
Genomic and Functional Characterization of Novel Phages Targeting Multidrug-Resistant Acinetobacter baumannii

 ,
,  ,
,  , , and
, , and
Abstract
1. Introduction
2. Results
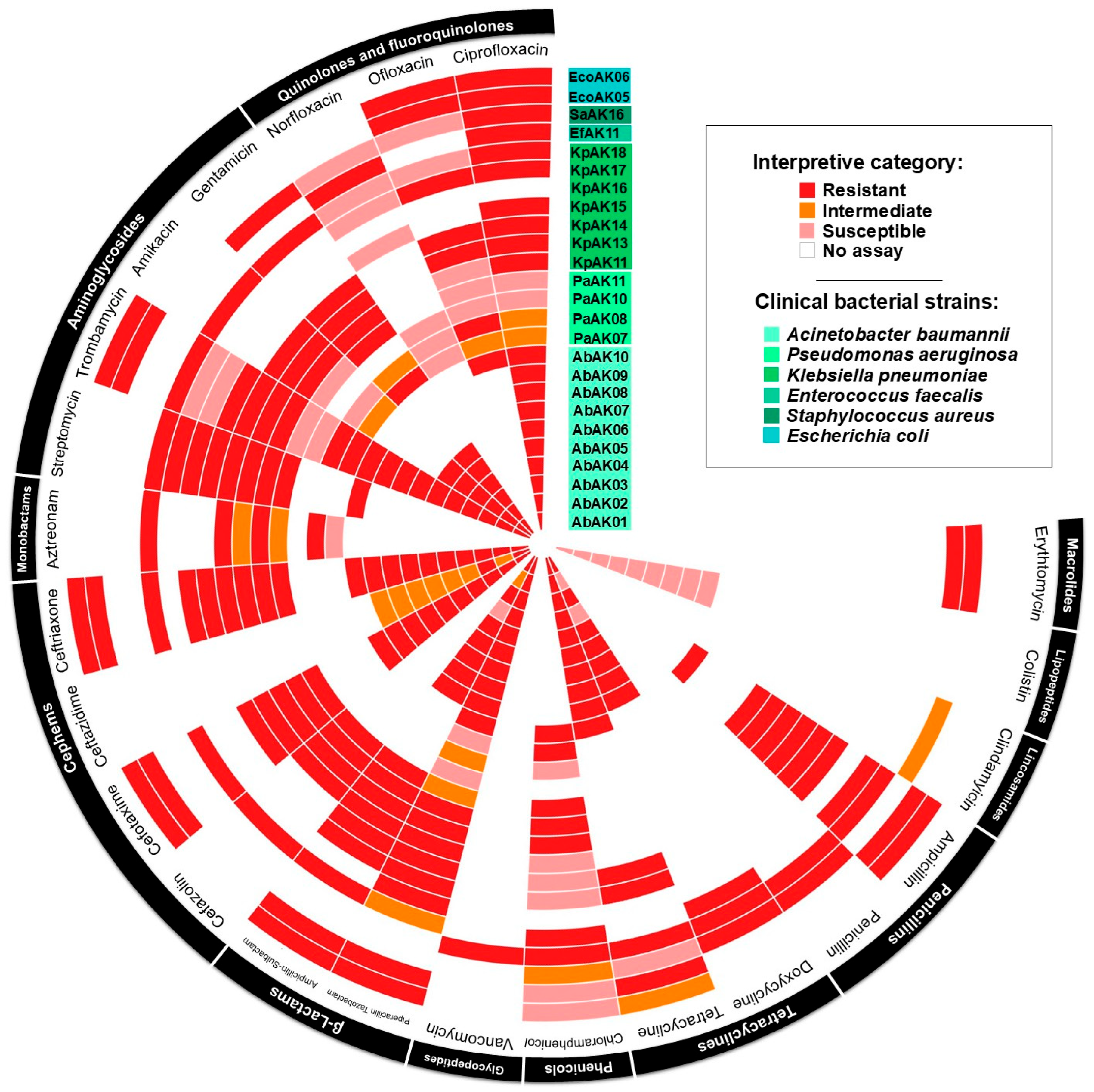
2.1. Antimicrobial Susceptibility Testing
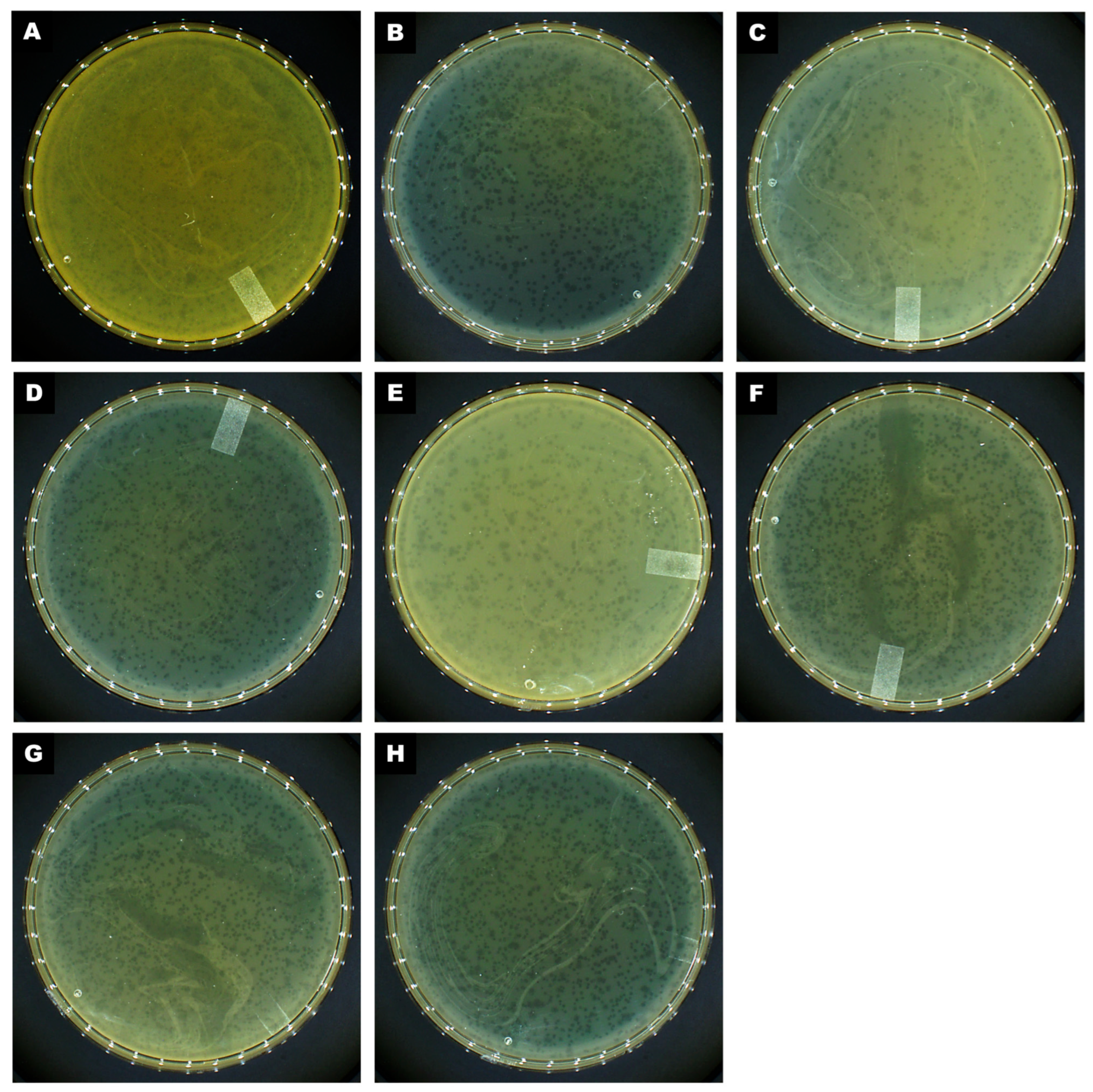
2.2. Phage Isolation, Plaque Characterization, and Host Range Analysis
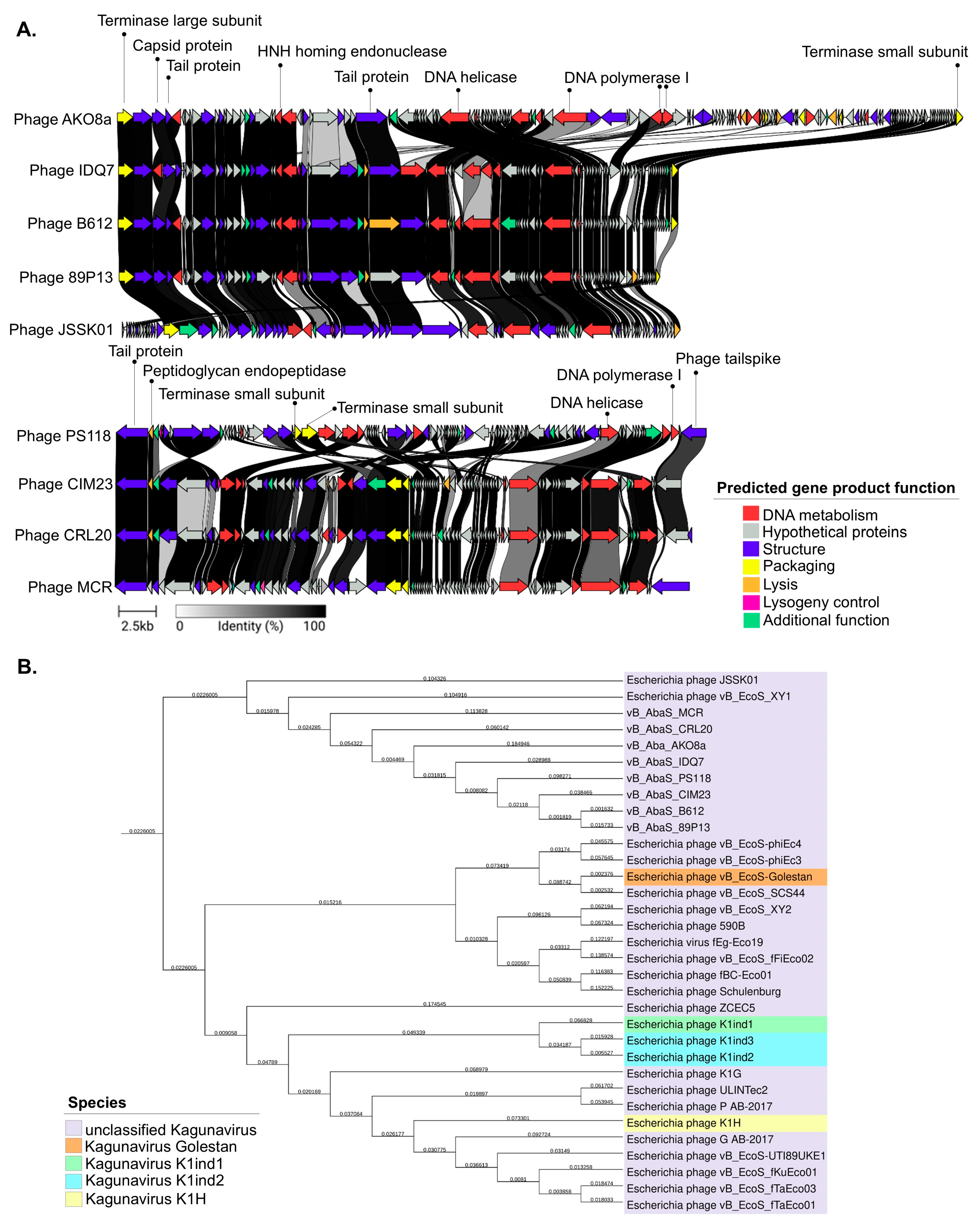
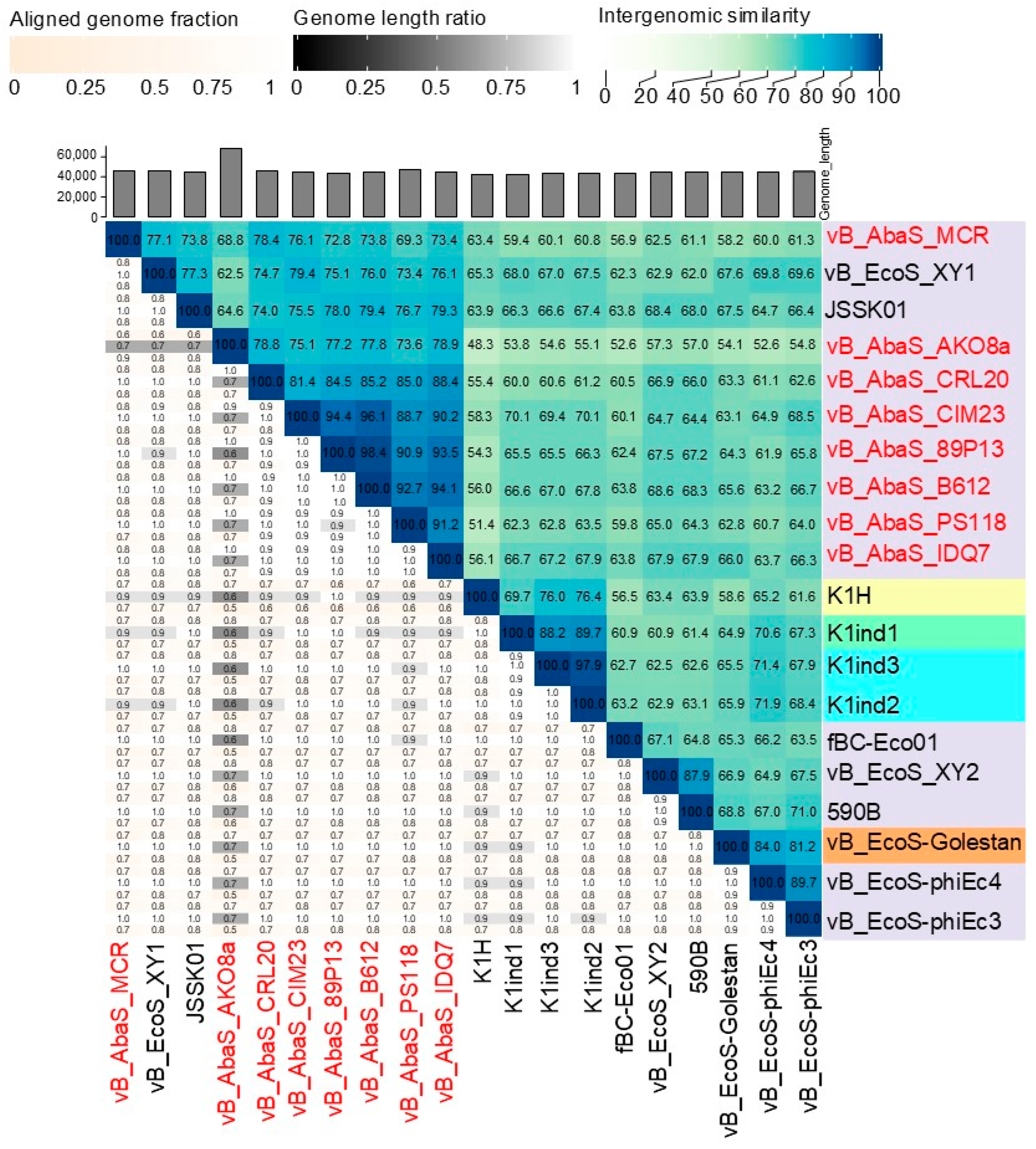
2.3. Comparative Genomics and Phylogenetic Analysis of Acinetobacter baumannii-Targeting Phages as a Promising Alternative Intervention
2.4. Evaluating Lysogenic Activity in Phages
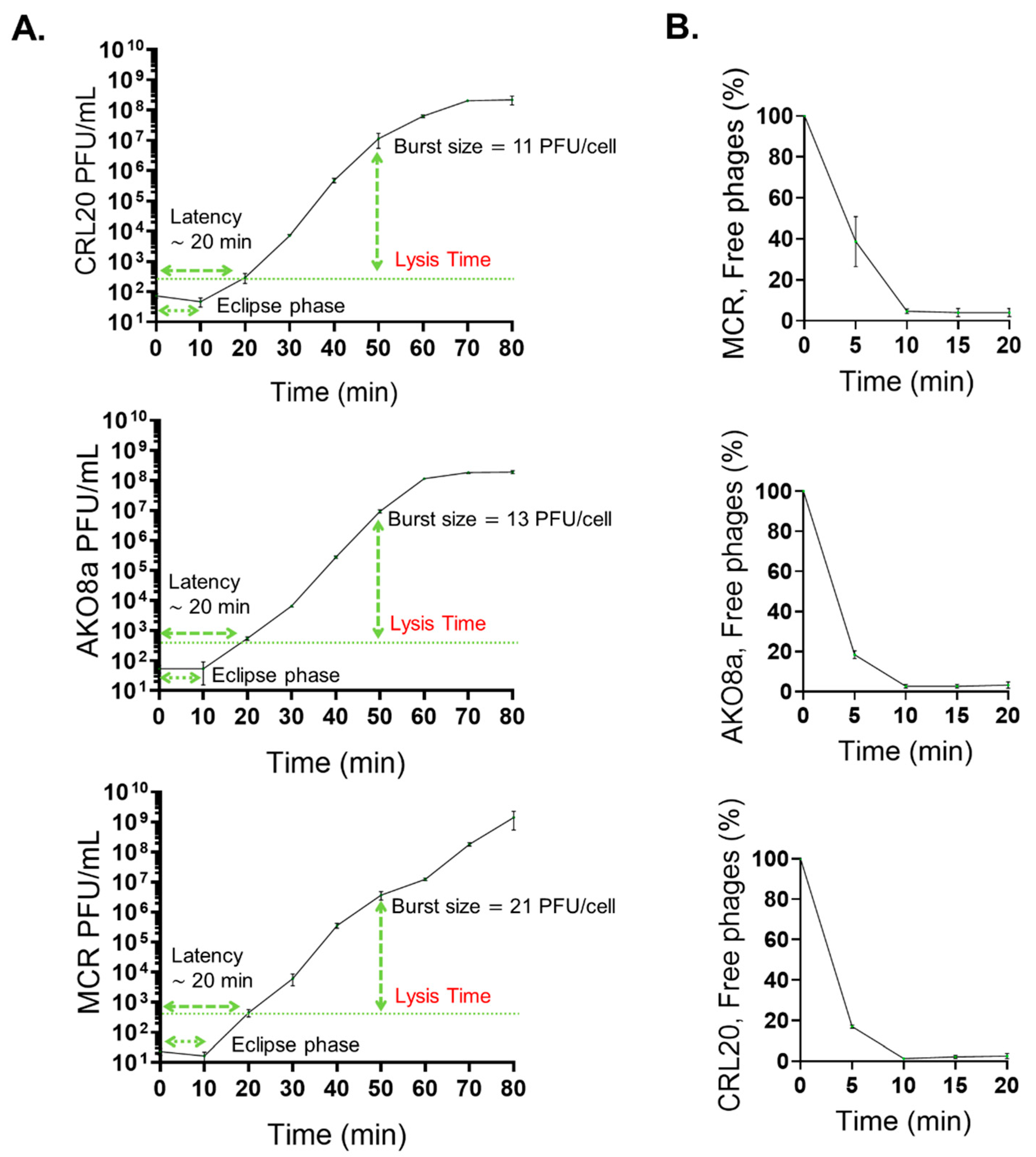
2.5. Antibacterial Activity, One-Step Growth Curve, and Phage Adsorption Rate
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains, Culture Conditions, and Storage
4.2. Antimicrobial Susceptibility Profile
4.3. Phage Isolation, Propagation, and Titration
4.4. Host Range of Phage and Efficiency of Plating
4.5. Phage DNA Extraction, Sequencing and Genome Assembly, Detection of Lysogeny
4.6. Bioinformatics Analysis
4.7. Phage-Mediated Antibacterial Activity Assays
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lobanovska, M.P.; Pilla, G. Penicillin’s Discovery and Antibiotic Resistance: Lessons for the Future? Yale J. Biol. Med. 2017, 90, 135–145. [Google Scholar] [PubMed]
- Salam, M.A.; Al-Amin, M.Y.; Salam, M.T.; Pawar, J.S.; Akhter, N.; Rabaan, A.A.; Alqumber, M.A.A. Antimicrobial Resistance: A Growing Serious Threat for Global Public Health. Healthcare 2023, 11, 1946. [Google Scholar] [CrossRef] [PubMed]
- Coque, T.M.; Canton, R.; Perez-Cobas, A.E.; Fernandez-de-Bobadilla, M.D.; Baquero, F. Antimicrobial Resistance in the Global Health Network: Known Unknowns and Challenges for Efficient Responses in the 21st Century. Microorganisms 2023, 11, 1050. [Google Scholar] [CrossRef]
- Ahmed, S.K.; Hussein, S.; Qurbani, K.; Ibrahim, R.H.; Fareeq, A.; Mahmood, K.A.; Mohamed, M.G. Antimicrobial resistance: Impacts, challenges, and future prospects. J. Med. Surg. Public Health 2024, 2, 100081. [Google Scholar] [CrossRef]
- Naghavi, M.; Vollset, S.E.; Ikuta, K.S.; Swetschinski, L.R.; Gray, A.P.; E Wool, E.; Aguilar, G.R.; Mestrovic, T.; Smith, G.; Han, C.; et al. Global burden of bacterial antimicrobial resistance 1990–2021: A systematic analysis with forecasts to 2050. Lancet 2024, 404, 1199–1226. [Google Scholar] [CrossRef]
- Attili, A.R.; Nocera, F.P.; Sisto, M.; Linardi, M.; Gigli, F.; Ngwa, V.N.; Fiorito, F.; Cerracchio, C.; Meligrana, M.C.T.; Bonacucina, E.; et al. Evidence and antibiotic resistance profiles of clinical Acinetobacter calcoaceticus-Acinetobacter baumannii (ACB) and non-ACB complex members in companion animals: A 2020–2022 retrospective study. Comp. Immunol. Microbiol. Infect. Dis. 2024, 109, 102185. [Google Scholar] [CrossRef]
- Thacharodi, A.; Vithlani, A.; Hassan, S.; Alqahtani, A.; Pugazhendhi, A. Carbapenem-resistant Acinetobacter baumannii raises global alarm for new antibiotic regimens. iScience 2024, 27, 111367. [Google Scholar] [CrossRef]
- Sati, H.; Carrara, E.; Savoldi, A.; Hansen, P.; Garlasco, J.; Campagnaro, E.; Boccia, S.; Castillo-Polo, J.A.; Magrini, E.; Garcia-Vello, P.; et al. The WHO Bacterial Priority Pathogens List 2024: A prioritisation study to guide research, development, and public health strategies against antimicrobial resistance. Lancet Infect. Dis. 2025. [Google Scholar] [CrossRef]
