programs from homeostatic apoptosis toward inflammatory forms of regulated cell death, including necroptosis, pyroptosis, and ferroptosis. Comparative expression patterns of key regulatory molecules in cells from young versus aged conditions are indicated at the top, highlighting increased Bcl-2/Bcl-XL, RIPK3 and NLRP3 expression (the red upward arrow) and reduced GPX4 levels in aged cells (the blue downward arrow). During aging, reduced efficiency of apoptosis and a decline in efferocytosis lead to the accumulation of dysfunctional cells and cellular debris within tissues. These uncleared apoptotic cells progressively undergo secondary necrosis, resulting in the release of damage-associated molecular patterns (DAMPs) and pro-inflammatory cytokines. The persistent presence of these signals sustains NF-κB activation and reinforces a chronic inflammatory milieu. In parallel, mitochondrial dysfunction–derived reactive oxygen species (ROS) and iron dysregulation further exacerbate cellular stress, shifting cell fate decisions away from immunologically silent apoptosis toward inflammatory forms of cell death, such as necroptosis, pyroptosis and ferroptosis. The ensuing release of additional inflammatory mediators amplifies tissue damage, accelerates immune cell dysfunction and senescence, and establishes a self-perpetuating vicious cycle that underlies immune aging and inflammaging. Created in BioRender. Kang, T. (2026). https://BioRender.com/ua8m688 (accessed on 2 February 2026).
3. Apoptosis in Immune Aging
Apoptosis is the dominant homeostatic cell death mechanism in the immune system, ensuring the safe elimination of excess, damaged, or autoreactive cells without affecting tissue integrity and immunological quiescence [60,61,62]. Although intrinsic and extrinsic pathways are tightly regulated, apoptosis maintains immune cell homeostasis, shapes lymphocyte repertoires, and facilitates the resolution of immune responses without triggering inflammation. However, aging profoundly alters the regulation, execution, and resolution of apoptosis, fundamentally changing its functional outcome within the immune system [50,63,64].
Importantly, immune aging does not simply increase or decrease apoptotic frequency in a uniform manner. Rather, aging introduces context-dependent dysregulation, in which certain immune cell populations become resistant to apoptosis, while others show heightened sensitivity. This imbalance disrupts immune composition and compromises immune competence [3,6,65]. In adaptive immunity, repeated antigen exposure, telomere attrition, epigenetic drift, and metabolic insufficiency collectively drive T cell exhaustion. Exhausted and terminally differentiated memory T cells frequently acquire resistance to apoptosis, allowing them to persist despite functional impairment [66,67,68,69,70]. The accumulation of such dysfunctional memory T cells contributes to reduced immune plasticity and diminished responsiveness to new antigens.
In contrast, naïve T cells in aged individuals often display increased susceptibility to apoptosis [71,72]. Heightened sensitivity to cytokine withdrawal, oxidative stress, and mitochondrial damage leads to accelerated loss of naïve T cells, resulting in contraction of the T cell receptor repertoire. This selective depletion compromises adaptive immune diversity and represents a defining characteristic of immunosenescence. Thus, aging reshapes apoptosis in a population-specific manner, promoting the survival of dysfunctional immune cells while eliminating those required for robust immune renewal [73,74,75,76].
In innate immune cells, aging-related changes are characterized by insufficient apoptotic turnover instead of simple resistance to pro-apoptotic signaling. Many studies indicate that aged innate immune cells exhibit impaired activation of some caspases, altered mitochondrial outer membrane permeabilization, and dysregulated death receptor signaling. These defects do not necessarily prevent cell death but instead delay or incompletely execute the apoptotic program. Such incomplete or defective apoptosis leads to a permissive state in which cells remain metabolically active despite being marked for death signals [36,77,78,79,80].
This incomplete apoptotic state carries significant functional implications [17,81,82,83,84]. Under conditions of chronic inflammatory signaling—such as persistent exposure to TNF, IL-1β, or type I interferons—aged immune cells with compromised apoptotic machinery tend to redirect death signaling toward alternative, inflammatory pathways [15,85]. In particular, reduced caspase-8 activity removes a critical checkpoint that normally suppresses necroptosis [86,87,88]. As a result, aging transforms apoptosis from a terminal, anti-inflammatory endpoint into an unstable intermediate that facilitates engagement of necrotic death programs.
In addition to execution defects in apoptosis, aging also profoundly impairs the resolution phase of apoptosis, namely the efficient clearance of apoptotic cells by phagocytes [51]. Efferocytosis is essential for maintaining immune tolerance and preventing secondary inflammation [50]. However, aged macrophages exhibit reduced phagocytic capacity, altered expression of “eat-me” receptors, and diminished responsiveness to apoptotic signals, consequently leading to the prolonged persistence of apoptotic cells in tissues [51].
Persisting apoptotic cells frequently undergo secondary necrosis, leading to plasma membrane rupture and release of damage-associated molecular patterns (DAMPs), including nuclear DNA, HMGB1, and oxidized lipids [54,89,90]. This process converts an otherwise immunologically silent form of cell death into a potent inflammatory stimulus [91]. The accumulation of secondary necrotic cells reinforces innate immune activation, perpetuates cytokine production, and further lowers the threshold for inflammatory cell death in neighboring immune cells [50,54,92,93]. Therefore, defective apoptotic clearance acts as a self-reinforcing driver of chronic inflammation.
Collectively, these alterations suggest that apoptosis in the aging immune system is not simply a matter of being diminished or enhanced; rather, it is fundamentally reconfigured and functionally compromised. Aging weakens apoptosis as a protective barrier that normally constrains inflammation and prevents inappropriate activation of necrotic death pathways. Instead, apoptotic dysregulation creates the cellular environment to favor the inflammatory cell death pathway (Figure 2) [25,26,50,94,95].
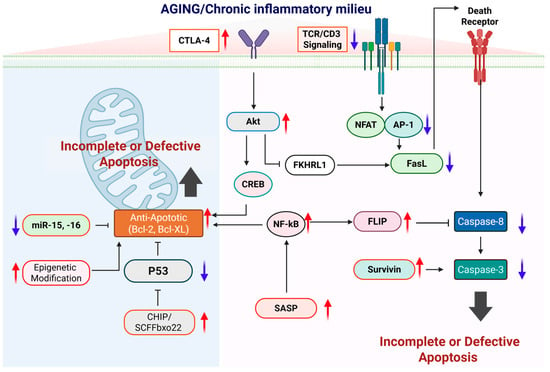
Figure 2.
Molecular mechanisms underlying impaired apoptotic control during aging. This schematic depicts how aging and a chronic inflammatory milieu progressively disrupt apoptotic regulation at multiple molecular checkpoints, leading to incomplete or defective apoptosis and the accumulation of apoptosis-resistant cells. In aged immune cells, increased expression of inhibitory receptors such as CTLA-4 and attenuated TCR/CD3 signaling weaken pro-apoptotic transcriptional programs mediated by NFAT and AP-1, resulting in reduced FasL expression and impaired activation of the extrinsic apoptotic pathway. At the same time, pro-survival signaling pathways, including Akt–CREB and persistent NF-κB activation, are enhanced and drive the upregulation of anti-apoptotic molecules such as Bcl-2, Bcl-XL, FLIP, and survivin, while suppressing caspase-8 and caspase-3 activation downstream of death receptor signaling. Within the intrinsic pathway, aging-associated mitochondrial alterations, epigenetic modifications, reduced expression of pro-apoptotic microRNAs (miR-15 and miR-16), and enhanced CHIP/SCFFbx22-mediated p53 degradation or functional suppression further reinforce resistance to apoptosis. In parallel, chronic inflammatory cues and senescence-associated secretory phenotype (SASP) signaling sustain NF-κB–dependent survival programs, thereby stabilizing an anti-apoptotic state. Red upward arrows indicate pathways or factors that are increased with aging, whereas blue downward arrows denote those that are diminished. Created in BioRender. Kang, T. (2026) https://BioRender.com/uk3uq46 (accessed on 2 February 2026).
Thus, apoptosis occupies a pivotal role in immune aging—not only as a victim of age-associated cellular stress, but also as a gatekeeper whose failure enables the emergence of necroptosis, pyroptosis, and other inflammatory death pathways. Understanding how apoptotic integrity is lost with age is therefore essential for deciphering the broader reprogramming of cell death pathways that underlie immune dysfunction and inflammaging [6,95,96,97].
4. Necroptosis as an Inflammatory Amplifier in Aging
Necroptosis is a regulated form of necrotic cell death mediated by the RIPK1–RIPK3–MLKL signaling axis [88,98,99]. Under physiological conditions, necroptosis functions as a protective host defense mechanism, eliminating infected cells when pathogens actively suppress apoptotic machinery [100,101,102]. However, accumulating evidence suggests that aging creates a cellular and tissue environment in which necroptosis is aberrantly activated, even in the absence of infection [82,103,104]. Rather than serving as a backup defense pathway, necroptosis increasingly operates as a maladaptive death program that amplifies inflammation in aged tissues [58,105] (Figure 3).
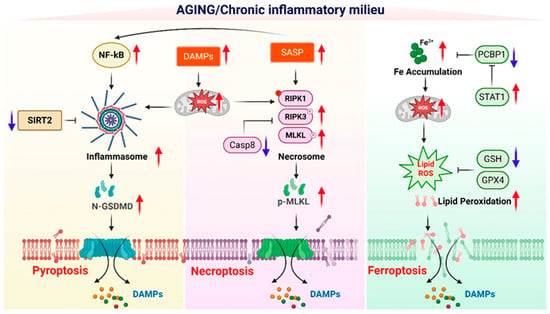
Figure 3.
Molecular reprogramming of inflammatory cell death pathways in the aging environment. This schematic illustrates how the aging-associated chronic inflammatory milieu selectively reprograms molecular pathways to promote distinct forms of inflammation-regulated cell death, including pyroptosis, necroptosis, and ferroptosis. During aging, sustained activation of NF-κB signaling, increased accumulation of damage-associated molecular patterns (DAMPs), and enhanced senescence-associated secretory phenotype (SASP) collectively amplify mitochondrial dysfunction and reactive oxygen species (ROS) production, creating a permissive environment for inflammatory cell death. In the pyroptotic pathway, reduced SIRT2 activity and heightened inflammasome activation facilitate caspase-dependent cleavage of gasdermin D (GSDMD), leading to membrane pore formation and the release of DAMPs. In parallel, aging-associated suppression of caspase-8 activity, together with increased RIPK1, RIPK3, and MLKL phosphorylation, favors necrosome assembly and execution of necroptosis via MLKL–mediated membrane disruption. Ferroptosis is promoted by age-related iron accumulation, enhanced Fe2+-driven mitochondrial ROS generation, impaired iron chaperoning by PCBP1, and altered STAT1 signaling, which collectively exacerbate lipid peroxidation. Concurrent depletion of antioxidant defenses, including reduced glutathione (GSH) levels and diminished GPX4 activity, further sensitizes cells to ferroptotic death. Red upward arrows indicate molecular pathways or factors that are increased in the aging or chronic inflammatory environment, whereas blue downward arrows denote pathways that are diminished with aging. The release of DAMPs from each inflammatory cell death modality reinforces chronic inflammation, thereby sustaining a feed-forward loop that exacerbates tissue dysfunction and immune aging. Created in BioRender. Kang, T. (2026) https://BioRender.com/3i5cyox (accessed on 2 February 2026).
