


Comparative Analysis of Melatonin and Polydeoxyribonucleotide: Possible Benefits of Co-Treatment Effects and Potential Synergistic Applicability


Abstract
1. Introduction
2. Mechanisms of Action and Effects of Melatonin
2.1. Melatonin: A Well-Studied and Versatile Regulatory Molecule
2.2. Melatonin as a Medicine or Dietary Supplement
2.3. Mechanisms of Action and Effects of Melatonin
2.3.1. Receptor-Mediated Actions
2.3.2. Antioxidant Activity
2.3.3. Anti-Inflammatory Effects
2.3.4. Modulation of Inflammasome Activity
2.3.5. Analgesic Properties
2.3.6. Anti-Cancer Effects
2.3.7. Collagen Synthesis
2.3.8. Wound Healing
2.3.9. Alleviation of Neuropathic Pain
3. Mechanisms of Action and Effects of PDRN
3.1. PDRN: A DNA-Derived Biologic Agent or Biopolymer
3.2. Mechanisms of Action and Effects of PDRN
3.2.1. Adenosine A2A Receptor Activation
3.2.2. Inhibition of MAPK Signaling Pathway
3.2.3. Enhancement of Collagen Production
3.2.4. Promotion of Angiogenesis
3.2.5. Stimulation of Tissue Repair
3.2.6. Melanin Reduction
4. Comparison of the Effects of Melatonin and PDRN

{kind=link}
| Melatonin [References 1] | PDRN [References 1] | |
|---|---|---|
| Main roles |
|
|
| Mechanisms of action |
|
|
| Main effects |
|
|
| Applications |
|
|
| Main sources of supply |
|
|
5. Comprehensive Research Framework
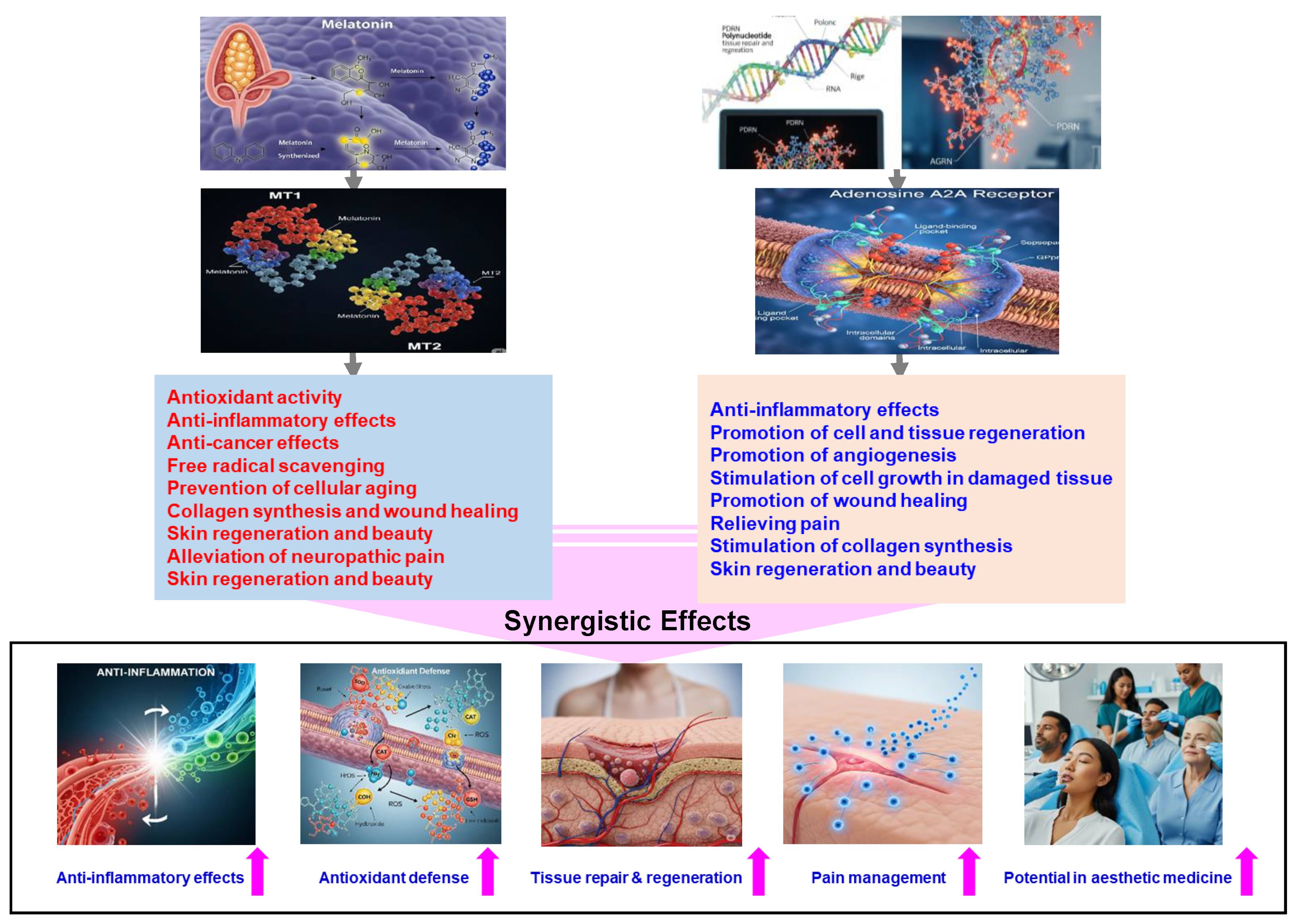
5.1. Potential Synergistic Effects
- Enhanced Anti-inflammatory Response: Melatonin is known to suppress pro-inflammatory cytokines, while PDRN activates adenosine A2A receptors to reduce inflammation. The combined action of both compounds could potentially result in a more potent anti-inflammatory effect.
- Augmented Anti-Inflammatory Effects: Both melatonin and PDRN independently exhibit anti-inflammatory properties. Utilizing them together could potentially provide a more comprehensive reduction in inflammation, which would be beneficial in conditions characterized by excessive inflammatory responses.
