Abstract
Singlet oxygen (1O2), the excitation stage of the ground-state molecular oxygen, is a fundamental reactive oxygen species (ROS) with important functions in plant growth, development, and stress responses. In plant cells, 1O2 is mainly generated in the chloroplast due to photosensitizing activity of tetrapyrroles. Moreover, 1O2 can be generated in non-photosynthetic tissues when plants suffer environmental stresses. Although 1O2 was initially considered as a cytotoxin—causing merely photooxidative damages, more recent work suggests that 1O2 also acts as a signal that either triggers a programmed cell death response or promotes acclimation. The 1O2 signaling pathway is distinct and operates independently of other ROS signaling cascades. In Arabidopsis, EXECUTER1 (EX1) protein has been identified as a crucial signaling component that perceives and relays 1O2 signals to the nucleus, thereby initiating extensive transcriptional reprogramming. Additionally, oxidative products of carotenoids, such as β-cyclocitral, are also recognized as 1O2-derived signaling molecules. Through specific chloroplast-to-nucleus signaling and cross talk with hormone signaling networks—including jasmonic acid (JA) and salicylic acid (SA)—1O2 helps finely coordinate plant growth, defense responses, and cell fate decisions under fluctuating environmental conditions. This review aims to summarize current knowledge on 1O2 generation and signaling, 1O2-induced chloroplast changes under diverse stress conditions, and cross talk between 1O2 and phytohormone signaling.
1. Introduction
Plants performing oxygenic photosynthesis inevitably generate a certain amount of ROS. While a basal level of ROS is necessary for various metabolic processes, a sudden increase in ROS is detrimental to plants [1]. Under stress conditions such as drought, high temperature, and high light that interfere with photosynthetic energy or electron transportation, a large amount of ROS including hydrogen peroxide (H2O2), singlet oxygen (1O2), superoxide radical (), and hydroxyl radical (OH•) are generated in the chloroplast [2,3]. Although many kinds of ROS are generated simultaneously, 1O2 has been proved the most detrimental to the chloroplast as more than 80% of photooxidative damages are caused by it directly [4]. Singlet oxygen is the excitation stage of the ground-state triplet molecular oxygen [5,6,7]. Energy transfer from an excited photosensitizer (e.g., 3Chl) could reverse the spin direction of one of the two outermost valence electrons of the triplet-state oxygen that occupy separate orbitals with parallel spins, allowing the pairing of the two electrons and formation of 1O2 [7]. Singlet oxygen could rapidly oxidize proteins, lipids, carbohydrates, and nucleic acids, and in this way it may irreversibly inactivate/destroy the targets [8,9,10]. Thus, for a long time, 1O2 had been considered as a cytotoxin because of its extreme high reactivity and short lifetime—about 1 µs in living cells [11,12,13]. However, an increasing body of recent studies clearly demonstrate that 1O2, in most cases, functions as a versatile signal that induces various stress responses via activating sequential signaling transduction cascades [14,15,16,17,18]. In the review, we summarize current understanding of 1O2 generation, perception, and signaling and discuss the role of 1O2 in plant environmental resilience.
2. Generation of Singlet Oxygen
In plant cells, 1O2 is mainly generated in the chloroplast by transferring the excitation energy to the ground-state triplet molecular oxygen (3O2) from a photosensitizer either metabolically or photochemically. Within the chloroplast, 1O2 is mainly generated during photosynthesis and from intermediate products of chlorophyll biosynthesis and degradation [19].
2.1. Singlet Oxygen Generation During Photosynthesis
In the light-harvesting antenna complex (LHC), chlorophyll (Chl) is excited from the ground state to its singlet excited state (1Chl*) after absorbing a photon, and the fate of its excitation energy is different [20,21]. At normal growth conditions, most (80~85%) of the excitation energy is gradually transferred to the reaction center of PSII to drive the photosynthetic electron transfer chain, and the rest of the light energy will be dissipated in the form of heat or fluorescence, a process known as non-photochemical quenching (NPQ) [22,23]. 1Chl * is a rather short-lived (about 10−8 s in living cells) molecule [20]. If the excitation energy in 1Chl * is not immediately used, 1Chl * will release a small portion of energy, decaying to its longer-lifespan (about 10−3 s) triplet excitation state, 3Chl*, via charge recombination [24]. 3Chl * is a strong photosensitizer that can transfer its excitation energy to the ground-state oxygen, leading to the generation of 1O2 [25,26,27].
Though 3Chl * is produced, generation of 1O2 in LHC is rather limited. In the antenna, chlorophylls are surrounded by carotenoids, such as lutein and zeaxanthin, and the generated 3Chl * is mostly (about 95%) quenched by the carotenoid molecules presented nearby. Thus, only a trace amount of 1O2 is produced in LHC, even for plants that are grown under high light [28,29,30,31]. The Arabidopsis ch1 mutant is devoid of chlorophyll b (Chl b) due to the inactivation of chlorophyll a oxygenase (CAO) that catalyzes the conversion of Chl a to Chl b [17,32]. Since Chl b is an essential component of the light-harvesting complex associated with PSII, the absence of Chl b disrupts the formation and stability of the PSII antenna complex, which in turn impairs excitation energy transfer within LHC and leads to enhanced 1O2 production under high light [33,34,35].
In the PSII reaction center (RC), it is a rather different scenario as a large amount of 1O2 is produced when plants suffer from high light [36,37]. The PSII RC contains six chlorophyll a (Chl a) molecules, two pheophytin (Pheo), two β-carotenes (Car), and two quinones arranged along the D1 and D2 branches [38]. Among the six Chl a molecules, two of the four Chl a molecules located in the center are specialized and paired as primary electron donor, P680 [39]. Once it absorbs light energy, P680 is excited to its singlet excitation state, 1P680 *, releasing an energetic electron that is transferred to Pheo and quinone A (QA) sequentially [40,41]. Under stress conditions, it frequently happens that the electron acceptor QA remains reduced due to the blocking of forward electron transfer (also called the closed state of the reaction center), making it unable to accept electrons [2,42]. In such a condition, the excited 1P680 * could decay to the long-lived triplet excited state 3P680 * via the formation of two intermediates, 1P680+Pheo− and 3P680+Pheo− [2,20]. Unlike LHC, P680 in PSII RC is not surrounded by carotenoids. The two β-carotene molecules in PSII RC are too far way to quench 3P680 * directly, that eventually leads to the production of 1O2 [25,27,43,44,45].
Plastoquinone (PQ) acts as an electron carrier between PSII and Photosystem I (PSI) during photosynthesis, existing in interconvertible oxidized and reduced forms [46]. Excitation of PSII enhances the electron transport rate, leading to the reduction in the PQ pool. In addition to being an electron carrier, PQ also acts as a scavenger of 1O2 [47,48]. Two plastoglobuli-localized kinases, ABC1K1 and ABC1K3, play a crucial role in maintaining the redox balance of the PQ pool. In the abc1k1 mutant, the diminished PQ pool compromises photosynthetic electron flow due to insufficient electron acceptors in PSII, and may also impair its capacity to scavenge 1O2. The abc1k1 mutant exhibits distinct photosynthetic and metabolic phenotypes—the contents of PQ, β-carotene, and xanthophyll lutein are reduced, and the metabolic process of the membrane antioxidant tocopherol is disrupted [49]. Consequently, the reduction in the PQ pool of the abc1k1 mutant results in excessive accumulation of 1O2 and severe cell death phenotypes when grown under red or white light [46].
2.2. Singlet Oxygen Generation from Chlorophyll Biosynthesis Intermediates
Another important source of 1O2 in the chloroplast is the biosynthesis intermediates of tetrapyrroles. In higher plants, tetrapyrroles are mainly divided into three categories, chlorophyll, heme, and phycobilin (Figure 1). Tetrapyrrole biosynthesis originates from glutamic acid and shares a common biosynthetic pathway until the formation of protoporphyrin IX (Proto IX) [50,51]. Afterwards, the pathway diverges into two branches, with the Mg branch leading the formation of chlorophylls and the Fe branch the formation of hemes and phycobilins [52,53]. The first step of the Mg branch is catalyzed by magnesium chelatase (MgCh) to insert an Mg2+ ion into Proto IX, forming Mg-Proto IX. Mg-Proto IX was then catalyzed by Mg-Proto IX methyltransferase (MgMT/CHLM) and Mg-Proto IX monomethylester cyclase (MgCY/CHL27) continuously to form protochlorophyllide a (Pchlide) in the dark [54,55]. The Mg branch stops when the Pchlide amount reaches a threshold level and restarts when Pchlide is reduced to chlorophyllide a (Chlide) by the Pchlide oxidoreductase (POR) under light. The resulting divinyl Chlide a is esterified with a long chain polyisoprenyl (geranylgeraniol or phytol) to synthesize Chl a [56]. The Fe branch is firstly catalyzed by ferrochelatase (FeCh) to insert Fe2+ into Proto IX to form heme b which is then converted to other hemes or eventually to phytochromobilins or phycobilins [53].
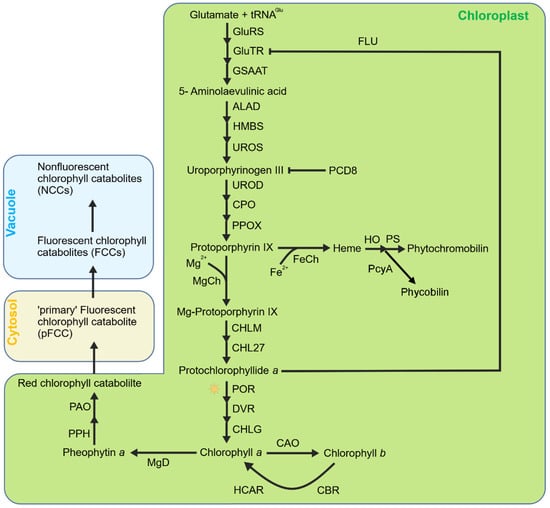
Figure 1.
Simplified model of tetrapyrrole metabolism in chloroplasts. Tetrapyrrole biosynthesis originates from glutamic acid and shares a common biosynthetic pathway until the formation of protoporphyrin IX (Proto IX). Then, this pathway diverges to the Mg branch (leading to the formation of Chls) and Fe branch (leading to the formation of heme and the subsequent phytochromobilin or phycobilin). Mg–protoporphyrin is subsequently converted to chlorophyll a, which can be further converted to chlorophyll b by CAO. Chlorophyll degradation begins with the conversion of chlorophyll a to pheophytin a by releasing the Mg2+ ion via MgD (Mg–dechelatase), followed by cleavage by PPH and PAO to produce red chlorophyll catabolite (RCC) and subsequently fluorescent chlorophyll catabolites (FCCs), which are finally converted to non-fluorescent chlorophyll catabolites (NCCs). Heme is the precursor of phytochromoblins and phycobilins.
Free chlorophylls, hemes, and phycobilins are strong photosensitizers due to their structural and biophysical properties [57]. In plant cells, these tetrapyrroles are usually bound with proteins or surrounded by carotenoids to dissipate excess absorbed light energy using various quenching mechanisms. However, the biosynthetic intermediates of tetrapyrroles mostly occurred in free form and are thus more detrimental when illuminated. To prevent overaccumulation, plants have evolved sophisticated strategies to exert strict regulations on tetrapyrrole biosynthesis and degradation, and the most important regulation is the negative feedback inhibition of the initial step of tetrapyrrole biosynthesis, i.e., the formation of δ-aminolevulinic acid (ALA) by two effector molecules, heme and the FLU protein, that both interact with glutamyl-tRNA reductase (Glu-TR), the first enzyme of tetrapyrrole biosynthesis [58,59]. Heme has been implicated in inhibiting mainly the Fe branch [60]. Increased levels of heme bind to the N-terminal part of Glu-TR that blocks Glu-TR activity and inhibits ALA synthesis [60,61,62]. FLU seems to selectively affect the Mg branch of tetrapyrrole biosynthesis [57]. In the dark, Chl biosynthesis ends with the formation of Pchlide. When Pchlide level reaches a critical level, the synthesis of ALA turns down through a yet unknown mechanism that relies on the FLU protein [61,62,63]. The Arabidopsis flu mutant, lacking this negative feedback regulation mechanism, is unable to restrain Pchlide accumulation in the dark and thus accumulates an excessive amount of Pchlide [57,61,64]. Once exposed to light, excessive Pchlide transfers light energy to the ground molecular oxygen to produce a large amount of 1O2, which leads to plant growth inhibition and even cell death [65,66,67]. However, in the flu mutant, the heme content is similar to that of wild-type plants, indicating that FLU acts independently of heme [57].
Two ferrochelatases (FeCh), FC1 and FC2, catalyze the formation of heme by inserting Fe2+ into protoporphyrin IX [68]. In fc1 knockout mutants, the level of heme is only slightly decreased in shoot and root, as well as the contents of chlorophylls and carotenoids, and the efficiency of PSII is basically unaffected, indicating that FC1 does not play a major role in tetrapyrrole biosynthesis [69]. However, the fc2 mutant forms abnormally small, pale green rosette leaves, and is low in chlorophylls and carotenoids, underscoring its major role in tetrapyrrole metabolism [68]. The fc2 mutant produces an overdose amount of 1O2 when grown under dark/light cycles that leads to cell death of young seedlings and growth inhibition of mature plants [70,71,72]. However, the underlying reason is still under debate. Woodson et al. reported that the fc2 mutants accumulate Proto IX after dawn [18], but Scharfenberg et al. pointed that this mutant does not accumulate Proto IX but Pchlide instead in the dark [68].
Homeostasis of tetrapyrrole is important for plants’ developmental programs and plants’ response to environmental cues. Impaired tetrapyrrole biosynthesis or catabolism leads to the generation of 1O2 and the following cell death responses. PROGRAMMED CELL DEATH 8 (PCD8) is a newly identified thylakoid-localized protein that controls proteolysis of tetrapyrrole biosynthesis proteins by directing Clp protease to these enzymes [73]. Knockdown of the PCD8 gene leads to the accumulation of ALA, uroporphyrinogen III (Uro III), Proto IX, Mg–protoporphyrin IX (MgP), and Pchlide, resulting in severe chloroplast damage and a necrotic phenotype under short-day conditions. It is proposed that the burst of 1O2, triggered by excessive accumulation of tetrapyrrole intermediates—particularly uroporphyrinogen III under light conditions—is responsible for the observed cell death in these knockdown mutants [73].
