Abstract
Parkinson’s disease (PD) is a progressive, irreversible neurodegenerative disorder characterized by motor impairments and a wide spectrum of non-motor symptoms, including gastrointestinal dysfunction, sleep disturbances, depression, and cognitive decline. These manifestations arise from disturbances across multiple systems—gastrointestinal, neuroendocrine, immune, enteric, and central nervous systems. Alterations in the gut microbiota may play a causal role in PD onset and frequently accompany disease progression. The gut–brain axis, particularly the vagus nerve, is increasingly recognized as a key communication pathway whose dysregulation contributes to systemic dysfunction and the breakdown of homeostasis, ultimately driving PD pathology. Currently, there is no cure for PD, and existing treatments primarily target symptom relief. Effective management of PD requires a comprehensive approach that integrates multiple pharmacologically active agents aimed at restoring impaired organ functions and, when possible, neutralizing toxic factors that accelerate disease progression. One promising therapeutic avenue lies in functional gut bacteria, which form the basis for developing live biotherapeutic products, postbiotics, and bacterial vesicles. In this review, we summarize current data on the effects of probiotics in PD, drawing on both animal models and clinical studies. We highlight the role of probiotics in modulating PD pathophysiology and discuss their potential as adjunctive therapeutic agents. To provide a broader perspective, we also include sections describing the clinical manifestations of PD, gut microbiota alterations associated with the disease, and the role of artificial intelligence, particularly machine learning, in constructing functional models of PD.
1. Introduction
Parkinson’s disease (PD) is a progressive, irreversible neurodegenerative disorder that affects multiple organ systems, including the gastrointestinal, neuroendocrine, immune, enteric, and central nervous systems (CNS) [1,2]. A key biomarker of PD is the formation of Lewy bodies in the neurons of the substantia nigra, resulting from the misfolding and aggregation of the alpha-synuclein protein [3]. The gut–brain axis is considered one of the body’s most important communication systems [4,5]. According to the Braak hypothesis, the initial trigger of PD may be an unknown pathogenic factor in the gut [6]. This factor could induce alpha-synuclein misfolding in enteric neurons, followed by the translocation of misfolded proteins and their mediators into the CNS [7]. In recent years, gut microbiota bacteria have been investigated as potential mediators of alpha-synuclein misfolding [8], and neuronal and behavioral dysfunctions [9,10] and prion-like mechanisms have also been proposed [11]. Strategies to prevent this process include immunotherapeutic approaches such as active immunization with peptide vaccines designed to elicit an immune response against insoluble or oligomeric alpha-synuclein and passive immunization with antibodies that directly target alpha-synuclein aggregates (e.g., Prasinezumab, Cinpanemab) [12].
However, current findings suggest the existence of additional mechanisms driving protein misfolding and transfer. It is now established that bacterial vesicles from the gut microbiota and human exosomes can package and transport proteins, metabolites, small RNAs, and other signaling molecules across the intestinal and blood–brain barriers [13,14,15]. This emerging knowledge highlights new avenues for identifying therapeutic targets in PD. In recent years, increasing attention has been directed toward the therapeutic potential of probiotic strains. Several probiotics and their formulations are currently in clinical trials as candidate treatments for PD.
In this review, we present an overview of current findings on the effects of probiotics in PD, based on evidence from animal models and clinical studies, with the aim of clarifying mechanisms of bacterial action and evaluating their therapeutic potential. To provide better context, we also include sections summarizing the pathology of PD, characteristic alterations in the gut microbiota, and the contribution of artificial intelligence, particularly machine learning, in developing functional PD models. We critically analyze recent experimental studies in this field, with a focus on the development of therapeutic products derived from functional bacteria with neuromodulatory, immunomodulatory, and anti-inflammatory (including neuroinflammatory) properties.
2. Current Understanding of the Development of Parkinson’s Disease
2.1. Etiology of Parkinson’s Disease
PD is a slowly progressing chronic neurodegenerative disorder that affects 1–4% of individuals over the age of 60. It occurs in both sporadic and hereditary forms, with contributing factors including genetic mutations and environmental exposures. PD is classified into primary (idiopathic), secondary (symptomatic), and neurodegenerative PD subtypes [16,17]. A hallmark of PD is the degeneration of neurons in the substantia nigra pars compacta (SN) and the accumulation of α-synuclein–containing protein aggregates (Lewy bodies). Damage to the nigrostriatal pathway leads to dopamine depletion in the striatum, which underlies the characteristic motor symptoms of the disease, such as hypokinesia, rigidity, and resting tremor. Involvement of additional neuronal pathways contributes to cognitive and autonomic dysfunctions [18,19].
Peripheral nervous system involvement has also been reported, including α-synuclein accumulation in the enteric nervous plexuses and salivary glands, which is associated with gastrointestinal dysmotility and impaired salivation [20]. Both exogenous (environmental) and endogenous (genetic) factors are implicated in PD pathogenesis. Endogenous factors include Mendelian-inherited genetic mutations. Monogenic forms of PD are linked to six of the 28 chromosomal regions associated with the disease. However, incomplete penetrance, variable expression, and phenocopies complicate assessment of whether PD is exclusively genetic or influenced by environmental modifiers [21].
Environmental exposure, particularly to pesticides and industrial chemicals, is associated with increased PD risk. Chronic exposure to rotenone, paraquat, and solvents such as trichloroethylene and perchloroethylene in humans has been shown to promote Lewy body formation, α-synuclein misfolding, and mitochondrial dysfunction, ultimately triggering oxidative stress and apoptosis. Irreversible parkinsonism can also be induced by neurotoxins such as 6-hydroxydopamine (6-OHDA) and 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP), which are metabolized into toxic compounds that accumulate in dopaminergic neurons and lead to their degeneration. Traumatic brain injury is another contributing factor, as it can damage the basal ganglia and their connections, disrupt the blood–brain barrier, and initiate prolonged neuroinflammation, mitochondrial dysfunction, excessive glutamate release, and α-synuclein aggregation in the brain. Together, these mechanisms may contribute to the development of secondary parkinsonism [17,22].
2.2. Motor and Non-Motor Symptoms in Parkinson’s Disease
Parkinson’s disease manifests through motor disturbances such as hypokinesia, muscle rigidity, resting tremor, and postural instability. The characteristic “petitioner’s posture” in PD is associated with increased tone in the flexor musculature [23,24]. Disease progression involves not only the dopaminergic system but also the cholinergic, serotonergic, and noradrenergic systems of the brain. Damage to these systems contributes to the development of non-motor symptoms (NMS), which persist throughout the disease course.
NMS are classified as autonomic, sensory, sleep-related, and neuropsychiatric. Autonomic dysfunction can affect nearly all functional systems of the body. Among the most common are gastrointestinal disturbances, which often appear in the early stages of PD—sometimes even before motor symptoms. These include impaired intestinal motility, dysphagia (difficulty swallowing), and gastroparesis (delayed gastric emptying), which hinder the absorption of medications, particularly levodopa, thereby reducing therapeutic efficacy. Patients may also experience heartburn, reflux, and other digestive problems caused by altered gastric and sphincter motility [20,25].
Olfactory dysfunction (hyposmia) is another frequent early marker of PD [26]. The disease also disrupts the regulation of sleep and wakefulness, leading to insomnia, excessive daytime sleepiness, and impaired motor control during REM sleep [27]. Cognitive impairments and dementia typically emerge in later stages, manifesting as reduced attention, memory deficits, and impaired visuospatial orientation. Common psychiatric disturbances include depression, apathy, hallucinations, and impulse control disorders, the latter often induced by pharmacological therapy [28]. Some NMS may also arise directly as side effects of treatment.
Despite their key role in PD, NMS remain underdiagnosed and insufficiently studied.
2.3. Pathogenesis of Parkinson’s Disease
Catecholamines, including dopamine, are synthesized from L-tyrosine through the action of tyrosine hydroxylase (TH), the rate-limiting enzyme in dopamine biosynthesis. In patients with PD, TH activity is significantly reduced in the nigrostriatal system, as well as in other brain regions, reflecting the systemic nature of neurodegeneration [29,30]. Dopamine metabolism generates reactive oxygen species (ROS) and quinones, which further exacerbate mitochondrial dysfunction and promote aggregation of α-synuclein—a protein central to PD pathogenesis [31,32].
Morphologically, PD is characterized by Lewy bodies—intracellular aggregates of abnormal, insoluble α-synuclein—found in both central and peripheral nervous system structures. Their widespread presence, particularly in advanced stages, confirms the progressive nature of the disease. The extent of dopaminergic cell loss correlates with Lewy body burden [33,34].
A hallmark of PD is oxidative stress, driven by mitochondrial dysfunction and ROS accumulation. Dopaminergic neurons are especially vulnerable due to their high oxygen metabolism, low antioxidant capacity, and elevated iron content. Increasing evidence highlights oxidative stress as a major driver of dopaminergic neurodegeneration across all forms of PD [35,36].
Iron plays a central role in free radical–producing oxidative reactions. Under physiological conditions, the SN is among the brain regions with the highest iron content. Abnormally elevated iron levels in the SN have been demonstrated in both genetic and idiopathic PD, linking the disease to disrupted iron metabolism [37]. With aging, glutathione levels and other antioxidant defense components decline, rendering neurons less capable of mounting stress responses [38].
Chronic neuroinflammation is another key factor in PD pathogenesis. Damaged neurons release α-synuclein, which activates astrocytes and microglia. These glial cells, in turn, produce proinflammatory cytokines, chemokines, and ROS, further amplifying tissue injury and neurodegeneration [39,40,41].
Taken together, these findings support the view of PD as a multifactorial disorder driven by diverse pathogenic processes at the cellular level. The human microbiota—particularly the gut microbiota—also contributes to PD onset and progression. Alterations in gut microbial composition may influence disease mechanisms, exacerbate inflammation, and aggravate symptoms. Moreover, the microbiota is increasingly studied as a potential source of therapeutic agents or adjuvants in PD treatment. These aspects will be discussed in the following sections of this review.
2.4. Modeling Parkinson’s Disease in Rodents
Postmortem analysis of PD-affected human brain tissue provides insights primarily into the terminal stages of the disease and is unsuitable for investigating early neurodegenerative events. Therefore, experimental model systems are essential for studying different aspects of PD. Commonly used models include rodents (rats and mice), non-human primates, and model organisms such as Caenorhabditis elegans, Drosophila melanogaster, and zebrafish. Among these, rodents are the most widely used and accessible. To reproduce disease symptoms, researchers employ both genetic models (e.g., knockout mice for genes such as α-synuclein, Parkin, UCH-L1, PINK1, DJ1, LRRK2, or overexpression of mutant human α-synuclein) and neurotoxic models [42]. Exogenous neurotoxins remain the most frequently applied approach [43].
The first PD animal model was established through intracerebral injection of 6-OHDA in rats [44]. 6-OHDA is a structural analog of catecholamines (dopamine and norepinephrine) that selectively destroys dopaminergic neurons by generating ROS. Since it cannot cross the blood–brain barrier (BBB), stereotactic surgery is required to deliver it directly into the SN [45,46]. A more widely used model involves MPTP, which readily crosses the BBB and therefore does not require surgical administration. MPTP is metabolized into the toxic compound MPP+, which damages dopaminergic neurons and replicates both motor and non-motor symptoms of PD [47]. MPTP administration in monkeys is considered the gold standard, though similar studies are also conducted in rodents [48].
Agricultural pesticides such as paraquat and rotenone also target the dopaminergic system. Paraquat, a nitrogen-based herbicide widely used to control broadleaf weeds, is structurally similar to MPP+, the toxic MPTP metabolite. Rotenone, used to eliminate invasive fish in lakes and reservoirs, is a potent dopaminergic neurotoxin with high lipophilicity that allows it to cross the BBB and enter cells. However, its application is limited by systemic toxicity. Both paraquat and rotenone disrupt mitochondrial function, increase ROS production, and lead to dopaminergic neuron loss and motor deficits [49,50,51].
3. Gut Microbiome and Parkinson’s Disease
The study of the human gut microbiome has become one of the most rapidly advancing interdisciplinary fields in biomedical science. A central objective—identifying key parameters and characterizing the microbiome in its healthy state—has been set and is being addressed with increasing success [52,53,54]. Over the past decade, the idea has emerged that the microbiome functions as an independent organ, integrating environmental factors with major physiological systems of the human body through the ENS and CNS via the gut–brain axis [55] (Figure 1).
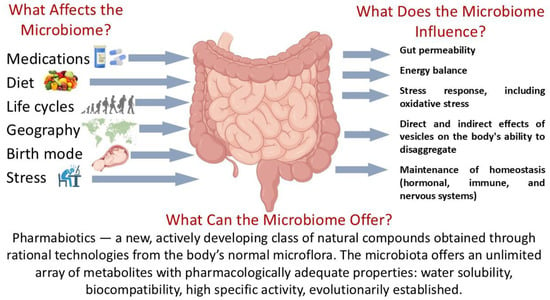
Figure 1.
The human gut microbiome: functional activity in the norm. Parts of this figure were created in BioRender (2025) https://BioRender.com/135e66q (accessed on 1 July 2025).
The relationship between gut microbiota and PD is complex. The microbiome may serve as a reservoir, a trigger, and even a source of therapeutic agents for PD. The Braak hypothesis and its supporting evidence were discussed in the Introduction. Additional confirmation that gut microbiota can act as a disease trigger comes from studies showing that fecal transplantation from PD donors to recipient mice induces not only disruptions of gut homeostasis but also motor and pathological features characteristic of PD [56].
Further studies have distinguished gut-first and brain-first PD subtypes. In 2019, Borghammer and Van Den Berge demonstrated different distributions of alpha-synuclein aggregates in postmortem PD patients: in one group, aggregates were predominantly located in the brain (brain-first), while in the other, they were mainly in the gut (gut-first) [57]. This led to the hypothesis that PD pathology originates either in the gut or in the brain. Gut-first PD is characterized by symmetric motor symptoms and bilateral brain involvement, whereas brain-first PD typically begins in one hemisphere, producing asymmetric motor symptoms [58]. Differences in the severity of gut microbiota alterations among PD patients are likely attributable to subtype. Indeed, the most pronounced microbiota changes are observed in patients with symmetric motor symptoms [59].
A significant portion of patients with PD exhibit gut dysfunction, including inflammatory processes, increased intestinal permeability, and dysbiosis [60]. Figure 2 schematically illustrates the relationship between negative changes in microbiome composition (dysbiosis) and the subsequent stages in the development of PD. Key mechanisms involved include oxidative stress and changes in the enteric nervous system, which, through neuroinflammation, lead to aggregation of α-synuclein proteins and the formation of Lewy bodies [61]. Simultaneously, there is immune system dysregulation, resulting in persistently elevated levels of pro-inflammatory cytokines, as well as dysfunction of the vagus nerve, disrupting communication between the enteric nervous system and the brain [7]. At later stages, these processes culminate in neurodegeneration, particularly the degeneration of dopaminergic neurons and a decline in dopamine levels.