Multiple age-associated factors converge to lower the threshold for necroptotic activation in immune cells [103]. Reduced caspase-8 expression or activity, a hallmark of aging immune cells, removes a critical inhibitory checkpoint that normally suppresses RIPK3 activation [86,106]. At the same time, chronic exposure to inflammatory cytokines—particularly TNF—provides sustained upstream signaling via death receptors [107]. Oxidative stress and mitochondrial dysfunction lower the threshold for RIPK1 activation by undermining the robustness of survival signaling [108,109]. Together, these conditions favor the assembly of the necrosome complex and execution of necroptosis in aged immune cells.
Macrophages and tissue-resident immune cells appear particularly susceptible to necroptotic reprogramming during aging [104]. These cells are chronically exposed to inflammatory cues and metabolic stress within aging tissues and often exhibit impaired apoptotic resolution, as discussed in the previous section. In this context, necroptosis becomes a dominant alternative fate when apoptotic checkpoints fail. Execution of necroptosis leads to MLKL-mediated plasma membrane permeabilization, resulting in rapid cell lysis and uncontrolled release of intracellular contents [110,111,112].
Necroptosis is highly immunogenic, characterized by the extensive release of damage-associated molecular patterns (DAMPs). Key inflammatory mediators such as HMGB1, mitochondrial DNA, ATP, and oxidized lipids are released, driving a pro-inflammatory phenotype [113,114]. These molecules engage pattern recognition receptors such as TLRs and cytosolic DNA sensors in neighboring immune and stromal cells, triggering further production of pro-inflammatory cytokines [115,116,117]. Thus, necroptosis acts as both a result and a driver of inflammatory stress, creating a vicious cycle.
Evidence from aged animal models supports a direct link between necroptosis and chronic inflammation. Increased expression and activation of RIPK3 and MLKL have been observed in aged tissues prone to inflammatory degeneration, including the intestine, liver, brain, and kidney [82,104]. Genetic or pharmacological inhibition of necroptotic signaling in these models often attenuates inflammatory pathology and tissue damage, suggesting that necroptosis is not merely a marker of immune aging but a functional driver of age-associated inflammation [58,104].
Importantly, necroptosis exhibits extensive crosstalk with other programmed cell death pathways in the context of aging. RIPK3 signaling serves as a bridge, triggering inflammasome activation and IL-1β release, which integrates necroptotic and pyroptotic responses [118,119,120]. In aged immune cells, such crosstalk further amplifies inflammatory output and reinforces inflammaging. Thus, necroptosis is not an isolated process but a central hub that connects various inflammatory death programs.
Collectively, these findings position necroptosis as a potent inflammatory amplifier in the aging immune system. Through the induction of membrane rupture and subsequent DAMP release, necroptosis transforms immune aging from passive attrition to active inflammatory progression. Understanding how necroptotic signaling is engaged and regulated in aged immune cells is, therefore, essential for identifying therapeutic strategies aimed at breaking the cycle of inflammaging.
5. Pyroptosis and Inflammasome Activation in Immune Aging
Pyroptosis is a highly inflammatory form of programmed cell death driven by inflammasome activation and gasdermin-mediated membrane pore formation [121,122,123]. Unlike apoptosis, pyroptosis is intrinsically pro-inflammatory, coupling cell death with the rapid release of cytokines and intracellular danger signals. Accumulating evidence suggests that aging markedly lowers the threshold for inflammasome activation, rendering aged tissues highly susceptible to pyroptosis-driven immune decline (Figure 3) [124,125,126].
Aging is characterized by an elevated basal priming of the NLRP3 inflammasome [124,127]. At the transcriptional level, aged immune cells often exhibit sustained NF-κB activation due to chronic exposure to inflammatory cytokines and endogenous danger signals [128,129]. This persistent priming state ensures elevated expression of inflammasome components such as NLRP3 and pro–IL-1β, effectively sensitizing aged cells to secondary activation signals [126,130]. Consequently, stimuli that would be insufficient to trigger inflammasome activation in young immune cells can readily induce pyroptotic responses in aged cells [127].
Various hallmarks of cellular aging act synergistically to facilitate inflammasome activation. In particular, mitochondrial dysfunction triggers a pro-inflammatory cascade by releasing reactive oxygen species (ROS) and mitochondrial DNA into the cytoplasm [131,132]. In parallel, age-associated lysosomal instability facilitates leakage of cathepsins, while dysregulated ion homeostasis—particularly potassium efflux—further lowers the activation threshold [133,134,135]. Together, these intracellular stress signals create a permissive environment for the formation of inflammasome assembly and caspase-1 activation.
Macrophages and microglia are particularly vulnerable to pyroptotic reprogramming during aging. As long-lived, tissue-resident immune cells, they gradually accumulate metabolic stress, damaged organelles, and persistent inflammatory cues over time [126,136,137]. In aged macrophages, enhanced inflammasome activation leads to excessive cleavage of gasdermin D, resulting in membrane pore formation and release of IL-1β and IL-18. Similarly, aged microglia display exaggerated inflammasome responses that contribute to neuroinflammation and neurodegenerative pathology [138,139]. Rather than promoting efficient pathogen clearance or tissue repair, repeated or sustained pyroptotic activation drives tissue dysfunction and reinforces chronic inflammation.
Inflammaging associated with senescence primes and sustains inflammasome activity, driving chronic IL-1β/IL-18 signaling. Through positive-feedback loops, this persistent signaling transforms pyroptosis from an acute, self-limiting defense mechanism into a chronic, feed-forward inflammatory cycle. Cytokines released from pyroptotic cells further prime neighboring immune cells, amplify local cytokine networks, and exacerbate tissue damage [124,140,141,142,143]. As a result, this persistent activation state obscures the line separating beneficial immunity from destructive inflammation, encapsulating the essence of inflammaging [126].
Emerging evidence also highlights extensive crosstalk between pyroptosis and other inflammatory cell death pathways in aged immune systems [144,145]. Inflammasome activation can promote necroptotic signaling through RIPK3-dependent mechanisms, while necroptosis-associated DAMP release further enhances inflammasome activation in surrounding cells [120,146,147]. In aged immune cells, where apoptotic checkpoints are weakened, this coordinated activation of pyroptosis and necroptosis amplifies inflammatory output far beyond that produced by either pathway alone [82,148]. Consequently, aging results in an integrated network of inflammatory cell death rather than the activation of discrete, independent pathways.
Collectively, these observations define inflammasome-dependent pyroptosis as a key effector mechanism that integrates immune aging with systemic inflammation. By linking the detection of cellular stress to explosive membrane rupture and cytokine release, pyroptosis transforms age-associated immune dysfunction into persistent inflammatory signaling. Understanding how inflammasome activation is dysregulated with age and how pyroptosis integrates with parallel death pathways is, therefore, essential for deciphering the molecular basis of immune aging and identifying strategies to mitigate inflammaging.
6. Ferroptosis and Metabolic Vulnerability of Aged Immune Cells
Ferroptosis is an iron-dependent, lipid peroxidation–driven form of regulated cell death that has recently emerged as a critical contributor to immune dysfunction. Unlike apoptosis or inflammasome-driven cell death, ferroptosis is tightly linked to cellular metabolism, redox balance, and iron homeostasis [59,149]. Aging profoundly alters these metabolic parameters, rendering aged cells increasingly vulnerable to ferroptotic stress (Figure 3) [150,151].
Aged immune cells frequently exhibit increased intracellular iron accumulation, impaired antioxidant capacity, and dysregulated lipid metabolism [152,153,154]. Systemic iron homeostasis becomes less tightly regulated with age, leading to iron deposition in multiple tissues. At the cellular level, reduced expression or activity of antioxidant systems—such as glutathione, GPX4, and NADPH-generating pathways—limits the ability of aged immune cells to detoxify lipid peroxides [59,155]. Concurrently, age-associated changes in lipid composition, including increased polyunsaturated fatty acid content in cellular membranes, further sensitize cells to lipid peroxidation. Together, these alterations create a metabolic landscape that strongly favors ferroptotic cell death [23,151,156].
Macrophages play a pivotal role in this process. As key regulators of iron recycling and storage, they are continuously exposed to iron flux and oxidative stress [157,158]. In aging tissues, macrophages accumulate excess iron and display impaired mitochondrial function and redox control, making them particularly susceptible to ferroptosis [157]. Ferroptotic death of macrophages not only compromises innate immune capacity but also disrupts local iron homeostasis, further amplifying oxidative stress within the tissue microenvironment [158,159,160].
Emerging evidence suggests that ferroptosis also affects adaptive immune cells [161]. T cells depend on strictly controlled redox signaling for activation, proliferation, and survival [162]. In aged T cells, compromised antioxidant defenses and mitochondrial dysfunction increase susceptibility to lipid peroxidation–induced damage [73,75,163]. Ferroptotic loss of T cells may therefore contribute to immune exhaustion, impaired effector function, and reduced immune surveillance in aging organisms [164,165].
Importantly, the impact of ferroptosis extends beyond depletion of immune cell populations, as the resulting oxidized phospholipids and lipid-derived mediators act as potent inflammatory signals [166,167]. These oxidized lipids can activate pattern recognition receptors (PRRs), modulate macrophage polarization, and amplify cytokine production in neighboring immune and stromal cells [168,169,170]. Thus, ferroptosis contributes to inflammaging not only by reducing immune cell viability but also by actively shaping a pro-inflammatory tissue milieu.
Although ferroptosis remains less extensively studied in the context of immune aging than apoptosis, necroptosis, or pyroptosis, accumulating evidence supports its relevance to age-associated inflammation and immune dysfunction. Notably, ferroptosis exhibits mechanistic crosstalk with other cell death pathways through shared upstream drivers, including mitochondrial dysfunction, ROS accumulation, and metabolic insufficiency [167,171,172]. These shared stressors suggest that ferroptosis is integrated into a broader network of inflammatory and metabolic cell death programs that are rewired during aging.
Collectively, ferroptosis shows how metabolic changes contribute to the aging of the immune system. By coupling iron dysregulation, oxidative stress, and lipid peroxidation to immune cell loss and inflammatory signaling, ferroptosis provides a mechanistic link between metabolic aging and chronic inflammation. Further investigation into ferroptotic regulation in immune cells will be essential for fully understanding immune aging and identifying metabolic interventions capable of restoring immune resilience in aged tissues.
7. Crosstalk Among Cell Death Pathways in Immune Aging
Programmed cell death pathways do not function as isolated, linear cascades; instead, they form a tightly interconnected regulatory network whose structure and functional balance are profoundly remodeled during aging. In young and healthy immune systems, apoptosis serves as the dominant and protective default, effectively resolving cellular stress and limiting inflammatory signaling [173]. Alternative death programs such as necroptosis, pyroptosis, and ferroptosis are tightly constrained and typically engaged only under specific pathological conditions. Aging disrupts this hierarchy, weakening apoptotic checkpoints and enabling coordinated activation of inflammatory and metabolic cell death pathways.
A master regulator in this network is caspase-8, which functions as a molecular gatekeeper between apoptosis and necroptosis [95,174]. Under homeostatic conditions, caspase-8 activity ensures apoptotic execution while actively suppressing RIPK3-dependent necroptotic signaling. In aged immune cells, however, reduced caspase-8 expression or activity—driven by chronic inflammatory stress, mitochondrial dysfunction, and altered redox balance—destabilizes this checkpoint [25,145]. Consequently, death receptor signaling that would normally lead to apoptosis is shunted toward necroptosis, particularly in macrophages and tissue-resident immune cells exposed to persistent TNF signaling.
Mitochondrial dysfunction serves as another critical hub linking multiple cell death modalities within the context of immune aging [39,175]. Mitochondria integrate metabolic status, redox signaling, and innate immune sensing, placing them at the intersection of apoptosis, pyroptosis, and ferroptosis. In aged immune cells, mitochondrial damage leads to impaired apoptotic signaling, excessive production of reactive oxygen species, and release of mitochondrial DNA into the cytosol [75,176,177]. These events simultaneously promote inflammasome activation and reinforce the sensitivity to oxidative damage. Thus, mitochondrial aging appears to shift cell death decisions toward inflammatory and oxidative outcomes by bridging disparate death programs.