- Luo, J.; Xie, L.; Yang, M.; Liu, M.; Li, Q.; Wang, P.; Fan, J.; Jin, J.; Luo, C. Synergistic Antibacterial Effect of Phage pB3074 in Combination with Antibiotics Targeting Cell Wall against Multidrug-Resistant Acinetobacter baumannii. In Vitro and Ex Vivo. Microbiol. Spectr. 2023, 11, e0034123. [Google Scholar] [CrossRef]
- RHOVE. Boletin Estatal Panorama Epidemiológico de las Infecciones Asociadas a la Atención de la Salud; Secretaría de Salud México. Red Hospitalaria de Vigilancia Epidemiológica (RHOVE): Mexico City, Mexico, 2023. [Google Scholar]
- RHOVE-Sinaloa. Boletin Estatal Panorama Epidemiológico de las Infecciones Asociadas a la Atención de la Salud; Secretaría de Salud de Sinaloa: Culiacán, Mexico, 2025. [Google Scholar]
- Gordillo Altamirano, F.; Forsyth, J.H.; Patwa, R.; Kostoulias, X.; Trim, M.; Subedi, D.; Archer, S.K.; Morris, F.C.; Oliveira, C.; Kielty, L.; et al. Bacteriophage-resistant Acinetobacter baumannii are resensitized to antimicrobials. Nat. Microbiol. 2021, 6, 157–161. [Google Scholar] [CrossRef]
- Petrovic Fabijan, A.; Iredell, J.; Danis-Wlodarczyk, K.; Kebriaei, R.; Abedon, S.T. Translating phage therapy into the clinic: Recent accomplishments but continuing challenges. PLoS Biol. 2023, 21, e3002119. [Google Scholar] [CrossRef] [PubMed]
- Green, S.I.; Clark, J.R.; Santos, H.H.; Weesner, K.E.; Salazar, K.C.; Aslam, S.; Campbell, J.W.; Doernberg, S.B.; Blodget, E.; Morris, M.I.; et al. A Retrospective, Observational Study of 12 Cases of Expanded-Access Customized Phage Therapy: Production, Characteristics, and Clinical Outcomes. Clin. Infect. Dis. 2023, 77, 1079–1091. [Google Scholar] [CrossRef] [PubMed]
- Pirnay, J.P.; Djebara, S.; Steurs, G.; Griselain, J.; Cochez, C.; De Soir, S.; Glonti, T.; Spiessens, A.; Vanden Berghe, E.; Green, S.; et al. Personalized bacteriophage therapy outcomes for 100 consecutive cases: A multicentre, multinational, retrospective observational study. Nat. Microbiol. 2024, 9, 1434–1453. [Google Scholar] [CrossRef]
- Pirnay, J.P.; Verbeken, G. Magistral Phage Preparations: Is This the Model for Everyone? Clin. Infect. Dis. 2023, 77, S360–S369. [Google Scholar] [CrossRef]
- Clokie, M.R.; Millard, A.D.; Letarov, A.V.; Heaphy, S. Phages in nature. Bacteriophage 2011, 1, 31–45. [Google Scholar] [CrossRef]
- Howard-Varona, C.; Hargreaves, K.R.; Abedon, S.T.; Sullivan, M.B. Lysogeny in nature: Mechanisms, impact and ecology of temperate phages. ISME J. 2017, 11, 1511–1520. [Google Scholar] [CrossRef]
- Meng, L.H.; Ke, F.; Zhang, Q.Y.; Zhao, Z. Functional Analysis of the Endopeptidase and Holin From Planktothrix. agardhii. Cyanophage PaV-LD. Front. Microbiol. 2022, 13, 849492. [Google Scholar] [CrossRef]
- Al-Anany, A.M.; Fatima, R.; Hynes, A.P. Temperate phage-antibiotic synergy eradicates bacteria through depletion of lysogens. Cell Rep. 2021, 35, 109172. [Google Scholar] [CrossRef]
- Al-Anany, A.M.; Fatima, R.; Nair, G.; Mayol, J.T.; Hynes, A.P. Temperate phage-antibiotic synergy across antibiotic classes reveals new mechanism for preventing lysogeny. mBio 2024, 15, e0050424. [Google Scholar] [CrossRef]
- Blum, M.; Andreeva, A.; Florentino, L.C.; Chuguransky, S.R.; Grego, T.; Hobbs, E.; Pinto, B.L.; Orr, A.; Paysan-Lafosse, T.; Ponamareva, I.; et al. InterPro: The protein sequence classification resource in 2025. Nucleic Acids Res. 2025, 53, D444–D456. [Google Scholar] [CrossRef]