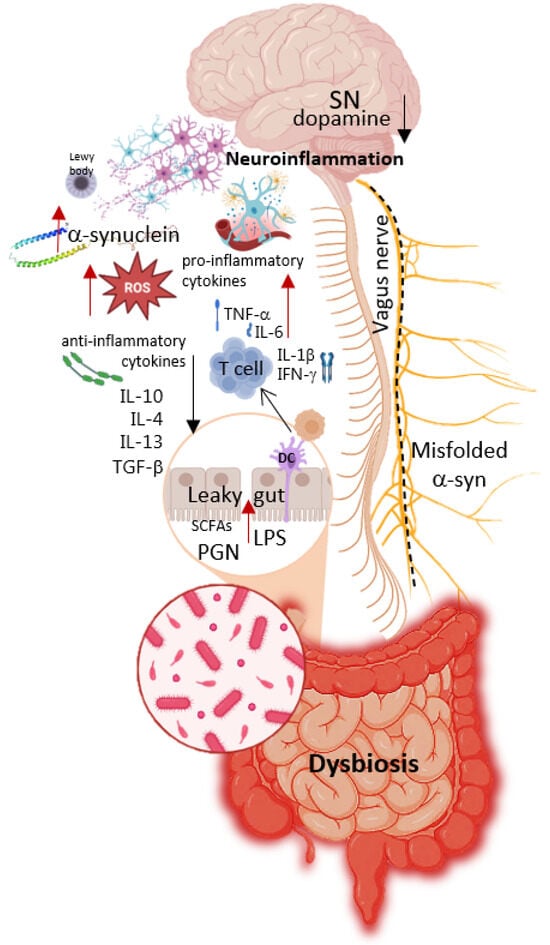
Figure 2.
Relationship between negative changes in microbiome composition (dysbiosis) and subsequent stages of Parkinson’s Disease development. Parts of this figure were created in BioRender (2025) https://BioRender.com/8bysho5 (accessed on 1 July 2025).
PD is also associated with alterations in the structure and metabolic activity of the microbiota. Dysbiosis is common, though not universal—approximately half of patients show no microbial imbalance. Compared with healthy controls, PD patients exhibit differences in beta [62] and alpha [63] diversity, as well as shifts in taxonomic composition, though results vary across studies. At the genus level, the most frequent changes include increases in Christensenella, Oscillospira, Clostridium, Enterococcus, Streptococcus, Lactobacillus, Alistipes, Parabacteroides, Bifidobacterium, Escherichia/Shigella, Bilophila, Desulfovibrio, and Akkermansia, along with decreases in Blautia, Roseburia, Fusicatenibacter, Faecalibacterium, Streptococcus, Prevotella, and Haemophilus [62]. Many studies consistently report reductions in butyrate-producing genera such as Faecalibacterium and Roseburia, together with increased abundances of Akkermansia and Bilophila, as microbial signatures of PD [64]. Although Bifidobacterium, Lactobacillus, and Akkermansia are generally considered beneficial, their elevated levels in PD may reflect confounding factors: Bifidobacterium and Lactobacillus increases are often associated with PD medications, particularly COMT (catechol-O-methyltransferase) inhibitors, while higher Akkermansia levels may relate to aging [62]. Importantly, Akkermansia muciniphila can degrade the intestinal mucus layer, thereby increasing gut permeability and potentially driving inflammation and oxidative stress [65]. Identifying microbiota changes directly linked to PD remains challenging. More robust insights can be obtained through statistical analyses of bacterial groups. For instance, analysis of 35 bacterial groups distinguished individuals with and without PD with an accuracy of just over 84% [64]. The most effective machine-learning approaches for such microbiome analyses are described in Section 5.
At the species level, most studies report that Blautia wexlerae, Faecalibacterium prausnitzii, and Roseburia are depleted in PD patients, whereas Clostridium leptum, Bifidobacterium bifidum, Bifidobacterium dentium, Eisenbergiella tayi, Lactobacillus salivarius, and A. muciniphila are increased. Another noteworthy finding is the higher prevalence of the poorly studied species Ruthenibacterium lactatiformans in PD. This species shares a high degree of genetic similarity with F. prausnitzii, and competition between the two may underlie the observed PD associations [66,67].
Beyond taxonomic composition, the metabolic activity of gut microbiota is of critical importance. PD is associated with a more pro-inflammatory microbiome. The phylum Proteobacteria, which contains many pro-inflammatory taxa, is frequently overabundant in PD [62]. The pro-inflammatory effects are largely attributed to lipopolysaccharides (LPS)—endotoxins that form part of the outer membrane of Gram-negative bacteria [68]. LPS activate TLR4 signaling, leading to increased cytokine production, oxidative stress, and microglial activation. According to the endotoxin hypothesis of PD, gut dysfunction elevates LPS levels in both the gut and bloodstream, thereby promoting α-synuclein aggregation in enteric neurons and triggering peripheral—and subsequently systemic—inflammation [69].
Pro-inflammatory activity may also result from reduced short-chain fatty acids (SCFAs). PD patients often exhibit decreased fecal SCFA levels [63], but elevated plasma SCFAs, with these alterations correlating with disease severity [70]. Collectively, these findings suggest that SCFAs are implicated in both the onset and progression of PD [71]. SCFAs—primarily acetic, propionic, and butyric acids—are produced by bacterial anaerobic fermentation of dietary fiber in the gut. They represent the main energy source for colonocytes, can cross the blood–brain barrier, possess anti-inflammatory properties, and play a direct or indirect role in microbiota–gut–brain axis signaling [72,73]. Many SCFA-producing bacteria (Blautia, Roseburia, Faecalibacterium, Eubacterium, and Prevotella) are consistently depleted in PD, whereas others (Bifidobacterium and Akkermansia) are increased [62,74]. Under certain conditions, however, SCFAs may also modulate signaling cascades that exacerbate PD pathology [71,75]. A comprehensive evaluation of SCFA effects is therefore needed—one that accounts for the diversity of SCFA types, their concentrations, and the host cellular environment—together with an assessment of the “optimal range” of SCFA-producing bacteria.
PD-associated microbes also perform protein fermentation, generating toxic metabolites. One such metabolite is p-cresol, a unique bacterial product not synthesized by human enzymes. Derived from tyrosine and phenylalanine catabolism, p-cresol promotes reactive oxygen species (ROS) production, thereby driving inflammation and multiple physiological abnormalities [76]. Elevated p-cresol metabolites have been detected in the serum of PD patients [77]. Their production has been linked to Blautia obeum, a bacterium associated with PD [59].
Five of the six major categories of carbohydrate-active enzymes (CAZymes) were decreased in PD. Pathway analysis further revealed a marked reduction in genes involved in riboflavin and biotin biosynthesis. Metabolomic profiling of fecal samples also demonstrated significantly lower polyamine levels in PD patients [63].
When assessing microbiota composition in PD, it is essential to account for the fact that patients are invariably receiving medication. Many drugs, including non-antibiotics, can alter the gut microbiota by modifying its composition and activity. In turn, gut microbes may influence drug bioavailability, bioactivity, or toxicity [78]. Levodopa itself appears to exert only minor effects on microbiota composition [79]. By contrast, COMT inhibitors—administered alongside levodopa and carbidopa to prevent levodopa degradation—have a more pronounced impact. Entacapone, a COMT inhibitor, alters microbial activity by chelating and depleting available iron, thereby reshaping gut microbial composition and function [80]. Conversely, certain gut bacteria (Enterococcus and Bifidobacterium) can metabolize levodopa through deamination or decarboxylation, reducing its bioavailability [81,82].
PD patients also frequently present with altered oral health [83] and oral dysbiosis [84], suggesting that oral inflammation may contribute to disease development.
The gut microbiome is increasingly recognized not only as a therapeutic target but also as a source of novel therapeutic agents. The most promising candidates are functional bacteria present in the microbiomes of healthy individuals. Computational algorithms for the discovery of such agents have already been applied to various psychotropic compounds [5,53,85]. The first step in this strategy involves a comprehensive analysis of published datasets to define a reliable reference set of bacterial genes and create catalogs. These catalogs enable the characterization of specific microbiome functions—such as neuromodulatory, immunomodulatory, or antioxidant activities—and facilitate the identification of relevant genes and metabolites.
One promising approach is the use of metagenomic signatures: matrices that include information not only on genes but also on the bacterial species harboring them. This strategy allows researchers to identify and isolate specific strains from the human gut metagenome and characterize them through multi-omics technologies. Genomic, transcriptomic, proteomic, and metabolomic profiling can then be applied to select strains with desirable functional properties from existing probiotic collections. Such strains may subsequently be developed into live biotherapeutic products or pharmabiotics. The ultimate goal is to generate pharmabiotics with defined pharmacological activities that restore gut function, repair damage to other organs and systems (Figure 2), and alleviate Parkinsonian symptoms [86,87].
4. Probiotics and Metabiotics: Existing Potential for the Development of Live Biotherapeutic Products and Pharmabiotics
4.1. Probiotics in Parkinson’s Disease: Analysis of Neuroinflammatory, Immunomodulatory, and Neuromodulatory Activity in Animal Studies
Symbiotic bacteria from the human gut microbiota are widely used to alleviate symptoms and treat various human diseases [88]. A substantial number of studies have explored the potential use of these bacteria as adjuvants in PD therapy. A search in PubMed NCBI using the query “probiotics Parkinson Parkinson’s” returns more than 300 publications from 2020 to April 2025. Most experimental studies have been conducted in animal models, primarily rodents. The findings of many of these studies have been summarized in reviews [89,90,91,92,93,94]. This article focuses primarily on experimental studies conducted between 2024 and April 2025, as well as earlier investigations employing alternative animal models. Regarding clinical studies on probiotic interventions in PD patients, we aimed to include all available reports. Relevant articles were identified through searches in Google Scholar and PubMed using the keywords “probiotic(s) Parkinson’s (Parkinson),” “Bifidobacterium Parkinson’s (Parkinson),” and “Lactobacillus Parkinson’s (Parkinson).” Studies were then selected according to two criteria of interest: (i) probiotic use in animal models of PD, and (ii) probiotic interventions in clinical trials. The overarching goal was to evaluate the mechanisms of bacterial action and assess their potential for clinical application.
Table 1 provides a list and a brief summary of studies from 2024–2025 using bacterial preparations in animal models of PD. Various bacterial species were tested: Lactocaseibacillus (Lactobacillus) rhamnosus, Lactiplantibacillus (Lactobacillus) plantarum, Limosilactobacillus (Lactobacillus) fermentum, Limosilactobacillus (Lactobacillus) reuteri, Lactobacillus delbrueckii, Lactobacillus acidophilus, Ligilactobacillus (Lactobacillus) murinus, Bifidobacterium animalis subsp. lactis, B. bifidum, Bifidobacterium longum, Bifidobacterium breve, Lactococcus lactis, Escherichia coli, Streptococcus thermophilus, Bacillus subtilis, Bacillus coagulans, Clostridium butyricum, and A. muciniphila. Many of these bacteria have been well-characterized in other models (e.g., L. rhamnosus GG, L. plantarum PS128, A. muciniphila Akk11). Both individual strains and multi-component mixtures, as well as pharmaceutical formulations (e.g., VSL#3®, a probiotic used for IBS treatment), were tested.

Table 1.
Investigations of bacterial interventions in Parkinson’s disease animal models (2024–2025).
The biological effects of probiotic strains are generally species- and strain-specific. This is clearly demonstrated in cell culture experiments. For example, 48 bifidobacterial strains were tested on an LPS-induced BV-2 murine microglial cell line (a neuroinflammation model) for NO signaling, Nrf2/HO-1/NQO1 pathway gene expression, and cytokine production. Only one strain, B. longum subsp. longum C7, showed promising protective properties [107]. Testing many strains in animal models is more complex, but strain specificity was also observed in such models [109]. In some cases, probiotic effects were found to be species-specific rather than strain-specific [99]. Genetically engineered strains were also used in some studies [110,111]
The administered bacterial doses ranged from 108 to 1010 CFU, and it was observed that higher doses yielded stronger effects [96,104]. The duration of experiments varied from 2 to 8 weeks. Prolonged administration was considered most effective [112].
Some studies combined bacterial preparations with medications or dietary supplements. L. rhamnosus increased the bioavailability of curcumin [96]. Vinpocetine (a synthetic alkaloid believed to improve cerebral circulation) showed effects similar to the bacterial preparation [97]. Liraglutide (a GLP-1R agonist that protects dopaminergic neurons by reducing inflammation) exhibited neuroprotective and anti-inflammatory effects and regulated gut microbiota dysbiosis, like L. plantarum, but did not affect intestinal transit, unlike the bacterial preparation [101]. The effects of B. longum on several markers were comparable to L-DOPA, and it outperformed L-DOPA in reducing inflammation [107]. When a bacterial mixture was co-administered with L-DOPA–benserazide, the neuroprotective effects of the bacteria did not interfere with L-DOPA therapy; in fact, the pharmacological efficacy of L-DOPA was maintained and, in some cases, improved [103]. Thus, in all studies, bacterial treatments either mirrored or enhanced the effects of conventional drugs.
In addition to variations in bacterial formulations, experimental designs also differed. Some studies used an acute model with a single high dose of a neurotoxin [95,98], while others employed a chronic model with repeated low doses [96,97,103]. Bacterial preparations were administered before the neurotoxin [101,102,105,107], after the neurotoxin [95,100,104], or simultaneously [96,97,98,103]. The experimental design most closely resembling human disease progression was employed by [100], in which 6-OHDA was injected into the SN, followed by a one-month latency period until PD symptoms emerged. Only then were bacterial therapies initiated. Animals used were generally 8–10-week-old mice, though [103] used 1-year-old mice, making the model more representative of the human disease. Most studies used male animals, with female subjects rarely included [104]. When both sexes were used, sex-specific differences in probiotic effects were observed. For instance, L. plantarum improved memory and reduced TNF-α levels in female mice, but these benefits were not seen in males [113].
All the reviewed studies clearly demonstrate the undeniable effect of bacterial preparations in alleviating PD symptoms. However, it is not possible to compare the efficacy of individual preparations due to differences across studies in all of the listed parameters.
Neurotoxins such as MPTP, paraquat, rotenone, and 6-OHDA were used as PD inducers and administered intraperitoneally, subcutaneously, or intranigrally. Neurotoxins are widely used to produce PD in animal models, triggering oxidative stress, neuronal loss, and neuroinflammation. The main signs of PD induced by these toxins are motor deficits caused by the almost complete loss of dopaminergic cells in the SN. Almost all studies conducted behavioral tests and noted that bacteria significantly improved motor deficits: motor performance and balance, movement coordination, and strength [96,97,100,101,102,103,104,105,108]. The positive effect of bacterial preparations was also noted in behavioral tests for cognitive deficits, various aspects of learning, memory, anxiety-like behaviors, and exploratory activity [103,104,107]. Almost all animal model studies of PD indicate that various bacterial preparations can positively affect dopaminergic neuron preservation and prevent neuronal damage in the SN [95,97,98,99,100,101,102,103,104,105,106,107]. An increase in brain dopamine [95,96,97,102,107] and acetylcholinesterase [96,97] levels has also been noted—two major neuromodulators regulating basal ganglia activity.
The root cause of altered dopaminergic neuron activity in PD is the agglomeration of misfolded protein aggregates known as α-synuclein in the brain and the gut. Bacterial preparations are capable of reducing the expression of the α-synuclein gene in the intestine [100], α-synuclein and tau proteins in the striatum [97] and SN [101], and the level of α-synuclein phosphorylation in the SN [108].