2.3. Singlet Oxygen Generation from Chlorophyll Breakdown Products
Another important source of 1O2 comes from the breakdown process of Chls, especially during senescence. In plant tissues, Chls are usually bound with proteins and carotenoids, and, in this way, the excitation energy absorbed by Chls are readily used for photosynthesis or dissipated as heat or fluorescence [74]. During senescence, Chls are frequently disassociated from these Chl-binding proteins, and the removal of free Chls and its breakdown intermediates is crucial since these molecules readily generate 1O2 and are potentially toxic to plants [12,75]. Within plastids, a series of enzymes catalyze the conversion of Chl to linear colorless tetrapyrrole derivatives referred to as ‘primary’ fluorescence Chl catabolites (pFCC). pFCC molecules are then exported to the vacuole and finally breakdown to monopyrrole molecules which might be further metabolized to smaller molecules [76,77,78]. Among the Chl breakdown process, the opening of the tetrapyrrole ring that forms the red chlorophyll catabolite (RCC) is recognized as a crucial step, and this step is catalyzed by phenylalanine oxygenase (PAO) [79]. The Arabidopsis acd1 mutant and maize lls1 mutant, lacking functional PAO protein, accumulate pheophorbide a in the plastids and generate 1O2 when illuminated [79]. Then RCC is reduced to a ‘primary’ fluorescent chlorophyll catabolite (pFCC) via RCC reductase (RCCR) [80]. The Arabidopsis acd2 mutant, bearing mutation in the RCCR gene, is defective in converting RCC to pFCC, and thus accumulates RCCs and RCC-like pigments in the dark, favoring 1O2 production when transferred to the light and causing cell death [71,81,82].
2.4. Singlet Oxygen Generation During Seedling De-Etiolation
When seeds are geminated and grown in complete darkness, etioplasts are developed in plant tissues that would have chloroplasts if subjected to light. Etioplasts do not contain chlorophyll or stacked thylakoid membranes, but rather have a paracrystalline lipid–pigment–protein structure known as the prolamellar body (PLB) [83]. The PLB consists largely of plastid lipids (mainly MGDG and DGDG), the chlorophyll precursor Pchlide, the light-dependent POR, and its cofactor NADPH [84]. However, if the darkness continues, the level of Pchlide will increase and lead to 1O2 production after illumination [85]. In etiolated seedlings, the expression of genes required for carotenoid (e.g., AtPSY) and chlorophyll biosynthesis (e.g., AtPOR) is repressed by the PIF transcription factors [86,87]. Deficiency in either PIF1 or PIF3 leads to elevated levels of Pchlide in darkness and results in photobleaching upon transfer to light [86,88]. Upon light exposure, the etiolated pif5 seedlings also develop a photobleaching phenotype [89]. The photobleaching phenotype observed in seedlings during the dark-to-light transition is largely attributed to ROS [90]. Therefore, researchers observed that, following 24 h of light treatment, 4-day-old dark-grown seedlings of the pif1 pif3 double mutant and the pifq (quadruple) mutant exhibited increased fluorescence from H2DCFDA—a dye sensitive to reactive oxygen species—as well as detectable 1O2 production, as measured by the singlet oxygen sensor green (SOSG) fluorescent probe. These findings indicate that PIFs play a significant role in suppressing 1O2 generation during seedling de-etiolation [91,92].
2.5. Singlet Oxygen Generation in Non-Photosynthetic Conditions
Recent studies indicate that 1O2 can be generated through multiple stress pathways and may originate from organelles other than chloroplasts in a light-independent manner [93,94,95]. Transcriptional analysis of leaves subjected to various stresses, including drought, reveals 1O2-induced responses under both dark and light conditions [96]. Researchers observed that the transcriptomic profiles under various stress conditions—whether induced by light or under dark treatments—closely resembled the transcriptome of the flu mutant in Arabidopsis [94]. When Arabidopsis roots are subjected to diverse biotic and abiotic stresses in darkness, 1O2 is rapidly produced and accumulated in mitochondria, peroxisomes, and nuclei. These findings demonstrate that 1O2 can be produced via multiple stress pathways and may originate from non-chloroplastic organelles without the need for light [94].
Under drought stress, plants exhibit a high osmotic potential. A research team reported that the effect of polyethylene glycol on root growth is strongly correlated with the generation of 1O2. Once 1O2 is produced, cell death occurs [97]. Similarly, simulating drought stress through leaf dehydration treatment can rapidly induce 1O2 production. This result was quantified by measuring the amounts of free radicals captured with the 1O2-specific spin-trapping agent 4-hydroxy-tetramethylpiperidine [98]. Another study also showed that 1O2 generates in the wounded Arabidopsis leaves. In the Arabidopsis lox2 mutant lacking the functional chloroplast lipoxygenase, the production of 1O2 was suppressed. This effect is due to the absence of lipoxygenase-initiated lipid peroxidation, indicating that chloroplast-localized lipoxygenase plays a role in 1O2 generation [93]. Moreover, in the absence of light, salicylic acid can increase the level of 1O2 through lipid peroxidation [95]. A summary for 1O2 generation in higher plants is shown in Table 1.

Table 1.
A summary of singlet oxygen generation in plant.
3. Singlet Oxygen-Induced Signaling
Singlet oxygen has long been regarded as a cytotoxin because of its short lifetime (0.5–1 µs in biological tissues) and high reactivity [11]. However, more evidence shows that 1O2 could also act as a multifunctional signal molecule, triggering various stress responses via activating signal transduction cascades [14,17]. The signaling role of 1O2 was first reported in C. reinhardtii in 2001. Leisinger et al. reported that transcripts of C. reinhardtii Glutathione peroxidase homologous (Gpxh), a thioredoxin-dependent peroxidase catalyzing the reduction of hydrogen peroxide and organic hydroperoxides, was specifically and rapidly induced by 1O2 produced by photosensitizers including neutral red, methylene blue, and rose bengal [99]. However, most important breakthroughs on 1O2 signaling came from studies of the Arabidopsis conditional mutants, including but not limited to the ch1, flu, and fc2 mutants.
3.1. Singlet Oxygen Signaling from Grana Core
Based on the sites of 1O2 generation, 1O2-induced signaling in the chloroplast can originate from the grana core (GC, the inner region of the grana stacks) or grana margin (GM, the marginal region of the grana stacks). The ch1 mutant produces 1O2 specifically in GC and is thus widely used as a tool to explore 1O2 signaling originating from GC. The D1 protein in GC could scavenge 1O2 at the expense of its own oxidation, but there is no evidence to support that the degradation products of D1 act as signal molecules in plants [24]. Using the Arabidopsis ch1 mutant, Ramel et al. found that 1O2 produced in GC could oxidize carotenoids, like β-carotene, and produce four volatile derivatives including β-cyclocitral (β-CC), β-ionone (β-I), dihydroactinidiolide (dhA), and α-ionene [100]. Among the four chemicals, β-CC was found to induce the expression of many 1O2-responsive genes (SORGs), like genes induced by 1O2 in the flu mutant, but had no obvious effect on the expression of H2O2-responsive genes. Thus, β-CC is considered as a second messenger of 1O2 in the chloroplast. β-CC-induced gene expression changes were associated with an increased tolerance to photooxidative stress—an external application of β-CC enhances plants’ tolerance to high light stress. In addition, a small number of genes induced by β-CC are also induced by dhA, indicating that β-CC is not the only β-carotenoid-derived messenger involved in the 1O2 signaling pathway, even though dhA may not be as important as β-CC in mediating chloroplast-to-nucleus retrograde signaling [20,101].
MBS1 is a small zinc finger protein, which was first discovered in the genetic screening of C. reinhardtii mutants for early components of 1O2 signaling. MBS1 is essential for the induction of SORGs. Upon oxidative stress, it accumulates in distinct granules and the cytosol [102]. Function loss of MBS1 leads to deregulation of 1O2-induced genes after β-CC treatment, indicating that MBS1 is a new downstream intermediate in β-CC signaling pathway that ultimately regulates the photoprotection process of Arabidopsis [103].
In Arabidopsis, a remarkable feature of β-CC-induced transcriptional changes is that it induces various detoxification-related genes, including several GSTs and UDP–glycosyltransferases [100]. The transcriptional cofactor SCARECROW LIKE 14 (SCL14) was proven to play a key role in the retrograde signal transduction triggered by β-CC under photooxidative stress. β-CC binds to SCL14, relocating it to the nucleus and regulating the expression of NAC transcription factors, enhancing the expression of ANAC002, ANAC032, and ANAC081. These transcription factors regulate detoxification-related gene expression, enhancing plant defense [103,104,105]. Meanwhile, β-CC can down-regulate the methyl erythritol-4-phosphate (MEP) pathway in terpenoid biosynthesis, which provides precursors for many important metabolites in photosynthesis, such as chlorophyll, carotenoids, plastoquinone, phylloquinone, and tocopherol, slowing down 1O2 generation from these tetrapyrroles and providing another layer of protection for plants under strong high light stress [106].
3.2. Singlet Oxygen Signaling from the Grana Margin
Unlike that in the GC, 1O2 generated in GM activates distinct signaling transduction pathways, based mostly on the study of the conditional mutants that generate 1O2 in GM, i.e., the Arabidopsis flu and fc2 mutant. In the flu mutant, excessive photosensitizer Pchlide is accumulated in the dark, allowing 1O2 generation when plants are transferred to light. Thus, the flu mutant provides an excellent system that can produce 1O2 in a controlled and non-invasive manner [14]. Furthermore, when grown under continuous light, i.e., without Pchlide accumulation and 1O2 generation, the flu mutant shows no visible growth defects, like the wild-type Col-0 plants. After the 1O2 burst, the mature flu plants stop growing immediately, while the seedlings bleach and die. However, all these drastic phenotypic changes are suppressed if EXECUTER1 (EX1) gene is inactivated, even though flu ex1 mutant generates a similar amount of 1O2 to its parental flu plant upon a dark-to-light shift. In the flu mutant, enhanced levels of 1O2 generated in the thylakoids oxidize the Trp643 residue of the EX1 protein, and the subsequent cleavage of the oxidized EX1 protein is necessary for induction of this signaling transduction pathway. The FtsH2 (also called VAR2) protease, localized in the thylakoid of the chloroplast, has been reported in the repair of damaged PSII and turnover of the EX1 protein. In flu var2, not only 1O2-induced cellular responses, but also 1O2-induced degradation of the EX1 protein is suppressed, underlying that proteolysis of EX1 is crucial for initiating 1O2-induced and EX1-mediated retrograde signaling [19,107]. Based on genome-wide transcriptome analysis, inactivation of FtsH2 inhibited most (85%) EX1-dependent 1O2 response genes (SORGs), providing further evidence for the key role of FtsH2 in mediating 1O2-induced signaling. Although Pchlide is accumulated and 1O2 is generated non-selectively all over the thylakoid membrane, the 1O2 sensor EX1 protein is localized in the GM region, making GM an origin of 1O2 signaling. A combined study of the flu and flu-related mutants has proved that 1) 1O2 can also be a highly versatile signal, 2) EX1 is a sensor of 1O2 signal, and 3) FtsH2-dependent proteolysis of EX1 is necessary for mediating 1O2 signaling [19,58,108]. A paper published recently reported that the mature EX1 protein relocates from chloroplasts to the nucleus upon 1O2 release, where it interacts with WRKY (WRKY18 and WRKY40) transcription factors to regulate SORG expression [92]. However, this study is questioned by another study published in 2024 which showed that the mature EX1 protein does not accumulate in the nucleus and does not interact with nuclear WRKY18 or WRKY40 either [109]. Subsequently, a recent study reports that mutation of the TOC33 gene in the flu mutant can suppress largely 1O2-induced stress responses, even when 1O2 levels are comparable to those in the flu [110]. TOC33 interacts with the UVR domain of EX1 (EX1-UVR) in the chloroplast envelope, promoting an 1O2-dependent reduction of EX1-UVR in the chloroplasts and an increase in EX1-UVR in the nucleus. The UVR domain in the nucleus interacts with WRKY18 and WRKY40 transcription factors to modulate the expression of SORGs [110].
In addition to EX1, its homolog, EXECUTER2 (EX2), is also involved in mediating 1O2 signals in Arabidopsis. Function loss of the chloroplastic EX1 protein suppresses largely the up-regulation of SORGs, but does not eliminate these changes. However, inactivation of both EX1 and EX2 led to almost complete inhibition of SORGs [107,111]. Like EX1, EX2 undergoes oxidative post-translational modification at the Trp530 of the DUF domain. EX2 localizes in close vicinity to the EX1 protein in the GM region and functions as a negative regulator of the EX1 signalosome through its own 1O2-dependent oxidation [104,112].
In addition, EX1 and EX2 have been reported to play an important role in plant–pathogen interactions, plastid development, and seed dormancy. During the invasion process, the genus Alternaria and other phytopathogenic fungi secrete mycotoxin tenuazonic acid (TeA) that inhibits photosynthesis and causes burst of photosynthetic 1O2. The functional loss of EX1, or both EX1 and EX2, significantly compromises the expression of 1O2-responsive nuclear genes and foliar lesions caused by the Alternaria pathogens [113]. During the de-etiolation process, EX1 and EX2 perceive and mediate the 1O2 signal produced in etiolated seedlings, and they orchestrate the expression of photosynthetic gene expression [114]. In addition, EX1 and EX2 also play an important role in seed dormancy and gemination via orchestrating plastid formation [115].
Except for the EX1-dependent pathway, 1O2 generated on GMs also initiates an EX1-independent pathway, the 1O2-SAFE1 pathway. SAFE1 was identified in a screening for suppressor mutants in EMS-mutagenized flu ex1 seeds that restored a flu-like phenotype. SAFE1 localizes in the chloroplast stroma, and the release of 1O2 induces SAFE1 degradation via chloroplast-originated vesicles. Without SAFE1, GMs of chloroplast thylakoids are specifically damaged upon 1O2 generation [116]. Thus, SAFE1 is recognized as a negative regulator of an 1O2-induced and EX1-independent pathway, although the 1O2 sensor of this pathway is still unknown.
However, the 1O2-induced signaling pathway in the fc2 mutant does not rely on the EX1 protein [18]. The fc2 mutant is defective in plastid ferrochelatases 2 that catalyze the conversion of protoporphyrin IX (Proto IX) to heme in the chloroplast tetrapyrrole biosynthesis pathways [117]. When grown under diurnal light cycles, the fc2 mutant starts to accumulate Proto IX at dawn and peaks shortly. Proto IX is a strong photosensitizer, generating 1O2 that leads to chloroplast damage and eventually PCD [18]. However, all these severe phenotypes can be suppressed by mutation of the PUB4 gene without impairing Proto IX accumulation and the following 1O2 generation. The PUB4 gene encodes the Plant U-box 4 (PUB4) E3 ubiquitin ligase that ubiquitinates the outer envelope of the damaged chloroplast, facilitating its degradation in the vacuole [18,118]. Since in the fc2 pub4 double mutants, much less damaged chloroplasts are observed, PUB4 has been proposed to promote the formation or maturation of damaged chloroplasts. In contrast to flu, 1O2-induced signaling does not rely on the EX1 protein in the fc2 mutant, in which mutation of the EX1 gene cannot suppress the 1O2-induced stress phenotypes [18]. However, whether PUB4 is involved in 1O2-induced signaling in the flu mutant is still unclear. A representative scheme illustrating chloroplast 1O2 generation and signaling in these mutants is shown in Figure 2.