- Monteiro, R.; Pires, D.P.; Costa, A.R.; Azeredo, J. Phage Therapy: Going Temperate? Trends Microbiol. 2019, 27, 368–378. [Google Scholar] [CrossRef] [PubMed]
- Gummalla, V.S.; Zhang, Y.; Liao, Y.T.; Wu, V.C.H. The Role of Temperate Phages in Bacterial Pathogenicity. Microorganisms 2023, 11, 54. [Google Scholar] [CrossRef] [PubMed]
- Sander, M.; Schmieger, H. Method for host-independent detection of generalized transducing bacteriophages in natural habitats. Appl. Environ. Microbiol. 2001, 67, 1490–1493. [Google Scholar] [CrossRef] [PubMed]
- Altamirano, F.L.G.; Barr, J.J. Screening for Lysogen Activity in Therapeutically Relevant Bacteriophages. Bio. Protoc. 2021, 11, e3997. [Google Scholar] [CrossRef]
- Mardiana, M.; Teh, S.H.; Tsai, Y.C.; Yang, H.H.; Lin, L.C.; Lin, N.T. Characterization of a novel and active temperate phage vB_AbaM_ABMM1 with antibacterial activity against Acinetobacter baumannii infection. Sci. Rep. 2023, 13, 11347. [Google Scholar] [CrossRef]
- Peng, W.; Zeng, F.; Wu, Z.; Jin, Z.; Li, W.; Zhu, M.; Wang, Q.; Tong, Y.; Chen, L.; Bai, Q. Isolation and genomic analysis of temperate phage 5W targeting multidrug-resistant Acinetobacter baumannii. Arch. Microbiol. 2021, 204, 58. [Google Scholar] [CrossRef]
- Gordillo Altamirano, F.L.; Kostoulias, X.; Subedi, D.; Korneev, D.; Peleg, A.Y.; Barr, J.J. Phage-antibiotic combination is a superior treatment against Acinetobacter baumannii. in a preclinical study. EBioMedicine 2022, 80, 104045. [Google Scholar] [CrossRef]
- Baginska, N.; Harhala, M.A.; Cieslik, M.; Orwat, F.; Weber-Dabrowska, B.; Dabrowska, K.; Gorski, A.; Jonczyk-Matysiak, E. Biological Properties of 12 Newly Isolated Acinetobacter baumannii-Specific Bacteriophages. Viruses 2023, 15, 231. [Google Scholar] [CrossRef]
- Orozco-Ochoa, A.K.; Gonzalez-Gomez, J.P.; Quinones, B.; Castro-Del Campo, N.; Valdez-Torres, J.B.; Chaidez-Quiroz, C. Bacteriophage Indie resensitizes multidrug-resistant Acinetobacter baumannii to antibiotics in vitro. Sci. Rep. 2025, 15, 11578. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, T.; Yu, M.; Chen, Y.L.; Jin, M. The Life Cycle Transitions of Temperate Phages: Regulating Factors and Potential Ecological Implications. Viruses 2022, 14, 1904. [Google Scholar] [CrossRef]
- Gu Liu, C.; Green, S.I.; Min, L.; Clark, J.R.; Salazar, K.C.; Terwilliger, A.L.; Kaplan, H.B.; Trautner, B.W.; Ramig, R.F.; Maresso, A.W. Phage-Antibiotic Synergy Is Driven by a Unique Combination of Antibacterial Mechanism of Action and Stoichiometry. mBio 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- North, O.I.; Brown, E.D. Phage-antibiotic combinations: A promising approach to constrain resistance evolution in bacteria. Ann. N. Y. Acad. Sci. 2021, 1496, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Suh, G.A.; Ferry, T.; Abdel, M.P. Phage Therapy as a Novel Therapeutic for the Treatment of Bone and Joint Infections. Clin. Infect. Dis. 2023, 77, S407–S415. [Google Scholar] [CrossRef]
- Engeman, E.; Freyberger, H.R.; Corey, B.W.; Ward, A.M.; He, Y.; Nikolich, M.P.; Filippov, A.A.; Tyner, S.D.; Jacobs, A.C. Synergistic Killing and Re-Sensitization of Pseudomonas aeruginosa to Antibiotics by Phage-Antibiotic Combination Treatment. Pharmaceuticals 2021, 14, 184. [Google Scholar] [CrossRef]