Neuroinflammation plays a critical role in PD progression and is mediated and facilitated by various pro-inflammatory cytokines. A growing body of evidence suggests that gut inflammation is the initiator of neuroinflammation. Neurotoxins used to model PD induce not only neuroinflammation but also gut and systemic inflammation. Bacterial preparations reduce inflammatory processes: the expression of pro-inflammatory cytokines decreases while anti-inflammatory cytokines increase in the brain [95,97,98,101,103,104,105,107]. The expression of the C-reactive protein is also reduced [104]. Glial activation decreases [98,101,105,107], and inhibition of the microglial NLRP3 inflammasome is observed [98]. Bacterial preparations restore levels of anti-inflammatory mediators GDNF and BDNF [95,107]. A reduction in inflammation due to bacterial preparations is also evidenced by changes in inflammatory markers in the brain: decreased levels of MDA [96,97,104], nitric oxide synthase [104], ROS [102], and increased antioxidant capacity via SOD, catalase, and GSH expression [96,97,104]. The anti-inflammatory effect of bacterial preparations is observed at both systemic and intestinal levels. They reduce the expression of pro-inflammatory cytokines and increase the expression of anti-inflammatory cytokines in the intestine and colon [95,98,107], the serum [95,103], and the liver [95], and inhibit the intestinal NLRP3 inflammasome [95]. Bacterial interventions improve BBB integrity [101,105,107].
The gut–brain axis plays a crucial role in the development of neurological and neurodegenerative disorders, including PD [1]. Gut-related symptoms are commonly observed in PD. Alterations in the composition of the gut microbiota and intestinal barrier permeability may contribute to the development of PD, at least in some subsets of the disease [67]. Neurotoxins used in animal models of PD, along with other physiological changes, significantly alter the composition and properties of the gut microbiota. The impact on microbiota was investigated in only five of the studies listed in Table 1. The results vary considerably. An improvement in microbiota richness and diversity was observed [101,107]. Both neurotoxin exposure alone and combined with probiotic administration led to changes in microbiota composition compared to control, but these changes did not compensate for each other. Changes induced by probiotics compared to neurotoxins at the phylum level differed between studies: a reduction in Firmicutes [101], an increased Firmicutes/Bacteroidetes ratio [102], and a reduction in Bacteroidetes [105]. At the family level, the following changes were noted: reductions in Prevotellaceae and Desulfovibrio [101]; Alloprevotella, Prevotellaceae Ga6a1 group (associated with anxiety-like behavior in rats), and Lachnospiraceae [103]; Muribaculaceae, Bacteroides, and Prevotellaceae [105]; and increases in Oscillibacter, Lactobacillus, Butyricicoccus (a butyrate producer) [102], Akkermansia, Roseburia, and Dubosiella [105]. While specific microbiota alterations varied, the overall effect was a reduction in pathogenic and pro-inflammatory taxa and the restoration of potentially beneficial bacteria.
The positive effects of bacterial preparations were also evident in the correlation between the relative abundance of specific fecal bacteria and certain traits: Bifidobacterium in feces showed a significant positive correlation with fecal levels of acetic and propionic acids; the relative abundance of the Eubacterium ruminantium group was significantly positively correlated with BDNF and GDNF expression in the striatum and TH-positive cells in the SN; and there was a negative correlation between fecal Eubacterium ruminantium group abundance and IL-6 and TNF-α expression in the striatum [107]. Analyzing bacterial metabolites may be an even more informative indicator. Bacterial intervention significantly increased the levels of beneficial bacterial metabolites. In feces of animals from the bacterial intervention group, compared to the PD group, levels of indole-3-acrylic acid, docosatrienoic acid, and indole-3-butyric acid increased, while levels of N-acetylhistamine, indole-3-acetic acid, and speridine decreased. A significant increase in SCFAs was also observed [107]. The increase in SCFAs in feces following bacterial intervention was confirmed in other studies as well [104,105].
Bacterial intervention also improves gastrointestinal motility [101,102,103,104,105], intestinal barrier integrity [102,104,107], and small intestine histology [103].
Bacterial strains such as B. animalis subsp. lactis NJ241 and L. plantarum SG5 enhance GLP-1 expression in the colon and increase serum GLP-1 levels, as well as GLP-1 receptor (GLP-1R) and PGC-1α expression in the SN of mice. GLP-1 (glucagon-like peptide-1) can promote insulin secretion, regulate gastrointestinal motility, and exert protective effects on the nervous system. The neuroprotective effects of bacterial preparations were attenuated when a GLP-1R antagonist was applied. These data indicate that the action of probiotics is at least partially mediated through the GLP-1/PGC-1α signaling pathway [101,105]. Additionally, genetically engineered bacterial strains delivering GLP-1 have also demonstrated neuroprotective effects in PD animal models [110,111].
Bacterial preparations are administered orally and primarily act on the gut microbiota, altering its composition, reducing the pathogenic component, and changing the spectrum of produced metabolites. The microbiota bacteria synthesize or facilitate the synthesis of metabolites in the host organism, such as neurotransmitters (dopamine, norepinephrine, serotonin, GABA, glutamate, and histamine), exopopolysaccharides, vitamins, antimicrobial peptides, secondary bile acids, and hydrogen sulfide [67]. Bacteria also produce SCFAs through microbial fermentation of dietary fiber in the colon, which are crucial for gastrointestinal and immune health [70]. In the studies reviewed, only some authors linked the activity of bacterial preparations to the production of specific products, specifically suggesting the synthesis of vitamins (riboflavin and folic acid) [103] and exopolysaccharides [102].
In a study of the L. fermentum U-21 strain, which exhibits anti-Parkinsonian activity [108], a 76 kDa protein was found in the cell-free supernatant and extracellular vesicles (EVs) of the strain, which was absent in the supernatant of the L. fermentum 279 strain that lacks such properties. This protein was identified as ClpL, the ATP-binding subunit of the Clp ATP-dependent protease. ClpL belongs to the HSP100 heat shock protein family and interacts with the DnaKJ/GrpE chaperone complex involved in the disaggregation of denatured proteins. In vivo in E. coli cells and in vitro, the ClpL protein demonstrated the ability to refold luciferase proteins. The cell-free culture fluid of the strain also had disaggregating activity [114]. Disaggregases are indeed being considered as potential drug candidates for the treatment of neurodegenerative diseases [115]. It is possible that the antioxidant and anti-Parkinsonian effects of the L. fermentum U-21 strain are partly due to the ClpL protein and its ability to refold protein aggregates.
The possibility of intranasal use of bacterial preparations based on commensal bacteria has also been demonstrated. Compared to the gut microbiota, bacteria of the nasal microbiota—especially those in the olfactory mucosa—are geographically closer to the brain and potentially have a tighter connection with the CNS via metabolite exchange. This offers additional advantages for the treatment of neurodegenerative diseases [116,117]. Recombinant commensal bacteria, particularly lactobacilli, can be used as intranasal drug delivery vehicles, highlighting their potential for brain-targeted therapies [118].
The nematode C. elegans is also used as a model to study the properties of functionally active bacteria [119]. Models of PD have been developed in C. elegans. In the study by Goya et al. [120], it was shown that diverse B. subtilis strains inhibit and clear human α-synuclein aggregation in transgenic C. elegans worms. The bacterium acts through metabolites and biofilm formation to activate protective host pathways, including DAF-16/FOXO and sphingolipid metabolism. In a more detailed study by Francisco and Grau [99], it was shown that B. subtilis strains used as gut commensals in C. elegans prevented the aggregation of GFP-tagged human α-synuclein in worms. To exert this effect, the strains needed to maintain biofilm formation proficiency and the QS peptide CSF. Biofilm-forming B. subtilis also protects against age-related and 6-OHDA-induced dopaminergic neurodegeneration, improves dopamine-dependent behaviors, and extends the lifespan of worms. The protection of C. elegans from 6-OHDA-induced dopaminergic neuron damage occurred not via insulin/IGF-1 signaling, as in the case of α-synuclein aggregation inhibition in the previous study, but through activation of PMK-1/SKN-1 signaling, which are major regulators of genes involved in detoxification and antioxidant defense. In the studies mentioned, the beneficial properties exhibited by Bacillus strains were not strain-specific but were shared by several strains capable of forming biofilms. It is likely that the ability to produce nitric oxide is also important for the probiotic effects of Bacillus. Previously, it was shown that the increased lifespan and stress resistance of C. elegans caused by B. subtilis were due to nitric oxide synthesized by the bacteria via NO synthase [121]. Lactobacilli are poorly suited to serve as gut commensals in C. elegans, but some strains have been found to increase the nematode’s lifespan after exposure to a neurotoxin [122].
All the reviewed studies clearly demonstrate that certain bacterial strains are capable of alleviating or even completely eliminating PD symptoms induced by neurotoxins, acting similarly to pharmaceutical drugs. The application of probiotic bacterial strains in PD therapy represents a highly promising direction. However, several important limitations must be considered. Probiotics are live microorganisms and, in rare cases, may trigger pathological inflammatory processes. This restricts their use in critically ill or immunocompromised patients, as well as in individuals with vascular catheters. Since most PD patients are elderly and often present with multiple comorbidities, these safety concerns are particularly relevant. Certain strains of Lactobacillus and Bifidobacterium produce both L- and D-lactate. The latter can accumulate in the intestine and cause D-lactic acidosis. In rare instances, probiotic intake has been linked to symptoms of D-lactic acidosis, such as brain fog or somnolence. Discontinuation of probiotics, in combination with antibiotic therapy, typically resolves acidosis and alleviates neurological symptoms [123,124]. Careful strain-specific characterization of candidate probiotic preparations is therefore essential. In addition, the development of postbiotics and paraprobiotics offers an important complementary strategy. Postbiotics (also called metabiotics) are metabolic products secreted by probiotics and their complex mixtures, while paraprobiotics are inactivated microbial cells of probiotics or their cellular components [125,126]. There are some data on the use of paraprobiotics in PD models. Heat-killed L. murinus improved 6-OHDA-induced motor impairments and prevented the loss of dopaminergic neurons in the SN, while live L. murinus showed no protective effect. The heat-killed bacteria also reduced NLRP3 inflammasome activation in microglia and the secretion of proinflammatory factors, thereby inhibiting neuroinflammation [127]. Certain bacterial cell components, such as SCFAs, spermine, spermidine, indole, and exopolysaccharides have neuroprotective effects in animal models [128]. EVs are of particular interest as postbiotics.
EVs are nanosized particles released from prokaryotic and eukaryotic cells, enclosed in a lipid bilayer. The cargo of bacterial EVs includes proteins, neurotransmitters, lipids, polysaccharides, phages, DNA, RNA, and microRNAs. Bacterial EVs perform a wide range of functions: exporting misfolded proteins and peptidoglycan fragments from cells; binding and transporting cytosolic metabolites, phages, antibiotics, and bioactive peptides; and transferring DNA. EVs mediate interactions between microbiome bacteria and host cells [129]. Bacterial EVs are absorbed by intestinal epithelial cells and nerve endings, enter the bloodstream, and cross the blood–brain barrier. EVs of Gram-negative pathogenic bacteria can shuttle bacterial toxins and virulence factors throughout the body, triggering inflammatory responses. There is evidence that these EVs may contribute to PD by initiating local, systemic, and neuroinflammation [130]. EVs from probiotic bacteria possess biological activity and are being considered as potential postbiotics [131,132]. Bacterial EVs can serve as vectors for the targeted delivery of specific metabolites and proteins to target cells. There is evidence that EVs from probiotic bacteria suppress neuroinflammation, reduce neuronal damage, and influence serotonin signaling [133]. In a model of Aβ phagocytosis in BV2 microglia, EVs from B. coagulans lilac-01 and E. coli DH5α significantly increased Aβ uptake in a dose-dependent manner [134]. EVs from L. rhamnosus ATCC 7469 reduced lipopolysaccharide-induced neuroinflammation in mice [135]. Lactobacillus-derived EVs have attracted significant attention due to their diverse physiological functions [136]. Biologically active proteins and metabolites have been identified in EVs from the strain L. fermentum U-21 [108], which may account for its biological activity [137,138]. In a study by Sheikh et al., 2025 [100], it was shown for the first time that EVs from the probiotic L. acidophilus PTCC 1643 reduce movement disorders and increase serotonin (5-HT) and TH levels in the SN in a 6-OHDA rat model of PD.
EVs are considered promising therapeutic agents because, unlike live bacteria, they carry a complex of bioactive proteins and metabolites and can be used as vectors for drug delivery.
4.2. Probiotics in Parkinson’s Disease: Human Studies
Animal models of PD have made it possible to identify bacterial strains and compositions with neuroprotective and anti-inflammatory effects, protecting animals from neurotoxins. These studies represent only the first stage in the search for probiotics aimed at alleviating PD symptoms. Due to the characteristics of PD (its long-term progression and unclear origin), there are no animal models that fully replicate the human disease.
Nevertheless, the success in identifying antiparkinsonian effects of bacterial preparations has led to their use in clinical trials. Results from these studies are summarized in Table 2.
Table 2.
Randomized controlled trials of bacterial interventions for the treatment of Parkinson’s disease.
The reviewed studies used various bacterial species: S. thermophilus, Enterococcus faecium, B. coagulans, and various Lactobacillus strains (L. rhamnosus, L. acidophilus, L. plantarum, L. paracasei, L. casei, L. delbrueckii, L. reuteri, and L. fermentum), as well as Bifidobacteria (B. breve, B. animalis, B. infantis, B. bifidum, and B. longum). It is not merely the species but the specific strain of bacteria that determines their effects on human health. In some studies, unfortunately, the strains were not specified. However, most studies did report the strain identity. Many of the strains used had already proven effective in previous studies unrelated to PD, such as L. paracasei strain Shirota [139,148], B. animalis subsp. lactis BS01, B. longum 03, B. adolescentis BA02 [152], L. plantarum PS128 [145,155], and B. animalis subsp. lactis Probio-M8 [147]. Commercial probiotic formulations were also used, such as Hexbio® (B-Crobes Laboratory, Malaysia) [143], Symprove (Symprove Ltd.) [144], Enterolactis Duo L. paracasei DG (DSM 34154) [151], Comflor® (Fara Daroo Fanavar Mehr Co) [150], BioZen D [153], and Livia® (Pharma Zad, Egypt) [154]. Prebiotics such as inulin, fructooligosaccharides, and vitamins D and B-group were also used. It is worth noting that the L. paracasei (Shirota) strain was successfully tested in two studies, and L. plantarum PS128 showed positive results both in clinical trials and animal experiments. The typical duration of probiotic administration was 3 months, with high daily doses (109–1010 CFU).
The conducted studies were predominantly randomized, double-blind, placebo-controlled clinical trials (RCTs). The number of participants ranged from 25 to 128, with average participant age between 56 and 76 years. All studies included both men and women. The probiotic treatments lasted between 4 and 12 weeks, most commonly 12 weeks. Probiotics were administered in the form of fermented milk, capsules, or powder sachets. In all studies, probiotics were used alongside standard medications (levodopa, dopamine agonists, and MAO-B inhibitors).
The functional bacteria were generally well tolerated by patients, with rare and mild adverse effects. For instance, Ibrahim et al. [43] reported two cases of abdominal symptoms and two of dizziness; Tan et al. [146] mentioned one case of lethargy after probiotic treatment.
All studies reported improvements in PD patients’ health parameters, though the nature of these improvements varied. Regarding patient condition and behavior, general improvement in quality of life [145,149,153] prolonged ON periods, and reduced OFF periods [145] were noted.