Inflammasome signaling further reinforces this network-level crosstalk. Activation of inflammasomes, particularly NLRP3, not only drives pyroptotic cell death but also intersects with necroptotic signaling [178,179,180]. RIPK3 and MLKL have been shown to modulate inflammasome activation, while necroptosis-associated DAMP release provides potent secondary signals that amplify inflammasome priming in neighboring cells [146,147,181,182]. In aging tissues, where basal inflammasome activity is already heightened [125,126], this bidirectional interaction establishes self-sustaining inflammatory loops that propagate cell death and inflammation across immune populations.
Finally, the metabolic dimension of ferroptosis is integrated into this broader stress–death–inflammation axis. Oxidative stress derived from ferroptotic processes can exacerbate the mitochondrial dysfunction and inflammasome activation mentioned above, while inflammatory cytokines generated by pyroptosis and necroptosis further disrupt cellular redox balance [120,132,183]. In this way, ferroptosis does not operate independently but is integrated into a broader stress–death–inflammation axis characteristic of immune aging.
Viewing immune aging through this network-based framework reframes immunosenescence as a failure of cell death coordination rather than a simple change in cell quantity. Understanding how aging rewires the crosstalk among apoptosis, necroptosis, pyroptosis, and ferroptosis is therefore essential for developing strategies that restore immune balance in aged tissues. Importantly, accumulating evidence from neurodegenerative disorders such as Alzheimer’s disease demonstrates that these death pathways frequently coexist and dynamically switch, providing a disease-relevant framework for the network-based model of immune aging (Table 1).

Table 1.
Evidence for the coexistence of apoptosis, necroptosis, pyroptosis, and ferroptosis in Alzheimer’s disease and their switching mechanisms.
8. Therapeutic Implications
Targeting programmed cell death pathways offers novel and conceptually distinct opportunities to modulate immune aging. Unlike conventional anti-inflammatory approaches that primarily suppress downstream cytokine signaling, interventions aimed at cell death decisions act upstream, at the level of cellular fate determination. By restoring appropriate death pathway selection, such strategies have the potential to rebalance immune homeostasis while preserving essential host defense mechanisms.
Pharmacological inhibition of necroptotic signaling, particularly through targeting RIPK1, represents a promising approach to limit inflammatory cell death in aged tissues [58,195,196]. Preclinical studies in inflammatory and degenerative disease models have demonstrated that RIPK1 inhibition can attenuate tissue damage and chronic inflammation without broadly suppressing immune responsiveness [197,198]. In the context of immune aging, such interventions may prevent the inappropriate engagement of necroptosis that arises from apoptotic checkpoint failure [104,199].
Similarly, suppression of inflammasome activation offers therapeutic potential for mitigating age-associated inflammation [126,200,201,202]. Targeting key components of inflammasome signaling, such as NLRP3 or caspase-1, may reduce excessive pyroptosis and cytokine release in aged innate immune cells. Importantly, selective modulation of inflammasome activity—rather than complete inhibition—may be required to maintain protective immune responses while limiting chronic inflammatory signaling.
Modulation of ferroptosis introduces a complementary metabolic strategy for immune rejuvenation [59,151,203,204]. Enhancing antioxidant capacity, regulating iron homeostasis, or stabilizing lipid metabolism may reduce ferroptotic vulnerability in aged immune cells. Such approaches are particularly attractive in the aging setting, as they address upstream metabolic and redox imbalances that simultaneously influence multiple cell death pathways.
Beyond direct pharmacological interventions, micronutrient supplementation and metabolic modulation represent additional, potentially safer strategies for long-term application in aging populations. Nutrients that support mitochondrial integrity, redox balance, and cellular stress resilience may indirectly reprogram death pathway selection, restoring the dominance of non-inflammatory apoptotic resolution. These interventions may be especially valuable for preventing the gradual escalation of inflammaging rather than treating advanced inflammatory pathology.
Collectively, these therapeutic strategies highlight a shift from symptom-oriented anti-inflammatory treatments toward mechanism-based modulation of immune cell fate. Targeting the core controls of cell death can lower aging-related inflammation while keeping the immune competence. This offers a lasting solution for healthy aging.
Despite the compelling mechanistic rationale, translating cell death–based interventions into clinical practice remains challenging. Programmed cell death pathways are highly context- and cell type–dependent, and their long-term or systemic modulation may compromise host defense, tissue repair, or immune balance in aged individuals. In addition, the lack of reliable biomarkers to monitor death pathway engagement in vivo and the pronounced heterogeneity of immune aging complicate patient stratification and therapeutic optimization. Addressing these limitations will be critical for bridging mechanistic insights with safe and effective clinical applications.
9. Future Perspectives
Despite significant progress in understanding how programmed cell death pathways are rewired during immune aging, many fundamental questions remain unresolved.
A key challenge is determining how cell death decisions vary at the single-cell level within aged immune populations. Aging increases cellular heterogeneity, raising the possibility that distinct subpopulations within the same immune lineage may preferentially engage different death pathways in response to identical stimuli. Dissecting this heterogeneity will be essential for understanding why inflammatory cell death becomes dominant in aging tissues.
Tissue context represents another major knowledge gap. Immune cells reside in highly diverse microenvironments, each characterized by distinct metabolic conditions, stromal interactions, and inflammatory cues. How apoptosis, necroptosis, pyroptosis, and ferroptosis are differentially engaged across tissues such as barrier organs, lymphoid sites, and immune-privileged regions remains largely unexplored.
An especially intriguing and clinically relevant question is whether immune aging can be reversed or reprogrammed by resetting cell death thresholds. If immune dysfunction reflects dysregulated death pathway coordination rather than irreversible damage, restoring apoptotic control or limiting inflammatory cell death may partially rejuvenate immune function.
Addressing these issues will require systems-level approaches that integrate single-cell, spatial, and in vivo analyses to move beyond pathway-centric models of cell death in aging immunity.
10. Conclusions
Aging transforms immune cell death from a quiet process to a loud, inflammatory one, converting physiological cell turnover into a persistent source of inflammation and tissue damage. By integrating apoptosis, necroptosis, pyroptosis, and ferroptosis into a unified framework, this review proposes that immune aging reflects a breakdown in the coordination of the cell death program rather than a simple accumulation of cell damage. The convergence of mitochondrial dysfunction, redox imbalance, chronic inflammatory signaling, and impaired clearance mechanisms rewires death pathway selection toward inflammatory outcomes, thereby providing a mechanistic foundation for inflammaging.
This perspective also carries important therapeutic implications. Targeting upstream regulatory nodes that govern cell death decisions, rather than suppressing downstream inflammatory mediators alone, may offer a more effective strategy to restore immune homeostasis while preserving host defense. Interventions that stabilize apoptotic checkpoints, limit inappropriate inflammatory cell death, or correct underlying metabolic vulnerabilities may enable partial reprogramming of immune aging.
Ultimately, understanding immune aging will require moving beyond single, linear pathways and instead examining how cell death networks are regulated as an integrated system across different cell types and stages of aging. This broader view suggests that immune aging is not fixed or inevitable, but rather a modifiable state shaped by how cell fate decisions are controlled. Reframing immune aging in this way may open new opportunities to reduce chronic inflammation and preserve healthy immune function throughout life.
Author Contributions
Conceptualization, T.-B.K.; writing—original draft preparation, H.-M.L. and T.-B.K.; writing—review and editing, E.-J.K., A.H. and T.-B.K.; visualization, H.-M.L. and T.-B.K.; supervision, T.-B.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Konkuk University, grant number 2023-A019-0130.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study.