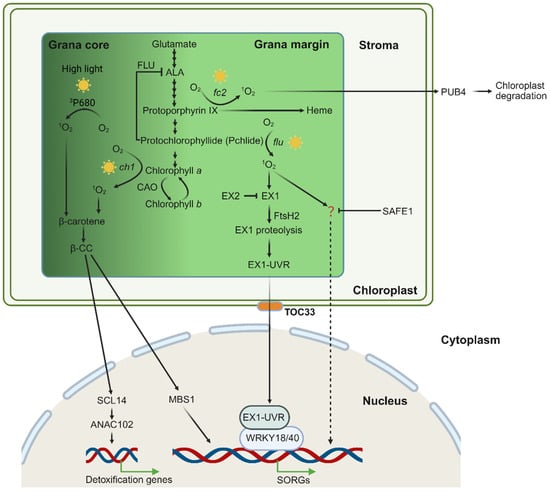
Figure 2.
Scheme of chloroplast 1O2 generation and signaling in the grana core and grana margin. Chloroplast 1O2 is primarily generated during the process of chlorophyll biosynthesis. Under high light, excess energy at PS II leads to the formation of 1O2 from excited chlorophyll (3P680 *). This 1O2 can be quenched by β-carotene, forming β-CC. In the ch1 mutant, there is 1O2 generation in the grana core (GC). Excessive 1O2 oxidizes β-carotene, yielding the volatile signaling molecule β-CC. This compound activates photoprotective gene expression via the zinc finger protein MBS1 and the transcription factor SCL14. The Arabidopsis mutant fc2 and flu, in which the chlorophyll biosynthesis is impaired, accumulates photosensitizing chlorophyll precursors, protoporphyrin IX and Pchlide, respectively. Upon light exposure, these compounds absorb light energy and generate highly reactive 1O2. The flu-produced 1O2 in the grana margin (GMs) initiates stress-related retrograde signaling (RS) pathways to regulate nuclear gene expression. The sensor protein EX1 detects elevated levels of 1O2 through oxidation at the W643 residue, triggering its hydrolysis by FtsH2. This process releases the UVR domain that is eventually transmitted via the TOC33-dependent mechanism to the nucleus, where it interacts with WRKY transcription factors to regulate the expression of SORGs. The EX2 protein localizes in close vicinity with EX1 and neutralizes 1O2 via its W530 residue oxidation. Additionally, 1O2 activates an EX1-independent signaling pathway which is suppressed by SAFE1, although the sensor is still uncovered. In the fc2 mutant, 1O2 causes chloroplast damage, leading to chloroplast outer envelope ubiquitination by the E3 ligase PUB4 and subsequent vacuolar degradation.
4. Cross Talk Between Phytohormones and 1O2-Induced Signaling
In addition to photosynthesis, chloroplasts are also major biosynthesis sites of phytohormones such as salicylic acid (SA), jasmonic acid (JA), abscisic acid (ABA), gibberellins, and ethylene [119]. An increased production of ROS in the chloroplast not only directly affects the photosynthetic efficiency, but also affects the biosynthesis and signal transduction of the above hormones [120,121,122].
4.1. Salicylic Acid and 1O2 Signaling
SA is a key phytohormone that regulates diverse signaling pathways, particularly plant immune responses [123]. In plants, SA is synthesized through two distinct pathways—the ICS pathway and the PAL pathway, both originating from chorismate in the chloroplast.
In the ICS pathway, chorismate is first isomerized to isochorismate by isochorismate synthase (ICS) enzymes. Isochorismate is then transported into the cytosol via the EDS5 transporter located on the chloroplast envelope. There, it is conjugated with L-glutamate by a cytosolic amidotransferase, forming isochorismate-9-glutamate (ICS-Glu), which spontaneously decomposes into SA and 2-hydroxy-acryloyl-N-glutamate. In the PAL pathway, chorismate is converted into phenylalanine within plastids. Phenylalanine is subsequently exported to the cytosol and deaminated to trans-cinnamic acid (t-CA) by phenylalanine ammonia-lyase (PAL). t-CA is further metabolized into SA via intermediates such as ortho-coumaric acid and benzaldehyde [124,125,126].
Under high light stress, β-CC generated in the PSII RC promotes SA biosynthesis by upregulating ICS1 expression in Arabidopsis [127]. This β-CC-induced SA accumulation, along with associated changes in nuclear gene expression, enhances plant tolerance to excessive light. During this acclimation process, EDS1 and NPR1 (Nonexpressor of Pathogenesis-Related Genes 1), a central component of SA signaling, positively regulate SA biosynthesis and signaling, respectively [127]. In the cytoplasm, NPR1 typically forms oligomers stabilized by disulfide bonds between cysteine residues in its monomers [128,129]. SA accumulation alters the cellular redox state, reducing these disulfide bonds and liberating NPR1 monomers, which then translocate into the nucleus. There, NPR1 interacts with TGA transcription factors to activate defense-related gene expression [128,130].
In the flu mutant, SA-responsive genes such as EDS1, PATHOGENESIS RELATED1 (PR1), and PR5 are strongly induced upon 1O2 release [131]. Inactivation of EDS1 in the flu mutant does not affect 1O2-induced oxylipin generation, growth inhibition, and the initiation of PCD, but it does allow plants to recover much faster from 1O2-induced growth inhibition and it also suppresses the spread of necrotic lesions in leaves [131]. It is proposed that 1O2 activates a complex stress-response program with EDS1 playing a key role in initiating and modulating several steps of it. This program includes not only responses to oxidative stress, but also responses known to be activated during plant–pathogen interactions and wounding [131]. Moreover, a chloroplast-localized protein, named the calcium-sensing receptor (CAS), has been shown to act upstream in 1O2-triggered retrograde signaling, influencing plant immunity and SA biosynthesis [132]. It was shown that pathogen-associated molecular pattern (PAMP) signals are quickly relayed to chloroplasts and evoke CAS-dependent transient Ca2+ signals in the chloroplast that are responsible for both the PAMP-induced basal resistance and R gene-mediated hypersensitive cell death. Transcriptome analysis demonstrates that CAS-mediated transcriptional reprogramming is achieved via chloroplast 1O2-mediated retrograde signaling [132].
Recent studies suggest that GUN1-mediated retrograde signaling also modulates the expression of defense-related genes associated with SA signaling [133,134]. GUN1 signaling transcriptionally represses GOLDEN2-LIKEs (GLKs), key regulators of chloroplast development [135]. GLKs are involved in SA signaling, and both SA and the SA-induced protein SIGMA FACTOR BINDING PROTEIN 1 (SIB1) can directly influence GLK activity [136]. Prolonged SA exposure negatively affects tetrapyrrole biosynthesis, impairing chloroplast development. Notably, this SA-induced defect in chloroplast biogenesis is more pronounced in gun1 mutants, implying that GUN1-mediated retrograde signaling may protect chloroplast development by suppressing GLK-driven defense responses linked to SA [133]. These findings point to a likely interconnection between chloroplast-derived reactive oxygen species (ROS) and SA-mediated signaling. Recently, a study reported that in dark conditions, salicylic acid-mediated lipid peroxidation leads to elevated levels of 1O2, which subsequently trigger EX1-dependent retrograde signaling. Loss of EX1 function partially alleviates the SA-induced hypocotyl growth inhibition in the npr1 mutant. This mechanism may involve the reactivation of SAUR gene expression at the transcriptional level in the npr1 genetic background [95]. Although the roles of chloroplast ROS and SA in plant stress responses are well established, the molecular and genetic interactions between these components, as well as the potential function of SA in retrograde signaling, remain incompletely understood. A simplified model for the relationship of 1O2 and JA is shown in Figure 3.
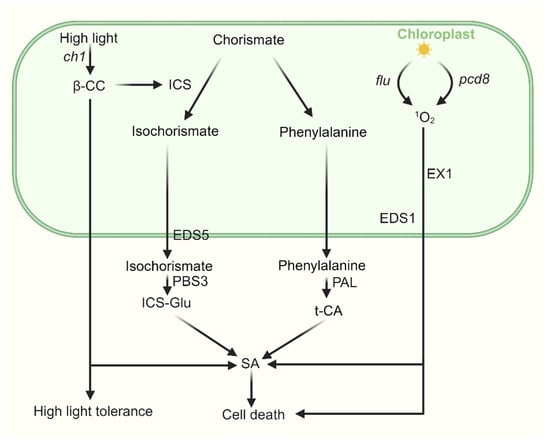
Figure 3.
The cross talk between SA and 1O2-induced signaling. SA is synthesized through two distinct pathways—the ICS pathway and the PAL pathway—both originating from chorismate in the chloroplast, with final biosynthesis occurring in the cytosol. Under high light stress, β-CC enhances plant tolerance by upregulating ICS1 expression and promoting SA biosynthesis, accompanied by associated changes in nuclear gene expression. In pcd8 RNAi mutants, the accumulation of 1O2 stimulates biosynthesis of SA in an EX1-dependent manner. In the flu mutant, upon 1O2 release, SA-responsive genes are strongly induced, activating a complex stress-response program in which EDS1 plays a key role.
4.2. Jasmonate and 1O2 Signaling
Jasmonates (JAs) are lipid-derived cyclopentanones that include JA and its various derivatives. As the major immunity hormone, the jasmonate family of oxylipins promote plant defense to necrotrophic pathogens, chewing insects, and mechanical wounding. Within chloroplasts, PHOSPHOLIPASE A1 (PLDA1) acts on membrane lipids to release α-linolenic acid, which is subsequently oxidized and cyclized by 13-LIPOXYGENASE (13-LOX), ALLENE OXIDE SYNTHASE (AOS), and ALLENE OXIDE CYCLASE (AOC) to form the JA precursor 12-oxo-phytodienoic acid (OPDA) [137,138,139]. OPDA is then transported via the channel formed by the JASSY protein on the chloroplast envelope to peroxisomes [140], where it is catalyzed to form JA by OPDA REDUCTASE 3 (OPR3), ACYL-CoA OXIDASE (ACX), MULTIFUNCTIONAL PROTEIN (MFP), and L-3-KETOACYL CoA THIOLASE (KAT) [139].
Studies on the flu mutant have shown that, when re-exposed to light after dark adaptation, it accumulates OPDA and JA in an EXECUTER 1 (EX1)-dependent manner [141]. When the flu mutant was crossed with the JA-depleted opr3 mutant, the cell death phenotype in the resulting flu opr3 was largely suppressed, indicating that JA promotes 1O2-induced cell death in flu. Whereas other oxylipins such as OPDA and dnOPDA antagonize this cell death-inducing activity of JA [142]. Similarly, during the invision process of necrotrophic Alternaria fungal pathogens on plant species, genes implicated in JA biosynthesis and signaling are induced rapidly, and mutation of the EX1 gene compromises the necrotrophic disease in Arabidopsis. Moreover, the foliar lesion formation caused by Alternaria pathogen in JA-deficient mutants lox3, dde2, aoc3, and jar1 is much smaller than that in wild-type controls, indicating that JA promotes Alternaria pathogen-induced cell death and disease development [113].
Interestingly, under moderate high light stress that does not induce cell death, β-CC-mediated acclimation does not influence JA biosynthesis or signaling. However, under severe high light stress that eventually leads to cell death, JA biosynthesis genes, such as LOX2, AOC1, AOS, and OPR1, are upregulated [16,100,143,144]. When ch1 mutants were exposed to intense high light (1400 μmol m−2 s−1), the expression of LOX2 and LOX3 was induced, JA accumulated, and cell death occurred. Conversely, when ch1 mutants were first acclimated (short exposure to medium light intensity), JA biosynthesis and cell death were suppressed under the same high light conditions [17]. Moreover, ch1 aos double mutants exhibited enhanced tolerance to photooxidative stress under high light [17]. These findings indicate a potential interaction between β-CC and JA biosynthesis/signaling under extreme high light, where JA levels influence both programmed cell death (PCD) and acclimation. Under milder stress, β-CC-mediated retrograde signaling promotes acclimation and suppresses JA biosynthesis.
The gene OXI1, which encodes an AGC kinase family protein localized at the cell periphery, is strongly upregulated in ch1 mutants under cell death-inducing high light [145]. The ch1 oxi1 double mutants showed a coordinated reduction in JA levels and PCD, resulting in a marked suppression of both photooxidative damage and necrosis phenotypes [145]. Conversely, overexpression of OXI1 increased the JA levels and sensitivity to high light stress. Exogenous JA application restored photooxidative damage and cell death in ch1 oxi1 mutants, indicating that OXI1 regulates 1O2-induced PCD via JA. Further studies revealed that DEFENDER AGAINST CELL DEATH 1 (DAD1) and DAD2, two negative regulators of PCD, modulate OXI1 expression under high light [146]. Overexpression of DAD1 and DAD2 suppressed OXI1 expression and reduced JA levels, thereby decreasing sensitivity to photooxidative damage. The relationship of 1O2 and JA is simply summarized in Figure 4.
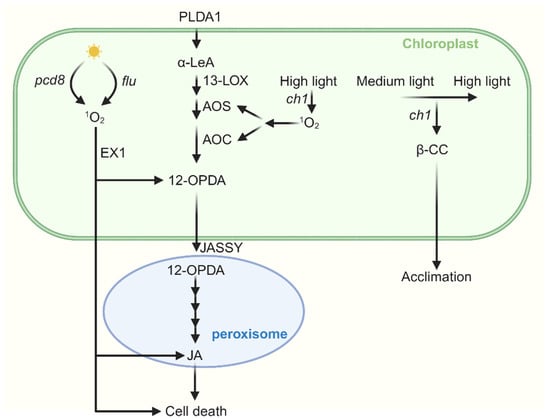
Figure 4.