Constipation is a predominant NMS of PD, often appearing years before motor symptoms. Its pathophysiology involves multiple factors: PD-related GI dysfunction, side effects of antiparkinsonian medications, sedentary lifestyle, reduced intestinal motility, and impaired anal sphincter function. Probiotics have proven effective in treating constipation in non-PD adult patients [156]. In PD-related probiotic studies, the following improvements were observed: improved stool frequency [140,143,146,147,148,150,151], improved whole gut transit time [145], and improved stool consistency [139,147,148,150,151]. These findings are supported by reduced laxative use [140]. However, some studies did not show statistically significant improvements in constipation symptoms [141,152].
An analysis of data from a single patient with recently diagnosed PD, who began a diet enriched with plant-based foods and dietary fiber, along with probiotic supplementation six months after diagnosis, showed a rapid disappearance of constipation symptoms. It is possible that the use of probiotics is most effective in the early stages of the disease [157].
Probiotic intake also decreases gastrointestinal problems such as bloating and abdominal pain [139,141,151,152,153]. The effects of probiotics on the gastrointestinal tract in PD patients may be multifaceted, including general improvements in well-being and possibly influencing the formation of synuclein aggregates. Furthermore, by promoting intestinal transit, probiotics may enhance the absorption of PD therapy, thereby increasing its clinical effectiveness.
Many studies have also noted improvements in other NMS. These include enhanced cognitive function [147,151,152], reductions in alexithymia and dysautonomia [151], improved sleep quality [147], and improvements in mental health parameters such as depression, anxiety, and panic disorder [147,148,149,151,153]. The effects of different probiotic strains on depression and anxiety symptoms have been studied previously [158,159].
Some studies report improvements in motor function, UPDRS-III scores, and akinesia [145,147,152]. However, other studies did not find improvements in motor symptoms with probiotic use [143,150,151].
Biochemical studies have also been conducted. Investigations among PD patients demonstrated a reduced endogenous antioxidant system [35]. In the studies reviewed, probiotic intake increased total antioxidant capacity (TAC) [149,153], reduced oxidative stress index [149], and improved markers of oxidative stress in serum—lowering levels of malondialdehyde [149,153,154] and myeloperoxidase [145]. In the study by Borzabadi et al. [142], however, no change in GSH levels was observed with probiotic intake.
Neuroinflammation is one of the key factors in PD pathogenesis, along with alpha-synuclein accumulation, synaptic dysfunction, and dopaminergic neuronal loss. Aggregated alpha-synuclein binds to Toll-like receptors and activates microglia, initiating the production and secretion of pro-inflammatory cytokines [160]. Regulating inflammatory markers may reduce central inflammation and help restore nervous system functions. Probiotic intake may reduce gene expression levels of inflammatory markers [161]. Studies have reported downregulation of pro-inflammatory cytokines (IL-1β, IL-8, IFN-γ, TNF-α) in peripheral blood mononuclear cells [142] and serum [152,153,154], as well as upregulation of the anti-inflammatory cytokine IL-10 in serum [153].
Probiotics may also affect glycemic status and insulin sensitivity. In animal studies, insulin resistance increased after probiotic use [162]. In clinical trials, probiotics reduced blood glucose levels [163]. Among the studies reviewed, only one investigated this aspect. These reported upregulation of peroxisome proliferator-activated receptor gamma in peripheral blood mononuclear cells [142]. This receptor, which acts as a transcription factor, is involved in fatty acid storage and glucose metabolism [164].
Other serum markers also changed—there was an increase in acetic acid and dopamine and a decrease in glutamine and tryptophan [147]. Yang et al. noted an increase in serum tyrosine and a decrease in fecal tyrosine [148]. Probiotics may improve impaired digestion and absorption of L-tyrosine in the upper digestive tract, increasing its level in the serum. A statistically significant increase in the butyrate/acetate ratio in fecal samples was also observed after synbiotic intake [151].
The primary action of probiotics is on the gut microbiota, and understanding how it changes in response to probiotics is essential. Unfortunately, only four of the reviewed studies examined microbiota composition. These studies reported that probiotic supplementation did not cause drastic changes in gut microbiota diversity or structure [147,148], though it did alter the microbiota composition. These changes likely occur rapidly: in an in vitro gut model using PD patient microbiota, changes in microbial composition and an increase in SCFAs and lactate production were observed within 48 h of probiotic addition [144]. The specific changes varied across studies: suppression of L. fermentum and potentially pathogenic taxa (Klebsiella) and increases in beneficial taxa (Ruminococcaceae, Lachnospira, Butyricimonas) [147]; increased Lacticaseibacillus [148]; and increases in Oscillospiraceae, particularly F. prausnitzii, a major butyrate producer, along with an increase in the butyrate/acetate ratio in fecal samples [151]. In the same in vitro model, probiotic dosing improved colonic media tight junction integrity. The probiotic itself did not directly affect tight junctions—rather, the effect was mediated by the altered microbiota and increased SCFA levels [144].
A number of studies have conducted systematic reviews and meta-analyses of existing research on the effects of probiotics in patients with PD. These analyses include various publications, and the conclusions drawn by authors are not always consistent. Overall, such studies confirm the effectiveness of probiotics in improving PD-related symptoms such as constipation [156,165,166,167,168], UPDRS-III motor scores [166,169], and depression [169]. These studies also highlight that the efficacy of probiotics appears to be greater when administered in capsule form compared to fermented milk [156,169].
Although early clinical trials investigating bacterial preparations in PD patients have yielded encouraging results, further research is essential. It should be noted that the number of available clinical studies remains limited; most involve small patient cohorts and report heterogeneous outcomes. Large-scale trials with well-defined, homogeneous patient groups are urgently needed. Early intervention and long-term administration of bacterial products may prove especially beneficial in slowing disease progression and alleviating symptoms. In particular, detailed investigation of bacterial formulations and the development of live biotherapeutic products (LBPs) based on them is necessary. This includes identifying suitable PD models, exploring novel probiotics and postbiotics—especially bacterial vesicles—and searching for early PD biomarkers [170,171,172].
Table 3 provides examples of bacterial preparations aimed at treating neurodegenerative diseases, including PD.
Table 3.
Developers and manufacturers of LBPs aimed at the treatment of Parkinson’s disease and other neurodegenerative disorders.
An important but underexplored microbial intervention for PD is fecal microbiota transplantation (FMT). FMT involves the transfer of fecal material from a healthy donor into the gastrointestinal tract of a patient via colonoscopy, gastroscopy through a nasojejunal tube, or oral capsules containing the microbiota. Unlike probiotics, FMT delivers a complex microbial community rather than a single strain. FMT has proven highly effective in treating recurrent Clostridium difficile infections [177] and is occasionally used in other conditions linked to gut dysbiosis. In rodent models of PD, FMT was shown to correct dysbiosis, reduce inflammation in the gut and brain, and alleviate disease symptoms [178,179]. To date, four randomized controlled trials have been published examining the effects of FMT in PD patients [180,181,182,183]. The study methodologies and outcomes varied, but in general, mild-to-moderate adverse events were transient and gastrointestinal in nature. Modest but statistically significant improvements were observed in gut motility/constipation, motor and NMS, and overall PD severity scores [67,184]. However, such studies are extremely difficult to standardize. Outcomes depend heavily on the microbiota profiles of both patients and donors, the method and formulation of FMT administration, and the quantity of transplanted material. Additionally, the potential risk of serious infections associated with FMT should not be underestimated.
5. Neurodegenerative Diseases—Parkinson’s Disease—The Potential of Machine Learning for Diagnosis and Treatment
In recent years, artificial intelligence (AI) technologies—particularly neural networks and machine learning—have entered nearly all areas of science, including biomedicine. Disorders of the nervous system, such as PD, have increasingly drawn the attention of AI researchers. Several publications have applied machine learning approaches to classify different forms of PD, and attempts have been made to refine biomarkers specific to PD [185]. Other studies have proposed machine learning–based methods to support PD diagnosis [186].
A vast amount of information generated from gut microbiome studies and digitized in databases cannot be effectively analyzed without the use of AI technologies (Figure 3). The integration of omics approaches, including metagenomic sequencing, with modern machine learning methods for studying the microbiota of both healthy individuals and patients with neurological disorders provides unique insights into the functional characteristics of microorganisms. In this context, machine learning can serve as an in silico diagnostic tool, as well as a means of identifying key microbiota biomarkers and therapeutic targets. Machine learning methods are increasingly applied to microbiota data to predict the risk of developing neurodegenerative diseases [187]. Distinct microbiota profiles have already been identified in patients with PD [188,189]. Several studies have reported the application of machine learning to predictive modeling of microbiome biomarkers (at the bacterial genus level) based on comparative analyses of cohorts of PD patients and healthy individuals [64,190].
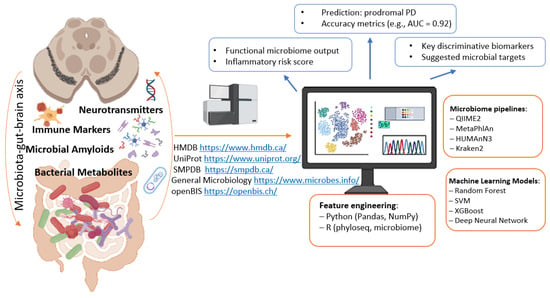
Figure 3.
Digital parameters of the microbiome—indicators for intervention. Parts of this figure were created in BioRender (2025) https://BioRender.com/ocetej3 (accessed on 1 July 2025).
Research combining neural networks and machine learning in the context of PD appears promising in two main directions: first, in the development and analysis of neural networks that integrate and model the functional architecture of PD; a schematic overview of relevant biomarkers and algorithms for constructing such networks is presented in Figure 4, with methodological approaches described in [191]; second, machine learning can be applied to mine gut microbiome databases for pharmabiotics and their metabolites with targeted antiparkinsonian effects—similar to how such approaches have already been employed to identify microbiome-derived pharmabiotics with antidepressant properties [158,192,193].
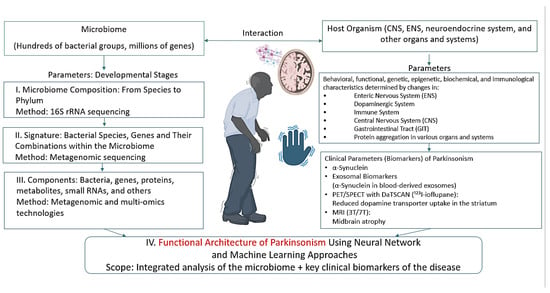
Figure 4.
Parameters characterizing the microbiome–host organism system in PD. Parts of this figure were created in BioRender (2025) https://BioRender.com/vs5aep9 (accessed on 1 July 2025).
6. Conclusions
PD is a complex, multifactorial disease, both in terms of its clinical manifestations and the range of organs and systems it affects during development and progression. Currently, no drugs are available that can cure the disease. Medications used in clinical practice generally target only one of the systems affected. Drug development strategies evolve with advances in chemistry, biochemistry, genetics, molecular biology, systems biology, bioinformatics, mathematical modeling, AI (including machine learning), and related fields.
Over the past decades, target-specific drugs have been developed—sometimes successfully—through chemical synthesis or by isolation from animal and plant sources, soil bacteria, and other origins. PD, like other neurodegenerative disorders, is characterized by dysfunction across multiple biological targets within the networks that maintain overall homeostasis. Based on this understanding, effective drugs for PD should ideally consist of combinations of pharmacologically active substances in precise compositions and ratios.
The biological targets of these drugs are already partially known and are expected to be further refined. Insights into the mechanisms underlying PD and the affected targets may also be gained from studies on post-COVID PD [194,195]. A promising source of such drugs is the microbiota of healthy individuals—a stably balanced system that supports homeostasis, even under stress.
Which microbiota-based bacteria are suitable for developing drugs targeting PD mechanisms? These should be functional bacteria-possessing genes and their products in combinations that match the biological targets they are intended to influence. This is supported, in part, by the demonstrated ability of certain probiotics to exhibit properties relevant to PD therapy. The next step involves selecting LBPs based on these strains, which are considered medicinal products under modern standards [196]. Postbiotics and vesicle-based preparations may also be promising therapeutic options.
LBPs and their derived vesicles can serve as carriers for proteins, low-molecular-weight metabolites, neurotransmitters, immunomodulators, anti-inflammatory agents, and small RNAs capable of modulating gene and receptor activity, including in neurons and immune cells. These interventions may, for example, promote the production of antibodies against misfolded proteins such as α-synuclein.
The human microbiome represents a vast resource for developing nature-inspired medicinal products for PD therapy. Such drugs have the potential to eliminate pathogenic gut bacteria, restore intestinal barrier integrity, recover bacterial communities that produce neurotransmitters, neurohormones, and their precursors, halt α-synuclein formation in both the enteric and central nervous systems, neutralize free radicals and reduce oxidative stress, and alleviate associated depressive symptoms and cognitive dysfunctions.
Author Contributions
Conceptualization, S.I. and V.D.; validation, S.I., V.D. and E.U.P.; data curation, E.U.P., M.M. and A.P.; writing—original draft preparation, V.D., E.U.P., A.S., A.P., R.Y., M.M. and T.K.; writing—review and editing, S.I., V.D., E.U.P. and R.Y.; project administration, T.K. All authors have read and agreed to the published version of the manuscript.
Funding
The study was supported by a grant from the Ministry of Science and Higher Education of the Russian Federation for major scientific projects in priority areas of scientific and technological development (No. 075-15-2024-638).