- Azam, A.H.; Tanji, Y. Bacteriophage-host arm race: An update on the mechanism of phage resistance in bacteria and revenge of the phage with the perspective for phage therapy. Appl. Microbiol. Biotechnol. 2019, 103, 2121–2131. [Google Scholar] [CrossRef]
- Ambroa, A.; Blasco, L.; Lopez, M.; Pacios, O.; Bleriot, I.; Fernandez-Garcia, L.; Gonzalez de Aledo, M.; Ortiz-Cartagena, C.; Millard, A.; Tomas, M. Genomic Analysis of Molecular Bacterial Mechanisms of Resistance to Phage Infection. Front. Microbiol. 2021, 12, 784949. [Google Scholar] [CrossRef]
- Tu, Q.; Pu, M.; Li, Y.; Wang, Y.; Li, M.; Song, L.; Li, M.; An, X.; Fan, H.; Tong, Y. Acinetobacter Baumannii Phages: Past, Present and Future. Viruses 2023, 15, 673. [Google Scholar] [CrossRef]
- Leptihn, S.; Loh, B. Complexity, challenges and costs of implementing phage therapy. Future Microbiol. 2022, 17, 643–646. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, X.; Hua, X.; Yu, Y.; Leptihn, S.; Loh, B. Therapeutic evaluation of the Acinetobacter baumannii. phage Phab24 for clinical use. Virus Res. 2022, 320, 198889. [Google Scholar] [CrossRef]
- Badawy, S.; Baka, Z.A.M.; Abou-Dobara, M.I.; El-Sayed, A.K.A.; Skurnik, M. Biological and molecular characterization of fEg-Eco19, a lytic bacteriophage active against an antibiotic-resistant clinical Escherichia coli isolate. Arch. Virol. 2022, 167, 1333–1341. [Google Scholar] [CrossRef]
- Ross, A.; Ward, S.; Hyman, P. More Is Better: Selecting for Broad Host Range Bacteriophages. Front. Microbiol. 2016, 7, 1352. [Google Scholar] [CrossRef] [PubMed]
- Palma, M.; Qi, B. Advancing Phage Therapy: A Comprehensive Review of the Safety, Efficacy, and Future Prospects for the Targeted Treatment of Bacterial Infections. Infect. Dis. Rep. 2024, 16, 1127–1181. [Google Scholar] [CrossRef] [PubMed]
- Fong, K.; Wong, C.W.Y.; Wang, S.; Delaquis, P. How Broad Is Enough: The Host Range of Bacteriophages and Its Impact on the Agri-Food Sector. Phage 2021, 2, 83–91. [Google Scholar] [CrossRef]
- Lopez-Cuevas, O.; Castro-Del Campo, N.; Leon-Felix, J.; Gonzalez-Robles, A.; Chaidez, C. Characterization of bacteriophages with a lytic effect on various Salmonella serotypes and Escherichia coli O157:H7. Can. J. Microbiol. 2011, 57, 1042–1051. [Google Scholar] [CrossRef]
- Turner, D.; Kropinski, A.M.; Adriaenssens, E.M. A Roadmap for Genome-Based Phage Taxonomy. Viruses 2021, 13. [Google Scholar] [CrossRef]
- Orozco-Ochoa, A.K.; Gonzalez-Gomez, J.P.; Castro-Del Campo, N.; Lira-Morales, J.D.; Martinez-Rodriguez, C.I.; Gomez-Gil, B.; Chaidez, C. Characterization and genome analysis of six novel Vibrio parahaemolyticus phages associated with acute hepatopancreatic necrosis disease (AHPND). Virus Res. 2023, 323, 198973. [Google Scholar] [CrossRef]
- Fastrez, J. Phage lysozymes. EXS 1996, 75, 35–64. [Google Scholar] [CrossRef]
- Petrzik, K. Peptidoglycan Endopeptidase from Novel Adaiavirus Bacteriophage Lyses Pseudomonas aeruginosa Strains as Well as Arthrobacter. globiformis. and A. pascens. Bacteria. Microorganisms. 2023, 11, 1888. [Google Scholar] [CrossRef]
- Kongari, R.; Rajaure, M.; Cahill, J.; Rasche, E.; Mijalis, E.; Berry, J.; Young, R. Phage spanins: Diversity, topological dynamics and gene convergence. BMC Bioinform. 2018, 19, 326. [Google Scholar] [CrossRef]