- Comprehensive Inflammatory Pathway Modulation: Melatonin acts by inhibiting inflammasome activity, whereas PDRN suppresses MAPK signaling. Together, they can address inflammation at multiple regulatory points, potentially leading to a more effective overall anti-inflammatory response.
- Enhanced Antioxidant Defense: Melatonin’s ability to boost antioxidant enzyme activity combined with PDRN’s role in promoting tissue repair through VEGF induction could synergize to reduce oxidative stress more effectively and facilitate healing processes.
- Accelerated Tissue Repair: Melatonin’s antioxidant properties protect tissues from oxidative damage, while PDRN promotes tissue regeneration. Together, these compounds may enhance healing processes in conditions such as skin injuries or degenerative diseases.
- Enhanced Tissue Regeneration: Melatonin’s antioxidant properties, coupled with PDRN’s promotion of angiogenesis and cell proliferation, could synergistically accelerate tissue repair processes.
- Improved Pain Management: Both melatonin and PDRN possess analgesic properties: melatonin modulates pain perception, while PDRN addresses inflammation-induced pain. Utilizing them in combination could potentially provide comprehensive pain relief.
- Potential in Esthetic Medicine: Given PDRN’s capacity to reduce melanin synthesis and melatonin’s regulatory effects on skin cells, their combination might offer innovative solutions in cosmetic treatments aimed at skin rejuvenation and mitigating pigmentation disorders.
5.2. Potential Clinical Applications
- Chronic Pain Conditions: The integration of melatonin’s analgesic effects with PDRN’s anti-inflammatory properties may offer effective relief in chronic pain syndromes.
- Degenerative Joint Diseases: In conditions like osteoarthritis, where inflammation and tissue degeneration prevail, this combination could reduce inflammation and promote cartilage repair.