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in the study are included in the article; further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| 6-OHDA | 6-hydroxydopamine |
| BBB | Blood-Brain Barrier |
| CFU | Colony Forming Units |
| CNS | Central Nervous System |
| COMT | Catechol-O-methyltransferase |
| ENS | Enteric Nervous System |
| EVs | Extracellular Vesicles |
| FMT | Fecal Microbiota Transplantation |
| GABA | Gamma-Aminobutyric Acid |
| GSH | Reduced Glutathione |
| IL-6 | Interleukin-6 |
| INF-γ | Interferon-gamma |
| LBPs | Live Biotherapeutic Products |
| LPS | Lipopolysaccharides |
| MPTP | 1-methyl-4-phenyl-1:2,3,6-tetrahydropyridine |
| NMS | Non-Motor Symptoms |
| PD | Parkinson’s Disease |
| QS | Quorum Sensing |
| ROS | Reactive Oxygen Species |
| SCFAs | Short-Chain Fatty Acids |
| SN | Substantia Nigra |
| SOD | Superoxide Dismutase |
| TH | Tyrosine Hydroxylase |
| TNF-α | Tumor Necrosis Factor-alpha |
References
- Morais, L.H.; Schreiber, H.L., 4th; Mazmanian, S.K. The gut microbiota-brain axis in behaviour and brain disorders. Nat. Rev. Microbiol. 2021, 19, 241–255. [Google Scholar] [CrossRef]
- Houser, M.C.; Tansey, M.G. The gut-brain axis: Is intestinal inflammation a silent driver of Parkinson’s disease pathogenesis? npj Park. Dis. 2017, 3, 3. [Google Scholar] [CrossRef]
- Goedert, M.; Spillantini, M.G.; Del Tredici, K.; Braak, H. 100 years of Lewy pathology. Nat. Rev. Neurol. 2013, 9, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Cryan, J.F.; O’Riordan, K.J.; Cowan, C.S.M.; Sandhu, K.V.; Bastiaanssen, T.F.S.; Boehme, M.; Codagnone, M.G.; Cussotto, S.; Fulling, C.; Golubev, A.V.; et al. The Microbiota-Gut-Brain Axis. Physiol. Rev. 2019, 99, 1877–2013. [Google Scholar] [CrossRef] [PubMed]
- Valles-Colomer, M.; Falony, G.; Darzi, Y.; Tigchelaar, E.F.; Wang, J.; Tito, R.Y.; Schiweck, C.; Kurilshikov, A.; Joossens, M.; Wijmenga, C.; et al. The neuroactive potential of the human gut microbiota in quality of life and depression. Nat. Microbiol. 2019, 4, 623–632. [Google Scholar] [CrossRef] [PubMed]
- Braak, H.; de Vos, R.A.; Bohl, J.; Del Tredici, K. Gastric alpha-synuclein immunoreactive inclusions in Meissner’s and Auerbach’s plexuses in cases staged for Parkinson’s disease-related brain pathology. Neurosci. Lett. 2006, 396, 67–72. [Google Scholar] [CrossRef]
- Kim, S.; Kwon, S.H.; Kam, T.I.; Panicker, N.; Karuppagounder, S.S.; Lee, S.; Lee, J.H.; Kim, W.R.; Kook, M.; Foss, C.A.; et al. Transneuronal Propagation of Pathologic α-Synuclein from the Gut to the Brain Models Parkinson’s Disease. Neuron 2019, 103, 627–641.e7. [Google Scholar] [CrossRef]
- Huh, E.; Choi, J.G.; Choi, Y.; Ju, I.G.; Kim, B.; Shin, Y.J.; An, J.M.; Park, M.G.; Yim, S.V.; Chung, S.J.; et al. P. mirabilis-derived pore-forming haemolysin, HpmA drives intestinal alpha-synuclein aggregation in a mouse model of neurodegeneration. EBioMedicine 2023, 98, 104887. [Google Scholar] [CrossRef]
- Xu, Y.; Xie, M.; Xue, J.; Xiang, L.; Li, Y.; Xiao, J.; Xiao, G.; Wang, H.L. EGCG ameliorates neuronal and behavioral defects by remodeling gut microbiota and TotM expression in Drosophila models of Parkinson’s disease. FASEB J. 2020, 34, 5931–5950. [Google Scholar] [CrossRef]
- He, Y.; Zhao, J.; Ma, Y.; Yan, X.; Duan, Y.; Zhang, X.; Dong, H.; Fang, R.; Zhang, Y.; Li, Q.; et al. Citrobacter rodentium infection impairs dopamine metabolism and exacerbates the pathology of Parkinson’s disease in mice. J. Neuroinflamm. 2024, 21, 153. [Google Scholar] [CrossRef]
- Hasegawa, M.; Nonaka, T.; Masuda-Suzukake, M. Prion-like mechanisms and potential therapeutic targets in neurodegenerative disorders. Pharmacol. Ther. 2017, 172, 22–33. [Google Scholar] [CrossRef]
- Alfaidi, M.; Barker, R.A.; Kuan, W.L. An update on immune-based alpha-synuclein trials in Parkinson’s disease. J. Neurol. 2024, 272, 21. [Google Scholar] [CrossRef]
- Haas-Neill, S.; Forsythe, P. A Budding Relationship: Bacterial Extracellular Vesicles in the Microbiota-Gut-Brain Axis. Int. J. Mol. Sci. 2020, 21, 8899. [Google Scholar] [CrossRef] [PubMed]
- Bleibel, L.; Dziomba, S.; Waleron, K.F.; Kowalczyk, E.; Karbownik, M.S. Deciphering psychobiotics’ mechanism of action: Bacterial extracellular vesicles in the spotlight. Front. Microbiol. 2023, 14, 1211447. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Yin, Y.; Zhou, X. Insights into Microbiota-Host Crosstalk in the Intestinal Diseases Mediated by Extracellular Vesicles and Their Encapsulated MicroRNAs. Int. J. Mol. Sci. 2024, 25, 13001. [Google Scholar] [CrossRef] [PubMed]
- Illarioshkin, S.N. Modern concepts of Parkinson’s disease etiology. Nevrol. Zhurnal 2015, 20, 4–13. (In Russian) [Google Scholar] [CrossRef]
- Xiao, B.; Zhou, Z.; Chao, Y.; Tan, E.K. Pathogenesis of Parkinson’s Disease. Neurol. Clin. 2025, 43, 185–207. [Google Scholar] [CrossRef]
- Salmina, A.B.; Kapkaeva, M.R.; Vetchinova, A.S.; Illarioshkin, S.N. Novel Approaches Used to Examine and Control Neurogenesis in Parkinson’s Disease. Int. J. Mol. Sci. 2021, 22, 9608. [Google Scholar] [CrossRef]
- Chen, H.; Li, J.; Huang, Z.; Fan, X.; Wang, X.; Chen, X.; Guo, H.; Liu, H.; Li, S.; Yu, S.; et al. Dopaminergic system and neurons: Role in multiple neurological diseases. Neuropharmacology 2024, 260, 110133. [Google Scholar] [CrossRef]
- Chen, Z.; Li, G.; Liu, J. Autonomic dysfunction in Parkinson’s disease: Implications for pathophysiology, diagnosis, and treatment. Neurobiol. Dis. 2020, 134, 104700. [Google Scholar] [CrossRef]
- Westenberger, A.; Brüggemann, N.; Klein, C. Genetics of Parkinson’s Disease: From Causes to Treatment. Cold Spring Harb. Perspect. Med. 2025, 15, a041774. [Google Scholar] [CrossRef]
- Ascherio, A.; Schwarzschild, M.A. The epidemiology of Parkinson’s disease: Risk factors and prevention. Lancet Neurol. 2016, 15, 1257–1272. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, M.J.; Okun, M.S. Diagnosis and Treatment of Parkinson Disease: A Review. JAMA 2020, 323, 548–560. [Google Scholar] [CrossRef] [PubMed]
- Jankovic, J. Parkinson’s disease: Clinical features and diagnosis. J. Neurol. Neurosurg. Psychiatry 2008, 79, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Marinus, J.; Zhu, K.; Marras, C.; Aarsland, D.; van Hilten, J.J. Risk factors for non-motor symptoms in Parkinson’s disease. Lancet Neurol. 2018, 17, 559–568. [Google Scholar] [CrossRef]
- Fullard, M.E.; Morley, J.F.; Duda, J.E. Olfactory Dysfunction as an Early Biomarker in Parkinson’s Disease. Neurosci. Bull. 2017, 33, 515–525. [Google Scholar] [CrossRef]
- Stefani, A.; Högl, B. Sleep in Parkinson’s disease. Neuropsychopharmacology 2020, 45, 121–128. [Google Scholar] [CrossRef]
- Weintraub, D.; Aarsland, D.; Chaudhuri, K.R.; Dobkin, R.D.; Leentjens, A.F.; Rodriguez-Violante, M.; Schrag, A. The neuropsychiatry of Parkinson’s disease: Advances and challenges. Lancet Neurol. 2022, 21, 89–102. [Google Scholar] [CrossRef]
- Kawahata, I.; Fukunaga, K. Degradation of Tyrosine Hydroxylase by the Ubiquitin-Proteasome System in the Pathogenesis of Parkinson’s Disease and Dopa-Responsive Dystonia. Int. J. Mol. Sci. 2020, 21, 3779. [Google Scholar] [CrossRef]
- Rausch, W.D.; Wang, F.; Radad, K. From the tyrosine hydroxylase hypothesis of Parkinson’s disease to modern strategies: A short historical overview. J. Neural. Transm. 2022, 129, 487–495. [Google Scholar] [CrossRef]
- Segura-Aguilar, J.; Paris, I.; Muñoz, P.; Ferrari, E.; Zecca, L.; Zucca, F.A. Protective and toxic roles of dopamine in Parkinson’s disease. J. Neurochem. 2014, 129, 898–915. [Google Scholar] [CrossRef]
- Calabresi, P.; Di Lazzaro, G.; Marino, G.; Campanelli, F.; Ghiglieri, V. Advances in understanding the function of alpha-synuclein: Implications for Parkinson’s disease. Brain 2023, 146, 3587–3597. [Google Scholar] [CrossRef]
- Wakabayashi, K.; Tanji, K.; Odagiri, S.; Miki, Y.; Mori, F.; Takahashi, H. The Lewy body in Parkinson’s disease and related neurodegenerative disorders. Mol. Neurobiol. 2013, 47, 495–508. [Google Scholar] [CrossRef]
- Atik, A.; Stewart, T.; Zhang, J. Alpha-Synuclein as a Biomarker for Parkinson’s Disease. Brain Pathol. 2016, 26, 410–418. [Google Scholar] [CrossRef] [PubMed]
- Fedorova, T.N.; Logvinenko, A.A.; Poleshchuk, V.V.; Illarioshkin, S.N. The state of systemic oxidative stress during Parkinson’s disease. Neurochem. J. 2017, 11, 340–345. [Google Scholar] [CrossRef]
- Caproni, S.; Di Fonzo, A.; Colosimo, C. Oxidative Stress: A New Pathophysiological Pathway in Parkinson’s Disease and a Potential Target of the Brain-Sport Crosstalk. Park. Dis. 2025, 2025, 6691390. [Google Scholar] [CrossRef] [PubMed]
- Mahoney-Sánchez, L.; Bouchaoui, H.; Ayton, S.; Devos, D.; Duce, J.A.; Devedjian, J.C. Ferroptosis and its potential role in the physiopathology of Parkinson’s Disease. Prog. Neurobiol. 2021, 196, 101890. [Google Scholar] [CrossRef]
- Chen, Y.; Luo, X.; Yin, Y.; Thomas, E.R.; Liu, K.; Wang, W.; Li, X. The interplay of iron, oxidative stress, and α-synuclein in Parkinson’s disease progression. Mol. Med. 2025, 31, 154. [Google Scholar] [CrossRef]
- Pajares, M.; Rojo, A.I.; Manda, G.; Boscá, L.; Cuadrado, A. Inflammation in Parkinson’s Disease: Mechanisms and Therapeutic Implications. Cells 2020, 9, 1687. [Google Scholar] [CrossRef]
- Wang, C.; Yang, T.; Liang, M.; Xie, J.; Song, N. Astrocyte dysfunction in Parkinson’s disease: From the perspectives of transmitted α-synuclein and genetic modulation. Transl. Neurodegener. 2021, 10, 39. [Google Scholar] [CrossRef]
- Isik, S.; Yeman Kiyak, B.; Akbayir, R.; Seyhali, R.; Arpaci, T. Microglia Mediated Neuroinflammation in Parkinson’s Disease. Cells 2023, 127, 1012. [Google Scholar] [CrossRef]
- Khan, E.; Hasan, I.; Haque, M.E. Parkinson’s Disease: Exploring Different Animal Model Systems. Int. J. Mol. Sci. 2023, 24, 9088. [Google Scholar] [CrossRef]
- Okyere, S.K.; Zeng, C.; Yue, D.; Hu, Y. Neurotoxic Mechanism and Shortcomings of MPTP, 6-OHDA, Rotenone and Paraquat-induced Parkinson’s Disease Animal Models. Venoms Toxins 2021, 1, 27–40. [Google Scholar] [CrossRef]
- Ungerstedt, U. 6-Hydroxy-dopamine induced degeneration of central monoamine neurons. Eur. J. Pharmacol. 1968, 5, 107–110. [Google Scholar] [CrossRef] [PubMed]
- Simola, N.; Morelli, M.; Carta, A.R. The 6-hydroxydopamine model of Parkinson’s disease. Neurotox. Res. 2007, 11, 151–167. [Google Scholar] [CrossRef] [PubMed]
- Masini, D.; Plewnia, C.; Bertho, M.; Scalbert, N.; Caggiano, V.; Fisone, G. A Guide to the Generation of a 6-Hydroxydopamine Mouse Model of Parkinson’s Disease for the Study of Non-Motor Symptoms. Biomedicines 2021, 9, 598. [Google Scholar] [CrossRef]
- Mondal, S.; Firdous, S.M. Unrevealing the molecular mechanisms of MPTP-induced Parkinson’s in experimental animals. Med. Chem. Res. 2025. [Google Scholar] [CrossRef]
- Jackson-Lewis, V.; Przedborski, S. Protocol for the MPTP mouse model of Parkinson’s disease. Nat. Protoc. 2007, 2, 141–151. [Google Scholar] [CrossRef]
- Dinis-Oliveira, R.J.; Remião, F.; Carmo, H.; Duarte, J.A.; Navarro, A.S.; Bastos, M.L.; Carvalho, F. Paraquat exposure as an etiological factor of Parkinson’s disease. Neurotoxicology 2006, 27, 1110–1122. [Google Scholar] [CrossRef]
- Johnson, M.E.; Bobrovskaya, L. An update on the rotenone models of Parkinson’s disease: Their ability to reproduce the features of clinical disease and model gene-environment interactions. Neurotoxicology 2015, 46, 101–116. [Google Scholar] [CrossRef]
- Sharma, P.; Mittal, P. Paraquat (herbicide) as a cause of Parkinson’s Disease. Park. Relat. Disord. 2024, 119, 105932. [Google Scholar] [CrossRef]
- Kovtun, A.S.; Averina, O.V.; Zakharevich, N.V.; Kasianov, A.S.; Danilenko, V.N. In silico identification of metagenomic signature describing neurometabolic potential of normal human gut microbiota. Russ. J. Genet. 2018, 54, 1101–1110. [Google Scholar] [CrossRef]
- Averina, O.V.; Poluektova, E.U.; Marsova, M.V.; Danilenko, V.N. Biomarkers and Utility of the Antioxidant Potential of Probiotic Lactobacilli and Bifidobacteria as Representatives of the Human Gut Microbiota. Biomedicines 2021, 9, 1340. [Google Scholar] [CrossRef] [PubMed]