- Greive, S.J.; Fung, H.K.; Chechik, M.; Jenkins, H.T.; Weitzel, S.E.; Aguiar, P.M.; Brentnall, A.S.; Glousieau, M.; Gladyshev, G.V.; Potts, J.R.; et al. DNA recognition for virus assembly through multiple sequence-independent interactions with a helix-turn-helix motif. Nucleic Acids Res. 2016, 44, 776–789. [Google Scholar] [CrossRef]
- Zinoviev, V.V.; Evdokimov, A.A.; Gorbunov, Y.A.; Malygin, E.G.; Kossykh, V.G.; Hattman, S. Phage T4 DNA [N6-adenine] methyltransferase: Kinetic studies using oligonucleotides containing native or modified recognition sites. Biol. Chem. 1998, 379, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Xu, D.; Huang, Y.; Zhu, X.; Rui, M.; Wan, T.; Zheng, X.; Shen, Y.; Chen, X.; Ma, K.; et al. Structural and functional characterization of deep-sea thermophilic bacteriophage GVE2 HNH endonuclease. Sci. Rep. 2017, 7, 42542. [Google Scholar] [CrossRef]
- Mascolo, E.; Adhikari, S.; Caruso, S.M.; deCarvalho, T.; Folch Salvador, A.; Serra-Sagrista, J.; Young, R.; Erill, I.; Curtis, P.D. The transcriptional regulator CtrA controls gene expression in Alphaproteobacteria phages: Evidence for a lytic deferment pathway. Front. Microbiol. 2022, 13, 918015. [Google Scholar] [CrossRef]
- Choi, Y.J.; Kim, S.; Shin, M.; Kim, J. Synergistic Antimicrobial Effects of Phage vB_AbaSi_W9 and Antibiotics against Acinetobacter baumannii. Infection. Antibiotics 2024, 13, 680. [Google Scholar] [CrossRef]
- Donnelly-Wu, M.K.; Jacobs, W.R., Jr.; Hatfull, G.F. Superinfection immunity of mycobacteriophage L5: Applications for genetic transformation of mycobacteria. Mol. Microbiol. 1993, 7, 407–417. [Google Scholar] [CrossRef]
- Elois, M.A.; Silva, R.D.; Pilati, G.V.T.; Rodriguez-Lazaro, D.; Fongaro, G. Bacteriophages as Biotechnological Tools. Viruses 2023, 15, 349. [Google Scholar] [CrossRef]
- Mavrich, T.N.; Hatfull, G.F. Evolution of Superinfection Immunity in Cluster A Mycobacteriophages. mBio 2019, 10. [Google Scholar] [CrossRef]
- Fowoyo, P.T. Phage Therapy: Clinical Applications, Efficacy, and Implementation Hurdles. Open Microbiol. J. 2024, 18. [Google Scholar] [CrossRef]
- Strathdee, S.A.; Hatfull, G.F.; Mutalik, V.K.; Schooley, R.T. Phage therapy: From biological mechanisms to future directions. Cell 2023, 186, 17–31. [Google Scholar] [CrossRef]
- Zhang, S.; Di, L.; Qi, Y.; Qian, X.; Wang, S. Treatment of infections caused by carbapenem-resistant Acinetobacter baumannii. Front. Cell. Infect. Microbiol. 2024, 14, 1395260. [Google Scholar] [CrossRef]
- Gordillo Altamirano, F.L.; Barr, J.J. Phage Therapy in the Postantibiotic Era. Clin. Microbiol. Rev. 2019, 32. [Google Scholar] [CrossRef] [PubMed]
- Merabishvili, M.; Pirnay, J.P.; De Vos, D. Chapter 4 Guidelines to Compose an Ideal Bacteriophage Cocktail. In Bacteriophage Therapy; Springer: New York, NY, USA, 2024. [Google Scholar] [CrossRef]
- Huang, L.; Huang, S.; Jiang, L.; Tan, J.; Yan, X.; Gou, C.; Chen, X.; Xiang, L.; Wang, D.; Huang, G.; et al. Characterisation and sequencing of the novel phage Abp95, which is effective against multi-genotypes of carbapenem-resistant Acinetobacter baumannii. Sci. Rep. 2023, 13, 188. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Liu, X.; Zhou, J.; Wang, L.; Wang, Q.; Qi, X.; Liu, J.; Zhao, D.; Hsiang, T.; Jiang, Y. Isolation, characterization and therapeutic evaluation of a new Acinetobacter virus Abgy202141 lysing Acinetobacter baumannii. Front. Microbiol. 2024, 15, 1379400. [Google Scholar] [CrossRef] [PubMed]