Acknowledgments
This paper was supported by Konkuk University in 2023.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Goronzy, J.J.; Weyand, C.M. Understanding Immunosenescence to Improve Responses to Vaccines. Nat. Immunol. 2013, 14, 428–436. [Google Scholar] [CrossRef]
- Liu, Z.; Liang, Q.; Ren, Y.; Guo, C.; Ge, X.; Wang, L.; Cheng, Q.; Luo, P.; Zhang, Y.; Han, X. Immunosenescence: Molecular Mechanisms and Diseases. Signal. Transduct. Target. Ther. 2023, 8, 200. [Google Scholar] [CrossRef] [PubMed]
- Nikolich-Zugich, J. The Twilight of Immunity: Emerging Concepts in Aging of the Immune System. Nat. Immunol. 2018, 19, 10–19. [Google Scholar] [CrossRef]
- Franceschi, C.; Garagnani, P.; Parini, P.; Giuliani, C.; Santoro, A. Inflammaging: A New Immune-Metabolic Viewpoint for Age-Related Diseases. Nat. Rev. Endocrinol. 2018, 14, 576–590. [Google Scholar] [CrossRef] [PubMed]
- Chambers, E.S.; Akbar, A.N. Can Blocking Inflammation Enhance Immunity during Aging? J. Allergy Clin. Immunol. 2020, 145, 1323–1331. [Google Scholar] [CrossRef]
- Fulop, T.; Larbi, A.; Dupuis, G.; Le Page, A.; Frost, E.H.; Cohen, A.A.; Witkowski, J.M.; Franceschi, C. Immunosenescence and Inflamm-Aging as Two Sides of the Same Coin: Friends or Foes? Front. Immunol. 2018, 8, 1960. [Google Scholar] [CrossRef]
- Ferrucci, L.; Fabbri, E. Inflammageing: Chronic Inflammation in Ageing, Cardiovascular Disease, and Frailty. Nat. Rev. Cardiol. 2018, 15, 505–522. [Google Scholar] [CrossRef]
- Hotamisligil, G.S. Inflammation and Metabolic Disorders. Nature 2006, 444, 860–867. [Google Scholar] [CrossRef]
- Wyss-Coray, T. Ageing, Neurodegeneration and Brain Rejuvenation. Nature 2016, 539, 180–186. [Google Scholar] [CrossRef]
- Franceschi, C.; Capri, M.; Monti, D.; Giunta, S.; Olivieri, F.; Sevini, F.; Panourgia, M.P.; Invidia, L.; Celani, L.; Scurti, M.; et al. Inflammaging and Anti-Inflammaging: A Systemic Perspective on Aging and Longevity Emerged from Studies in Humans. Mech. Ageing Dev. 2007, 128, 92–105. [Google Scholar] [CrossRef] [PubMed]
- Mittelbrunn, M.; Kroemer, G. Hallmarks of T Cell Aging. Nat. Immunol. 2021, 22, 687–698. [Google Scholar] [CrossRef]
- Nguyen, T.Q.T.; Cho, K.A. Targeting Immunosenescence and Inflammaging: Advancing Longevity Research. Exp. Mol. Med. 2025, 57, 1881–1892. [Google Scholar] [CrossRef]
- Lin, H.; Xiong, W.; Fu, L.; Yi, J.; Yang, J. Damage-Associated Molecular Patterns (DAMPs) in Diseases: Implications for Therapy. Mol. Biomed. 2025, 6, 60–63. [Google Scholar] [CrossRef]
- Ma, M.; Jiang, W.; Zhou, R. DAMPs and DAMP-Sensing Receptors in Inflammation and Diseases. Immunity 2024, 57, 752–771. [Google Scholar] [CrossRef]
- Baechle, J.J.; Chen, N.; Makhijani, P.; Winer, S.; Furman, D.; Winer, D.A. Chronic Inflammation and the Hallmarks of Aging. Mol. Metab. 2023, 74, 101755. [Google Scholar] [CrossRef] [PubMed]
- Furman, D.; Campisi, J.; Verdin, E.; Carrera-Bastos, P.; Targ, S.; Franceschi, C.; Ferrucci, L.; Gilroy, D.W.; Fasano, A.; Miller, G.W.; et al. Chronic Inflammation in the Etiology of Disease Across the Life Span. Nat. Med. 2019, 25, 1822–1832. [Google Scholar] [CrossRef] [PubMed]
- Gielecinska, A.; Kciuk, M.; Yahya, E.; Ainane, T.; Mujwar, S.; Kontek, R. Apoptosis, Necroptosis, and Pyroptosis as Alternative Cell Death Pathways Induced by Chemotherapeutic Agents? Biochim. Biophys. Acta Rev. Cancer 2023, 1878, 189024. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Kanneganti, T. Inflammatory Cell Death in Intestinal Pathologies. Immunol. Rev. 2017, 280, 57–73. [Google Scholar] [CrossRef]
- Korovila, I.; Hugo, M.; Castro, J.P.; Weber, D.; Hohn, A.; Grune, T.; Jung, T. Proteostasis, Oxidative Stress and Aging. Redox Biol. 2017, 13, 550–567. [Google Scholar] [CrossRef]
- Calabrese, V.; Cornelius, C.; Mancuso, C.; Lentile, R.; Stella, A.M.G.; Butterfield, D.A. Redox Homeostasis and Cellular Stress Response in Aging and Neurodegeneration. Methods Mol. Biol. 2010, 610, 285–308. [Google Scholar]
- Srivastava, S. The Mitochondrial Basis of Aging and Age-Related Disorders. Genes 2017, 8, 398. [Google Scholar] [CrossRef] [PubMed]
- Haigis, M.C.; Yankner, B.A. The Aging Stress Response. Mol. Cell 2010, 40, 333–344. [Google Scholar] [CrossRef]
- Hamidu, S.; Amponsah, S.K.; Aning, A.; Adams, L.; Kumi, J.; Ampem-Danso, E.; Hamidu, F.; Mumin Mohammed, M.A.; Ador, G.T.; Khatun, S. Modulating Ferroptosis in Aging: The Therapeutic Potential of Natural Products. J. Aging Res. 2025, 2025, 8832992. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Wang, J.; Zhao, Z.; Xin, T.; Fan, X.; Shen, Q.; Raheem, A.; Lee, C.R.; Jiang, H.; Ding, J. Regulated Necrosis, a Proinflammatory Cell Death, Potentially Counteracts Pathogenic Infections. Cell Death Dis. 2022, 13, 637. [Google Scholar] [CrossRef]
- Li, X.; Li, C.; Zhang, W.; Wang, Y.; Qian, P.; Huang, H. Inflammation and Aging: Signaling Pathways and Intervention Therapies. Signal Transduct. Target. Ther. 2023, 8, 239. [Google Scholar] [CrossRef]
- Royce, G.H.; Brown-Borg, H.M.; Deepa, S.S. The Potential Role of Necroptosis in Inflammaging and Aging. Geroscience 2019, 41, 795–811. [Google Scholar] [CrossRef]
- Su, T.; Hauenstein, J.; Somuncular, E.; Dumral, O.; Leonard, E.; Gustafsson, C.; Tzortzis, E.; Forlani, A.; Johansson, A.; Qian, H.; et al. Aging is Associated with Functional and Molecular Changes in Distinct Hematopoietic Stem Cell Subsets. Nat. Commun. 2024, 15, 7966. [Google Scholar] [CrossRef]
- Kasbekar, M.; Mitchell, C.A.; Proven, M.A.; Passegue, E. Hematopoietic Stem Cells through the Ages: A Lifetime of Adaptation to Organismal Demands. Cell Stem Cell 2023, 30, 1403–1420. [Google Scholar] [CrossRef]
- Mejia-Ramirez, E.; Florian, M.C. Understanding Intrinsic Hematopoietic Stem Cell Aging. Haematologica 2020, 105, 22–37. [Google Scholar] [CrossRef]
- Sun, D.; Luo, M.; Jeong, M.; Rodriguez, B.; Xia, Z.; Hannah, R.; Wang, H.; Le, T.; Faull, K.F.; Chen, R.; et al. Epigenomic Profiling of Young and Aged HSCs Reveals Concerted Changes during Aging that Reinforce Self-Renewal. Cell Stem Cell 2014, 14, 673–688. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.; Nerlov, C. Epigenetic Regulation of Hematopoietic Stem Cell Fate. Trends Cell Biol. 2025, 35, 217–229. [Google Scholar] [CrossRef] [PubMed]
- Mansell, E.; Sigurdsson, V.; Deltcheva, E.; Brown, J.; James, C.; Miharada, K.; Soneji, S.; Larsson, J.; Enver, T. Mitochondrial Potentiation Ameliorates Age-Related Heterogeneity in Hematopoietic Stem Cell Function. Cell Stem Cell 2021, 28, 241–256.e6. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Octaviani, S.; Tian, Z.; Wang, S.R.; Huang, C.; Huang, J. Mitochondrial Quality Control in Hematopoietic Stem Cells: Mechanisms, Implications, and Therapeutic Opportunities. Stem Cell Res. Ther. 2025, 16, 180. [Google Scholar] [CrossRef]
- Morganti, C.; Ito, K. Mitochondrial Contributions to Hematopoietic Stem Cell Aging. Int. J. Mol. Sci. 2021, 22, 11117. [Google Scholar] [CrossRef]
- Puri, G.; Blanc, R.S. The Systemic Costs of Hematopoietic Stem Cell Aging. Development 2025, 152, dev205103. [Google Scholar] [CrossRef]
- Xu, X.; Pang, Y.; Fan, X. Mitochondria in Oxidative Stress, Inflammation and Aging: From Mechanisms to Therapeutic Advances. Signal Transduct. Target. Ther. 2025, 10, 190. [Google Scholar] [CrossRef]
- Nissanka, N.; Moraes, C.T. Mitochondrial DNA Damage and Reactive Oxygen Species in Neurodegenerative Disease. FEBS Lett. 2018, 592, 728–742. [Google Scholar] [CrossRef]
- Yarosz, E.L.; Chang, C. The Role of Reactive Oxygen Species in Regulating T Cell-Mediated Immunity and Disease. Immune Netw. 2018, 18, e14. [Google Scholar] [CrossRef]
- Zhang, X.; Gao, Y.; Zhang, S.; Wang, Y.; Pei, X.; Chen, Y.; Zhang, J.; Zhang, Y.; Du, Y.; Hao, S.; et al. Mitochondrial Dysfunction in the Regulation of Aging and Aging-Related Diseases. Cell Commun. Signal. 2025, 23, 290. [Google Scholar] [CrossRef] [PubMed]
- Su, L.; Zhang, J.; Gomez, H.; Murugan, R.; Hong, X.; Xu, D.; Jiang, F.; Peng, Z. Reactive Oxygen Species-Induced Lipid Peroxidation in Apoptosis, Autophagy, and Ferroptosis. Oxidative Med. Cell. Longev. 2019, 2019, 5080843. [Google Scholar] [CrossRef]
- Ru, Q.; Li, Y.; Chen, L.; Wu, Y.; Min, J.; Wang, F. Iron Homeostasis and Ferroptosis in Human Diseases: Mechanisms and Therapeutic Prospects. Signal Transduct. Target. Ther. 2024, 9, 271. [Google Scholar] [CrossRef]
- Gupta, S.; Cassel, S.L.; Sutterwala, F.S.; Dagvadorj, J. Regulation of the NLRP3 Inflammasome by Autophagy and Mitophagy. Immunol. Rev. 2025, 329, e13410. [Google Scholar] [CrossRef]
- Kim, S.R.; Kim, D.I.; Kim, S.H.; Lee, H.; Lee, K.S.; Cho, S.H.; Lee, Y.C. NLRP3 Inflammasome Activation by Mitochondrial ROS in Bronchial Epithelial Cells is Required for Allergic Inflammation. Cell Death Dis. 2014, 5, e1498. [Google Scholar] [CrossRef]
- Zhao, P.; Yin, S.; Qiu, Y.; Sun, C.; Yu, H. Ferroptosis and Pyroptosis are Connected through Autophagy: A New Perspective of Overcoming Drug Resistance. Mol. Cancer 2025, 24, 23. [Google Scholar] [CrossRef]