- Joos, R.; Boucher, K.; Lavelle, A.; Arumugam, M.; Blaser, M.J.; Claesson, M.J.; Clarke, G.; Cotter, P.D.; De Sordi, L.; Dominguez-Bello, M.G.; et al. Examining the healthy human microbiome concept. Nat. Rev. Microbiol. 2025, 23, 192–205, Erratum in Nat. Rev. Microbiol. 2025, 23, 206. https://doi.org/10.1038/s41579-024-01145-8. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Wang, K.; Su, N.; Yuan, C.; Zhang, N.; Hu, X.; Fu, Y.; Zhao, F. Microbiota-gut-brain axis in health and neurological disease: Interactions between gut microbiota and the nervous system. J. Cell. Mol. Med. 2024, 28, e70099. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Shao, Y.; Hu, Y.; Qian, J.; Wang, P.; Tian, L.; Ni, Y.; Li, S.; Al-Nusaif, M.; Liu, C.; et al. Fecal microbiota from patients with Parkinson’s disease intensifies inflammation and neurodegeneration in A53T mice. CNS Neurosci. Ther. 2024, 30, e70003. [Google Scholar] [CrossRef]
- Borghammer, P.; Van Den Berge, N. Brain-first versus gut-first Parkinson’s disease: A hypothesis. J. Park. Dis. 2019, 9, S281–S295. [Google Scholar] [CrossRef]
- Horsager, J.; Borghammer, P. Brain-first vs. body-first Parkinson’s disease: An update on recent evidence. Park. Relat. Disord. 2024, 122, 106101. [Google Scholar] [CrossRef]
- Metcalfe-Roach, A.; Cirstea, M.S.; Yu, A.C.; Ramay, H.R.; Coker, O.; Boroomand, S.; Kharazyan, F.; Martino, D.; Sycuro, L.K.; Appel-Cresswell, S.; et al. Metagenomic analysis reveals large-scale disruptions of the gut microbiome in Parkinson’s disease. Mov. Disord. 2024, 39, 1740–1751. [Google Scholar] [CrossRef]
- Suresh, S.B.; Malireddi, A.; Abera, M.; Noor, K.; Ansar, M.; Boddeti, S.; Nath, T.S. Gut Microbiome and Its Role in Parkinson’s Disease. Cureus 2024, 16, e73150. [Google Scholar] [CrossRef]
- Gallop, A.; Weagley, J.; Paracha, S.U.; Grossberg, G. The Role of The Gut Microbiome in Parkinson’s Disease. J. Geriatr. Psychiatry Neurol. 2021, 34, 253–262. [Google Scholar] [CrossRef]
- Li, Z.; Liang, H.; Hu, Y.; Lu, L.; Zheng, C.; Fan, Y.; Wu, B.; Zou, T.; Luo, X.; Zhang, X.; et al. Gut bacterial profiles in Parkinson’s disease: A systematic review. CNS Neurosci. Ther. 2023, 29, 140–157. [Google Scholar] [CrossRef]
- Nishiwaki, H.; Ueyama, J.; Ito, M.; Hamaguchi, T.; Takimoto, K.; Maeda, T.; Kashihara, K.; Tsuboi, Y.; Mori, H.; Kurokawa, K.; et al. Meta-analysis of shotgun sequencing of gut microbiota in Parkinson’s disease. npj Park. Dis. 2024, 10, 106. [Google Scholar] [CrossRef]
- Zhao, Z.; Chen, J.; Zhao, D.; Chen, B.; Wang, Q.; Li, Y.; Chen, J.; Bai, C.; Guo, X.; Hu, N.; et al. Microbial biomarker discovery in Parkinson’s disease through a network-based approach. npj Park. Dis. 2024, 10, 203. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.J.; Yang, J.Y.; Yan, Z.H.; Hu, S.; Li, J.Q.; Xu, Z.X.; Jian, Y.P. Recent findings in Akkermansia muciniphila-regulated metabolism and its role in intestinal diseases. Clin. Nutr. 2022, 41, 2333–2344. [Google Scholar] [CrossRef] [PubMed]
- Shkoporov, A.N.; Chaplin, A.V.; Shcherbakova, V.A.; Suzina, N.E.; Kafarskaia, L.I.; Bozhenko, V.K.; Efimov, B.A. Ruthenibacterium lactatiformans gen. nov., sp. nov., an anaerobic, lactate-producing member of the family Ruminococcaceae isolated from human faeces. Int. J. Syst. Evol. Microbiol. 2016, 66, 3041–3049. [Google Scholar] [CrossRef] [PubMed]
- Metcalfe-Roach, A.; Finlay, B.B. The role of the gut microbiome in Parkinson’s disease. Future Neurol. 2025, 20, 2494981. [Google Scholar] [CrossRef]
- VanOtterloo, L.M.; Trent, M.S. Microbial Primer: Lipopolysaccharide—A remarkable component of the Gram-negative bacterial surface. Microbiology 2024, 170, 1439. [Google Scholar] [CrossRef]
- Brown, G.C.; Camacho, M.; Williams-Gray, C.H. The Endotoxin Hypothesis of Parkinson’s Disease. Mov. Disord. 2023, 38, 1143–1155. [Google Scholar] [CrossRef]
- Chen, S.J.; Chen, C.C.; Liao, H.Y.; Lin, Y.T.; Wu, Y.W.; Liou, J.M.; Wu, M.S.; Kuo, C.H.; Lin, C.H. Association of Fecal and Plasma Levels of Short-Chain Fatty Acids with Gut Microbiota and Clinical Severity in Patients with Parkinson Disease. Neurology 2022, 98, e848–e858. [Google Scholar] [CrossRef]
- Liu, J.; Chen, Q.; Su, R. Interplay of human gastrointestinal microbiota metabolites: Short-chain fatty acids and their correlation with Parkinson’s disease. Medicine 2024, 103, e37960. [Google Scholar] [CrossRef]
- Dalile, B.; Van Oudenhove, L.; Vervliet, B.; Verbeke, K. The role of short-chain fatty acids in microbiota-gut-brain communication. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 461–478. [Google Scholar] [CrossRef]
- Caetano-Silva, M.E.; Rund, L.; Hutchinson, N.T.; Woods, J.A.; Steelman, A.J.; Johnson, R.W. Inhibition of inflammatory microglia by dietary fiber and short-chain fatty acids. Sci. Rep. 2023, 13, 2819. [Google Scholar] [CrossRef] [PubMed]
- Krueger, M.E.; Boles, J.S.; Simon, Z.D.; Alvarez, S.D.; McFarland, N.R.; Okun, M.S.; Zimmermann, E.M.; Forsmark, C.E.; Tansey, M.G. Comparative analysis of Parkinson’s and inflammatory bowel disease gut microbiomes reveals shared butyrate-producing bacteria depletion. npj Park. Dis. 2025, 11, 50. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; An, K.; Wang, D.; Yu, H.; Li, J.; Min, Z.; Xiong, Y.; Xue, Z.; Mao, Z. Short-Chain Fatty Acid Aggregates Alpha-Synuclein Accumulation and Neuroinflammation via GPR43-NLRP3 Signaling Pathway in a Model Parkinson’s Disease. Mol. Neurobiol. 2025, 62, 6612–6625. [Google Scholar] [CrossRef] [PubMed]
- Renaldi, R.; Wiguna, T.; Persico, A.M.; Tanra, A.J. p-Cresol and p-Cresyl Sulphate Boost Oxidative Stress: A Systematic Review of Recent Evidence. Basic Clin. Pharmacol. Toxicol. 2025, 137, e70065. [Google Scholar] [CrossRef]
- Paul, K.C.; Zhang, K.; Walker, D.I.; Sinsheimer, J.; Yu, Y.; Kusters, C.; Del Rosario, I.; Folle, A.D.; Keener, A.M.; Bronstein, J.; et al. Untargeted serum metabolomics reveals novel metabolite associations and disruptions in amino acid and lipid metabolism in Parkinson’s disease. Mol. Neurodegener. 2023, 18, 100. [Google Scholar] [CrossRef]
- Weersma, R.K.; Zhernakova, A.; Fu, J. Interaction between drugs and the gut microbiome. Gut 2020, 69, 1510–1519. [Google Scholar] [CrossRef]
- Boertien, J.M.; Murtomäki, K.; Pereira, P.A.B.; van der Zee, S.; Mertsalmi, T.H.; Levo, R.; Nojonen, T.; Mäkinen, E.; Jaakkola, E.; Laine, P.; et al. Fecal microbiome alterations in treatment-naive de novo Parkinson’s disease. npj Park. Dis. 2022, 8, 129. [Google Scholar] [CrossRef]
- Pereira, F.C.; Ge, X.; Kristensen, J.M.; Kirkegaard, R.H.; Maritsch, K.; Szamosvári, D.; Imminger, S.; Seki, D.; Shazzad, J.B.; Zhu, Y.; et al. The Parkinson’s disease drug entacapone disrupts gut microbiome homoeostasis via iron sequestration. Nat. Microbiol. 2024, 9, 3165–3183. [Google Scholar] [CrossRef]
- Cirstea, M.S.; Creus-Cuadros, A.; Lo, C.; Yu, A.C.; Serapio-Palacios, A.; Neilson, S.; Appel-Cresswell, S.; Finlay, B.B. A novel pathway of levodopa metabolism by commensal Bifidobacteria. Sci. Rep. 2023, 13, 19155. [Google Scholar] [CrossRef]
- Xu, K.; Sheng, S.; Zhang, F. Relationship Between Gut Bacteria and Levodopa Metabolism. Curr. Neuropharmacol. 2023, 21, 1536–1547. [Google Scholar] [CrossRef] [PubMed]
- Auffret, M.; Meuric, V.; Boyer, E.; Bonnaure-Mallet, M.; Vérin, M. Oral Health Disorders in Parkinson’s Disease: More than Meets the Eye. J. Park. Dis. 2021, 11, 1507–1535. [Google Scholar] [CrossRef] [PubMed]
- Murcia-Flores, L.; Sánchez-García, A.; Pecci-Lloret, M.P.; Rodríguez-Lozano, F.J. Association between oral dysbiosis and Parkinson’s disease: A systematic review. Front. Cell. Infect. Microbiol. 2025, 15, 1564362. [Google Scholar] [CrossRef] [PubMed]
- Cussotto, S.; Clarke, G.; Dinan, T.G.; Cryan, J.F. Psychotropics and the Microbiome: A Chamber of Secrets. Psychopharmacology 2019, 236, 1411–1432. [Google Scholar] [CrossRef]
- Alam, M.; Abbas, K.; Mustafa, M.; Usmani, N.; Habib, S. Microbiome-based therapies for Parkinson’s disease. Front Nutr. 2024, 11, 1496616. [Google Scholar] [CrossRef]
- Ghosh, D.; Ghosh, S.; Sharma, M.; Datta, S. A review of gut microbiota as a therapeutic approach for Parkinson’s disease. MGM J. Med. Sci. 2024, 11, 763–771. [Google Scholar] [CrossRef]
- da Silva, T.F.; Glória, R.A.; Americo, M.F.; Freitas, A.D.S.; de Jesus, L.C.L.; Barroso, F.A.L.; Laguna, J.G.; Coelho-Rocha, N.D.; Tavares, L.M.; le Loir, Y.; et al. Unlocking the Potential of Probiotics: A Comprehensive Review on Research, Production, and Regulation of Probiotics. Probiotics Antimicrob. Proteins 2024, 16, 1687–1723. [Google Scholar] [CrossRef]
- Tan, A.H.; Hor, J.W.; Chong, C.W.; Lim, S.Y. Probiotics for Parkinson’s disease: Current evidence and future directions. JGH Open 2020, 5, 414–419. [Google Scholar] [CrossRef]
- Mirzaei, H.; Sedighi, S.; Kouchaki, E.; Barati, E.; Dadgostar, E.; Aschner, M.; Tamtaji, O.R. Probiotics and the Treatment of Parkinson’s Disease: An Update. Cell. Mol. Neurobiol. 2022, 42, 2449–2457. [Google Scholar] [CrossRef]
- Zhu, F.; Yin, S.; Wang, Y.; Zhong, Y.; Ji, Q.; Wu, J. Effects of Probiotics on Neurodegenerative Disease-Related Symptoms and Systemic Inflammation: A Systematic Review. Int. J. Gen. Med. 2024, 17, 5941–5958. [Google Scholar] [CrossRef]
- Rout, M.; Prusty, S.K.; Kar, D.M. Probiotic: A Gut Microbiota-Based Therapeutic Approaches for the Treatment of Parkinson’s Disease. Curr. Rev. Clin. Exp. Pharmacol. 2024, 20, 322–333. [Google Scholar] [CrossRef]
- Kim, J.H.; Choi, Y.; Lee, S.; Oh, M.S. Probiotics as Potential Treatments for Neurodegenerative Diseases: A Review of the Evidence from in vivo to Clinical Trial. Biomol. Ther. 2025, 33, 54–74. [Google Scholar] [CrossRef]
- Kumar, D.; Bishnoi, M.; Kondepudi, K.K.; Sharma, S.S. Gut Microbiota-Based Interventions for Parkinson’s Disease: Neuroprotective Mechanisms and Current Perspective. Probiotics Antimicrob. Proteins 2025, 17, 2438–2460. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Wong, K.Y.; Xie, H. Modulation of Intestinal Inflammation and Protection of Dopaminergic Neurons in Parkinson’s Disease Mice through a Probiotic Formulation Targeting NLRP3 Inflammasome. J. Neuroimmune Pharmacol. 2025, 20, 9. [Google Scholar] [CrossRef] [PubMed]
- Soni, A.; Patel, A.; Acharya, S.; Patel, A.; Shah, U.; Patel, S.; Solanki, N.; Patel, M. Novel approaches to improve systemic bioavailability of curcumin using probiotics for rotenone-induced Parkinson’s disease in rodents. Am. J. Transl. Res. 2025, 17, 868–877. [Google Scholar] [CrossRef] [PubMed]
- Hassan, H.M.; Abou-Hany, H.O.; Shata, A.; Hellal, D.; El-Baz, A.M.; ElSaid, Z.H.; Haleem, A.A.; Morsy, N.E.; Abozied, R.M.; Elbrolosy, B.M.; et al. Vinpocetine and Lactobacillus Attenuated Rotenone-Induced Parkinson’s Disease and Restored Dopamine Synthesis in Rats through Modulation of Oxidative Stress, Neuroinflammation, and Lewy Bodies Inclusion. J. Neuroimmune Pharmacol. 2025, 20, 22. [Google Scholar] [CrossRef]
- Wang, W.; Shi, S.; Su, M.; Li, Y.; Yao, X.; Jiang, J.; Yao, W.; Qin, X.; Wang, Z.; Tang, C. Akkermansia muciniphila Akk11 Supplementation Attenuates MPTP-Induced Neurodegeneration by Inhibiting Microglial NLRP3 Inflammasome. Probiotics Antimicrob. Proteins 2025, 17, 2348–2361. [Google Scholar] [CrossRef]
- Francisco, M.; Grau, R. Biofilm proficient Bacillus subtilis prevents neurodegeneration in Caenorhabditis elegans Parkinson’s disease models via PMK-1/p38 MAPK and SKN-1/Nrf2 signaling. Sci. Rep. 2025, 15, 9864. [Google Scholar] [CrossRef]
- Sheikh, K.; Arasteh, J.; Tajabadi Ebrahimi, M.; Hesampour, A. Membrane vesicles from Lactobacillus acidophilus reduce intestinal inflammation and increase 5-HT in the substantia nigra of rats with Parkinson’s disease. Arch. Med. Res. 2025, 56, 103143. [Google Scholar] [CrossRef]
- Qi, Y.; Dong, Y.; Chen, J.; Xie, S.; Ma, X.; Yu, X.; Yu, Y.; Wang, Y. Lactiplantibacillus plantarum SG5 inhibits neuroinflammation in MPTP-induced PD mice through GLP-1/PGC-1α pathway. Exp. Neurol. 2025, 383, 115001. [Google Scholar] [CrossRef]
- Aktas, B.; Aslim, B.; Ozdemir, D.A. A Neurotherapeutic Approach with Lacticaseibacillus rhamnosus E9 on Gut Microbiota and Intestinal Barrier in MPTP-Induced Mouse Model of Parkinson’s Disease. Sci. Rep. 2024, 14, 15460. [Google Scholar] [CrossRef]