- Esthetic Medicine: For skin rejuvenation, the antioxidant effect of melatonin combined with PDRN’s collagen-stimulating properties may enhance skin elasticity and reduce signs of aging.
5.3. Future Directions
- Dose Optimization: Determining the optimal dosages for combination therapy to maximize benefits and minimize potential side effects.
- Preclinical Studies: Utilizing animal models to assess the combined effects of melatonin and PDRN on tissue repair, inflammation, and markers of oxidative stress.
- Mechanistic Studies: Investigating the molecular and signaling pathways, such as SIRT1, Nrf2, and NF-κB, involved in their combined action to better understand the synergistic effects.
- Clinical Trials: Conducting randomized controlled trials to evaluate the safety and efficacy of combined melatonin and PDRN therapy in various patient populations, focusing on conditions such as neurodegenerative diseases, ischemic injuries, and dermatological conditions to comprehensively assess efficacy and safety.
- Delivery Systems: Developing advanced delivery mechanisms, such as nanocarriers, to enhance the bioavailability and targeted delivery of melatonin and PDRN, thereby maximizing therapeutic outcomes.
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| PDRN | Polydeoxyribonucleotide |
| VEGF | Vascular endothelial growth factor |
| SCN | Hypothalamic suprachiasmatic nucleus |
| cAMP | Cyclic adenosine monophosphate |
| cGMP | Cyclic guanosine monophosphate |
| AhR | Aryl hydrocarbon receptor |
| PPARγ | Peroxisome proliferator-activated receptor γ |
| CREB | cAMP response element-binding protein |
| NF-κB | Nuclear factor-kappa B |
| SOD | Superoxide dismutase |
| CAT | Catalase |
| GPx | Glutathione peroxidase |
| GSH | Glutathione |
| MDA | Malondialdehyde |
| IL-1β | Interleukin-1 beta |
| IL-6 | Interleukin-6 |
| IL-8 | Interleukin-8 |
| TNF-α | Tumor necrosis factor-alpha |
| CRP | C-reactive protein |
| TBI | Traumatic brain injury |
| IVDD | Intervertebral disk disease |
| α7 nAChRs | α7 nicotinic acetylcholine receptors |
| ACC | Anterior cingulate cortex |
| CDKs | Cyclin-dependent kinases |
| HIF-1α | Hypoxia-inducible factor-1α |
| miRNAs | microRNAs |
| EMT | Epithelial–mesenchymal transition |
| MMPs | Markers and matrix metalloproteinases |
| EGF | Epidermal growth factor |
| L-PDRN | Microbial-derived PDRN |
| FAK | Focal adhesion kinase |
| AKT | protein kinase B |
| MAPK | Mitogen-activated protein kinase |
| DMPX | 3,7-Dimethyl-1-propargylxanthine |
| ECM | Extracellular matrix |
| JAK | Janus Kinase |
| STAT | Signal Transducer and Activator of Transcription |
References
- Singh, S.K.; Mohammed, A.; Alghamdi, O.A.; Husain, S.M. New approaches for targeting drug resistance through drug combination. In Combination Therapy Against Multidrug Resistance; Academic Press: Cambridge, MA, USA; Elsevier Inc.: Amsterdam, The Netherlands, 2020; pp. 221–246. [Google Scholar]
- Plana, D.; Palmer, A.C.; Sorger, P.K. Independent Drug Action in Combination Therapy: Implications for Precision Oncology. Cancer Discov. 2022, 12, 606–624. [Google Scholar] [CrossRef] [PubMed]