- Ndiaye, I.; Debarbieux, L.; Sow, O.; Sambe Ba, B.; Diagne, M.M.; Cisse, A.; Fall, C.; Dieye, Y.; Dia, N.; Constantin de Magny, G.; et al. Isolation and characterization of Acinetobacter phage vAbaIN10 active against carbapenem-resistant Acinetobacter baumannii (CRAB) isolates from healthcare-associated infections in Dakar, Senegal. J. Glob. Antimicrob. Resist. 2025, 41, 151–158. [Google Scholar] [CrossRef]
- Badawy, S.; Pajunen, M.I.; Haiko, J.; Baka, Z.A.M.; Abou-Dobara, M.I.; El-Sayed, A.K.A.; Skurnik, M. Identification and Functional Analysis of Temperate Siphoviridae Bacteriophages of Acinetobacter baumannii. Viruses 2020, 12, 604. [Google Scholar] [CrossRef]
- Abedon, S.T. Bacteriophage Adsorption: Likelihood of Virion Encounter with Bacteria and Other Factors Affecting Rates. Antibiotics 2023, 12, 723. [Google Scholar] [CrossRef]
- Schwarzer, D.; Stummeyer, K.; Gerardy-Schahn, R.; Muhlenhoff, M. Characterization of a novel intramolecular chaperone domain conserved in endosialidases and other bacteriophage tail spike and fiber proteins. J. Biol. Chem. 2007, 282, 2821–2831. [Google Scholar] [CrossRef]
- Roshini, J.; Patro, L.P.P.; Sundaresan, S.; Rathinavelan, T. Structural diversity among Acinetobacter baumannii. K-antigens and its implication in the in silico serotyping. Front. Microbiol. 2023, 14, 1191542. [Google Scholar] [CrossRef]
- Singh, J.K.; Adams, F.G.; Brown, M.H. Diversity and Function of Capsular Polysaccharide in Acinetobacter baumannii. Front. Microbiol. 2018, 9, 3301. [Google Scholar] [CrossRef]
- Evseev, P.V.; Sukhova, A.S.; Tkachenko, N.A.; Skryabin, Y.P.; Popova, A.V. Lytic Capsule-Specific Acinetobacter Bacteriophages Encoding Polysaccharide-Degrading Enzymes. Viruses 2024, 16, 771. [Google Scholar] [CrossRef]
- Falah, F.; Shokoohizadeh, L.; Adabi, M. Molecular identification and genotyping of Acinetobacter baumannii. isolated from burn patients by PCR and ERIC-PCR. Scars Burn. Heal. 2019, 5, 2059513119831369. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.J.; Kim, M.H.; Cho, M.S.; Kim, B.K.; Kim, J.Y.; Kim, C.; Park, D.S. Improved PCR for identification of Pseudomonas aeruginosa. Appl. Microbiol. Biotechnol. 2013, 97, 3643–3651. [Google Scholar] [CrossRef] [PubMed]
- Kwon, K.H.; Hwang, S.Y.; Moon, B.Y.; Park, Y.K.; Shin, S.; Hwang, C.Y.; Park, Y.H. Occurrence of antimicrobial resistance and virulence genes, and distribution of enterococcal clonal complex 17 from animals and human beings in Korea. J. Vet. Diagn. Investig. 2012, 24, 924–931. [Google Scholar] [CrossRef]
- Hamdan-Partida, A.; González García, S.; Bustos-Martínez, J. Identificación de Staphylococcus aureus utilizando como marcadores los genes nucA y femB. Ciencias Clínicas 2015, 16, 37–41. [Google Scholar] [CrossRef]
- Badger-Emeka, L.I.; Emeka, P.M. Genetic fingerprinting and profile analysis of virulence genes in XDR clinical isolates of Klebsiella Pneumoniae. Eur. Rev. Med. Pharmacol. Sci. 2022, 26, 5233–5244. [Google Scholar] [CrossRef]
- Muhammed, N.a.A.; Farhan, M.B.; Shabeeb, Z.A. Detection of luxS Gene in Serratia marcescens and rpoS Gene in Enterobacter cloacae Isolates Using PCR Reaction. J. Al-Nahrain Univ. Sci. 2018, 21, 115–120. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing; CLSI Document M100; Clinical and Laboratory Standards Institute. Clinical and Laboratory Standards Institute: Malvern, PA, USA, 2020. [Google Scholar]