The cross talk between JA and 1O2-induced signaling. Jasmonic acid (JA) synthesis begins with α-LeA, which is converted to 12-OPDA by 13-LOX, AOS, and AOC. 12-OPDA is then transported from the chloroplast via the outer envelope protein JASSY into peroxisomes, where it is further catalyzed to JA. In the flu mutant, when dark-adapted plants are re-exposed to light, the accumulation of both OPDA and JA occurs in an EX1-dependent manner. Furthermore, JA promotes 1O2-induced cell death in flu. In pcd8 RNAi mutants, the accumulation of 1O2 stimulates biosynthesis of JA and cell death. Under severe high light stress, JA biosynthesis genes are upregulated in the ch1 mutant, leading to JA accumulation and subsequent cell death. Interestingly, when ch1 mutants are first acclimated to medium light and then transferred to the same high light conditions, both JA biosynthesis and cell death are suppressed.
Notably, JA treatment upregulated the SA biosynthesis gene, leading to more severe leaf damage—a phenomenon absent in the SA-deficient mutant ics1, indicating that SA acts downstream of JA in promoting PCD [146]. Additionally, exogenous SA treatment enhanced DAD expression while suppressing OXI1. These results suggest that DADs and OXI1 antagonistically regulate PCD through the modulation of JA and SA pathways. Overall, the relationships among JA, SA, and various retrograde signaling pathways are complex and need further investigation to fully elucidate their interactions.
5. Singlet Oxygen Induces Chloroplast Changes Under Stress
Under ideal environmental conditions, plant cells generate ROS at a low, steady rate, maintaining their concentration within a safe threshold. However, under stress conditions such as high light, high temperatures, drought, or salinity, the balance of photosynthesis is disrupted. This leads to over-reduction in electron transport chains and a consequent surge in ROS production. These highly reactive molecules inflict damage on essential cellular components—including lipids, carbohydrates, proteins, enzymes, and DNA—through oxidative degradation. Such cumulative harm disrupts molecular and cellular integrity, ultimately resulting in plant mortality [147,148].
Chloroplasts are sensitive to stress. Stress affects the structure and components of the thylakoid membrane. A significant transcriptomic correlation has been observed across multiple abiotic stress conditions and 1O2 accumulation [94]. These stresses—including drought, salinity, mechanical injury, and high light exposure—were commonly linked to 1O2-induced disruptions in mRNA translation. This effect is mediated through the direct oxidation of RNA by singlet oxygen [149].
5.1. Abiotic Stress
5.1.1. High Light
High light can induce the unstacking of thylakoids and randomization of pigment-protein complexes of chloroplasts [150]. The highly photosensitive mutant ch1 has less chlorophyll b and releases 1O2 in PSII under photooxidation stress [17]. Light stress damages PSI and PSII, decreases photosynthesis rate, and leads to photoinhibition and photooxidative stress [151]. PSII is one of the primary targets for damage induced by visible and ultraviolet light. High light and UVB can damage the PSII complex at Mn clusters, and induces modification or loss in the function of the QA and QB quinone electron acceptors [152,153]. However, there are some differences between high light stress and UVB stress. Under high light stress, the excessive light energy absorbed by chlorophyll and carotenoids can destroy the integrity of the PSII reaction center in the absence of a functional OEC (oxygen-evolving complex) [154]. Under ultraviolet light, the energy absorbed by the OEC leads to changes in the valency of Mn, which in turn can oxidize other components of PSII [155]. When plants are exposed to more light energy than they need, chlorophyll molecules become over-excited and generate an excess of excitation energy. The over-excitation of PSII promotes the formation of triplet-state excited chlorophyll, which can then excite oxygen to produce 1O2 [2,44].
5.1.2. High Temperature
High temperatures, high salinity, and drought stress do not directly damage PSII as high light stress does. However, they can affect the content of photosynthetic pigments and the repair of PSII [156]. In extreme temperatures, high salinity stress can inhibit the de novo synthesis of the D1 protein, and affect the phosphorylation dynamics within the PSII complex [157,158]. This process is crucial for initiating the degradation and reassembly of PSII [151]. Moreover, abiotic stresses (e.g., extreme temperatures and high salinity) can affect the grana stacking and thylakoid membrane heterogeneity, which in turn impacts the repair of PSII [159]. When exposed to heat stress, the thylakoid membranes have lower thermostability compared to the chloroplast membranes, and the membrane potential is disrupted [160]. High temperature stress induces significant changes in the structure of thylakoid membranes in plants, accompanied with an increase in peroxidation of thylakoid lipids [161,162]. Moderate heat stress does not damage PSII, but it reduces the photosynthetic rate and increases thylakoid leakiness. When plants were exposed to increasing temperature, researchers observed a dissociation of the light-harvesting units of Photosystem II, and phase-separated deaggregates of non-bilayer-forming lipids which can be restored under recovery conditions [163,164]. In addition, high temperatures inactivate Rubisco, thereby affecting carbon fixation [165]. When plants are subjected to low temperature and high light stress, species such as the grasses Sorghum, Paspalum, and the legume soybean, representing the C4 and C3 photosynthetic pathways, exhibit a reduction in the size of starch grains within their chloroplasts [166]. Extreme temperatures cause proteins to precipitate from the thylakoid membranes, affecting photosynthetic phosphorylation, leading to partial inactivation of electron transport through Photosystem II, and the types of precipitated proteins are related to osmolarities [161,167].
5.1.3. Drought and Salt Stress
Drought stress imposes multifaceted constraints on photosynthesis by affecting chloroplast development, reducing Chl content, damaging photosynthetic thylakoid membranes, limiting CO2 availability through stomatal closure, altering enzyme activities, changing the membrane lipid contents, and inducing oxidative stresses. When etiolated wheat seedlings are subjected to drought stress simulated by polyethylene glycol and then exposed to light, the number of chloroplasts formed is significantly reduced, and the number of grana thylakoids within the chloroplasts decrease [168]. Meanwhile, Chl production was significantly decreased in drought-stressed leaves, and swelling or dilation of thylakoid membranes become common during the subsequent greening process [168]. Under osmotic stress, the integrity of the chloroplast envelope remains unaffected, but the level of ATP synthesis is impacted, with a partial inactivation of electron transport through Photosystem II occurring. Drought stress affects the content of membrane lipids. When winter wheat encounters water stress, phosphatidylcholine (PC), phosphatidylethanolamine (PE), and phosphatidylglycerol (PG) are the main targets for degradation, with PC being particularly affected, while the molecular species of monogalactosyldiacylglycerol (MGDG) and digalactosyldiacylglycerol (DGDG) exhibit different degradation time courses [169]. Moreover, short-term drought stress triggered a rapid disassembly of the light-harvesting complex II (LHCII), the amount of the PSII-LHCII supercomplexes and LHCII assemblies are markedly reduced [170,171].
Different levels of salt stress have varying effects on plants, for instance, 50 mM NaCl treatment on Robinia pseudoacacia increases antioxidant enzyme activity and upregulates the expression of the ion transport-related genes and chloroplast development-related genes, while severe salt stress has the opposite effect [172]. Furthermore, the high salt stress also reduced the photochemical efficiency of PSI and PSII and decreased the supercomplexes such as PSII-LHCII in pea (Pisum sativum) [173].
5.1.4. Heavy Metal
Heavy metal toxicity can also damage the chloroplasts and photosynthetic systems of plants. When Oryza sativa was treated with high concentrations of copper (II) oxide nanoparticles (CuO NPs), researchers observed the accumulation of CuO NPs in chloroplasts and a lower number of thylakoids per granum [174]. Chromium (Cr) and lead (Pb) cause distortion of the thylakoid, distortion of chloroplast membrane, and changes in the chloroplast structure [175]. A summary of 1O2-induced chloroplast and photosystem changes under abiotic stresses is shown in Table 2.
Table 2.
Responses of plant chloroplasts and photosynthetic systems to different abiotic stresses.
5.2. Biotic Stress
Chloroplasts are among the most dynamic organelles in plant cells. Beyond their central role in photosynthesis and the synthesis of key plant hormones, they also actively contribute to defense signaling [176]. As a result, chloroplasts have become a critical target for numerous pathogens [177]. Both viral and bacterial invaders employ a variety of sophisticated strategies to disrupt chloroplast structure and function, thereby compromising plant immunity and facilitating their own colonization [176].
A previous study has reported that the infection of Eclipta yellow vein virus (EcYVV-IN) on Andrographis paniculata, resulting in the decrease in carotenoid content and 1O2 quenching, highlighted the alteration in redox status caused by virus-induced biotic stress on the plants [178]. Cercosporin, a toxin secreted by the necrotrophic fungus Cercospora, induces the expression of 1O2-responsive genes. This activation promotes programmed cell death and foliar tissue damage in host plants, ultimately enhancing fungal colonization [179,180]. Tenuazonic acid (TeA), produced by the genus Alternaria and other phytopathogenic fungi, binds to D1 proteins of the PSII reaction center and leads to 1O2 burst that is implicated in chloroplast damage and chloroplast-to-nucleus retrograde signaling.
In plant cells, one of the most significant challenges faced by viruses is the silencing of their RNA [181]. To evade this defense, viruses compartmentalize their replication within specialized vesicles and chloroplasts—structures believed to lack RNA silencing machinery [182,183]. Viral proteins directly target chloroplasts or interact with chloroplast-associated proteins, leading to structural and functional alterations within the organelle. These disruptions commonly manifest as visible symptoms in infected leaves, such as chlorosis, mottling, or mosaic patterning [184]. Virus infection can also inhibit the photosynthetic function of chloroplasts and activate genes related to defense response (genes related to JA and SA signaling pathway) [185,186].
Calcium and ROS serve as the initial signaling molecules in chloroplast-mediated immunity. Following the recognition of pathogen-associated molecular patterns (PAMPs) by pattern recognition receptors (PRRs), rapid phosphorylation cascades are triggered alongside H2O2 production, leading to a rise in cytoplasmic calcium ion (Ca2+) levels [187,188]. This calcium signal subsequently activates the thylakoid-localized calcium-sensing receptor (CAS), resulting in increased Ca2+ concentration within the chloroplast stroma [189]. Concurrently, H2O2 may potentiate early immune responses originating in the chloroplast [190]. The activation of pattern-triggered immunity (PTI) induces a robust burst of chloroplast-derived reactive oxygen species, which in turn stimulates the expression of defense-related genes and represses nuclear-encoded chloroplast genes [188]. This coordinated response ultimately attenuates photosynthetic activity and impairs plastid development. CAS contributes to both basal resistances triggered by PAMPs and hypersensitive cell death mediated by resistance (R) genes. Functioning upstream of SA accumulation, CAS is implicated in the activation of PAMP-induced defense-related genes and the suppression of chloroplast-encoded genes via 1O2-mediated retrograde signaling. This mechanism facilitates chloroplast-driven transcriptional reprogramming during the plant immune response [122,132].
In recent years, a plastid (chloroplast) extension structure named stromule has garnered increasing attention as an emerging focus in plant cell biology. These highly versatile structures extend from plastids and form intimate connections with the nucleus and other organelles, facilitating inter-organellar communication and material exchange [191]. In the plant immune response process, chloroplast-derived stromules play a role in promoting the movement of chloroplasts to the perinuclear region, transducing and amplifying pro-defense signals originating from chloroplasts, and participating in the induction of programmed cell death [192,193]. The formation of stromules during plant immunity is a multifaceted process that involves a complex interplay of signaling components. Initially, plants recognize effectors produced by pathogens through immune receptors, triggering Effector-Triggered Immunity (ETI). Then, stromules can emerge from the surface of chloroplasts as tubular structures, extend along microtubules, and anchor to actin filaments around the nucleus to promote perinuclear chloroplast clustering [194,195]. Stromules often form in response to abiotic and biotic stress, and their formation is associated with ROS, particularly H2O2 [196]. Caplan et al. found that stromules induction is a general response initiated by viral effectors such as the p50 helicase domain of TMV replicases and bacterial effectors such as AvrBS2 during the early phases of a hypersensitive response (HR-PCD). Moreover, stromule induction was not limited to the initially affected cells but also occurs in intact neighboring cells [196].
6. Conclusions Remarks
As a by-product of photosynthesis, 1O2 has been recognized from a simple toxic molecule into a central node within a complex signaling network. This review systematically elaborates on the generation of 1O2 in plant cells and elucidates the molecular mechanisms underlying its perception and downstream responses. More importantly, there exists an intricate cross talk between 1O2 and the signaling networks of two key defense hormones—JA and SA. Firstly, 1O2 production is often linked to photosynthetic disruption and early stress responses, which can activate JA biosynthesis and signaling. This interaction enables plants to initiate defense programs against herbivorous insects and pathogens. Secondly, 1O2 and SA signaling exhibit a complex relationship marked by both antagonism and coordinates. Under high light stress, for instance, 1O2 promotes SA accumulation and enhances plant tolerance. Conversely, 1O2 generated during the transition from dark to light can also induce SA accumulation and trigger cell death. This dynamic cross talk allows plants to adjust their defense strategies appropriately in response to varying 1O2 levels under stress conditions.
Despite significant progress, several key questions in this field remain to be explored. The precise intersection of 1O2 with JA/SA pathways is still not fully understood. Identifying “bridge proteins” that sense the redox state of 1O2 and regulate JA/SA signaling will be crucial for exploring the molecular basis of these interactions. Future studies should focus on identifying key components of 1O2 sensing and signaling, elucidating how 1O2 signaling integrates with JA/SA networks under combined stresses—to more accurately reflect plant survival strategies in natural environments, as well as exploring the usage of 1O2 in modern agriculture and medical fields. In summary, 1O2 constitutes a sophisticated signaling system that links photosynthesis, oxidative stress, and hormone-mediated defense networks. Further research into its production, transduction, and cross talk with phytohormones will not only deepen our understanding on plant environmental adaptation and immune balance but also provide novel theoretical insights and technical approaches for addressing agricultural challenges in the context of global climate change.
Author Contributions
Conceptualization: H.Z., X.W. and L.W.; writing—original draft preparation: H.Z., X.W. and L.W.; writing—review and editing: H.Z.; supervision: L.W.; project administration: L.W.; funding acquisition: L.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the National Key Research and Development Program of China (grant no. 2023YFF1000203-2) and the National Natural Science Foundation of China (NSFC) (grant no. 32170284) to L.W.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study.