- Kopustinskiene, D.M.; Bernatoniene, J. Molecular Mechanisms of Melatonin-Mediated Cell Protection and Signaling in Health and Disease. Pharmaceutics 2021, 13, 129. [Google Scholar] [CrossRef]
- Alshehri, F.S.; Althobaiti, Y.S. A review of the potential use of melatonin in cancer treatment: Data analysis from Clinicaltrials.gov. Medicine 2024, 103, e40517. [Google Scholar] [CrossRef]
- Ko, I.G.; Kim, S.E.; Jin, J.J.; Hwang, L.; Ji, E.S.; Kim, C.J.; Han, J.H.; Hong, I.T.; Kwak, M.S.; Yoon, J.Y.; et al. Combination therapy with polydeoxyribonucleotide and proton pump inhibitor enhances therapeutic effectiveness for gastric ulcer in rats. Life Sci. 2018, 203, 12–19. [Google Scholar] [CrossRef]
- Bitto, A.; Polito, F.; Altavilla, D.; Minutoli, L.; Migliorato, A.; Squadrito, F. Polydeoxyribonucleotide (PDRN) restores blood flow in an experimental model of peripheral artery occlusive disease. J. Vasc. Surg. 2008, 48, 1292–1300. [Google Scholar] [CrossRef]
- Lee, K.W.A.; Chan, K.W.L.; Lee, A.; Lee, C.H.; Wan, J.; Wong, S.; Yi, K.H. Polynucleotides in Aesthetic Medicine: A Review of Current Practices and Perceived Effectiveness. Int. J. Mol. Sci. 2024, 25, 8224. [Google Scholar] [CrossRef]
- Ebihara, S.; Marks, T.; Hudson, D.J.; Menaker, M. Genetic control of melatonin synthesis in the pineal gland of the mouse. Science 1986, 231, 491–493. [Google Scholar] [CrossRef]
- Klein, D.C.; Coon, S.L.; Roseboom, P.H.; Weller, J.L.; Bernard, M.; Gastel, J.A.; Zatz, M.; Iuvone, P.M.; Rodriguez, I.R.; Bégay, V.; et al. The melatonin rhythm-generating enzyme: Molecular regulation of serotonin N-acetyltransferase in the pineal gland. Recent Prog. Horm. Res. 1997, 52, 307–357. [Google Scholar]
- Reiter, R.J.; Tan, D.X.; Fuentes-Broto, L. Melatonin: A multitasking molecule. Prog. Brain Res. 2010, 181, 127–151. [Google Scholar] [CrossRef]
- Comai, S.; Gobbi, G. Melatonin, Melatonin Receptors and Sleep: Moving Beyond Traditional Views. J. Pineal Res. 2024, 76, e13011. [Google Scholar] [CrossRef] [PubMed]
- Vanecek, J. Cellular mechanisms of melatonin action. Physiol. Rev. 1998, 78, 687–721. [Google Scholar] [CrossRef] [PubMed]
- Erland, L.A.; Saxena, P.K. Melatonin Natural Health Products and Supplements: Presence of Serotonin and Significant Variability of Melatonin Content. J. Clin. Sleep Med. 2017, 13, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Grigg-Damberger, M.M.; Ianakieva, D. Poor Quality Control of Over-the-Counter Melatonin: What They Say Is Often Not What You Get. J. Clin. Sleep Med. 2017, 13, 163–165. [Google Scholar] [CrossRef] [PubMed]
- Sateia, M.J.; Buysse, D.J.; Krystal, A.D.; Neubauer, D.N.; Heald, J.L. Clinical Practice Guideline for the Pharmacologic Treatment of Chronic Insomnia in Adults: An American Academy of Sleep Medicine Clinical Practice Guideline. J. Clin. Sleep Med. 2017, 13, 307–349. [Google Scholar] [CrossRef]