- Fu, W.; Liu, S.; Xu, T.; Liu, Y.; Zhang, T.; Liu, D.; Wang, X.; Wang, J.; He, Y.; Ma, T. Emodin Inhibits NLRP3 Inflammasome Activation and Protects Against Sepsis Via Promoting FUNDC1-Mediated Mitophagy. Int. J. Biol. Sci. 2025, 21, 3631–3648. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, M.; Wang, R.; Lan, J.; Yu, L.; Gao, J.; Lu, H.; Fang, Q.; Wang, F. Mitophagy Protects Against Silver Nanoparticle-Induced Hepatotoxicity by Inhibiting Mitochondrial ROS and the NLRP3 Inflammasome. Ecotoxicol. Environ. Saf. 2024, 273, 116137. [Google Scholar] [CrossRef]
- Casey, A.M.; Ryan, D.G.; Prag, H.A.; Chowdhury, S.R.; Marques, E.; Turner, K.; Gruszczyk, A.V.; Yang, M.; Wolf, D.M.; Miljkovic, J.L.; et al. Pro-Inflammatory Macrophages Produce Mitochondria-Derived Superoxide by Reverse Electron Transport at Complex I that Regulates IL-1beta Release during NLRP3 Inflammasome Activation. Nat. Metab. 2025, 7, 493–507. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Xiang, Y.; Liu, S.; Li, C.; Dong, J.; Kong, X.; Ji, X.; Cheng, X.; Zhang, L. RIPK3 Signaling and its Role in Regulated Cell Death and Diseases. Cell Death Discov. 2024, 10, 200. [Google Scholar] [CrossRef] [PubMed]
- Coperchini, F.; Greco, A.; Teliti, M.; Croce, L.; Chytiris, S.; Magri, F.; Gaetano, C.; Rotondi, M. Inflamm-Ageing: How Cytokines and Nutrition Shape the Trajectory of Ageing. Cytokine Growth Factor Rev. 2025, 82, 31–42. [Google Scholar] [CrossRef]
- Poon, I.K.H.; Ravichandran, K.S. Targeting Efferocytosis in Inflammaging. Annu. Rev. Pharmacol. Toxicol. 2024, 64, 339–357. [Google Scholar] [CrossRef]
- Hu, H.; Cheng, X.; Li, F.; Guan, Z.; Xu, J.; Wu, D.; Gao, Y.; Zhan, X.; Wang, P.; Zhou, H.; et al. Defective Efferocytosis by Aged Macrophages Promotes STING Signaling Mediated Inflammatory Liver Injury. Cell Death Discov. 2023, 9, 236. [Google Scholar] [CrossRef]
- Quarato, E.R.; Salama, N.A.; Li, A.J.; Smith, C.O.; Zhang, J.; Kawano, Y.; McArthur, M.; Liesveld, J.L.; Becker, M.W.; Elliott, M.R.; et al. Efferocytosis by Bone Marrow Mesenchymal Stromal Cells Disrupts Osteoblastic Differentiation Via Mitochondrial Remodeling. Cell Death Dis. 2023, 14, 428. [Google Scholar] [CrossRef]
- Sachet, M.; Liang, Y.Y.; Oehler, R. The Immune Response to Secondary Necrotic Cells. Apoptosis 2017, 22, 1189–1204. [Google Scholar] [CrossRef]
- Ge, Y.; Huang, M.; Yao, Y. Efferocytosis and its Role in Inflammatory Disorders. Front. Cell Dev. Biol. 2022, 10, 839248. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Conrad, M. Ferroptosis: When Metabolism Meets Cell Death. Physiol. Rev. 2025, 105, 651–706. [Google Scholar] [CrossRef]
- Gao, J.; Xiong, A.; Liu, J.; Li, X.; Wang, J.; Zhang, L.; Liu, Y.; Xiong, Y.; Li, G.; He, X. PANoptosis: Bridging Apoptosis, Pyroptosis, and Necroptosis in Cancer Progression and Treatment. Cancer Gene Ther. 2024, 31, 970–983. [Google Scholar] [CrossRef]
- Bell, H.N.; Stockwell, B.R.; Zou, W. Ironing Out the Role of Ferroptosis in Immunity. Immunity 2024, 57, 941–956. [Google Scholar] [CrossRef]
- Mohammed, S.; Ohene-Marfo, P.; Jiang, C.; Peng, Z.; Thadathil, N.; Tran, A.; Nicklas, E.; Bhaskaran, S.; Wang, D.; Selvarani, R.; et al. Impact of Mlkl Or Ripk3 Deletion on Age-Associated Liver Inflammation, Metabolic Health, and Lifespan. Geroscience 2025, 47, 4465–4483. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Dong, R.; Yuan, B.; Xie, Y.; Feng, Z.; Zhou, S.; Luan, F.; Chen, Y. Immune Cells Dying from Ferroptosis: Mechanisms and Therapeutic Opportunities. Cell Death Dis. 2025, 16, 878. [Google Scholar] [CrossRef]
- Hildeman, D.; Jorgensen, T.; Kappler, J.; Marrack, P. Apoptosis and the Homeostatic Control of Immune Responses. Curr. Opin. Immunol. 2007, 19, 516–521. [Google Scholar] [CrossRef] [PubMed]
- Newton, K.; Strasser, A.; Kayagaki, N.; Dixit, V.M. Cell Death. Cell 2024, 187, 235–256. [Google Scholar] [CrossRef]
- Fadok, V.A.; Bratton, D.L.; Konowal, A.; Freed, P.W.; Westcott, J.Y.; Henson, P.M. Macrophages that have Ingested Apoptotic Cells in Vitro Inhibit Proinflammatory Cytokine Production through Autocrine/Paracrine Mechanisms Involving TGF-Beta, PGE2, and PAF. J. Clin. Investig. 1998, 101, 890–898. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Wang, B.; Alu, A.; Hong, W.; Lei, H.; He, X.; Shi, H.; Cheng, P.; Yang, X. Immunosenescence: Signaling Pathways, Diseases and Therapeutic Targets. Signal Transduct. Target. Ther. 2025, 10, 250. [Google Scholar] [CrossRef]
- Wrona, M.V.; Ghosh, R.; Coll, K.; Chun, C.; Yousefzadeh, M.J. The 3 I’s of Immunity and Aging: Immunosenescence, Inflammaging, and Immune Resilience. Front. Aging 2024, 5, 1490302. [Google Scholar] [CrossRef] [PubMed]
- Goronzy, J.J.; Weyand, C.M. Mechanisms Underlying T Cell Ageing. Nat. Rev. Immunol. 2019, 19, 573–583. [Google Scholar] [CrossRef]
- Quiros-Roldan, E.; Sottini, A.; Natali, P.G.; Imberti, L. The Impact of Immune System Aging on Infectious Diseases. Microorganisms 2024, 12, 775. [Google Scholar] [CrossRef] [PubMed]
- Soto-Heredero, G.; Gomez de Las Heras, M.M.; Escrig-Larena, J.I.; Mittelbrunn, M. Extremely Differentiated T Cell Subsets Contribute to Tissue Deterioration during Aging. Annu. Rev. Immunol. 2023, 41, 181–205. [Google Scholar] [CrossRef]
- Beltra, J.; Manne, S.; Abdel-Hakeem, M.S.; Kurachi, M.; Giles, J.R.; Chen, Z.; Casella, V.; Ngiow, S.F.; Khan, O.; Huang, Y.J.; et al. Developmental Relationships of Four Exhausted CD8(+) T Cell Subsets Reveals Underlying Transcriptional and Epigenetic Landscape Control Mechanisms. Immunity 2020, 52, 825–841.e8. [Google Scholar] [CrossRef]
- Wherry, E.J.; Kurachi, M. Molecular and Cellular Insights into T Cell Exhaustion. Nat. Rev. Immunol. 2015, 15, 486–499. [Google Scholar] [CrossRef]
- Goronzy, J.J.; Weyand, C.M. Successful and Maladaptive T Cell Aging. Immunity 2017, 46, 364–378. [Google Scholar] [CrossRef]
- Han, S.; Georgiev, P.; Ringel, A.E.; Sharpe, A.H.; Haigis, M.C. Age-Associated Remodeling of T Cell Immunity and Metabolism. Cell Metab. 2023, 35, 36–55. [Google Scholar] [CrossRef]
- Donnini, A.; Re, F.; Bollettini, M.; Moresi, R.; Tesei, S.; Bernardini, G.; Provinciali, M. Age-Related Susceptibility of Naive and Memory CD4 T Cells to Apoptosis Induced by IL-2 Deprivation Or PHA Addition. Biogerontology 2005, 6, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Lechuga-Vieco, A.V.; Sattentau, C.; Borsa, M.; Simon, A.K. Dysfunctional Mitochondria in Ageing T Cells: A Perspective on Mitochondrial Quality Control Mechanisms. EMBO Rep. 2025, 26, 4402–4418. [Google Scholar] [CrossRef]
- Weng, N. Numbers and Odds: TCR Repertoire Size and its Age Changes Impacting on T Cell Functions. Semin. Immunol. 2023, 69, 101810. [Google Scholar] [CrossRef]
- Escrig-Larena, J.I.; Delgado-Pulido, S.; Mittelbrunn, M. Mitochondria during T Cell Aging. Semin. Immunol. 2023, 69, 101808. [Google Scholar] [CrossRef] [PubMed]
- Koenen, P.; Heinzel, S.; Carrington, E.M.; Happo, L.; Alexander, W.S.; Zhang, J.; Herold, M.J.; Scott, C.L.; Lew, A.M.; Strasser, A.; et al. Mutually Exclusive Regulation of T Cell Survival by IL-7R and Antigen Receptor-Induced Signals. Nat. Commun. 2013, 4, 1735. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, A.; Sridharan, A.; Prakash, S.; Agrawal, H. Dendritic Cells and Aging: Consequences for Autoimmunity. Expert Rev. Clin. Immunol. 2012, 8, 73–80. [Google Scholar] [CrossRef]
- Dho, S.H.; Cho, M.; Woo, W.; Jeong, S.; Kim, L.K. Caspases as Master Regulators of Programmed Cell Death: Apoptosis, Pyroptosis and Beyond. Exp. Mol. Med. 2025, 57, 1121–1132. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Wei, S.; Nguyen, T.H.; Jo, Y.; Zhang, Y.; Park, W.; Gariani, K.; Oh, C.; Kim, H.H.; Ha, K.; et al. Mitochondria-Associated Programmed Cell Death as a Therapeutic Target for Age-Related Disease. Exp. Mol. Med. 2023, 55, 1595–1619. [Google Scholar] [CrossRef]
- Stutz, M.D.; Allison, C.C.; Ojaimi, S.; Preston, S.P.; Doerflinger, M.; Arandjelovic, P.; Whitehead, L.; Bader, S.M.; Batey, D.; Asselin-Labat, M.; et al. Macrophage and Neutrophil Death Programs Differentially Confer Resistance to Tuberculosis. Immunity 2021, 54, 1758–1771.e7. [Google Scholar] [CrossRef]
- Pang, J.; Vince, J.E. The Role of Caspase-8 in Inflammatory Signalling and Pyroptotic Cell Death. Semin. Immunol. 2023, 70, 101832. [Google Scholar] [CrossRef]
- Yang, Y.; Li, X.; Zhang, T.; Xu, D. RIP Kinases and Necroptosis in Aging and Aging-Related Diseases. Life Med. 2022, 1, 2–20. [Google Scholar] [CrossRef] [PubMed]
- Schwarzer, R.; Laurien, L.; Pasparakis, M. New Insights into the Regulation of Apoptosis, Necroptosis, and Pyroptosis by Receptor Interacting Protein Kinase 1 and Caspase-8. Curr. Opin. Cell Biol. 2020, 63, 186–193. [Google Scholar] [CrossRef]
- Gunther, C.; Martini, E.; Wittkopf, N.; Amann, K.; Weigmann, B.; Neumann, H.; Waldner, M.J.; Hedrick, S.M.; Tenzer, S.; Neurath, M.F.; et al. Caspase-8 Regulates TNF-Alpha-Induced Epithelial Necroptosis and Terminal Ileitis. Nature 2011, 477, 335–339. [Google Scholar] [CrossRef]