- Liu, Y.; Mi, Z.; Mi, L.; Huang, Y.; Li, P.; Liu, H.; Yuan, X.; Niu, W.; Jiang, N.; Bai, C.; et al. Identification and characterization of capsule depolymerase Dpo48 from Acinetobacter baumannii phage IME200. PeerJ 2019, 7, e6173. [Google Scholar] [CrossRef]
- Wang, Z.; Yang, X.; Wang, H.; Wang, S.; Fang, R.; Li, X.; Xing, J.; Wu, Q.; Li, Z.; Song, N. Characterization and efficacy against carbapenem-resistant Acinetobacter baumannii of a novel Friunavirus phage from sewage. Front. Cell. Infect. Microbiol. 2024, 14, 1382145. [Google Scholar] [CrossRef]
- Mirzaei, M.K.; Nilsson, A.S. Correction: Isolation of phages for phage therapy: A comparison of spot tests and efficiency of plating analyses for determination of host range and efficacy. PLoS ONE 2015, 10, e0127606. [Google Scholar] [CrossRef]
- Sambrook, J.; Russell, D.W. Extraction of Bacteriophage lambda DNA from Large-scale Cultures Using Proteinase K and SDS. Cold Spring Harb. Protoc. 2006, 2006, pdb.prot3972. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed]
- Chan, P.P.; Lowe, T.M. tRNAscan-SE: Searching for tRNA Genes in Genomic Sequences. Methods Mol. Biol. 2019, 1962, 1–14. [Google Scholar] [CrossRef]
- Carver, T.; Harris, S.R.; Berriman, M.; Parkhill, J.; McQuillan, J.A. Artemis: An integrated platform for visualization and analysis of high-throughput sequence-based experimental data. Bioinformatics 2012, 28, 464–469. [Google Scholar] [CrossRef]
- Carver, T.; Thomson, N.; Bleasby, A.; Berriman, M.; Parkhill, J. DNAPlotter: Circular and linear interactive genome visualization. Bioinformatics 2009, 25, 119–120. [Google Scholar] [CrossRef]
- Gilchrist, C.L.M.; Chooi, Y.H. clinker & clustermap.js: Automatic generation of gene cluster comparison figures. Bioinformatics 2021, 37, 2473–2475. [Google Scholar] [CrossRef]
- Gonzalez-Gomez, J.P.; Rodriguez-Arellano, S.N.; Gomez-Gil, B.; Vergara-Jimenez, M.J.; Chaidez, C. Genomic and biological characterization of bacteriophages against Enterobacter cloacae, a high-priority pathogen. Virology 2024, 595, 110100. [Google Scholar] [CrossRef]
- Park, D.W.; Park, J.H. Characterization and Food Application of the Novel Lytic Phage BECP10: Specifically Recognizes the O-polysaccharide of Escherichia coli O157:H7. Viruses 2021, 13, 1469. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Host Range (Spot Test)/Efficiency of Plating (EOP) a | Plaque Diameter (mm) c | |||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Phage Strain ID | Acinetobacter baumannii Strain | ESKAPE Pathogens | GNPs b | |||||||||||||||||||||||
| AbAK01 | AbAK02 | AbAK03 | AbAK04 | AbAK05 | AbAK06 | AbAK07 | AbAK08 | AbAK09 | AbAK10 | PaAK07 | PaAK08 | PaAK10 | PaAK11 | KpAK11 | KpAK13 | KpAK14 | KpAK15 | KpAK16 | KpAK17 | KpAK18 | EfAK11 | SaAK16 | EcoAK05 | EcoAK06 | ||
| AKO8a | - | - | - | +++ (1)H | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| 89P13 | - | - | - | +++ (1)H | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| κeane | - | - | - | +++ (1)H | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| C∞ldPlαy | - | - | - | +++ (1)H | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| MCR | - | - | - | +++ (0.7)H | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| PS118 | - | - | - | +++ (1)H | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 2 |