Acknowledgments
We thank the National Key Research and Development Program of China and the National Natural Science Foundation of China for providing fundings for this work.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
ABA, Abscisic acid; ACX, Acyl-CoA oxidase; ALA, δ-aminolevulinic acid; AOC, Allene oxide cyclase; AOS, Allene oxide synthase; CAO, Chlorophyll a oxygenase; Car, β-carotenes; CAS, Calcium-sensing receptor; ch1, chlorina1; Chl, Chlorophyll; Chl a, Chlorophyll a; Chl b, Chlorophyll b; 1Chl *, Singlet excited state of chlorophyll; 3Chl *, Triplet excitation state of chlorophyll; Chlide, Chlorophyllide; DAD1, Defender against cell death 1; DGDG, Digalactosyldiacylglycerol; dhA, Dihydroactinidiolide; EcYVV-IN, Eclipta yellow vein virus-[IN]; ETI, Effector-Triggered Immunity; EX1, EXECUTER1; FeCh, Ferrochelatase; GC, Grana core; GLKs, Golden 2-likes; Glu-TR, Glutamyl-tRNA reductase; GM, Grana margin; Gpxh, C. reinhardtii Glutathione peroxidase homologous; H2O2, Hydrogen peroxide; ICS, Isochorismate synthase; ICS-Glu, Isochorismate-9-glutamate; JA, Jasmonic acid; KAT, L-3-ketoacyl CoA thiolase; LHC, Light-harvesting antenna complex; MEP, Methyl erythritol-4-phosphate; MFP, Multifunctional protein; MgCh, Magnesium chelatase; MgCY, Mg-Proto IX monomethylester cyclase; MGDG, Monogalactosyldiacylglycerol; MgMT, Mg-Proto IX methyltransferase; MgP, Mg–protoporphyrin IX; NPQ, Non-photochemical quenching; NPR1, Nonexpressor of pathogenesis-related genes 1; 1O2, Singlet oxygen; 3O2, Triplet molecular oxygen; , Superoxide radical; OH•, Hydroxyl radical; OPDA, 12-Oxo-phytodienoic acid, OPR3, OPDA reductase 3; PAL, Phenylalanine ammonia-lyase; PAMP, Pathogen-associated molecular pattern; PAO, Phenylalanine oxygenase; PC, Phosphatidylcholine; PCD8, Programmed cell death 8; Pchlide, Protochlorophyllide a; PE, Phosphatidylethanolamine; pFCC, primary fluorescence Chl catabolites; PG, Phosphatidylglycerol; Pheo, Pheophytin; PLB, Prolamellar body; PLDA1, Phospholipase A1; POR, Pchlide oxidoreductase; PQ, Plastoquinone; PR1, Pathogenesis related 1; Proto IX, Protoporphyrin IX; PRRs, Pattern recognition receptors; PTI, Pattern-triggered immunity; PUB4, Plant U-box 4; QA, Quinone A; RC, PSII reaction center; RCC, Red chlorophyll catabolite; RCCR, RCC reductase; ROS, Reactive oxygen species; SA, Salicylic acid; SCL14, Scarecrow like 14; SIB1, Sigma factor binding protein 1; SORGs, 1O2-responsive genes; SOSG, Singlet oxygen sensor green; t-CA, trans-cinnamic acid; TeA, Tenuazonic acid; Uro III, Uroporphyrinogen III; β-CC, β-cyclocitral; β-I, β-ionone; 13-LOX, 13-Lipoxygenase.
References
- Mittler, R. ROS are good. Trends Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef]
- Krieger-Liszkay, A. Singlet oxygen production in photosynthesis. J. Exp. Bot. 2005, 56, 337–346. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.; Ahlawat, S.; Mohan, H.; Gill, S.S.; Sharma, K.K. Balancing reactive oxygen species generation by rebooting gut microbiota. J. Appl. Microbiol. 2022, 132, 4112–4129. [Google Scholar] [CrossRef]
- Triantaphylidès, C.; Krischke, M.; Hoeberichts, F.A.; Ksas, B.; Gresser, G.; Havaux, M.; Van Breusegem, F.; Mueller, M.J. Singlet oxygen is the major reactive oxygen species involved in photooxidative damage to plants. Plant Physiol. 2008, 148, 960–968. [Google Scholar] [CrossRef]
- Wang, Y.; Lin, Y.; He, S.; Wu, S.; Yang, C. Singlet oxygen: Properties, generation, detection, and environmental applications. J. Hazard. Mater. 2024, 461, 132538. [Google Scholar] [CrossRef]
- Winslow, M.; Hazelby, A.; Robinson, D. Spin-restricted descriptions of singlet oxygen reactions from XMS-CASPT2 benchmarks. J. Phys. Chem. A 2024, 128, 4128–4137. [Google Scholar] [CrossRef]
- Kashyap, A.; Ramasamy, E.; Ramalingam, V.; Pattabiraman, M. Supramolecular control of singlet oxygen generation. Molecules 2021, 26, 2673. [Google Scholar] [CrossRef]
- Di Mascio, P.; Martinez, G.R.; Miyamoto, S.; Ronsein, G.E.; Medeiros, M.H.G.; Cadet, J. Singlet molecular oxygen reactions with nucleic acids, lipids, and proteins. Chem. Rev. 2019, 119, 2043–2086. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Jan, S.; Singh, R.; Bhardwaj, R.; Ahmad, P.; Kapoor, D. Plant growth regulators: A sustainable approach to combat pesticide toxicity. 3 Biotech 2020, 10, 466. [Google Scholar] [CrossRef] [PubMed]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef]
- Wang, L.; Apel, K. Dose-dependent effects of 1O2 in chloroplasts are determined by its timing and localization of production. J. Exp. Bot. 2019, 70, 29–40. [Google Scholar] [CrossRef]
- Silva, J.N.; Filipe, P.; Morlière, P.; Mazière, J.C.; Freitas, J.P.; Gomes, M.M.; Santus, R. Photodynamic therapy: Dermatology and ophthalmology as main fields of current applications in clinic. Biomed. Mater. Eng. 2008, 18, 319–327. [Google Scholar] [CrossRef]
- op den Camp, R.G.; Przybyla, D.; Ochsenbein, C.; Laloi, C.; Kim, C.; Danon, A.; Wagner, D.; Hideg, E.; Göbel, C.; Feussner, I.; et al. Rapid induction of distinct stress responses after the release of singlet oxygen in Arabidopsis. Plant Cell 2003, 15, 2320–2332. [Google Scholar] [CrossRef]
- Wagner, D.; Przybyla, D.; Op den Camp, R.; Kim, C.; Landgraf, F.; Lee, K.P.; Würsch, M.; Laloi, C.; Nater, M.; Hideg, E.; et al. The genetic basis of singlet oxygen-induced stress responses of Arabidopsis thaliana. Science 2004, 306, 1183–1185. [Google Scholar] [CrossRef]
- Ramel, F.; Birtic, S.; Cuiné, S.; Triantaphylidès, C.; Ravanat, J.L.; Havaux, M. Chemical quenching of singlet oxygen by carotenoids in plants. Plant Physiol. 2012, 158, 1267–1278. [Google Scholar] [CrossRef]
- Ramel, F.; Ksas, B.; Akkari, E.; Mialoundama, A.S.; Monnet, F.; Krieger-Liszkay, A.; Ravanat, J.L.; Mueller, M.J.; Bouvier, F.; Havaux, M. Light-induced acclimation of the Arabidopsis chlorina1 mutant to singlet oxygen. Plant Cell 2013, 25, 1445–1462. [Google Scholar] [CrossRef] [PubMed]
- Woodson, J.D.; Joens, M.S.; Sinson, A.B.; Gilkerson, J.; Salomé, P.A.; Weigel, D.; Fitzpatrick, J.A.; Chory, J. Ubiquitin facilitates a quality-control pathway that removes damaged chloroplasts. Science 2015, 350, 450–454. [Google Scholar] [CrossRef] [PubMed]
- Dogra, V.; Li, M.; Singh, S.; Li, M.; Kim, C. Oxidative post-translational modification of EXECUTER1 is required for singlet oxygen sensing in plastids. Nat. Commun. 2019, 10, 2834. [Google Scholar] [CrossRef] [PubMed]
- Dogra, V.; Kim, C. Singlet oxygen metabolism: From genesis to signaling. Front. Plant Sci. 2019, 10, 1640. [Google Scholar] [CrossRef]
- Jahns, P.; Holzwarth, A.R. The role of the xanthophyll cycle and of lutein in photoprotection of photosystem II. Biochim. Biophys. Acta 2012, 1817, 182–193. [Google Scholar] [CrossRef]
- Dmitrieva, V.A.; Tyutereva, E.V.; Voitsekhovskaja, O.V. singlet oxygen in plants: Generation, detection, and signaling roles. Int. J. Mol. Sci. 2020, 21, 3237. [Google Scholar] [CrossRef]
- Demmig-Adams, B.; Adams, W.W., III. Harvesting sunlight safely. Nature 2000, 403, 371–373. [Google Scholar] [CrossRef] [PubMed]
- Triantaphylidès, C.; Havaux, M. Singlet oxygen in plants: Production, detoxification and signaling. Trends Plant Sci. 2009, 14, 219–228. [Google Scholar] [CrossRef] [PubMed]
- van Mieghem, F.; Brettel, K.; Hillmann, B.; Kamlowski, A.; Rutherford, A.W.; Schlodder, E. Charge recombination reactions in photosystem II. I. Yields, recombination pathways, and kinetics of the primary pair. Biochemistry 1995, 34, 4798–4813. [Google Scholar] [CrossRef]
- Rinalducci, S.; Pedersen, J.Z.; Zolla, L. Formation of radicals from singlet oxygen produced during photoinhibition of isolated light-harvesting proteins of photosystem II. Biochim. Biophys. Acta 2004, 1608, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Santabarbara, S.; Chen, M.; Larkum, A.W.; Evans, M.C. An electron paramagnetic resonance investigation of the electron transfer reactions in the chlorophyll d containing photosystem I of Acaryochloris marina. FEBS Lett. 2007, 581, 1567–1571. [Google Scholar] [CrossRef][Green Version]
- Kvíčalová, Z.; Alster, J.; Hofmann, E.; Khoroshyy, P.; Litvín, R.; Bína, D.; Polívka, T.; Pšenčík, J. Triplet-triplet energy transfer from chlorophylls to carotenoids in two antenna complexes from dinoflagellate Amphidinium carterae. Biochim. Biophys. Acta 2016, 1857, 341–349. [Google Scholar] [CrossRef]
- Magdaong, N.C.M.; Blankenship, R.E. Photoprotective, excited-state quenching mechanisms in diverse photosynthetic organisms. J. Biol. Chem. 2018, 293, 5018–5025. [Google Scholar] [CrossRef]
- Domonkos, I.; Kis, M.; Gombos, Z.; Ughy, B. Carotenoids, versatile components of oxygenic photosynthesis. Prog. Lipid Res. 2013, 52, 539–561. [Google Scholar] [CrossRef]
- Yamano, N.; Wang, P.; Dong, F.Q.; Zhang, J.P. Lipid-enhanced photoprotection of LHCII in membrane nanodisc by reducing chlorophyll triplet production. J. Phys. Chem. B 2022, 126, 2669–2676. [Google Scholar] [CrossRef]
- Wójtowicz, J.; Jagielski, A.K.; Mostowska, A.; Gieczewska, K.B. Compensation mechanism of the photosynthetic apparatus in arabidopsis thaliana ch1 mutants. Int. J. Mol. Sci. 2020, 22, 221. [Google Scholar] [CrossRef]
- Pålsson, L.O.; Spangfort, M.D.; Gulbinas, V.; Gillbro, T. Ultrafast chlorophyll b-chlorophyll a excitation energy transfer in the isolated light harvesting complex, LHC II, of green plants. Implications for the organisation of chlorophylls. FEBS Lett. 1994, 339, 134–138. [Google Scholar] [CrossRef]
- Yang, D.H.; Andersson, B.; Aro, E.M.; Ohad, I. The redox state of the plastoquinone pool controls the level of the light-harvesting chlorophyll a/b binding protein complex II (LHC II) during photoacclimation. Photosynth. Res. 2001, 68, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Guenther, J.E.; Nemson, J.A.; Melis, A. Development of Photosystem II in dark grown Chlamydomonas reinhardtii. A light-dependent conversion of PS IIβ, QB-nonreducing centers to the PS IIα, QB-reducing form. Photosynth. Res. 1990, 24, 35–46. [Google Scholar] [CrossRef]
- Roach, T.; Baur, T.; Stöggl, W.; Krieger-Liszkay, A. Chlamydomonas reinhardtii responding to high light: A role for 2-propenal (acrolein). Physiol. Plant 2017, 161, 75–87. [Google Scholar] [CrossRef]
- Havaux, M.; Ksas, B.; Szewczyk, A.; Rumeau, D.; Franck, F.; Caffarri, S.; Triantaphylidès, C. Vitamin B6 deficient plants display increased sensitivity to high light and photo-oxidative stress. BMC Plant Biol. 2009, 9, 130. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.H.; Song, Y.; Maret, E.L.; Silori, Y.; Willow, R.; Yocum, C.F.; Ogilvie, J.P. Charge separation in the photosystem II reaction center resolved by multispectral two-dimensional electronic spectroscopy. Sci. Adv. 2023, 9, eade7190. [Google Scholar] [CrossRef] [PubMed]
- Rigby, S.E.; Nugent, J.H.; O’Malley, P.J. ENDOR and special triple resonance studies of chlorophyll cation radicals in photosystem 2. Biochemistry 1994, 33, 10043–10050. [Google Scholar] [CrossRef]
- Nadtochenko, V.A.; Shelaev, I.V.; Mamedov, M.D.; Shkuropatov, A.Y.; Semenov, A.Y.; Shuvalov, V.A. Primary radical ion pairs in photosystem II core complexes. Biochemistry 2014, 79, 197–204. [Google Scholar] [CrossRef]
- Hillmann, B.; Brettel, K.; van Mieghem, F.; Kamlowski, A.; Rutherford, A.W.; Schlodder, E. Charge recombination reactions in photosystem II. 2. Transient absorbance difference spectra and their temperature dependence. Biochemistry 1995, 34, 4814–4827. [Google Scholar] [CrossRef]
- Miksovska, J.; Maróti, P.; Tandori, J.; Schiffer, M.; Hanson, D.K.; Sebban, P. Distant electrostatic interactions modulate the free energy level of QA− in the photosynthetic reaction center. Biochemistry 1996, 35, 15411–15417. [Google Scholar] [CrossRef]
- Kálai, T.; Hideg, E.; Vass, I.; Hideg, K. Double (fluorescent and spin) sensors for detection of reactive oxygen species in the thylakoid membrane. Free Radic. Biol. Med. 1998, 24, 649–652. [Google Scholar] [CrossRef]