- Zhang, Z.; Xue, P.; Bendlin, B.B.; Zetterberg, H.; De Felice, F.; Tan, X.; Benedict, C. Melatonin: A potential nighttime guardian against Alzheimer’s. Mol. Psychiatry 2025, 30, 237–250. [Google Scholar] [CrossRef]
- Zanif, U.; Lai, A.S.; Parks, J.; Roenningen, A.; McLeod, C.B.; Ayas, N.; Wang, X.; Lin, Y.; Zhang, J.J.; Bhatti, P. Melatonin supplementation and oxidative DNA damage repair capacity among night shift workers: A randomised placebo-controlled trial. Occup. Environ. Med. 2025, 82, 1–6. [Google Scholar] [CrossRef]
- Boivin, D.B.; Boudreau, P.; Kosmadopoulos, A. Disturbance of the Circadian System in Shift Work and Its Health Impact. J. Biol. Rhythm. 2022, 37, 3–28. [Google Scholar] [CrossRef]
- Di Vincenzo, A.; Zabeo, E.; Purificati, C.; Rossato, M. Growth Hormone Increase Induced by Oral Administration of Melatonin in a Young Woman With Sleep Disturbances. JCEM Case Rep. 2025, 3, luaf006. [Google Scholar] [CrossRef]
- Arnao, M.B.; Giraldo-Acosta, M.; Castejón-Castillejo, A.; Losada-Lorán, M.; Sánchez-Herrerías, P.; El Mihyaoui, A.; Cano, A.; Hernández-Ruiz, J. Melatonin from Microorganisms, Algae, and Plants as Possible Alternatives to Synthetic Melatonin. Metabolites 2023, 13, 72. [Google Scholar] [CrossRef]
- Kim, T.K.; Slominski, R.M.; Pyza, E.; Kleszczynski, K.; Tuckey, R.C.; Reiter, R.J.; Holick, M.F.; Slominski, A.T. Evolutionary formation of melatonin and vitamin D in early life forms: Insects take centre stage. Biol. Rev. Camb. Philos. Soc. 2024, 99, 1772–1790. [Google Scholar] [CrossRef]
- Minich, D.M.; Henning, M.; Darley, C.; Fahoum, M.; Schuler, C.B.; Frame, J. Is Melatonin the “Next Vitamin D”?: A Review of Emerging Science, Clinical Uses, Safety, and Dietary Supplements. Nutrients 2022, 14, 3934. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, J.L.; Dubocovich, M.L. Melatonin multifaceted pharmacological actions on melatonin receptors converging to abrogate COVID-19. J. Pineal Res. 2021, 71, e12732. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, H.H.; Cecon, E.; Nureki, O.; Rivara, S.; Jockers, R. Melatonin receptor structure and signaling. J. Pineal Res. 2024, 76, e12952. [Google Scholar] [CrossRef] [PubMed]
- McGowan, N.M.; Kim, D.S.; de Andres Crespo, M.; Bisdounis, L.; Kyle, S.D.; Saunders, K.E.A. Hypnotic and Melatonin/Melatonin-Receptor Agonist Treatment in Bipolar Disorder: A Systematic Review and Meta-Analysis. CNS Drugs 2022, 36, 345–363. [Google Scholar] [CrossRef]
- Klosen, P. Thirty-seven years of MT1 and MT2 melatonin receptor localization in the brain: Past and future challenges. J. Pineal Res. 2024, 76, e12955. [Google Scholar] [CrossRef]
- Wu, N.; Carpino, G.; Ceci, L.; Baiocchi, L.; Francis, H.; Kennedy, L.; Zhou, T.; Chen, L.; Sato, K.; Kyritsi, K.; et al. Melatonin receptor 1A, but not 1B, knockout decreases biliary damage and liver fibrosis during cholestatic liver injury. Hepatology 2022, 75, 797–813. [Google Scholar] [CrossRef]
- Liang, L.; Zeng, T.; Zhao, Y.; Lu, R.; Guo, B.; Xie, R.; Tang, W.; Zhang, L.; Mao, Z.; Yang, X.; et al. Melatonin pretreatment alleviates the long-term synaptic toxicity and dysmyelination induced by neonatal Sevoflurane exposure via MT1 receptor-mediated Wnt signaling modulation. J. Pineal Res. 2021, 71, e12771. [Google Scholar] [CrossRef]