- Pérez Visñuk, D.; LeBlanc, J.G.; de Moreno de LeBlanc, A. Neuroprotective Effects Exerted by a Combination of Selected Lactic Acid Bacteria in a Mouse Parkinsonism Model under Levodopa-Benserazide Treatment. Neurochem. Res. 2024, 49, 2940–2956. [Google Scholar] [CrossRef]
- Valvaikar, S.; Vaidya, B.; Sharma, S.; Bishnoi, M.; Kondepudi, K.K.; Sharma, S.S. Supplementation of probiotic Bifidobacterium breve Bif11 reverses neurobehavioural deficits, inflammatory changes and oxidative stress in Parkinson’s disease model. Neurochem. Int. 2024, 174, 05691. [Google Scholar] [CrossRef]
- Dong, Y.; Qi, Y.; Chen, J.; Han, S.; Su, W.; Ma, X.; Yu, Y.; Wang, Y. Neuroprotective Effects of Bifidobacterium animalis subsp. lactis NJ241 in a Mouse Model of Parkinson’s Disease: Implications for Gut Microbiota and PGC-1α. Mol. Neurobiol. 2024, 61, 7534–7548. [Google Scholar] [CrossRef] [PubMed]
- Tsao, S.P.; Yeh, T.H.; Lin, Y.T.; Pan, C.H.; Lee, Y.K.; Wu, C.H.; Huang, H.Y. Supplementation with Bifidobacterium animalis subsp. lactis MH-022 for remission of motor impairments in a 6-OHDA-induced Parkinson’s disease rat model by reducing inflammation, reshaping the gut microbiome, and fostering specific microbial taxa. Food Funct. 2024, 15, 9368–9389. [Google Scholar] [CrossRef]
- Li, T.; Zhang, C.; Qiannan Wang, Q.; Wang, S.; Yu, L.; Qixiao Zhai, Q.; Tian, F. Exploring the therapeutic potential of Bifidobacterium longum subsp. longum CCFM1029 in Parkinson’s disease: Insights from behavioral, neurophysiological, gut microbiota, and microbial metabolites analysis. Foods 2024, 5, 156. [Google Scholar] [CrossRef]
- Stavrovskaya, A.V.; Voronkov, D.N.; Marsova, M.V.; Olshansky, A.S.; Gushchina, A.S.; Danilenko, V.N.; Illarioshkin, S.N. Effects of the pharmabiotic U-21 in a combined neuroinflammatory model of Parkinson’s disease in rats. Bull. Exp. Biol. Med. 2024, 177, 193–199. (In Russian) [Google Scholar] [CrossRef]
- Li, T.; Chu, C.; Yu, L.; Zhai, Q.; Wang, S.; Zhao, J.; Zhang, H.; Chen, W.; Tian, F. Neuroprotective effects of Bifidobacterium breve CCFM1067 in MPTP-induced mouse models of Parkinson’s disease. Nutrients 2022, 14, 4678. [Google Scholar] [CrossRef]
- Wu, H.; Wei, J.; Zhao, X.; Liu, Y.; Chen, Z.; Wei, K.; Lu, J.; Chen, W.; Jiang, M.; Li, S.; et al. Neuroprotective effects of an engineered Escherichia coli Nissle 1917 on Parkinson’s disease in mice by delivering GLP-1 and modulating gut microbiota. Bioeng. Transl. Med. 2022, 8, e10351. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, W.J.; Han, Y.Y.; Xu, X.; Yang, A.X.; Wei, J.; Hong, D.J.; Fang, X.; Chen, T.T. Neuroprotective effect of engineered Clostridium butyricum-pMTL007-GLP-1 on Parkinson’s disease mice models via promoting mitophagy. Bioeng. Transl. Med. 2024, 8, e10505. [Google Scholar] [CrossRef]
- Poluektova, E.; Yunes, R.; Danilenko, V. The Putative Antidepressant Mechanisms of Probiotic Bacteria: Relevant Genes and Proteins. Nutrients 2021, 13, 1591. [Google Scholar] [CrossRef]
- Kaur, H.; Nookala, S.; Singh, S.; Mukundan, S.; Nagamoto-Combs, K.; Combs, C.K. Sex-Dependent Effects of Intestinal Microbiome Manipulation in a Mouse Model of Alzheimer’s Disease. Cells 2021, 10, 2370. [Google Scholar] [CrossRef]
- Al Ebrahim, R.N.; Alekseeva, M.G.; Bazhenov, S.V.; Fomin, V.V.; Mavletova, D.A.; Nesterov, A.A.; Poluektova, E.U.; Danilenko, V.N.; Manukhov, I.V. ClpL Chaperone as a Possible Component of the Disaggregase Activity of Limosilactobacillus fermentum U-21. Biology 2024, 3, 592. [Google Scholar] [CrossRef]
- Khan, A.N.; Khan, R.H. Protein Misfolding and Related Human Diseases: A Comprehensive Review of Toxicity, Proteins Involved, and Current Therapeutic Strategies. Int. J. Biol. Macromol. 2022, 223, 143–160. [Google Scholar] [CrossRef]
- Meredith, M.E.; Salameh, T.S.; Banks, W.A. Intranasal Delivery of Proteins and Peptides in the Treatment of Neurodegenerative Diseases. AAPS J. 2015, 17, 780–787. [Google Scholar] [CrossRef] [PubMed]
- Braschi, C.; Capsoni, S.; Narducci, R.; Poli, A.; Sansevero, G.; Brandi, R.; Maffei, L.; Cattaneo, A.; Berardi, N. Intranasal delivery of BDNF rescues memory deficits in AD11 mice and reduces brain microgliosis. Aging Clin. Exp. Res. 2021, 33, 1223–1238. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Aggarwal, N.; Cui, B.; Foo, G.W.; He, Y.; Srivastava, S.K.; Li, S.; Seah, M.Z.X.; Wun, K.S.; Ling, H.; et al. Engineered commensals for targeted nose-to-brain drug delivery. Cell 2025, 188, 1545–1562.e16. [Google Scholar] [CrossRef]
- Poupet, C.; Chassard, C.; Nivoliez, A.; Bornes, S. Caenorhabditis elegans, a Host to Investigate the Probiotic Properties of Beneficial Microorganisms. Front. Nutr. 2020, 7, 135. [Google Scholar] [CrossRef]
- Goya, M.E.; Xue, F.; Sampedro-Torres-Quevedo, C.; Arnaouteli, S.; Riquelme-Dominguez, L.; Romanowski, A.; Brydon, J.; Ball, K.L.; Stanley-Wall, N.R.; Doitsidou, M. Probiotic Bacillus subtilis Protects against α-Synuclein Aggregation in C. elegans. Cell Rep. 2020, 30, 367–380.e7. [Google Scholar] [CrossRef]
- Gusarov, I.; Gautier, L.; Smolentseva, O.; Shamovsky, I.; Eremina, S.; Mironov, A.; Nudler, E. Bacterial nitric oxide extends the lifespan of C. elegans. Cell 2013, 152, 818–830. [Google Scholar] [CrossRef]
- Marsova, M.; Poluektova, E.; Odorskaya, M.; Ambaryan, A.; Revishchin, A.; Pavlova, G.; Danilenko, V. Protective effects of Lactobacillus fermentum U-21 against paraquat-induced oxidative stress in Caenorhabditis elegans and mouse models. World J. Microbiol. Biotechnol. 2020, 36, 104. [Google Scholar] [CrossRef] [PubMed]
- Rao, S.S.C.; Rehman, A.; Yu, S.; Andino, N.M. Brain fogginess, gas and bloating: A link between SIBO, probiotics and metabolic acidosis. Clin. Transl. Gastroenterol. 2018, 9, 162. [Google Scholar] [CrossRef] [PubMed]
- Leung, B.P.Y.; Tsui, B.S.Y.; Kan, I. D-lactic acidosis in short bowel syndrome: Are probiotics friend or foe? A case report. Hong Kong Med. J. 2025, 31, 319–321. [Google Scholar] [CrossRef] [PubMed]
- Nataraj, B.H.; Ali, S.A.; Behare, P.V.; Yadav, H. Postbiotics-parabiotics: The new horizons in microbial biotherapy and functional foods. Microb. Cell. Fact. 2020, 19, 168. [Google Scholar] [CrossRef]
- Yunes, R.A.; Poluektova, E.U.; Belkina, T.V.; Danilenko, V.N. Lactobacilli: Legal regulation and prospects for new generation drugs. Appl. Biochem. Microbiol. 2022, 58, 652–664. [Google Scholar] [CrossRef]
- Fan, H.X.; Sheng, S.; Li, D.D.; Li, J.J.; Wang, G.Q.; Zhang, F. Heat-killed Lactobacillus murinus confers neuroprotection against dopamine neuronal loss by targeting NLRP3 inflammasome. Bioeng. Transl. Med. 2022, 8, e10455. [Google Scholar] [CrossRef]
- Głowacka, P.; Oszajca, K.; Pudlarz, A.; Szemraj, J.; Witusik-Perkowska, M. Postbiotics as Molecules Targeting Cellular Events of Aging Brain-The Role in Pathogenesis, Prophylaxis and Treatment of Neurodegenerative Diseases. Nutrients 2024, 16, 2244. [Google Scholar] [CrossRef]
- Toyofuku, M.; Schild, S.; Kaparakis-Liaskos, M.; Eberl, L. Composition and functions of bacterial membrane vesicles. Nat. Rev. Microbiol. 2023, 21, 415–430. [Google Scholar] [CrossRef]
- Koukoulis, T.F.; Beauchamp, L.C.; Kaparakis-Liaskos, M.; McQuade, R.M.; Purnianto, A.; Finkelstein, D.I.; Barnham, K.J.; Vella, L.J. Do Bacterial Outer Membrane Vesicles Contribute to Chronic Inflammation in Parkinson’s Disease? J. Park. Dis. 2024, 14, 227–244. [Google Scholar] [CrossRef]
- Krzyżek, P.; Marinacci, B.; Vitale, I.; Grande, R. Extracellular Vesicles of Probiotics: Shedding Light on the Biological Activity and Future Applications. Pharmaceutics 2023, 15, 522. [Google Scholar] [CrossRef]
- Xie, J.; Haesebrouck, F.; Van Hoecke, L.; Vandenbroucke, R.E. Bacterial extracellular vesicles: An emerging avenue to tackle diseases. Trends Microbiol. 2023, 31, 1206–1224. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, Y.E.Q.; Naveed, M.; Wang, X.; Liu, Y.; Li, M. The biological activity and potential of probiotics-derived extracellular vesicles as postbiotics in modulating microbiota-host communication. J. Nanobiotechnol. 2025, 23, 349. [Google Scholar] [CrossRef] [PubMed]
- Haasbroek, K.; Otaka, S.; Sakiyama, C.; Iwase, S.; Yagi, M.; Yonei, Y. Extracellular vesicles from probiotic microorganisms enhance microglia amyloid β phagocytosis. Glycative Stress Res. 2025, 12, 25–38. [Google Scholar] [CrossRef]
- Yang, Y.; Li, N.; Gao, Y.; Xu, F.; Chen, H.; Zhang, C.; Ni, X. The activation impact of lactobacillus-derived extracellular vesicles on lipopolysaccharide-induced microglial cell. BMC Microbiol. 2024, 24, 70. [Google Scholar] [CrossRef] [PubMed]
- Yadav, P.; Debnath, N.; Pradhan, D.; Mehta, P.K.; Kumar, A.; Yadav, M.L.; Yadav, A.K. Probiotic Lactobacillus-Derived Extracellular Vesicles: Insights Into Disease Prevention and Management. Mol. Nutr. Food Res. 2025, 69, e70013. [Google Scholar] [CrossRef]
- Grishina, Y.V.; Vatlin, A.A.; Mavletova, D.A.; Odorskaya, M.V.; Senkovenko, A.M.; Ilyasov, R.A.; Danilenko, V.N. Metabolites Potentially Determine the High Antioxidant Properties of Limosilactobacillus fermentum U-21. BioTech 2023, 12, 39. [Google Scholar] [CrossRef]
- Odorskaya, M.V.; Mavletova, D.A.; Nesterov, A.A.; Tikhonova, O.V.; Soloveva, N.A.; Reznikova, D.A.; Galanova, O.O.; Vatlin, A.A.; Slynko, N.M.; Vasilieva, A.R.; et al. The use of omics technologies in creating LBP and postbiotics based on the Limosilactobacillus fermentum U-21. Front. Microbiol. 2024, 15, 1416688. [Google Scholar] [CrossRef]
- Cassani, E.; Privitera, G.; Pezzoli, G.; Pusani, C.; Madio, C.; Iorio, L.; Barichella, M. Use of probiotics for the treatment of constipation in Parkinson’s disease patients. Minerva Gastroenterol. Dietol. 2011, 57, 117–121. [Google Scholar] [PubMed]
- Barichella, M.; Pacchetti, C.; Bolliri, C.; Cassani, E.; Iorio, L.; Pusani, C.; Pinelli, G.; Privitera, G.; Cesari, I.; Faierman, S.A.; et al. Probiotics and prebiotic fiber for constipation associated with Parkinson disease: An RCT. Neurology 2016, 87, 1274–1280. [Google Scholar] [CrossRef]
- Georgescu, D.; Ancusa, O.E.; Georgescu, L.A.; Ionita, I.; Reisz, D. Nonmotor gastrointestinal disorders in older patients with Parkinson’s disease: Is there hope? Clin. Interv. Aging 2016, 11, 1601–1608. [Google Scholar] [CrossRef] [PubMed]
- Borzabadi, S.; Oryan, S.; Eidi, A.; Aghadavod, E.; Daneshvar Kakhaki, R.; Tamtaji, O.R.; Taghizadeh, M.; Asemi, Z. The Effects of Probiotic Supplementation on Gene Expression Related to Inflammation, Insulin and Lipid in Patients with Parkinson’s Disease: A Randomized, Double-blind, PlaceboControlled Trial. Arch. Iran. Med. 2018, 21, 289–295. [Google Scholar] [PubMed]
- Ibrahim, A.; Ali, R.A.R.; Manaf, M.R.A.; Ahmad, N.; Tajurruddin, F.W.; Qin, W.Z.; Desa, S.H.M.; Ibrahim, N.M. Multistrain probiotics (Hexbio) containing MCP BCMC strains improved constipation and gut motility in Parkinson’s disease: A randomized controlled trial. PLoS ONE 2020, 15, e0244680. [Google Scholar] [CrossRef] [PubMed]
- Ghyselinck, J.; Verstrepen, L.; Moens, F.; Abbeele, P.V.D.; Bruggeman, A.; Said, J.; Smith, B.; Barker, L.A.; Jordan, C.; Leta, V.; et al. Influence of probiotic bacteria on gut -microbiota composition and gut wall function in an in-vitro model in patients with Parkinson’s disease. Int. J. Pharm. X 2021, 3, 100087. [Google Scholar] [CrossRef]
- Lu, C.-S.; Chang, H.-C.; Weng, Y.-H.; Chen, C.-C.; Kuo, Y.-S.; Tsai, Y.-C. The Add-On Effect of Lactobacillus plantarum PS128 in Patients With Parkinson’s Disease: A Pilot Study. Front. Nutr. 2021, 8, 650053. [Google Scholar] [CrossRef]
- Tan, A.H.; Lim, S.Y.; Chong, K.K.; Manap, M.A.A.A.; Hor, J.W.; Lim, J.L.; Low, S.C.; Chong, C.W.; Mahadeva, S.; Lang, A.E. Probiotics for Constipation in Parkinson Disease: A Randomized Placebo-Controlled Study. Neurology 2021, 96, e772–e782. [Google Scholar] [CrossRef]
- Sun, H.; Zhao, F.; Liu, Y.; Ma, T.; Jin, H.; Quan, K.; Leng, B.; Zhao, J.; Yuan, X.; Li, Z.; et al. Probiotics synergized with conventional regimen in managing Parkinson’s disease. npj Park. Dis. 2022, 8, 62. [Google Scholar] [CrossRef]