- Robinson, N.; McComb, S.; Mulligan, R.; Dudani, R.; Krishnan, L.; Sad, S. Type I Interferon Induces Necroptosis in Macrophages during Infection with Salmonella Enterica Serovar Typhimurium. Nat. Immunol. 2012, 13, 954–962. [Google Scholar] [CrossRef]
- Oberst, A.; Dillon, C.P.; Weinlich, R.; McCormick, L.L.; Fitzgerald, P.; Pop, C.; Hakem, R.; Salvesen, G.S.; Green, D.R. Catalytic Activity of the Caspase-8-FLIP(L) Complex Inhibits RIPK3-Dependent Necrosis. Nature 2011, 471, 363–367. [Google Scholar] [CrossRef]
- Rickard, J.A.; O’Donnell, J.A.; Evans, J.M.; Lalaoui, N.; Poh, A.R.; Rogers, T.; Vince, J.E.; Lawlor, K.E.; Ninnis, R.L.; Anderton, H.; et al. RIPK1 Regulates RIPK3-MLKL-Driven Systemic Inflammation and Emergency Hematopoiesis. Cell 2014, 157, 1175–1188. [Google Scholar] [CrossRef] [PubMed]
- Morgan, M.J.; Kim, Y. Roles of RIPK3 in Necroptosis, Cell Signaling, and Disease. Exp. Mol. Med. 2022, 54, 1695–1704. [Google Scholar] [CrossRef]
- Chen, R.; Zou, J.; Liu, J.; Kang, R.; Tang, D. DAMPs in the Immunogenicity of Cell Death. Mol. Cell 2025, 85, 3874–3889. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.T. Secondary Necrosis: The Natural Outcome of the Complete Apoptotic Program. FEBS Lett. 2010, 584, 4491–4499. [Google Scholar] [CrossRef]
- Szondy, Z.; Garabuczi, E.; Joos, G.; Tsay, G.J.; Sarang, Z. Impaired Clearance of Apoptotic Cells in Chronic Inflammatory Diseases: Therapeutic Implications. Front. Immunol. 2014, 5, 354. [Google Scholar] [CrossRef]
- Mann, V.; Sundaresan, A.; Shishodia, S. Overnutrition and Lipotoxicity: Impaired Efferocytosis and Chronic Inflammation as Precursors to Multifaceted Disease Pathogenesis. Biology 2024, 13, 241. [Google Scholar] [CrossRef]
- Shi, H.; Moore, M.P.; Wang, X.; Tabas, I. Efferocytosis in Liver Disease. JHEP Rep. 2023, 6, 100960. [Google Scholar] [CrossRef]
- Wang, Y.; Kanneganti, T. From Pyroptosis, Apoptosis and Necroptosis to PANoptosis: A Mechanistic Compendium of Programmed Cell Death Pathways. Comput. Struct. Biotechnol. J. 2021, 19, 4641–4657. [Google Scholar] [CrossRef] [PubMed]
- Bertheloot, D.; Latz, E.; Franklin, B.S. Necroptosis, Pyroptosis and Apoptosis: An Intricate Game of Cell Death. Cell. Mol. Immunol. 2021, 18, 1106–1121. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Otin, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. Hallmarks of Aging: An Expanding Universe. Cell 2023, 186, 243–278. [Google Scholar] [CrossRef]
- Karki, R.; Sharma, B.R.; Tuladhar, S.; Williams, E.P.; Zalduondo, L.; Samir, P.; Zheng, M.; Sundaram, B.; Banoth, B.; Malireddi, R.K.S.; et al. Synergism of TNF-Alpha and IFN-Gamma Triggers Inflammatory Cell Death, Tissue Damage, and Mortality in SARS-CoV-2 Infection and Cytokine Shock Syndromes. Cell 2021, 184, 149–168.e17. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Wang, H.; Wang, Z.; He, S.; Chen, S.; Liao, D.; Wang, L.; Yan, J.; Liu, W.; Lei, X.; et al. Mixed Lineage Kinase Domain-Like Protein Mediates Necrosis Signaling Downstream of RIP3 Kinase. Cell 2012, 148, 213–227. [Google Scholar] [CrossRef]
- Cho, Y.S.; Challa, S.; Moquin, D.; Genga, R.; Ray, T.D.; Guildford, M.; Chan, F.K. Phosphorylation-Driven Assembly of the RIP1-RIP3 Complex Regulates Programmed Necrosis and Virus-Induced Inflammation. Cell 2009, 137, 1112–1123. [Google Scholar] [CrossRef]
- Orzalli, M.H.; Kagan, J.C. Apoptosis and Necroptosis as Host Defense Strategies to Prevent Viral Infection. Trends Cell Biol. 2017, 27, 800–809. [Google Scholar] [CrossRef]
- Yu, X.; Yuan, J.; Shi, L.; Dai, S.; Yue, L.; Yan, M. Necroptosis in Bacterial Infections. Front. Immunol. 2024, 15, 1394857. [Google Scholar] [CrossRef]
- Nailwal, H.; Chan, F.K. Necroptosis in Anti-Viral Inflammation. Cell Death Differ. 2019, 26, 4–13. [Google Scholar] [CrossRef]
- Tovey Crutchfield, E.C.; Garnish, S.E.; Day, J.; Anderton, H.; Chiou, S.; Hempel, A.; Hall, C.; Patel, K.M.; Gangatirkar, P.; Martin, K.R.; et al. MLKL Deficiency Protects Against Low-Grade, Sterile Inflammation in Aged Mice. Cell Death Differ. 2023, 30, 1059–1071. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, S.; Thadathil, N.; Selvarani, R.; Nicklas, E.H.; Wang, D.; Miller, B.F.; Richardson, A.; Deepa, S.S. Necroptosis Contributes to Chronic Inflammation and Fibrosis in Aging Liver. Aging Cell 2021, 20, e13512. [Google Scholar] [CrossRef] [PubMed]
- Nair, B.; Menon, A.; Khader, M.A.; Sethi, G.; Nath, L.R. Linking Necroptosis with Liver Aging and Chronic Inflammation in Hepatic Pathology. Life Sci. 2025, 379, 123871. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, W.J.; Upton, J.W.; Long, A.B.; Livingston-Rosanoff, D.; Daley-Bauer, L.P.; Hakem, R.; Caspary, T.; Mocarski, E.S. RIP3 Mediates the Embryonic Lethality of Caspase-8-Deficient Mice. Nature 2011, 471, 368–372. [Google Scholar] [CrossRef]
- Vandenabeele, P.; Galluzzi, L.; Vanden Berghe, T.; Kroemer, G. Molecular Mechanisms of Necroptosis: An Ordered Cellular Explosion. Nat. Rev. Mol. Cell Biol. 2010, 11, 700–714. [Google Scholar] [CrossRef]
- Jantas, D.; Lason, W. Preclinical Evidence for the Interplay between Oxidative Stress and RIP1-Dependent Cell Death in Neurodegeneration: State of the Art and Possible Therapeutic Implications. Antioxidants 2021, 10, 1518. [Google Scholar] [CrossRef]
- Zhang, Y.; Su, S.S.; Zhao, S.; Yang, Z.; Zhong, C.; Chen, X.; Cai, Q.; Yang, Z.; Huang, D.; Wu, R.; et al. RIP1 Autophosphorylation is Promoted by Mitochondrial ROS and is Essential for RIP3 Recruitment into Necrosome. Nat. Commun. 2017, 8, 14329. [Google Scholar] [CrossRef]
- Wang, H.; Sun, L.; Su, L.; Rizo, J.; Liu, L.; Wang, L.; Wang, F.; Wang, X. Mixed Lineage Kinase Domain-Like Protein MLKL Causes Necrotic Membrane Disruption upon Phosphorylation by RIP3. Mol. Cell 2014, 54, 133–146. [Google Scholar] [CrossRef]
- Chen, X.; Li, W.; Ren, J.; Huang, D.; He, W.; Song, Y.; Yang, C.; Li, W.; Zheng, X.; Chen, P.; et al. Translocation of Mixed Lineage Kinase Domain-Like Protein to Plasma Membrane Leads to Necrotic Cell Death. Cell Res. 2014, 24, 105–121. [Google Scholar] [CrossRef]
- Cai, Z.; Jitkaew, S.; Zhao, J.; Chiang, H.; Choksi, S.; Liu, J.; Ward, Y.; Wu, L.; Liu, Z. Plasma Membrane Translocation of Trimerized MLKL Protein is Required for TNF-Induced Necroptosis. Nat. Cell Biol. 2014, 16, 55–65. [Google Scholar] [CrossRef]
- Schuermans, S.; Kestens, C.; Marques, P.E. Systemic Mechanisms of Necrotic Cell Debris Clearance. Cell Death Dis. 2024, 15, 557. [Google Scholar] [CrossRef] [PubMed]
- Kaczmarek, A.; Vandenabeele, P.; Krysko, D.V. Necroptosis: The Release of Damage-Associated Molecular Patterns and its Physiological Relevance. Immunity 2013, 38, 209–223. [Google Scholar] [CrossRef] [PubMed]
- Gong, T.; Liu, L.; Jiang, W.; Zhou, R. DAMP-Sensing Receptors in Sterile Inflammation and Inflammatory Diseases. Nat. Rev. Immunol. 2020, 20, 95–112. [Google Scholar] [CrossRef]
- Chen, G.Y.; Nunez, G. Sterile Inflammation: Sensing and Reacting to Damage. Nat. Rev. Immunol. 2010, 10, 826–837. [Google Scholar] [CrossRef]
- Takeuchi, O.; Akira, S. Pattern Recognition Receptors and Inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef]
- Lee, E.; Song, C.; Bae, S.; Ha, K.; Karki, R. Regulated Cell Death Pathways and their Roles in Homeostasis, Infection, Inflammation, and Tumorigenesis. Exp. Mol. Med. 2023, 55, 1632–1643. [Google Scholar] [CrossRef]
- Lee, K.; Kang, T. The Molecular Links between Cell Death and Inflammasome. Cells 2019, 8, 1057. [Google Scholar] [CrossRef]
- Frank, D.; Vince, J.E. Pyroptosis Versus Necroptosis: Similarities, Differences, and Crosstalk. Cell Death Differ. 2019, 26, 99–114. [Google Scholar] [CrossRef] [PubMed]
- Gaidt, M.M.; Hornung, V. Pore Formation by GSDMD is the Effector Mechanism of Pyroptosis. EMBO J. 2016, 35, 2167–2169. [Google Scholar] [CrossRef]
- Sborgi, L.; Ruhl, S.; Mulvihill, E.; Pipercevic, J.; Heilig, R.; Stahlberg, H.; Farady, C.J.; Muller, D.J.; Broz, P.; Hiller, S. GSDMD Membrane Pore Formation Constitutes the Mechanism of Pyroptotic Cell Death. EMBO J. 2016, 35, 1766–1778. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, Z.; Ruan, J.; Pan, Y.; Magupalli, V.G.; Wu, H.; Lieberman, J. Inflammasome-Activated Gasdermin D Causes Pyroptosis by Forming Membrane Pores. Nature 2016, 535, 153–158. [Google Scholar] [CrossRef]
- Latz, E.; Duewell, P. NLRP3 Inflammasome Activation in Inflammaging. Semin. Immunol. 2018, 40, 61–73. [Google Scholar] [CrossRef]
- Stout-Delgado, H.W.; Cho, S.J.; Chu, S.G.; Mitzel, D.N.; Villalba, J.; El-Chemaly, S.; Ryter, S.W.; Choi, A.M.K.; Rosas, I.O. Age-Dependent Susceptibility to Pulmonary Fibrosis is Associated with NLRP3 Inflammasome Activation. Am. J. Respir. Cell Mol. Biol. 2016, 55, 252–263. [Google Scholar] [CrossRef]
- Youm, Y.; Grant, R.W.; McCabe, L.R.; Albarado, D.C.; Nguyen, K.Y.; Ravussin, A.; Pistell, P.; Newman, S.; Carter, R.; Laque, A.; et al. Canonical Nlrp3 Inflammasome Links Systemic Low-Grade Inflammation to Functional Decline in Aging. Cell Metab. 2013, 18, 519–532. [Google Scholar] [CrossRef]
- Gritsenko, A.; Green, J.P.; Brough, D.; Lopez-Castejon, G. Mechanisms of NLRP3 Priming in Inflammaging and Age Related Diseases. Cytokine Growth Factor Rev. 2020, 55, 15–25. [Google Scholar] [CrossRef]