- Krieger-Liszkay, A.; Fufezan, C.; Trebst, A. Singlet oxygen production in photosystem II and related protection mechanism. Photosynth. Res. 2008, 98, 551–564. [Google Scholar] [CrossRef]
- Pospísil, P. Production of reactive oxygen species by photosystem II. Biochim. Biophys. Acta 2009, 1787, 1151–1160. [Google Scholar] [CrossRef]
- Yang, M.; Huang, H.; Xu, C.; Han, X.; Qin, G.; Chang, L.; Lin, F.; Wang, X.; He, H.; Deng, X.W. Redox-regulated plastoglobule ABC1K1-ABC1K3 kinase complex controls plastoquinone mobilization for chloroplast photosynthetic adaptation to red light. Mol. Plant 2025, 18, 2119–2133. [Google Scholar] [CrossRef]
- Ksas, B.; Légeret, B.; Ferretti, U.; Chevalier, A.; Pospíšil, P.; Alric, J.; Havaux, M. The plastoquinone pool outside the thylakoid membrane serves in plant photoprotection as a reservoir of singlet oxygen scavengers. Plant Cell Environ. 2018, 41, 2277–2287. [Google Scholar] [CrossRef]
- Havaux, M. Plastoquinone in and beyond photosynthesis. Trends Plant Sci. 2020, 25, 1252–1265. [Google Scholar] [CrossRef] [PubMed]
- Martinis, J.; Glauser, G.; Valimareanu, S.; Stettler, M.; Zeeman, S.C.; Yamamoto, H.; Shikanai, T.; Kessler, F. ABC1K1/PGR6 kinase: A regulatory link between photosynthetic activity and chloroplast metabolism. Plant J. 2014, 77, 269–283. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, R.; Tanaka, A. Tetrapyrrole biosynthesis in higher plants. Annu. Rev. Plant Biol. 2007, 58, 321–346. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, R.; Kobayashi, K.; Masuda, T. Tetrapyrrole metabolism in Arabidopsis thaliana. Arab. Book 2011, 9, e0145. [Google Scholar] [CrossRef]
- Davison, P.A.; Schubert, H.L.; Reid, J.D.; Iorg, C.D.; Heroux, A.; Hill, C.P.; Hunter, C.N. Structural and biochemical characterization of Gun4 suggests a mechanism for its role in chlorophyll biosynthesis. Biochemistry 2005, 44, 7603–7612. [Google Scholar] [CrossRef]
- Ilag, L.L.; Kumar, A.M.; Söll, D. Light regulation of chlorophyll biosynthesis at the level of 5-aminolevulinate formation in Arabidopsis. Plant Cell 1994, 6, 265–275. [Google Scholar] [CrossRef]
- Xue, Y.; Dong, H.; Huang, H.; Li, S.; Shan, X.; Li, H.; Liu, H.; Xia, D.; Su, S.; Yuan, Y. Mutation in Mg-protoporphyrin IX monomethyl ester (oxidative) cyclase gene ZmCRD1 causes chlorophyll-deficiency in Maize. Front. Plant Sci. 2022, 13, 912215. [Google Scholar] [CrossRef]
- Li, C.Y.; Hu, S.Y.; Yang, W.T.; Yang, H.Z.; Zhang, W.W.; Ye, J.H.; Zheng, X.Q.; Liang, Y.R.; Dong, Z.B.; Lu, J.L. Conversion obstacle from Mg-protoporphyrin IX to protochlorophyllide might be responsible for chlorophyll-deficient phenotype of the Huangjinya’s albino offspring. Plant Physiol. Biochem. 2024, 212, 108778. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.E.; Canniffe, D.P.; Barnett, S.F.H.; Hollingshead, S.; Brindley, A.A.; Vasilev, C.; Bryant, D.A.; Hunter, C.N. Complete enzyme set for chlorophyll biosynthesis in Escherichia coli. Sci. Adv. 2018, 4, eaaq1407. [Google Scholar] [CrossRef] [PubMed]
- Meskauskiene, R.; Nater, M.; Goslings, D.; Kessler, F.; op den Camp, R.; Apel, K. FLU: A negative regulator of chlorophyll biosynthesis in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2001, 98, 12826–12831. [Google Scholar] [CrossRef]
- Wang, L.; Kim, C.; Xu, X.; Piskurewicz, U.; Dogra, V.; Singh, S.; Mahler, H.; Apel, K. Singlet oxygen- and EXECUTER1-mediated signaling is initiated in grana margins and depends on the protease FtsH2. Proc. Natl. Acad. Sci. USA 2016, 113, E3792–E3800. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhang, F.; Fang, Y.; Chen, X.; Chen, Y.; Zhang, W.; Dai, H.E.; Lin, R.; Liu, L. The non-canonical tetratricopeptide repeat (TPR) domain of Fluorescent (FLU) mediates complex formation with glutamyl-tRNA reductase. J. Biol. Chem. 2015, 290, 17559–17565. [Google Scholar] [CrossRef]
- Vothknecht, U.C.; Kannangara, C.G.; von Wettstein, D. Barley glutamyl tRNAGlu reductase: Mutations affecting haem inhibition and enzyme activity. Phytochemistry 1998, 47, 513–519. [Google Scholar] [CrossRef]
- Kauss, D.; Bischof, S.; Steiner, S.; Apel, K.; Meskauskiene, R. FLU, a negative feedback regulator of tetrapyrrole biosynthesis, is physically linked to the final steps of the Mg(++)-branch of this pathway. FEBS Lett. 2012, 586, 211–216. [Google Scholar] [CrossRef]
- Terry, M.J.; Kendrick, R.E. Feedback inhibition of chlorophyll synthesis in the phytochrome chromophore-deficient aurea and yellow-green-2 mutants of tomato. Plant Physiol. 1999, 119, 143–152. [Google Scholar] [CrossRef]
- Hou, Z.; Pang, X.; Hedtke, B.; Grimm, B. In vivo functional analysis of the structural domains of FLUORESCENT (FLU). Plant J. 2021, 107, 360–376. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.; Apel, K. Singlet oxygen-mediated signaling in plants: Moving from flu to wild type reveals an increasing complexity. Photosynth. Res. 2013, 116, 455–464. [Google Scholar] [CrossRef] [PubMed]
- Khandal, D.; Samol, I.; Buhr, F.; Pollmann, S.; Schmidt, H.; Clemens, S.; Reinbothe, S.; Reinbothe, C. Singlet oxygen-dependent translational control in the tigrina-d.12 mutant of barley. Proc. Natl. Acad. Sci. USA 2009, 106, 13112–13117. [Google Scholar] [CrossRef]
- Zhang, Z.W.; Li, M.X.; Huang, B.; Feng, L.Y.; Wu, F.; Fu, Y.F.; Zheng, X.J.; Peng, H.Q.; Chen, Y.E.; Yang, H.N.; et al. Nitric oxide regulates chlorophyllide biosynthesis and singlet oxygen generation differently between Arabidopsis and barley. Nitric Oxide 2018, 76, 6–15. [Google Scholar] [CrossRef] [PubMed]
- Coll, N.S.; Danon, A.; Meurer, J.; Cho, W.K.; Apel, K. Characterization of soldat8, a suppressor of singlet oxygen-induced cell death in Arabidopsis seedlings. Plant Cell Physiol. 2009, 50, 707–718. [Google Scholar] [CrossRef][Green Version]
- Scharfenberg, M.; Mittermayr, L.; Von Roepenack-Lahaye, E.; Schlicke, H.; Grimm, B.; Leister, D.; Kleine, T. Functional characterization of the two ferrochelatases in Arabidopsis thaliana. Plant Cell Environ. 2015, 38, 280–298. [Google Scholar] [CrossRef]
- Nagai, S.; Koide, M.; Takahashi, S.; Kikuta, A.; Aono, M.; Sasaki-Sekimoto, Y.; Ohta, H.; Takamiya, K.; Masuda, T. Induction of isoforms of tetrapyrrole biosynthetic enzymes, AtHEMA2 and AtFC1, under stress conditions and their physiological functions in Arabidopsis. Plant Physiol. 2007, 144, 1039–1051. [Google Scholar] [CrossRef]
- Lemke, M.D.; Abate, A.N.; Woodson, J.D. Investigating the mechanism of chloroplast singlet oxygen signaling in the Arabidopsis thaliana accelerated cell death 2 mutant. Plant Signal. Behav. 2024, 19, 2347783. [Google Scholar] [CrossRef]
- Tano, D.W.; Kozlowska, M.A.; Easter, R.A.; Woodson, J.D. Multiple pathways mediate chloroplast singlet oxygen stress signaling. Plant Mol. Biol. 2023, 111, 167–187. [Google Scholar] [CrossRef]
- Lemke, M.D.; Woodson, J.D. A genetic screen for dominant chloroplast reactive oxygen species signaling mutants reveals life stage-specific singlet oxygen signaling networks. Front. Plant Sci. 2023, 14, 1331346. [Google Scholar] [CrossRef]
- Geng, R.; Pang, X.; Li, X.; Shi, S.; Hedtke, B.; Grimm, B.; Bock, R.; Huang, J.; Zhou, W. Programmed Cell DEATH8 interacts with tetrapyrrole biosynthesis enzymes and ClpC1 to maintain homeostasis of tetrapyrrole metabolites in Arabidopsis. New Phytol. 2023, 238, 2545–2560. [Google Scholar] [CrossRef]
- Balevičius, V., Jr.; Fox, K.F.; Bricker, W.P.; Jurinovich, S.; Prandi, I.G.; Mennucci, B.; Duffy, C.D.P. Fine control of chlorophyll-carotenoid interactions defines the functionality of light-harvesting proteins in plants. Sci. Rep. 2017, 7, 13956. [Google Scholar] [CrossRef]
- Wang, P.; Grimm, B. Connecting Chlorophyll Metabolism with Accumulation of the Photosynthetic Apparatus. Trends Plant Sci. 2021, 26, 484–495. [Google Scholar] [CrossRef]
- Tanaka, A.; Ito, H. Chlorophyll degradation and its physiological function. Plant Cell Physiol. 2024, 66, 139–152. [Google Scholar] [CrossRef] [PubMed]
- Hörtensteiner, S.; Wüthrich, K.L.; Matile, P.; Ongania, K.H.; Kräutler, B. The key step in chlorophyll breakdown in higher plants. Cleavage of pheophorbide a macrocycle by a monooxygenase. J. Biol. Chem. 1998, 273, 15335–15339. [Google Scholar] [CrossRef]
- Hörtensteiner, S. Chlorophyll breakdown in higher plants and algae. Cell. Mol. Life Sci. 1999, 56, 330–347. [Google Scholar] [CrossRef]
- Wüthrich, K.L.; Bovet, L.; Hunziker, P.E.; Donnison, I.S.; Hörtensteiner, S. Molecular cloning, functional expression and characterisation of RCC reductase involved in chlorophyll catabolism. Plant J. 2000, 21, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Pruzinská, A.; Anders, I.; Aubry, S.; Schenk, N.; Tapernoux-Lüthi, E.; Müller, T.; Kräutler, B.; Hörtensteiner, S. In vivo participation of red chlorophyll catabolite reductase in chlorophyll breakdown. Plant Cell 2007, 19, 369–387. [Google Scholar] [CrossRef] [PubMed]
- Pruzinská, A.; Tanner, G.; Anders, I.; Roca, M.; Hörtensteiner, S. Chlorophyll breakdown: Pheophorbide a oxygenase is a Rieske-type iron-sulfur protein, encoded by the accelerated cell death 1 gene. Proc. Natl. Acad. Sci. USA 2003, 100, 15259–15264. [Google Scholar] [CrossRef]
- Sindhu, A.; Janick-Buckner, D.; Buckner, B.; Gray, J.; Zehr, U.; Dilkes, B.P.; Johal, G.S. Propagation of cell death in dropdead1, a sorghum ortholog of the maize lls1 mutant. PLoS ONE 2018, 13, e0201359. [Google Scholar] [CrossRef]
- Armarego-Marriott, T.; Sandoval-Ibanez, O.; Kowalewska, L. Beyond the darkness: Recent lessons from etiolation and de-etiolation studies. J. Exp. Bot. 2020, 71, 1215–1225. [Google Scholar] [CrossRef]
- Kowalewska, L.; Bykowski, M.; Mostowska, A. Spatial organization of thylakoid network in higher plants. Bot. Lett. 2019, 166, 326–343. [Google Scholar] [CrossRef]
- Stephenson, P.G.; Fankhauser, C.; Terry, M.J. PIF3 is a repressor of chloroplast development. Proc. Natl. Acad. Sci. USA 2009, 106, 7654–7659. [Google Scholar] [CrossRef] [PubMed]
- Moon, J.; Zhu, L.; Shen, H.; Huq, E. PIF1 directly and indirectly regulates chlorophyll biosynthesis to optimize the greening process in Arabidopsis. Proc. Natl. Acad. Sci. USA 2008, 105, 9433–9438. [Google Scholar] [CrossRef]
- Shi, H.; Wang, X.; Mo, X.; Tang, C.; Zhong, S.; Deng, X.W. Arabidopsis DET1 degrades HFR1 but stabilizes PIF1 to precisely regulate seed germination. Proc. Natl. Acad. Sci. USA 2015, 112, 3817–3822. [Google Scholar] [CrossRef]
- Huq, E.; Al-Sady, B.; Hudson, M.; Kim, C.; Apel, K.; Quail, P.H. Phytochrome-interacting factor 1 is a critical bHLH regulator of chlorophyll biosynthesis. Science 2004, 305, 1937–1941. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.; Kim, K.; Kang, H.; Zulfugarov, I.S.; Bae, G.; Lee, C.H.; Lee, D.; Choi, G. Phytochromes promote seedling light responses by inhibiting four negatively-acting phytochrome-interacting factors. Proc. Natl. Acad. Sci. USA 2009, 106, 7660–7665. [Google Scholar] [CrossRef]
- Reinbothe, S.; Reinbothe, C.; Apel, K.; Lebedev, N. Evolution of chlorophyll biosynthesis—The challenge to survive photooxidation. Cell 1996, 86, 703–705. [Google Scholar] [CrossRef]
- Chen, D.; Xu, G.; Tang, W.; Jing, Y.; Ji, Q.; Fei, Z.; Lin, R. Antagonistic basic helix-loop-helix/bZIP transcription factors form transcriptional modules that integrate light and reactive oxygen species signaling in Arabidopsis. Plant Cell 2013, 25, 1657–1673. [Google Scholar] [CrossRef]
- Li, Y.; Liu, H.; Ma, T.; Li, J.; Yuan, J.; Xu, Y.C.; Sun, R.; Zhang, X.; Jing, Y.; Guo, Y.L.; et al. Arabidopsis EXECUTER1 interacts with WRKY transcription factors to mediate plastid-to-nucleus singlet oxygen signaling. Plant Cell 2023, 35, 827–851. [Google Scholar] [CrossRef] [PubMed]
- Prasad, A.; Sedlářová, M.; Kale, R.S.; Pospíšil, P. Lipoxygenase in singlet oxygen generation as a response to wounding: In vivo imaging in Arabidopsis thaliana. Sci. Rep. 2017, 7, 9831. [Google Scholar] [CrossRef] [PubMed]