- Yang, X.; He, X.; Xu, S.; Zhang, Y.; Mo, C.; Lai, Y.; Song, Y.; Yan, Z.; Ai, P.; Qian, Y.; et al. Effect of Lacticaseibacillus paracasei strain Shirota supplementation on clinical responses and gut microbiome in Parkinson’s disease. Food Funct. 2023, 14, 6828–6839. [Google Scholar] [CrossRef]
- Mehrabani, S.; Khorvash, F.; Heidari, Z.; Tajabadi-Ebrahimi, M.; Amani, R. The effects of synbiotic supplementation on oxidative stress markers, mental status, and quality of life in patients with Parkinson’s disease: A double-blind, placebo-controlled, randomized controlled trial. J. Funct. Foods 2023, 100, 105397. [Google Scholar] [CrossRef]
- Ghalandari, N.; Assarzadegan, F.; Habibi, S.A.H.; Esmaily, H.; Malekpour, H. Efficacy of probiotics in improving motor functionand alleviating constipation in Parkinson’s disease: A randomized controlled trial. Iran. J. Pharm. Res. 2023, 22, e137840. [Google Scholar] [CrossRef]
- Andreozzi, V.; Cuoco, S.; Balestrieri, M.; Fierro, F.; Ferrara, N.; Erro, R.; Di Filippo, M.; Barbella, G.; Memoli, M.C.; Silvestri, A.; et al. Synbiotic supplementation may globally improve non-motor symptoms in patients with stable Parkinson’s disease: Results from an open label single-arm study. Sci. Rep. 2024, 14, 23095. [Google Scholar] [CrossRef]
- Magistrelli, L.; Contaldi, E.; Visciglia, A.; Deusebio, G.; Pane, M.; Amoruso, A. The Impact of Probiotics on Clinical Symptoms and Peripheral Cytokines Levels in Parkinson’s Disease: Preliminary In Vivo Data. Brain Sci. 2024, 14, 1147. [Google Scholar] [CrossRef]
- Zali, A.; Hajyani, S.; Salari, M.; Tajabadi-Ebrahimi, M.; Mortazavian, A.M.; Pakpour, B. Co-administration of probiotics and vitamin D reduced disease severity and complications in patients with Parkinson’s disease: A randomized controlled clinical trial. Psychopharmacology 2024, 241, 1905–1914. [Google Scholar] [CrossRef]
- Ramadan, M.E.; Mostafa, T.M.; Ghali, A.A.; El-Afify, D.R. Randomized controlled trial evaluating synbiotic supplementation as an adjuvant therapy in the treatment of Parkinson’s disease. Inflammopharmacology 2025, 33, 3897–3908. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.Z.; Cheng, S.H.; Chang, M.Y.; Lin, Y.F.; Wu, C.C.; Tsai, Y.C. Neuroprotective Effects of Lactobacillus plantarum PS128 in a Mouse Model of Parkinson’s Disease: The Role of Gut Microbiota and MicroRNAs. Int. J. Mol. Sci. 2023, 24, 6794. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Wu, M.; Zhao, W.; Kwok, L.-Y.; Zhang, W. Effects of probiotics and its fermented milk on constipation: A systematic review. Food Sci. Hum. Well. 2023, 12, 2124–2134. [Google Scholar] [CrossRef]
- Neiworth-Petshow, E.M.; Baldwin-Sayre, C. Naturopathic Treatment of Gastrointestinal Dysfunction in the Setting of Parkinson’s Disease. Integr. Med. 2018, 17, 44–50. [Google Scholar] [PubMed]
- Averina, O.V.; Poluektova, E.U.; Zorkina, Y.A.; Kovtun, A.S.; Danilenko, V.N. Human Gut Microbiota for Diagnosis and Treatment of Depression. Int. J. Mol. Sci. 2024, 25, 5782. [Google Scholar] [CrossRef]
- Ferrari, S.; Mulè, S.; Parini, F.; Galla, R.; Ruga, S.; Rosso, G.; Brovero, A.; Molinari, C.; Uberti, F. The influence of the gut-brain axis on anxiety and depression: A review of the literature on the use of probiotics. J. Tradit. Complement. Med. 2024, 14, 237–255. [Google Scholar] [CrossRef]
- Çınar, E.; Tel, B.C.; Şahin, G. Neuroinflammation in Parkinson’s Disease and its Treatment Opportunities. Balk. Med. J. 2022, 39, 318–333. [Google Scholar] [CrossRef]
- Chen, X.; Yan, L.; Yang, J.; Xu, C.; Yang, L. The impact of probiotics on oxidative stress and inflammatory markers in patients with diabetes: A meta-research of meta-analysis studies. Front. Nutr. 2025, 12, 1552358. [Google Scholar] [CrossRef] [PubMed]
- Salles, B.I.M.; Cioffi, D.; Ferreira, S.R.G. Probiotics supplementation and insulin resistance: A systematic review. Diabetol. Metab. Syndr. 2020, 12, 98. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, M.C.; Garbim Junior, E.E.; Jahnke, V.S.; Lisbôa Moura, J.G.; Brasil, C.S.; Schimitt da Cunha, P.H.; Lora, P.S.; Gemelli, T. Impact of probiotic supplementation in a patient with type 2 diabetes on glycemic and lipid profile. Clin. Nutr. ESPEN 2022, 49, 264–269. [Google Scholar] [CrossRef] [PubMed]
- Mal, S.; Dwivedi, A.R.; Kumar, V.; Kumar, N.; Kumar, B.; Kumar, V. Role of Peroxisome Proliferator-Activated Receptor Gamma (PPARγ) in Different Disease States: Recent Updates. Curr. Med. Chem. 2021, 28, 3193–3215. [Google Scholar] [CrossRef]
- Hong, C.T.; Chen, J.H.; Huang, T.W. Probiotics treatment for Parkinson disease: A systematic review and meta-analysis of clinical trials. Aging 2022, 14, 7014–7025. [Google Scholar] [CrossRef]
- Xie, L.; Chen, D.; Zhu, X.; Cheng, C. Efficacy and safety of probiotics in Parkinson’s constipation: A systematic review and meta-analysis. Front. Pharmacol. 2023, 13, 1007654. [Google Scholar] [CrossRef]
- Ghalandari, N.; Assarzadegan, F.; Mahdavi, H.; Jamshidi, E.; Esmaily, H. Evaluating the effectiveness of probiotics in relieving constipation in Parkinson’s disease: A systematic review and meta-analysis. Heliyon 2023, 9, e14312. [Google Scholar] [CrossRef]
- Jin, X.; Dong, W.; Chang, K. Efficacy of probiotic supplements on Parkinson’s disease and meta-analysis. Complement. Ther. Med. 2024, 82, 103045. [Google Scholar] [CrossRef]
- Park, J.M.; Lee, S.C.; Ham, C.; Kim, Y.W. Effect of probiotic supplementation on gastrointestinal motility, inflammation, motor, non-motor symptoms and mental health in Parkinson’s disease: A meta-analysis of randomized controlled trials. Gut Pathog. 2023, 15, 9. [Google Scholar] [CrossRef]
- Ugrumov, M. Development of early diagnosis of Parkinson’s disease: Illusion or reality? CNS Neurosci. Ther. 2020, 26, 997–1009. [Google Scholar] [CrossRef]
- Katunina, E.A.; Blokhin, V.; Nodel, M.R.; Pavlova, E.N.; Kalinkin, A.L.; Kucheryanu, V.G.; Alekperova, L.; Selikhova, M.V.; Martynov, M.Y.; Ugrumov, M.V. Searching for Biomarkers in the Blood of Patients at Risk of Developing Parkinson’s Disease at the Prodromal Stage. Int. J. Mol. Sci. 2023, 24, 1842. [Google Scholar] [CrossRef]
- Blokhin, V.; Pavlova, E.N.; Katunina, E.A.; Nodel, M.R.; Kataeva, G.V.; Moskalets, E.R.; Pronina, T.S.; Ugrumov, M.V. Dopamine Synthesis in the Nigrostriatal Dopaminergic System in Patients at Risk of Developing Parkinson’s Disease at the Prodromal Stage. J. Clin. Med. 2024, 13, 875. [Google Scholar] [CrossRef] [PubMed]
- Axial Therapeutics|United States, Biotechnology Company|HOME. Available online: https://www.axialbiotherapeutics.com (accessed on 1 August 2025).
- Research|Bened Biomedical Co., Ltd. Available online: https://www.benedbiomed.com/research (accessed on 1 August 2025).
- 4D Pharma Plc—Developing Science, Delivering Therapies. Available online: https://www.4dpharmaplc.com/ (accessed on 1 August 2025).
- ADS024. Available online: https://www.ads024.com/ads024 (accessed on 1 August 2025).
- Ianiro, G.; Bibbò, S.; Porcari, S.; Settanni, C.R.; Giambò, F.; Curta, A.R.; Quaranta, G.; Scaldaferri, F.; Masucci, L.; Sanguinetti, M.; et al. Fecal microbiota transplantation for recurrent infection in patients with inflammatory bowel disease: Experience of a large-volume European FMT center. Gut Microbes 2021, 13, 1994834. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Ning, J.; Bao, X.Q.; Shang, M.; Ma, J.; Li, G.; Zhang, D. Fecal microbiota transplantation protects rotenone-induced Parkinson’s disease mice via suppressing inflammation mediated by the lipopolysaccharide-TLR4 signaling pathway through the microbiota-gut-brain axis. Microbiome 2021, 9, 226. [Google Scholar] [CrossRef] [PubMed]
- Ni, Y.; Tong, Q.; Xu, M.; Gu, J.; Ye, H. Gut Microbiota-Induced Modulation of the Central Nervous System Function in Parkinson’s Disease Through the Gut-Brain Axis and Short-Chain Fatty Acids. Mol. Neurobiol. 2025, 62, 2480–2492. [Google Scholar] [CrossRef]
- Cheng, Y.; Tan, G.; Zhu, Q.; Wang, C.; Ruan, G.; Ying, S.; Qie, J.; Hu, X.; Xiao, Z.; Xu, F.; et al. Efficacy of fecal microbiota transplantation in patients with Parkinson’s disease: Clinical trial results from a randomized, placebo-controlled design. Gut Microbes 2023, 15, 2284247. [Google Scholar] [CrossRef]
- DuPont, H.L.; Suescun, J.; Jiang, Z.D.; Brown, E.L.; Essigmann, H.T.; Alexander, A.S.; DuPont, A.W.; Iqbal, T.; Utay, N.S.; Newmark, M.; et al. Fecal microbiota transplantation in Parkinson’s disease-A randomized repeat-dose, placebo-controlled clinical pilot study. Front. Neurol. 2023, 14, 1104759. [Google Scholar] [CrossRef]
- Bruggeman, A.; Vandendriessche, C.; Hamerlinck, H.; De Looze, D.; Tate, D.J.; Vuylsteke, M.; De Commer, L.; Devolder, L.; Raes, J.; Verhasselt, B.; et al. Safety and efficacy of faecal microbiota transplantation in patients with mild to moderate Parkinson’s disease (GUT-PARFECT): A double-blind, placebo-controlled, randomised, phase 2 trial. EClinicalMedicine 2024, 71, 102563. [Google Scholar] [CrossRef]
- Scheperjans, F.; Levo, R.; Bosch, B.; Lääperi, M.; Pereira, P.A.B.; Smolander, O.P.; Aho, V.T.E.; Vetkas, N.; Toivio, L.; Kainulainen, V.; et al. Fecal Microbiota Transplantation for Treatment of Parkinson Disease: A Randomized Clinical Trial. JAMA Neurol. 2024, 81, 925–938. [Google Scholar] [CrossRef]
- Sampson, T.R. Fecal Microbiome Transplants For Parkinson Disease. JAMA Neurol. 2024, 81, 911–913. [Google Scholar] [CrossRef]
- Rojas-Velazquez, D.; Kidwai, S.; Liu, T.C.; El-Yacoubi, M.A.; Garssen, J.; Tonda, A.; Lopez-Rincon, A. Understanding Parkinson’s: The microbiome and machine learning approach. Maturitas 2025, 193, 108185. [Google Scholar] [CrossRef] [PubMed]
- Rabie, H.; Akhloufi, M.A. A review of machine learning and deep learning for Parkinson’s disease detection. Discov. Artif. Intell. 2025, 5, 24. [Google Scholar] [CrossRef] [PubMed]
- Pietrucci, D.; Teofani, A.; Unida, V.; Cerroni, R.; Biocca, S.; Stefani, A.; Desideri, A. Can Gut Microbiota Be a Good Predictor for Parkinson’s Disease? A Machine Learning Approach. Brain Sci. 2020, 10, 242. [Google Scholar] [CrossRef] [PubMed]
- Romano, S.; Wirbel, J.; Ansorge, R.; Schudoma, C.; Ducarmon, Q.R.; Narbad, A.; Zeller, G. Machine learning-based meta-analysis reveals gut microbiome alterations associated with Parkinson’s disease. Nat. Commun. 2025, 16, 4227. [Google Scholar] [CrossRef]
- Yu, B.; Zhang, H.; Zhang, M. Deep learning-based differential gut flora for prediction of Parkinson’s. PLoS ONE 2025, 20, e0310005. [Google Scholar] [CrossRef]
- Nie, S.; Wang, J.; Deng, Y.; Ye, Z.; Ge, Y. Inflammatory microbes and genes as potential biomarkers of Parkinson’s disease. Npj Biofilms Microbiomes 2022, 8, 101. [Google Scholar] [CrossRef]
- Morton, J.T.; Jin, D.M.; Mills, R.H.; Shao, Y.; Rahman, G.; McDonald, D.; Zhu, Q.; Balaban, M.; Jiang, Y.; Cantrell, K.; et al. Multi-level analysis of the gut-brain axis shows autism spectrum disorder-associated molecular and microbial profiles. Nat. Neurosci. 2023, 26, 1208–1217. [Google Scholar] [CrossRef]
- Kovtun, A.S.; Averina, O.V.; Angelova, I.Y.; Yunes, R.A.; Zorkina, Y.A.; Morozova, A.Y.; Pavlichenko, A.V.; Syunyakov, T.S.; Karpenko, O.A.; Kostyuk, G.P.; et al. Alterations of the Composition and Neurometabolic Profile of Human Gut Microbiota in Major Depressive Disorder. Biomedicines 2022, 10, 2162. [Google Scholar] [CrossRef]
- Angelova, I.Y.; Kovtun, A.S.; Averina, O.V.; Koshenko, T.A.; Danilenko, V.N. Unveiling the Connection between Microbiota and Depressive Disorder through Machine Learning. Int. J. Mol. Sci. 2023, 24, 16459. [Google Scholar] [CrossRef]
- Danilenko, V.; Devyatkin, A.; Marsova, M.; Shibilova, M.; Ilyasov, R.; Shmyrev, V. Common Inflammatory Mechanisms in COVID-19 and Parkinson’s Diseases: The Role of Microbiome, Pharmabiotics and Postbiotics in Their Prevention. J. Inflamm. Res. 2021, 14, 6349–6381. [Google Scholar] [CrossRef]
- Zhang, J.; Bishir, M.; Barbhuiya, S.; Chang, S.L. Meta-Analysis of the Mechanisms Underlying COVID-19 Modulation of Parkinson’s Disease. Int. J. Mol. Sci. 2023, 24, 13554. [Google Scholar] [CrossRef]
- Cordaillat-Simmons, M.; Rouanet, A.; Pot, B. Live biotherapeutic products: The importance of a defined regulatory framework. Exp. Mol. Med. 2020, 52, 1397–1406. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).