- Garcia-Garcia, V.A.; Alameda, J.P.; Page, A.; Casanova, M.L. Role of NF-kappaB in Ageing and Age-Related Diseases: Lessons from Genetically Modified Mouse Models. Cells 2021, 10, 1906. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.Y.; Kim, D.H.; Lee, E.K.; Chung, K.W.; Chung, S.; Lee, B.; Seo, A.Y.; Chung, J.H.; Jung, Y.S.; Im, E.; et al. Redefining Chronic Inflammation in Aging and Age-Related Diseases: Proposal of the Senoinflammation Concept. Aging Dis. 2019, 10, 367–382. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S. NF-kappaB Signaling in Inflammation. Signal. Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef]
- Dubey, S.R.; Turnbull, C.; Pandey, A.; Zhao, A.; Kurera, M.; Al-Zidan, R.; Shen, C.; Gautam, M.; Mahajan, S.; Jadhav, P.S.; et al. Molecular Mechanisms and Regulation of Inflammasome Activation and Signaling: Sensing of Pathogens and Damage Molecular Patterns. Cell. Mol. Immunol. 2025, 22, 1313–1344. [Google Scholar] [CrossRef]
- Salminen, A.; Ojala, J.; Kaarniranta, K.; Kauppinen, A. Mitochondrial Dysfunction and Oxidative Stress Activate Inflammasomes: Impact on the Aging Process and Age-Related Diseases. Cell. Mol. Life Sci. 2012, 69, 2999–3013. [Google Scholar] [CrossRef] [PubMed]
- Chevriaux, A.; Pilot, T.; Derangere, V.; Simonin, H.; Martine, P.; Chalmin, F.; Ghiringhelli, F.; Rebe, C. Cathepsin B is Required for NLRP3 Inflammasome Activation in Macrophages, through NLRP3 Interaction. Front. Cell Dev. Biol. 2020, 8, 167. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Wang, H.; Kouadir, M.; Song, H.; Shi, F. Recent Advances in the Mechanisms of NLRP3 Inflammasome Activation and its Inhibitors. Cell Death Dis. 2019, 10, 128. [Google Scholar] [CrossRef]
- He, Y.; Hara, H.; Nunez, G. Mechanism and Regulation of NLRP3 Inflammasome Activation. Trends Biochem. Sci. 2016, 41, 1012–1021. [Google Scholar] [CrossRef]
- Liu, Y.; Pan, R.; Ouyang, Y.; Gu, W.; Xiao, T.; Yang, H.; Tang, L.; Wang, H.; Xiang, B.; Chen, P. Pyroptosis in Health and Disease: Mechanisms, Regulation and Clinical Perspective. Signal Transduct. Target. Ther. 2024, 9, 245. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Hong, W.; Zhu, H.; He, Q.; Yang, B.; Wang, J.; Weng, Q. Macrophage Senescence in Health and Diseases. Acta Pharm. Sin. B 2024, 14, 1508–1524. [Google Scholar] [CrossRef]
- McKenzie, B.A.; Mamik, M.K.; Saito, L.B.; Boghozian, R.; Monaco, M.C.; Major, E.O.; Lu, J.; Branton, W.G.; Power, C. Caspase-1 Inhibition Prevents Glial Inflammasome Activation and Pyroptosis in Models of Multiple Sclerosis. Proc. Natl. Acad. Sci. USA 2018, 115, E6065–E6074. [Google Scholar] [CrossRef]
- Heneka, M.T.; Kummer, M.P.; Stutz, A.; Delekate, A.; Schwartz, S.; Vieira-Saecker, A.; Griep, A.; Axt, D.; Remus, A.; Tzeng, T.; et al. NLRP3 is Activated in Alzheimer’s Disease and Contributes to Pathology in APP/PS1 Mice. Nature 2013, 493, 674–678. [Google Scholar] [CrossRef]
- Mittal, R.; Saavedra, D.; Mittal, M.; Lemos, J.R.N.; Hirani, K. Inflammasome Activation and Accelerated Immune Aging in Autoimmune Disorders. Front. Aging 2025, 6, 1688060. [Google Scholar] [CrossRef]
- Lee, G.; Lee, G. Potential Role of Inflammasomes in Aging. Int. J. Mol. Sci. 2025, 26, 6768. [Google Scholar] [CrossRef] [PubMed]
- Liang, R.; Qi, X.; Cai, Q.; Niu, L.; Huang, X.; Zhang, D.; Ling, J.; Wu, Y.; Chen, Y.; Yang, P.; et al. The Role of NLRP3 Inflammasome in Aging and Age-Related Diseases. Immun. Ageing 2024, 21, 14. [Google Scholar] [CrossRef]
- Romero, A.; Dongil, P.; Valencia, I.; Vallejo, S.; Hipolito-Luengo, A.S.; Diaz-Araya, G.; Bartha, J.L.; Gonzalez-Arlanzon, M.M.; Rivilla, F.; de la Cuesta, F.; et al. Pharmacological Blockade of NLRP3 Inflammasome/IL-1beta-Positive Loop Mitigates Endothelial Cell Senescence and Dysfunction. Aging Dis. 2022, 13, 284–297. [Google Scholar] [CrossRef]
- Kumar, S.; Budhathoki, S.; Oliveira, C.B.; Kahle, A.D.; Calhan, O.Y.; Lukens, J.R.; Deppmann, C.D. Role of the Caspase-8/RIPK3 Axis in Alzheimer’s Disease Pathogenesis and Abeta-Induced NLRP3 Inflammasome Activation. JCI Insight 2023, 8, e157433. [Google Scholar] [CrossRef] [PubMed]
- Imanishi, T.; Unno, M.; Yoneda, N.; Motomura, Y.; Mochizuki, M.; Sasaki, T.; Pasparakis, M.; Saito, T. RIPK1 Blocks T Cell Senescence Mediated by RIPK3 and Caspase-8. Sci. Adv. 2023, 9, eadd6097. [Google Scholar] [CrossRef] [PubMed]
- Conos, S.A.; Chen, K.W.; De Nardo, D.; Hara, H.; Whitehead, L.; Nunez, G.; Masters, S.L.; Murphy, J.M.; Schroder, K.; Vaux, D.L.; et al. Active MLKL Triggers the NLRP3 Inflammasome in a Cell-Intrinsic Manner. Proc. Natl. Acad. Sci. USA 2017, 114, E961–E969. [Google Scholar] [CrossRef]
- Lawlor, K.E.; Khan, N.; Mildenhall, A.; Gerlic, M.; Croker, B.A.; D’Cruz, A.A.; Hall, C.; Kaur Spall, S.; Anderton, H.; Masters, S.L.; et al. RIPK3 Promotes Cell Death and NLRP3 Inflammasome Activation in the Absence of MLKL. Nat. Commun. 2015, 6, 6282. [Google Scholar] [CrossRef]
- Thadathil, N.; Nicklas, E.H.; Mohammed, S.; Lewis, T.L.J.; Richardson, A.; Deepa, S.S. Necroptosis Increases with Age in the Brain and Contributes to Age-Related Neuroinflammation. Geroscience 2021, 43, 2345–2361. [Google Scholar] [CrossRef]
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S.; et al. Ferroptosis: An Iron-Dependent Form of Nonapoptotic Cell Death. Cell 2012, 149, 1060–1072. [Google Scholar] [CrossRef]
- Wang, Z.; Dai, Y.; Jin, R.; Mo, D.; Hu, Q.; Huang, Y.; Zhang, L.; Zhang, C.; Gao, H.; Yan, Q. Ferroptosis Activation Contributes to Kidney Aging in Mice by Promoting Tubular Cell Senescence. J. Gerontol. Ser. A 2025, 80, glaf042. [Google Scholar] [CrossRef]
- Mazhar, M.; Din, A.U.; Ali, H.; Yang, G.; Ren, W.; Wang, L.; Fan, X.; Yang, S. Implication of Ferroptosis in Aging. Cell Death Discov. 2021, 7, 149. [Google Scholar] [CrossRef]
- Ezuz, D.; Ombashe, H.; Watad, L.; Rakhymzhanova, A.; Pandey, S.; Atar, O.; Meyron-Holtz, E.G.; Ron-Harel, N. Heme and Iron Toxicity in the Aged Spleen Impairs T Cell Immunity through Iron Deprivation. Nat. Aging 2025, 5, 2247–2262. [Google Scholar] [CrossRef]
- Diaz-Del Cerro, E.; Martinez de Toda, I.; Felix, J.; Baca, A.; De la Fuente, M. Components of the Glutathione Cycle as Markers of Biological Age: An Approach to Clinical Application in Aging. Antioxidants 2023, 12, 1529. [Google Scholar] [CrossRef] [PubMed]
- Marschallinger, J.; Iram, T.; Zardeneta, M.; Lee, S.E.; Lehallier, B.; Haney, M.S.; Pluvinage, J.V.; Mathur, V.; Hahn, O.; Morgens, D.W.; et al. Lipid-Droplet-Accumulating Microglia Represent a Dysfunctional and Proinflammatory State in the Aging Brain. Nat. Neurosci. 2020, 23, 194–208. [Google Scholar] [CrossRef] [PubMed]
- Verduijn, J.; Coutant, K.; Fane, M.E.; Galluzzi, L. Molecular Mechanisms of Immune Cell Death in Immunosenescence. Cell Death Differ. 2025, 32, 1755–1762. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Yang, K.; Yu, G.; Hao, W.; Zhu, X.; Ge, A.; Chen, J.; Sun, L. Advances in Research on Immunocyte Iron Metabolism, Ferroptosis, and their Regulatory Roles in Autoimmune and Autoinflammatory Diseases. Cell Death Dis. 2024, 15, 481. [Google Scholar] [CrossRef]
- Recalcati, S.; Cairo, G. Macrophages and Iron: A Special Relationship. Biomedicines 2021, 9, 1585. [Google Scholar] [CrossRef]
- Winn, N.C.; Volk, K.M.; Hasty, A.H. Regulation of Tissue Iron Homeostasis: The Macrophage “Ferrostat”. JCI Insight 2020, 5, e132964. [Google Scholar] [CrossRef]
- Liu, Y.; Luo, X.; Chen, Y.; Dang, J.; Zeng, D.; Guo, X.; Weng, W.; Zhao, J.; Shi, X.; Chen, J.; et al. Heterogeneous Ferroptosis Susceptibility of Macrophages Caused by Focal Iron Overload Exacerbates Rheumatoid Arthritis. Redox Biol. 2024, 69, 103008. [Google Scholar] [CrossRef]
- Shen, L.; Chen, J.; Tou, J. Inhibition of Ferroptosis in Inflammatory Macrophages Alleviates Intestinal Injury in Neonatal Necrotizing Enterocolitis. Cell Death Discov. 2025, 11, 365. [Google Scholar] [CrossRef]
- Xu, H.; Lu, Y.; Zeng, Q.; Zhu, X.; Guan, W.; Liu, S. Ferroptosis of Immune Cells in Infection, Inflammation and Tumor Progression. Biomolecules 2025, 15, 1464. [Google Scholar] [CrossRef]
- Matsushita, M.; Freigang, S.; Schneider, C.; Conrad, M.; Bornkamm, G.W.; Kopf, M. T Cell Lipid Peroxidation Induces Ferroptosis and Prevents Immunity to Infection. J. Exp. Med. 2015, 212, 555–568. [Google Scholar] [CrossRef]
- Fetarayani, D.; Kahdina, M.; Waitupu, A.; Pratiwi, L.; Ningtyas, M.C.; Adytia, G.J.; Sutanto, H. Immunosenescence and the Geriatric Giants: Molecular Insights into Aging and Healthspan. Med. Sci. 2025, 13, 100. [Google Scholar] [CrossRef]
- Mbah, N.E.; Lyssiotis, C.A. Metabolic Regulation of Ferroptosis in the Tumor Microenvironment. J. Biol. Chem. 2022, 298, 101617. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Xiao, L.; Liu, L.; Ye, L.; Su, P.; Bi, E.; Wang, Q.; Yang, M.; Qian, J.; Yi, Q. CD36-Mediated Ferroptosis Dampens Intratumoral CD8(+) T Cell Effector Function and Impairs their Antitumor Ability. Cell Metab. 2021, 33, 1001–1012.e5. [Google Scholar] [CrossRef] [PubMed]