- Posa, L.; Lopez-Canul, M.; Rullo, L.; De Gregorio, D.; Dominguez-Lopez, S.; Kaba Aboud, M.; Caputi, F.F.; Candeletti, S.; Romualdi, P.; Gobbi, G. Nociceptive responses in melatonin MT2 receptor knockout mice compared to MT1 and double MT1 /MT2 receptor knockout mice. J. Pineal Res. 2020, 69, e12671. [Google Scholar] [CrossRef]
- Slominski, A.T.; Kim, T.K.; Slominski, R.M.; Song, Y.; Qayyum, S.; Placha, W.; Janjetovic, Z.; Kleszczyński, K.; Atigadda, V.; Song, Y.; et al. Melatonin and Its Metabolites Can Serve as Agonists on the Aryl Hydrocarbon Receptor and Peroxisome Proliferator-Activated Receptor Gamma. Int. J. Mol. Sci. 2023, 24, 15496. [Google Scholar] [CrossRef]
- Slominski, A.T.; Kim, T.K.; Janjetovic, Z.; Slominski, R.M.; Ganguli-Indra, G.; Athar, M.; Indra, A.K.; Reiter, R.J.; Kleszczyński, K. Melatonin and the Skin: Current Progress and Perspectives for Human Health. J. Invest. Dermatol. 2025, 145, 1345–1360.e2. [Google Scholar] [CrossRef]
- Tan, D.X.; Reiter, R.J.; Manchester, L.C.; Yan, M.T.; El-Sawi, M.; Sainz, R.M.; Mayo, J.C.; Kohen, R.; Allegra, M.; Hardeland, R. Chemical and physical properties and potential mechanisms: Melatonin as a broad spectrum antioxidant and free radical scavenger. Curr. Top. Med. Chem. 2002, 2, 181–197. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.X.; Terron, M.P.; Flores, L.J.; Czarnocki, Z. Melatonin and its metabolites: New findings regarding their production and their radical scavenging actions. Acta Biochim. Pol. 2007, 54, 1–9. [Google Scholar] [PubMed]
- Bonmatí-Carrión, M.Á.; Rol, M.A. Melatonin as a Mediator of the Gut Microbiota-Host Interaction: Implications for Health and Disease. Antioxidants 2023, 13, 34. [Google Scholar] [CrossRef]
- Monteiro, K.K.A.C.; Shiroma, M.E.; Damous, L.L.; Simões, M.J.; Simões, R.D.S.; Cipolla-Neto, J.; Baracat, E.C.; Soares, J.M., Jr. Antioxidant Actions of Melatonin: A Systematic Review of Animal Studies. Antioxidants 2024, 13, 439. [Google Scholar] [CrossRef]
- Korkmaz, A.; Reiter, R.J.; Topal, T.; Manchester, L.C.; Oter, S.; Tan, D.X. Melatonin: An established antioxidant worthy of use in clinical trials. Mol. Med. 2009, 15, 43–50. [Google Scholar] [CrossRef]
- Cho, J.H.; Bhutani, S.; Kim, C.H.; Irwin, M.R. Anti-inflammatory effects of melatonin: A systematic review and meta-analysis of clinical trials. Brain Behav. Immun. 2021, 93, 245–253. [Google Scholar] [CrossRef]
- Joseph, T.T.; Schuch, V.; Hossack, D.J.; Chakraborty, R.; Johnson, E.L. Melatonin: The placental antioxidant and anti-inflammatory. Front. Immunol. 2024, 15, 1339304. [Google Scholar] [CrossRef]
- Sieminski, M.; Reimus, M.; Kałas, M.; Stępniewska, E. Antioxidant and Anti-Inflammatory Properties of Melatonin in Secondary Traumatic Brain Injury. Antioxidants 2024, 14, 25. [Google Scholar] [CrossRef]