- Mor, A.; Koh, E.; Weiner, L.; Rosenwasser, S.; Sibony-Benyamini, H.; Fluhr, R. Singlet oxygen signatures are detected independent of light or chloroplasts in response to multiple stresses. Plant Physiol. 2014, 165, 249–261. [Google Scholar] [CrossRef]
- Li, M.; Li, M.; Qi, S.; Wang, L.; Kim, C. Salicylic acid and ROS signaling modulate hypocotyl elongation in darkness via NPR1 and EX1. Sci. Adv. 2025, 11, eadx4417. [Google Scholar] [CrossRef]
- Rosenwasser, S.; Fluhr, R.; Joshi, J.R.; Leviatan, N.; Sela, N.; Hetzroni, A.; Friedman, H. ROSMETER: A bioinformatic tool for the identification of transcriptomic imprints related to reactive oxygen species type and origin provides new insights into stress responses. Plant Physiol. 2013, 163, 1071–1083. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Fluhr, R. Singlet oxygen plays an essential role in the root’s response to osmotic stress. Plant Physiol. 2018, 177, 1717–1727. [Google Scholar] [CrossRef]
- Koh, E.; Carmieli, R.; Mor, A.; Fluhr, R. Singlet oxygen-induced membrane disruption and serpin-protease balance in vacuolar-driven cell death. Plant Physiol. 2016, 171, 1616–1625. [Google Scholar] [CrossRef]
- Leisinger, U.; Rüfenacht, K.; Fischer, B.; Pesaro, M.; Spengler, A.; Zehnder, A.J.; Eggen, R.I. The glutathione peroxidase homologous gene from Chlamydomonas reinhardtii is transcriptionally up-regulated by singlet oxygen. Plant Mol. Biol. 2001, 46, 395–408. [Google Scholar] [CrossRef]
- Ramel, F.; Birtic, S.; Ginies, C.; Soubigou-Taconnat, L.; Triantaphylides, C.; Havaux, M. Carotenoid oxidation products are stress signals that mediate gene responses to singlet oxygen in plants. Proc. Natl. Acad. Sci. USA 2012, 109, 5535–5540. [Google Scholar] [CrossRef]
- Shumbe, L.; Bott, R.; Havaux, M. Dihydroactinidiolide, a high light-induced β-carotene derivative that can regulate gene expression and photoacclimation in Arabidopsis. Mol. Plant 2014, 7, 1248–1251. [Google Scholar] [CrossRef]
- Shao, N.; Duan, G.Y.; Bock, R. A mediator of singlet oxygen responses in Chlamydomonas reinhardtii and Arabidopsis identified by a luciferase-based genetic screen in algal cells. Plant Cell 2013, 25, 4209–4226. [Google Scholar] [CrossRef]
- Shumbe, L.; D’Alessandro, S.; Shao, N.; Chevalier, A.; Ksas, B.; Bock, R.; Havaux, M. METHYLENE BLUE SENSITIVITY 1 (MBS1) is required for acclimation of Arabidopsis to singlet oxygen and acts downstream of β-cyclocitral. Plant Cell Environ. 2017, 40, 216–226. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.P.; Kim, C. Photosynthetic ROS and retrograde signaling pathways. New Phytol. 2024, 244, 1183–1198. [Google Scholar] [CrossRef] [PubMed]
- D’Alessandro, S.; Ksas, B.; Havaux, M. Decoding β-cyclocitral-mediated retrograde signaling reveals the role of a detoxification response in plant tolerance to photooxidative stress. Plant Cell 2018, 30, 2495–2511. [Google Scholar] [CrossRef] [PubMed]
- Mitra, S.; Estrada-Tejedor, R.; Volke, D.C.; Phillips, M.A.; Gershenzon, J.; Wright, L.P. Negative regulation of plastidial isoprenoid pathway by herbivore-induced β-cyclocitral in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2021, 118, e2008747118. [Google Scholar] [CrossRef]
- Dogra, V.; Duan, J.; Lee, K.P.; Lv, S.; Liu, R.; Kim, C. Ftsh2-dependent proteolysis of executer1 is essential in mediating singlet oxygen-triggered retrograde signaling in Arabidopsis thaliana. Front. Plant Sci. 2017, 8, 1145. [Google Scholar] [CrossRef]
- Kato, Y.; Sakamoto, W. Ftsh protease in the thylakoid membrane: Physiological functions and the regulation of protease activity. Front. Plant Sci. 2018, 9, 855. [Google Scholar] [CrossRef]
- Liu, K.; Zhao, H.; Lee, K.P.; Yu, Q.; Di, M.; Wang, L.; Kim, C. EXECUTER1 and singlet oxygen signaling: A reassessment of nuclear activity. Plant Cell 2024, 37, koae296. [Google Scholar] [CrossRef]
- Zhao, H.; Zhang, F.; Wang, X.; Liu, K.; Zhang, L.; Li, J.; Kim, C.; Wang, L. The chloroplast translocon subunit TOC33 relays singlet oxygen-induced chloroplast-to-nucleus retrograde signaling in Arabidopsis. Mol. Plant 2025, 18, 1706–1723. [Google Scholar] [CrossRef]
- Lee, K.P.; Kim, C.; Landgraf, F.; Apel, K. EXECUTER1- and EXECUTER2-dependent transfer of stress-related signals from the plastid to the nucleus of Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2007, 104, 10270–10275. [Google Scholar] [CrossRef]
- Dogra, V.; Singh, R.M.; Li, M.; Li, M.; Singh, S.; Kim, C. EXECUTER2 modulates the EXECUTER1 signalosome through its singlet oxygen-dependent oxidation. Mol. Plant 2022, 15, 438–453. [Google Scholar] [CrossRef]
- Shi, J.; Wang, H.; Li, M.; Mi, L.; Gao, Y.; Qiang, S.; Zhang, Y.; Chen, D.; Dai, X.; Ma, H.; et al. Alternaria TeA toxin activates a chloroplast retrograde signaling pathway to facilitate JA-dependent pathogenicity. Plant Commun. 2024, 5, 100775. [Google Scholar] [CrossRef] [PubMed]
- Page, M.T.; McCormac, A.C.; Smith, A.G.; Terry, M.J. Singlet oxygen initiates a plastid signal controlling photosynthetic gene expression. New Phytol. 2017, 213, 1168–1180. [Google Scholar] [CrossRef]
- Kim, C.; Lee, K.P.; Baruah, A.; Nater, M.; Göbel, C.; Feussner, I.; Apel, K. 1O2-mediated retrograde signaling during late embryogenesis predetermines plastid differentiation in seedlings by recruiting abscisic acid. Proc. Natl. Acad. Sci. USA 2009, 106, 9920–9924. [Google Scholar] [CrossRef]
- Wang, L.; Leister, D.; Guan, L.; Zheng, Y.; Schneider, K.; Lehmann, M.; Apel, K.; Kleine, T. The Arabidopsis SAFEGUARD1 suppresses singlet oxygen-induced stress responses by protecting grana margins. Proc. Natl. Acad. Sci. USA 2020, 117, 6918–6927. [Google Scholar] [CrossRef] [PubMed]
- Chow, K.S.; Singh, D.P.; Walker, A.R.; Smith, A.G. Two different genes encode ferrochelatase in Arabidopsis: Mapping, expression and subcellular targeting of the precursor proteins. Plant J. 1998, 15, 531–541. [Google Scholar] [CrossRef]
- Woodson, J.D. Chloroplast stress signals: Regulation of cellular degradation and chloroplast turnover. Curr. Opin. Plant Biol. 2019, 52, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.X.; Phua, S.Y.; Crisp, P.; McQuinn, R.; Pogson, B.J. Learning the languages of the chloroplast: Retrograde signaling and beyond. Annu. Rev. Plant Biol. 2016, 67, 25–53. [Google Scholar] [CrossRef]
- Dietz, K.J.; Turkan, I.; Krieger-Liszkay, A. Redox- and reactive oxygen species-dependent signaling into and out of the photosynthesizing chloroplast. Plant Physiol. 2016, 171, 1541–1550. [Google Scholar] [CrossRef]
- Dogra, V.; Kim, C. Chloroplast protein homeostasis is coupled with retrograde signaling. Plant Signal. Behav. 2019, 14, 1656037. [Google Scholar] [CrossRef]
- Kachroo, P.; Burch-Smith, T.M.; Grant, M. An emerging role for chloroplasts in disease and defense. Annu. Rev. Phytopathol. 2021, 59, 423–445. [Google Scholar] [CrossRef]
- An, C.; Mou, Z. Salicylic acid and its function in plant immunity. J. Integr. Plant Biol. 2011, 53, 412–428. [Google Scholar] [CrossRef]
- Chen, Z.; Zheng, Z.; Huang, J.; Lai, Z.; Fan, B. Biosynthesis of salicylic acid in plants. Plant Signal. Behav. 2009, 4, 493–496. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.I.; León, J.; Raskin, I. Biosynthesis and metabolism of salicylic acid. Proc. Natl. Acad. Sci. USA 1995, 92, 4076–4079. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.K.; Baek, K.H. Salicylic acid biosynthesis and metabolism: A divergent pathway for plants and bacteria. Biomolecules 2021, 11, 705. [Google Scholar] [CrossRef]
- Lv, F.; Zhou, J.; Zeng, L.; Xing, D. β-cyclocitral upregulates salicylic acid signalling to enhance excess light acclimation in Arabidopsis. J. Exp. Bot. 2015, 66, 4719–4732. [Google Scholar] [CrossRef]
- Mou, Z.; Fan, W.; Dong, X. Inducers of plant systemic acquired resistance regulate NPR1 function through redox changes. Cell 2003, 113, 935–944. [Google Scholar] [CrossRef] [PubMed]
- Kinkema, M.; Fan, W.; Dong, X. Nuclear localization of NPR1 is required for activation of PR gene expression. Plant Cell 2000, 12, 2339–2350. [Google Scholar] [CrossRef]
- Johnson, C.; Boden, E.; Arias, J. Salicylic acid and NPR1 induce the recruitment of trans-activating TGA factors to a defense gene promoter in Arabidopsis. Plant Cell 2003, 15, 1846–1858. [Google Scholar] [CrossRef]
- Ochsenbein, C.; Przybyla, D.; Danon, A.; Landgraf, F.; Göbel, C.; Imboden, A.; Feussner, I.; Apel, K. The role of EDS1 (enhanced disease susceptibility) during singlet oxygen-mediated stress responses of Arabidopsis. Plant J. 2006, 47, 445–456. [Google Scholar] [CrossRef]
- Nomura, H.; Komori, T.; Uemura, S.; Kanda, Y.; Shimotani, K.; Nakai, K.; Furuichi, T.; Takebayashi, K.; Sugimoto, T.; Sano, S.; et al. Chloroplast-mediated activation of plant immune signalling in Arabidopsis. Nat. Commun. 2012, 3, 926, https://doi.org/10.1038/ncomms1926. Correction in Nat. Commun.2013, 4, 1456.. [Google Scholar] [CrossRef]
- Lee, K.P.; Li, M.; Li, M.; Liu, K.; Medina-Puche, L.; Qi, S.; Cui, C.; Lozano-Duran, R.; Kim, C. Hierarchical regulatory module GENOMES UNCOUPLED1-GOLDEN2-LIKE1/2-WRKY18/40 modulates salicylic acid signaling. Plant Physiol. 2023, 192, 3120–3133. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Lee, K.P.; Liu, T.; Dogra, V.; Duan, J.; Li, M.; Xing, W.; Kim, C. Antagonistic modules regulate photosynthesis-associated nuclear genes via GOLDEN2-LIKE transcription factors. Plant Physiol. 2022, 188, 2308–2324. [Google Scholar] [CrossRef] [PubMed]
- Kakizaki, T.; Matsumura, H.; Nakayama, K.; Che, F.S.; Terauchi, R.; Inaba, T. Coordination of plastid protein import and nuclear gene expression by plastid-to-nucleus retrograde signaling. Plant Physiol. 2009, 151, 1339–1353. [Google Scholar] [CrossRef]
- Lv, R.; Li, Z.; Li, M.; Dogra, V.; Lv, S.; Liu, R.; Lee, K.P.; Kim, C. Uncoupled expression of nuclear and plastid photosynthesis-associated genes contributes to cell death in a lesion mimic mutant. Plant Cell 2019, 31, 210–230. [Google Scholar] [CrossRef]
- Reinbothe, C.; Springer, A.; Samol, I.; Reinbothe, S. Plant oxylipins: Role of jasmonic acid during programmed cell death, defence and leaf senescence. FEBS J. 2009, 276, 4666–4681. [Google Scholar] [CrossRef] [PubMed]
- Reymond, P.; Farmer, E.E. Jasmonate and salicylate as global signals for defense gene expression. Curr. Opin. Plant Biol. 1998, 1, 404–411. [Google Scholar] [CrossRef]
- Bali, S.; Gautam, A.; Dhiman, A.; Michael, R.; Dogra, V. Salicylate and jasmonate intertwine in ROS-triggered chloroplast-to-nucleus retrograde signaling. Physiol. Plant. 2023, 175, e14041. [Google Scholar] [CrossRef]
- Guan, L.; Denkert, N.; Eisa, A.; Lehmann, M.; Sjuts, I.; Weiberg, A.; Soll, J.; Meinecke, M.; Schwenkert, S. JASSY, a chloroplast outer membrane protein required for jasmonate biosynthesis. Proc. Natl. Acad. Sci. USA 2019, 116, 10568–10575. [Google Scholar] [CrossRef]
- Przybyla, D.; Göbel, C.; Imboden, A.; Hamberg, M.; Feussner, I.; Apel, K. Enzymatic, but not non-enzymatic, 1O2-mediated peroxidation of polyunsaturated fatty acids forms part of the EXECUTER1-dependent stress response program in the flu mutant of Arabidopsis thaliana. Plant J. 2008, 54, 236–248. [Google Scholar] [CrossRef]
- Danon, A.; Miersch, O.; Felix, G.; Camp, R.G.; Apel, K. Concurrent activation of cell death-regulating signaling pathways by singlet oxygen in Arabidopsis thaliana. Plant J. 2005, 41, 68–80. [Google Scholar] [CrossRef]
- Havaux, M. Carotenoid oxidation products as stress signals in plants. Plant J. 2014, 79, 597–606. [Google Scholar] [CrossRef]
- Ramel, F.; Mialoundama, A.S.; Havaux, M. Nonenzymic carotenoid oxidation and photooxidative stress signalling in plants. J. Exp. Bot. 2013, 64, 799–805. [Google Scholar] [CrossRef]