- Yapici, F.I.; Seidel, E.; Dahlhaus, A.; Weber, J.; Schmidt, C.; de Britto Chaves Filho, A.; Yang, M.; Nenchova, M.; Gungor, E.; Stroh, J.; et al. An Atlas of Ferroptosis-Induced Secretomes. Cell Death Differ. 2025, 32, 1986–2008. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.; Chen, X.; Kang, R.; Kroemer, G. Ferroptosis: Molecular Mechanisms and Health Implications. Cell Res. 2021, 31, 107–125. [Google Scholar] [CrossRef]
- Zhou, H.; Huang, J.; Liu, Y.; Wang, J.; Wang, J. The Emerging Role of Ferroptosis in the Pathological Development and Progression of Sepsis. Mil. Med. Res. 2025, 12, 81. [Google Scholar] [CrossRef]
- Choi, J.H.; Kagan, J.C. Oxidized Phospholipid Damage Signals as Modulators of Immunity. Open Biol. 2025, 15, 240391. [Google Scholar] [CrossRef]
- Kadl, A.; Sharma, P.R.; Chen, W.; Agrawal, R.; Meher, A.K.; Rudraiah, S.; Grubbs, N.; Sharma, R.; Leitinger, N. Oxidized Phospholipid-Induced Inflammation is Mediated by Toll-Like Receptor 2. Free Radic. Biol. Med. 2011, 51, 1903–1909. [Google Scholar]
- Eskander, G.; Abdelhamid, S.G.; Wahdan, S.A.; Radwan, S.M. Insights on the Crosstalk among Different Cell Death Mechanisms. Cell Death Discov. 2025, 11, 56. [Google Scholar] [CrossRef]
- Song, X.; Hao, X.; Zhu, B.T. Role of Mitochondrial Reactive Oxygen Species in Chemically-Induced Ferroptosis. Free Radic. Biol. Med. 2024, 223, 473–492. [Google Scholar] [CrossRef]
- Kayagaki, N.; Webster, J.D.; Newton, K. Control of Cell Death in Health and Disease. Annu. Rev. Pathol. Mech. Dis. 2024, 19, 157–180. [Google Scholar] [CrossRef] [PubMed]
- Fritsch, M.; Gunther, S.D.; Schwarzer, R.; Albert, M.; Schorn, F.; Werthenbach, J.P.; Schiffmann, L.M.; Stair, N.; Stocks, H.; Seeger, J.M.; et al. Caspase-8 is the Molecular Switch for Apoptosis, Necroptosis and Pyroptosis. Nature 2019, 575, 683–687. [Google Scholar] [CrossRef]
- McGuire, P.J. Mitochondrial Dysfunction and the Aging Immune System. Biology 2019, 8, 26. [Google Scholar] [CrossRef] [PubMed]
- Victorelli, S.; Salmonowicz, H.; Chapman, J.; Martini, H.; Vizioli, M.G.; Riley, J.S.; Cloix, C.; Hall-Younger, E.; Machado Espindola-Netto, J.; Jurk, D.; et al. Apoptotic Stress Causes mtDNA Release during Senescence and Drives the SASP. Nature 2023, 622, 627–636. [Google Scholar] [CrossRef] [PubMed]
- Miwa, S.; Kashyap, S.; Chini, E.; von Zglinicki, T. Mitochondrial Dysfunction in Cell Senescence and Aging. J. Clin. Investig. 2022, 132, e158447. [Google Scholar] [CrossRef]
- Li, H.; Guan, J.; Chen, J.; Sun, W.; Chen, H.; Wen, Y.; Chen, Q.; Xie, S.; Zhang, X.; Tao, A.; et al. Necroptosis Signaling and NLRP3 Inflammasome Cross-Talking in Epithelium Facilitate Pseudomonas Aeruginosa Mediated Lung Injury. Biochim. Biophys. Acta Mol. Basis Dis. 2023, 1869, 166613. [Google Scholar] [CrossRef]
- Huang, Y.; Xu, W.; Zhou, R. NLRP3 Inflammasome Activation and Cell Death. Cell. Mol. Immunol. 2021, 18, 2114–2127. [Google Scholar] [CrossRef]
- Kang, T.B.; Yang, S.H.; Toth, B.; Kovalenko, A.; Wallach, D. Caspase-8 Blocks Kinase RIPK3-Mediated Activation of the NLRP3 Inflammasome. Immunity 2013, 38, 27–40. [Google Scholar] [CrossRef]
- Doglio, M.G.; Verboom, L.; Ruilova Sosoranga, E.; Frising, U.C.; Asaoka, T.; Gansemans, Y.; Van Nieuwerburgh, F.; van Loo, G.; Wullaert, A. Myeloid OTULIN Deficiency Couples RIPK3-Dependent Cell Death to Nlrp3 Inflammasome Activation and IL-1beta Secretion. Sci. Immunol. 2023, 8, eadf4404. [Google Scholar] [CrossRef]
- Lei, X.; Chen, Y.; Lien, E.; Fitzgerald, K.A. MLKL-Driven Inflammasome Activation and Caspase-8 Mediate Inflammatory Cell Death in Influenza A Virus Infection. mBio 2023, 14, e0011023–23. [Google Scholar] [CrossRef]
- Li, J.; Li, L.; Zhang, Z.; Chen, P.; Shu, H.; Yang, C.; Chu, Y.; Liu, J. Ferroptosis: An Important Player in the Inflammatory Response in Diabetic Nephropathy. Front. Immunol. 2023, 14, 1294317. [Google Scholar] [CrossRef] [PubMed]
- Anderson, A.J.; Su, J.H.; Cotman, C.W. DNA Damage and Apoptosis in Alzheimer’s Disease: Colocalization with C-Jun Immunoreactivity, Relationship to Brain Area, and Effect of Postmortem Delay. J. Neurosci. 1996, 16, 1710–1719. [Google Scholar] [CrossRef]
- Jakaria, M.; Belaidi, A.A.; Bush, A.I.; Ayton, S. Ferroptosis as a Mechanism of Neurodegeneration in Alzheimer’s Disease. J. Neurochem. 2021, 159, 804–825. [Google Scholar] [CrossRef] [PubMed]
- Carrazana, E.; Salvadores, N. Therapeutic Implications of Necroptosis Activation in Alzheimer s Disease. Alzheimers Res. Ther. 2024, 16, 275. [Google Scholar] [CrossRef]
- Xu, C.; Wu, J.; Wu, Y.; Ren, Z.; Yao, Y.; Chen, G.; Fang, E.F.; Noh, J.H.; Liu, Y.U.; Wei, L.; et al. TNF-Alpha-Dependent Neuronal Necroptosis Regulated in Alzheimer’s Disease by Coordination of RIPK1-p62 Complex with Autophagic UVRAG. Theranostics 2021, 11, 9452–9469. [Google Scholar] [CrossRef]
- Shah, N.; Natesan, G.; Gupta, R. Uncovering Necroptosis in Alzheimer’s Disease: A Systematic Review of Evidence Across Experimental Models. Cell. Mol. Neurobiol. 2025, 45, 83. [Google Scholar] [CrossRef] [PubMed]
- Bao, W.; Pang, P.; Zhou, X.; Hu, F.; Xiong, W.; Chen, K.; Wang, J.; Wang, F.; Xie, D.; Hu, Y.; et al. Loss of Ferroportin Induces Memory Impairment by Promoting Ferroptosis in Alzheimer’s Disease. Cell Death Differ. 2021, 28, 1548–1562. [Google Scholar] [CrossRef]
- Vince, J.E.; Wong, W.W.; Gentle, I.; Lawlor, K.E.; Allam, R.; O’Reilly, L.; Mason, K.; Gross, O.; Ma, S.; Guarda, G.; et al. Inhibitor of Apoptosis Proteins Limit RIP3 Kinase-Dependent Interleukin-1 Activation. Immunity 2012, 36, 215–227. [Google Scholar] [CrossRef]
- Hambright, W.S.; Fonseca, R.S.; Chen, L.; Na, R.; Ran, Q. Ablation of Ferroptosis Regulator Glutathione Peroxidase 4 in Forebrain Neurons Promotes Cognitive Impairment and Neurodegeneration. Redox Biol. 2017, 12, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Hu, Z.; Li, Z.; Lin, Y. Crosstalk among Mitophagy, Pyroptosis, Ferroptosis, and Necroptosis in Central Nervous System Injuries. Neural Regen. Res. 2024, 19, 1660–1670. [Google Scholar] [CrossRef] [PubMed]
- Markesbery, W.R.; Carney, J.M. Oxidative Alterations in Alzheimer’s Disease. Brain Pathol. 1999, 9, 133–146. [Google Scholar] [CrossRef]
- Butterfield, D.A.; Lauderback, C.M. Lipid Peroxidation and Protein Oxidation in Alzheimer’s Disease Brain: Potential Causes and Consequences Involving Amyloid Beta-Peptide-Associated Free Radical Oxidative Stress. Free Radic. Biol. Med. 2002, 32, 1050–1060. [Google Scholar] [CrossRef]
- Mannion, J.; Gifford, V.; Bellenie, B.; Fernando, W.; Ramos Garcia, L.; Wilson, R.; John, S.W.; Udainiya, S.; Patin, E.C.; Tiu, C.; et al. A RIPK1-Specific PROTAC Degrader Achieves Potent Antitumor Activity by Enhancing Immunogenic Cell Death. Immunity 2024, 57, 1514–1532.e15. [Google Scholar] [CrossRef]
- Cao, L.; Mu, W. Necrostatin-1 and Necroptosis Inhibition: Pathophysiology and Therapeutic Implications. Pharmacol. Res. 2021, 163, 105297. [Google Scholar] [CrossRef] [PubMed]
- Degterev, A.; Ofengeim, D.; Yuan, J. Targeting RIPK1 for the Treatment of Human Diseases. Proc. Natl. Acad. Sci. USA 2019, 116, 9714–9722. [Google Scholar] [CrossRef]
- Zelic, M.; Pontarelli, F.; Woodworth, L.; Zhu, C.; Mahan, A.; Ren, Y.; LaMorte, M.; Gruber, R.; Keane, A.; Loring, P.; et al. RIPK1 Activation Mediates Neuroinflammation and Disease Progression in Multiple Sclerosis. Cell Rep. 2021, 35, 109112. [Google Scholar] [CrossRef]
- Bohgaki, T.; Mozo, J.; Salmena, L.; Matysiak-Zablocki, E.; Bohgaki, M.; Sanchez, O.; Strasser, A.; Hakem, A.; Hakem, R. Caspase-8 Inactivation in T Cells Increases Necroptosis and Suppresses Autoimmunity in Bim-/- Mice. J. Cell Biol. 2011, 195, 277–291. [Google Scholar] [CrossRef]
- Marin-Aguilar, F.; Castejon-Vega, B.; Alcocer-Gomez, E.; Lendines-Cordero, D.; Cooper, M.A.; de la Cruz, P.; Andujar-Pulido, E.; Perez-Alegre, M.; Muntane, J.; Perez-Pulido, A.J.; et al. NLRP3 Inflammasome Inhibition by MCC950 in Aged Mice Improves Health Via Enhanced Autophagy and PPARalpha Activity. J. Gerontol. Ser. A 2020, 75, 1457–1464. [Google Scholar] [CrossRef]
- Camell, C.D.; Gunther, P.; Lee, A.; Goldberg, E.L.; Spadaro, O.; Youm, Y.; Bartke, A.; Hubbard, G.B.; Ikeno, Y.; Ruddle, N.H.; et al. Aging Induces an Nlrp3 Inflammasome-Dependent Expansion of Adipose B Cells that Impairs Metabolic Homeostasis. Cell Metab. 2019, 30, 1024–1039.e6. [Google Scholar] [CrossRef]
- Youm, Y.H.; Nguyen, K.Y.; Grant, R.W.; Goldberg, E.L.; Bodogai, M.; Kim, D.; D’Agostino, D.; Planavsky, N.; Lupfer, C.; Kanneganti, T.D.; et al. The Ketone Metabolite Beta-Hydroxybutyrate Blocks NLRP3 Inflammasome-Mediated Inflammatory Disease. Nat. Med. 2015, 21, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Lin, H.; Li, S.; Huang, Y.; Sun, Y.; Shu, S.; Luo, T.; Liang, T.; Lai, W.; Rao, J.; et al. Macrophage Iron Dyshomeostasis Promotes Aging-Related Renal Fibrosis. Aging Cell 2024, 23, e14275. [Google Scholar] [CrossRef] [PubMed]
- Xiang, W.; Zhang, T.; Li, B.; Li, S.; Zhang, B.; Fang, S.; Chen, L.; Gong, Y.; Huang, B.; Feng, D.; et al. Senescent Macrophages Induce Ferroptosis in Skeletal Muscle and Accelerate Osteoarthritis-Related Muscle Atrophy. Nat. Aging 2025, 5, 1295–1316. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.