| KOL | - | - | - | +++ (0.4)M | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| B612 | - | - | - | +++ (1)H | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| Green-8 | - | - | - | +++ (0.1)L | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| PB14N | - | - | - | +++ (1)H | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| DA78 | - | - | - | +++ (1)H | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| TAM10 | - | - | - | +++ (1)H | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| IDQ7 | - | - | - | +++ (1)H | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 2 |
| PN1412 | - | - | - | +++ (1)H | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 2 |
| CRL20 | - | - | - | +++ (1)H | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| MRL18 | - | - | - | +++ (1)H | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 2 |
| CIM23 | - | - | - | +++ (1)H | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| AM | - | - | - | +++ (1)H | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| Pyro | - | - | - | +++ (0.4)M | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| dakota | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| Août | - | - | - | +++ (0.4)M | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| Δspirit | - | - | - | +++ (1)H | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| Phage Common Name | Phage Taxonomy Name | Genome Length (bp) | GC Content (%) | Total No. of ORFs a | No. of ORFs with Known Functions | tRNAs b | Antibiotic Resistance Genes | Virulence Genes | Lysogeny Genes |
|---|---|---|---|---|---|---|---|---|---|
| CIM23 | vB_AbaS_CIM23 | 44,512 | 50.6 | 145 | 35 | 0 | 0 | 0 | 1 |
| B612 | vB_AbaS_B612 | 44,702 | 50.4 | 146 | 34 | 0 | 0 | 0 | 1 |
| 89P13 | vB_AbaS_89P13 | 43,176 | 50.4 | 141 | 35 | 0 | 0 | 0 | 1 |
| AKO8a | vB_AbaS_AKO8a | 67,450 | 49.9 | 157 | 149 | 0 | 0 | 0 | 0 |
| PS118 | vB_AbaS_PS118 | 46,430 | 50.5 | 161 | 38 | 0 | 0 | 0 | 1 |
| MCR | vB_AbaS_MCR | 45,205 | 50.4 | 139 | 34 | 0 | 0 | 0 | 0 |
| CRL20 | vB_AbaS_CRL20 | 45,335 | 50.2 | 147 | 39 | 0 | 0 | 0 | 0 |
| IDQ7 | vB_AbaS_IDQ7 | 44,703 | 50.3 | 148 | 38 | 0 | 0 | 0 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Orozco-Ochoa, A.K.; Quiñones, B.; González-Gómez, J.P.; Castro-del Campo, N.; Valdez-Torres, J.B.; Chaidez-Quiroz, C. Genomic and Functional Characterization of Novel Phages Targeting Multidrug-Resistant Acinetobacter baumannii. Int. J. Mol. Sci. 2025, 26, 6141. https://doi.org/10.3390/ijms26136141
Orozco-Ochoa AK, Quiñones B, González-Gómez JP, Castro-del Campo N, Valdez-Torres JB, Chaidez-Quiroz C. Genomic and Functional Characterization of Novel Phages Targeting Multidrug-Resistant Acinetobacter baumannii. International Journal of Molecular Sciences. 2025; 26(13):6141. https://doi.org/10.3390/ijms26136141
Chicago/Turabian StyleOrozco-Ochoa, Alma Karen, Beatriz Quiñones, Jean Pierre González-Gómez, Nohelia Castro-del Campo, José Benigno Valdez-Torres, and Cristóbal Chaidez-Quiroz. 2025. "Genomic and Functional Characterization of Novel Phages Targeting Multidrug-Resistant Acinetobacter baumannii" International Journal of Molecular Sciences 26, no. 13: 6141. https://doi.org/10.3390/ijms26136141
APA StyleOrozco-Ochoa, A. K., Quiñones, B., González-Gómez, J. P., Castro-del Campo, N., Valdez-Torres, J. B., & Chaidez-Quiroz, C. (2025). Genomic and Functional Characterization of Novel Phages Targeting Multidrug-Resistant Acinetobacter baumannii. International Journal of Molecular Sciences, 26(13), 6141. https://doi.org/10.3390/ijms26136141