- Shumbe, L.; Chevalier, A.; Legeret, B.; Taconnat, L.; Monnet, F.; Havaux, M. Singlet oxygen-induced cell death in Arabidopsis under high-light stress is controlled by OXI1 Kinase. Plant Physiol. 2016, 170, 1757–1771. [Google Scholar] [CrossRef]
- Beaugelin, I.; Chevalier, A.; D’Alessandro, S.; Ksas, B.; Novák, O.; Strnad, M.; Forzani, C.; Hirt, H.; Havaux, M.; Monnet, F. OXI1 and DAD regulate light-induced cell death antagonistically through jasmonate and salicylate levels. Plant Physiol. 2019, 180, 1691–1708. [Google Scholar] [CrossRef] [PubMed]
- Sachdev, S.; Ansari, S.A.; Ansari, M.I.; Fujita, M.; Hasanuzzaman, M. Abiotic stress and reactive oxygen species: Generation, signaling, and defense mechanisms. Antioxidants 2021, 10, 277. [Google Scholar] [CrossRef]
- Zandi, P.; Schnug, E. Reactive Oxygen Species, Antioxidant responses and implications from a microbial modulation perspective. Biology 2022, 11, 155. [Google Scholar] [CrossRef] [PubMed]
- Koh, E.; Cohen, D.; Brandis, A.; Fluhr, R. Attenuation of cytosolic translation by RNA oxidation is involved in singlet oxygen-mediated transcriptomic responses. Plant Cell Environ. 2021, 44, 3597–3615. [Google Scholar] [CrossRef]
- Sharma, M.; Bhardwaj, U.; Bhardwaj, R. High light-induced unstacking of thylakoids and randomization of pigment-protein complexes of chloroplast. J. Plant Biochem. Biot. 1996, 5, 131–133. [Google Scholar] [CrossRef]
- Didaran, F.; Kordrostami, M.; Ghasemi-Soloklui, A.A.; Pashkovskiy, P.; Kreslavski, V.; Kuznetsov, V.; Allakhverdiev, S.I. The mechanisms of photoinhibition and repair in plants under high light conditions and interplay with abiotic stressors. J. Photochem. Photobiol. B 2024, 259, 113004. [Google Scholar] [CrossRef] [PubMed]
- Eberhard, S.; Finazzi, G.; Wollman, F.A. The dynamics of photosynthesis. Annu. Rev. Genet. 2008, 42, 463–515. [Google Scholar] [CrossRef]
- Sharma, N.; Nagar, S.; Thakur, M.; Suriyakumar, P.; Kataria, S.; Shanker, A.K.; Landi, M.; Anand, A. Photosystems under high light stress: Throwing light on mechanism and adaptation. Photosynthetica 2023, 61, 250–263. [Google Scholar] [CrossRef]
- Shi, Y.; Ke, X.; Yang, X.; Liu, Y.; Hou, X. Plants response to light stress. J. Genet. Genom. 2022, 49, 735–747. [Google Scholar] [CrossRef]
- Hou, X.; Hou, H.J.M. Roles of manganese in photosystem II dynamics to irradiations and temperatures. Front. Biol. 2013, 8, 312–322. [Google Scholar] [CrossRef]
- Muhammad, I.; Shalmani, A.; Ali, M.; Yang, Q.H.; Ahmad, H.; Li, F.B. Mechanisms regulating the dynamics of photosynthesis under abiotic stresses. Front. Plant Sci. 2020, 11, 615942. [Google Scholar] [CrossRef]
- Allakhverdiev, S.I.; Murata, N. Environmental stress inhibits the synthesis de novo of proteins involved in the photodamage-repair cycle of Photosystem II in Synechocystis sp. PCC 6803. Biochim. Biophys. Acta 2004, 1657, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Allakhverdiev, S.I.; Nishiyama, Y.; Miyairi, S.; Yamamoto, H.; Inagaki, N.; Kanesaki, Y.; Murata, N. Salt stress inhibits the repair of photodamaged photosystem II by suppressing the transcription and translation of psbA genes in Synechocystis. Plant Physiol. 2002, 130, 1443–1453. [Google Scholar] [CrossRef]
- Singh, D.P.; Kshatriya, K. NaCl-induced oxidative damage in the cyanobacterium Anabaena doliolum. Curr. Microbiol. 2002, 44, 411–417. [Google Scholar] [CrossRef]
- Krause, G.H.; Santarius, K.A. Relative thermostability of chloroplast envelope. Planta 1975, 127, 285–299. [Google Scholar] [CrossRef] [PubMed]
- Santarius, K.A.; Müller, M. Investigations on heat resistance of spinach leaves. Planta 1979, 146, 529–538. [Google Scholar] [CrossRef] [PubMed]
- Mishra, R.K.; Singhal, G.S. Function of photosynthetic apparatus of intact wheat leaves under high light and heat-stress and its relationship with peroxidation of thylakoid lipids. Plant Physiol. 1992, 98, 1–6. [Google Scholar] [CrossRef]
- Haldimann, P.; Feller, U. Growth at moderately elevated temperature alters the physiological response of the photosynthetic apparatus to heat stress in pea (Pisum sativum L.) leaves. Plant Cell Environ. 2005, 28, 302–317. [Google Scholar] [CrossRef]
- Rath, J.R.; Pandey, J.; Yadav, R.M.; Zamal, M.Y.; Ramachandran, P.; Mekala, N.R.; Allakhverdiev, S.I.; Subramanyam, R. Temperature-induced reversible changes in photosynthesis efficiency and organization of thylakoid membranes from pea (Pisum sativum). Plant Physiol. Biochem. 2022, 185, 144–154. [Google Scholar] [CrossRef] [PubMed]
- Salvucci, M.E.; Crafts-Brandner, S.J. Mechanism for deactivation of Rubisco under moderate heat stress. Physiol. Plant. 2004, 122, 513–519. [Google Scholar] [CrossRef]
- Taylor, A.O.; Craig, A.S. Plants under climatic stress: II. Low temperature, high light effects on chloroplast ultrastructure. Plant Physiol. 1971, 47, 719–725. [Google Scholar] [CrossRef]
- Hincha, D.K.; Schmidt, J.E.; Heber, U.; Schmitt, J.M. Colligative and non-colligative freezing damage to thylakoid membranes. Biochim. Biophys. Acta 1984, 769, 8–14. [Google Scholar] [CrossRef]
- Freeman, T.P.; Duysen, M.E. Effect of imposed water stress on development and ultrastructure of wheat chloroplasts. Protoplasma 1975, 83, 131–145. [Google Scholar] [CrossRef]
- Wang, Y.J.; Zhang, X.Y.; Huang, G.R.; Feng, F.; Liu, X.Y.; Guo, R.; Gu, F.X.; Zhong, X.L.; Mei, X.R. Dynamic changes in membrane lipid composition of leaves of winter wheat seedlings in response to PEG-induced water stress. BMC Plant Biol. 2020, 20, 84. [Google Scholar] [CrossRef]
- Chen, Y.E.; Liu, W.J.; Su, Y.Q.; Cui, J.M.; Zhang, Z.W.; Yuan, M.; Zhang, H.Y.; Yuan, S. Different response of photosystem II to short and long-term drought stress in Arabidopsis thaliana. Physiol. Plant. 2016, 158, 225–235. [Google Scholar] [CrossRef]
- Pandey, J.; Devadasu, E.; Saini, D.; Dhokne, K.; Marriboina, S.; Raghavendra, A.S.; Subramanyam, R. Reversible changes in structure and function of photosynthetic apparatus of pea (Pisum sativum) leaves under drought stress. Plant J. 2023, 113, 60–74. [Google Scholar] [CrossRef]
- Lu, C.X.; Li, L.Y.; Liu, X.L.; Chen, M.; Wan, S.B.; Li, G.W. Salt stress inhibits photosynthesis and destroys chloroplast structure by downregulating chloroplast development-related genes in Robinia pseudoacacia seedlings. Plants 2023, 12, 1283. [Google Scholar] [CrossRef] [PubMed]
- Dhokne, K.; Pandey, J.; Yadav, R.M.; Ramachandran, P.; Rath, J.R.; Subramanyam, R. Change in the photochemical and structural organization of thylakoids from pea (Pisum sativum) under salt stress. Plant Physiol. Biochem. 2022, 177, 46–60. [Google Scholar] [CrossRef]
- Da Costa, M.V.J.; Sharma, P.K. Effect of copper oxide nanoparticles on growth, morphology, photosynthesis, and antioxidant response in Oryza sativa. Photosynthetica 2016, 54, 110–119. [Google Scholar] [CrossRef]
- Choudhury, S.; Panda, S.K. Toxic effects, oxidative stress and ultrastructural changes in moss Taxithelium nepalense (Schwaegr.) Broth. Under chromium and lead phytotoxicity. Water Air Soil Pollut. 2005, 167, 73–90. [Google Scholar] [CrossRef]
- Bhattacharyya, D.; Chakraborty, S. Chloroplast: The Trojan horse in plant-virus interaction. Mol. Plant Pathol. 2018, 19, 504–518. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Xiao, K.; Pan, H.; Liu, J. Chloroplast: The emerging battlefield in plant-microbe interactions. Front. Plant Sci. 2021, 12, 637853. [Google Scholar] [CrossRef]
- Khan, A.; Luqman, S.; Masood, N.; Singh, D.K.; Saeed, S.T.; Samad, A. Eclipta yellow vein virus enhances chlorophyll destruction, singlet oxygen production and alters endogenous redox status in Andrographis paniculata. Plant Physiol. Biochem. 2016, 104, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Daub, M.E.; Ehrenshaft, M. The photoactivated cercospora toxin cercosporin: Contributions to plant disease and fundamental biology. Annu. Rev. Phytopathol. 2000, 38, 461–490. [Google Scholar] [CrossRef]
- Santos Rezende, J.; Zivanovic, M.; Costa de Novaes, M.I.; Chen, Z.Y. The AVR4 effector is involved in cercosporin biosynthesis and likely affects the virulence of Cercospora cf. flagellaris on soybean. Mol. Plant Pathol. 2020, 21, 53–65. [Google Scholar] [CrossRef]
- Pumplin, N.; Voinnet, O. RNA silencing suppression by plant pathogens: Defence, counter-defence and counter-counter-defence. Nat. Rev. Microbiol. 2013, 11, 745–760. [Google Scholar] [CrossRef]
- Wei, T.; Huang, T.S.; McNeil, J.; Laliberté, J.F.; Hong, J.; Nelson, R.S.; Wang, A. Sequential recruitment of the endoplasmic reticulum and chloroplasts for plant potyvirus replication. J. Virol. 2010, 84, 799–809. [Google Scholar] [CrossRef]
- Torrance, L.; Cowan, G.H.; Gillespie, T.; Ziegler, A.; Lacomme, C. Barley stripe mosaic virus-encoded proteins triple-gene block 2 and γb localize to chloroplasts in virus-infected monocot and dicot plants, revealing hitherto-unknown roles in virus replication. J. Gen. Virol. 2006, 87, 2403–2411. [Google Scholar] [CrossRef]
- Ershova, N.; Sheshukova, E.; Kamarova, K.; Arifulin, E.; Tashlitsky, V.; Serebryakova, M.; Komarova, T. Nicotiana benthamiana Kunitz peptidase inhibitor-like protein involved in chloroplast-to-nucleus regulatory pathway in plant-virus interaction. Front. Plant Sci. 2022, 13, 1041867. [Google Scholar] [CrossRef]
- Reinero, A.; Beachy, R.N. Reduced photosystem II activity and accumulation of viral coat protein in chloroplasts of leaves infected with tobacco mosaic virus. Plant Physiol. 1989, 89, 111–116. [Google Scholar] [CrossRef]
- Souza, P.F.N.; Garcia-Ruiz, H.; Carvalho, F.E.L. What proteomics can reveal about plant-virus interactions? Photosynthesis-related proteins on the spotlight. Theor. Exp. Plant Physiol. 2019, 31, 227–248. [Google Scholar] [CrossRef] [PubMed]
- Weralupitiya, C.; Eccersall, S.; Meisrimler, C.N. Shared signals, different fates: Calcium and ROS in plant PRR and NLR immunity. Cell Rep. 2024, 43, 114910. [Google Scholar] [CrossRef]
- Rui, L.; Yang, S.Q.; Zhou, X.H.; Wang, W. The important role of chloroplasts in plant immunity. Plant Commun. 2025, 6, 101420. [Google Scholar] [CrossRef]
- Li, B.; Hou, L.; Song, C.; Wang, Z.; Xue, Q.; Li, Y.; Qin, J.; Cao, N.; Jia, C.; Zhang, Y.; et al. Biological function of calcium-sensing receptor (CAS) and its coupling calcium signaling in plants. Plant Physiol. Biochem. 2022, 180, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhou, X.; Li, F. H2O2-driven plant immunity requires post-translational modification as a switch. Trends Plant Sci. 2025, 30, 591–593. [Google Scholar] [CrossRef] [PubMed]
- Itoh, R. Tubular extensions of plant organelles and their implications on retrograde signaling. J. Biol. Res. 2023, 96, 11724. [Google Scholar] [CrossRef]
- Brunkard, J.O.; Runkel, A.M.; Zambryski, P.C. Chloroplasts extend stromules independently and in response to internal redox signals. Proc. Natl. Acad. Sci. USA 2015, 112, 10044–10049. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.; Woo, J.; Park, E. Talk to your neighbors in an emergency: Stromule-mediated chloroplast-nucleus communication in plant immunity. Curr. Opin. Plant Biol. 2024, 79, 102529. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.S.; Park, E.; Nedo, A.; Alqarni, A.; Ren, L.; Hoban, K.; Modla, S.; McDonald, J.H.; Kambhamettu, C.; Dinesh-Kumar, S.P.; et al. Stromule extension along microtubules coordinated with actin-mediated anchoring guides perinuclear chloroplast movement during innate immunity. Elife 2018, 7, e23625. [Google Scholar] [CrossRef]
- Meier, N.D.; Seward, K.; Caplan, J.L.; Dinesh-Kumar, S.P. Calponin homology domain containing kinesin, KIS1, regulates chloroplast stromule formation and immunity. Sci. Adv. 2023, 9, eadi7407. [Google Scholar] [CrossRef]
- Caplan, J.L.; Kumar, A.S.; Park, E.; Padmanabhan, M.S.; Hoban, K.; Modla, S.; Czymmek, K.; Dinesh-Kumar, S.P. Chloroplast stromules function during innate immunity. Dev. Cell 2015, 34, 45–57. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.