- Schaefer, K.R.; Rogers, S.; Faber, Z.; Kelley, T.J. Efficacy of melatonin treatment in a cystic fibrosis mouse model of airway infection. Sci. Rep. 2025, 15, 1849. [Google Scholar] [CrossRef]
- Chen, F.; Jiang, G.; Liu, H.; Li, Z.; Pei, Y.; Wang, H.; Pan, H.; Cui, H.; Long, J.; Wang, J.; et al. Melatonin alleviates intervertebral disc degeneration by disrupting the IL-1β/NF-κB-NLRP3 inflammasome positive feedback loop. Bone Res. 2020, 8, 10. [Google Scholar] [CrossRef]
- Zhao, K.; Zhang, Y.; Liao, Z.; Zhang, W.; Li, G.; Shi, P.; Cheng, Z.; Chen, Y.; Li, S.; Wang, K.; et al. Melatonin mitigates intervertebral disc degeneration by suppressing NLRP3 inflammasome activation via the EGR1/DDX3X pathway. FASEB J. 2024, 38, e70143. [Google Scholar] [CrossRef] [PubMed]
- Saha, M.; Manna, K.; Das Saha, K. Melatonin Suppresses NLRP3 Inflammasome Activation via TLR4/NF-κB and P2X7R Signaling in High-Fat Diet-Induced Murine NASH Model. J. Inflamm. Res. 2022, 15, 3235–3258. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Suo, Z.; Wang, Z.; Cao, J.; Dong, Y.; Chen, Y. Melatonin modulates neuroinflammatory response and microglial activation in mice exposed to dim blue light at night. Front. Pharmacol. 2024, 15, 1416350. [Google Scholar] [CrossRef]
- Farré-Alins, V.; Narros-Fernández, P.; Palomino-Antolín, A.; Decouty-Pérez, C.; Lopez-Rodriguez, A.B.; Parada, E.; Muñoz-Montero, A.; Gómez-Rangel, V.; López-Muñoz, F.; Ramos, E.; et al. Melatonin Reduces NLRP3 Inflammasome Activation by Increasing α7 nAChR-Mediated Autophagic Flux. Antioxidants 2020, 9, 1299. [Google Scholar] [CrossRef]
- Nopparat, C.; Boontor, A.; Kutpruek, S.; Govitrapong, P. The role of melatonin in amyloid beta-induced inflammation mediated by inflammasome signaling in neuronal cell lines. Sci. Rep. 2023, 13, 17841. [Google Scholar] [CrossRef]
- Tanase, D.M.; Valasciuc, E.; Gosav, E.M.; Ouatu, A.; Buliga-Finis, O.N.; Floria, M.; Maranduca, M.A.; Serban, I.L. Portrayal of NLRP3 Inflammasome in Atherosclerosis: Current Knowledge and Therapeutic Targets. Int. J. Mol. Sci. 2023, 24, 8162. [Google Scholar] [CrossRef]
- Xie, S.; Fan, W.; He, H.; Huang, F. Role of Melatonin in the Regulation of Pain. J. Pain Res. 2020, 13, 331–343. [Google Scholar] [CrossRef]
- Wang, J.; Gu, J.; Ma, F.; Wei, Y.; Wang, P.; Yang, S.; Yan, X.; Xiao, Y.; Xing, K.; Lou, A.; et al. Melatonin Induces Analgesic Effects through MT2 Receptor-Mediated Neuroimmune Modulation in the Mice Anterior Cingulate Cortex. Research 2024, 7, 0493. [Google Scholar] [CrossRef]
- Oh, S.N.; Myung, S.K.; Jho, H.J. Analgesic Efficacy of Melatonin: A Meta-Analysis of Randomized, Double-Blind, Placebo-Controlled Trials. J. Clin. Med. 2020, 9, 1553. [Google Scholar] [CrossRef]
- Gilron, I.; DeBow, C.; Elkerdawy, H.; Khan, J.S.; Salomons, T.V.; Duggan, S.; Tu, D.; Holden, R.R.; Milev, R.; Buckley, D.N.; et al. PRECISE trial (Pain Relief Combination Intervention Strategies): Protocol for the clinical trial of a pregabalin-melatonin combination for fibromyalgia. BMJ Open 2024, 14, e087180. [Google Scholar] [CrossRef]
- Andersen, L.P.H.; Gögenur, I.; Fenger, A.Q.; Petersen, M.C.; Rosenberg, J.; Werner, M.U. Analgesic and antihyperalgesic effects of melatonin in a human inflammatory pain model: A randomized, double-blind, placebo-controlled, three-arm crossover study. Pain 2015, 156, 2286–2294. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; De Almeida Chuffa, L.G.; Simão, V.A.; Martín Giménez, V.M.; De Las Heras, N.; Spandidos, D.A.; Manucha, W. Melatonin and vitamin D as potential synergistic adjuvants for cancer therapy (Review). Int. J. Oncol. 2024, 65, 114. [Google Scholar] [CrossRef] [PubMed]
- Mehrzadi, S.; Pourhanifeh, M.H.; Mirzaei, A.; Moradian, F.; Hosseinzadeh, A. An updated review of mechanistic potentials of melatonin against cancer: Pivotal roles in angiogenesis, apoptosis, autophagy, endoplasmic reticulum stress and oxidative stress. Cancer Cell Int. 2021, 21, 188. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, S.; Zhou, Y.; Meng, X.; Zhang, J.J.; Xu, D.P.; Li, H.B. Melatonin for the prevention and treatment of cancer. Oncotarget 2017, 8, 39896–39921. [Google Scholar] [CrossRef]
- Sung, E.S.; Kim, J.Y.; Ahn, Y.T.; Lee, I.W.; Choi, S.W.; Jang, H.B.; Lee, J.C.; An, W.G. Melatonin Exerts Anticancer Effects in Human Tongue Squamous Cell Carcinoma Cells by Promoting Autophagy. Anticancer Res. 2020, 40, 6295–6303. [Google Scholar] [CrossRef]
- Bonmati-Carrion, M.A.; Tomas-Loba, A. Melatonin and Cancer: A Polyhedral Network Where the Source Matters. Antioxidants 2021, 10, 210. [Google Scholar] [CrossRef]
- Sohn, E.H.; Kim, S.N.; Lee, S.R. Melatonin’s Impact on Wound Healing. Antioxidants 2024, 13, 1197. [Google Scholar] [CrossRef]
- Drobnik, M.; Tomaszewska, A.; Ryżko, J.; Kędzia, A.; Gałdyszyńska, M.; Piera, L.; Rydel, J.; Szymański, J.; Drobnik, J. Melatonin increases collagen content accumulation and Fibroblast Growth Factor-2 secretion in cultured human cardiac fibroblasts. Pharmacol. Rep. 2023, 75, 560–569. [Google Scholar] [CrossRef]
- Greco, G.; Di Lorenzo, R.; Ricci, L.; Di Serio, T.; Vardaro, E.; Laneri, S. Clinical Studies Using Topical Melatonin. Int. J. Mol. Sci. 2024, 25, 5167. [Google Scholar] [CrossRef]
- Taheri, M.; Seirafianpour, F.; Fallahian, A.; Hosseinzadeh, A.; Reiter, R.J.; Mehrzadi, S. Exploring melatonin’s signalling pathways in the protection against age-related skin deterioration. Pharmacol. Rep. 2025, 77, 375–391. [Google Scholar] [CrossRef]
- Nakade, O.; Koyama, H.; Ariji, H.; Yajima, A.; Kaku, T. Melatonin stimulates proliferation and type I collagen synthesis in human bone cells in vitro. J. Pineal Res. 1999, 27, 106–110. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Yu, M.; Xie, D.; Wang, L.; Ye, C.; Zhu, Q.; Liu, F.; Yang, L. Melatonin-stimulated MSC-derived exosomes improve diabetic wound healing through regulating macrophage M1 and M2 polarization by targeting the PTEN/AKT pathway. Stem Cell Res. Ther. 2020, 11, 259. [Google Scholar] [CrossRef] [PubMed]
- Soriano, J.L.; Calpena, A.C.; Rincón, M.; Pérez, N.; Halbaut, L.; Rodríguez-Lagunas, M.J.; Clares, B. Melatonin nanogel promotes skin healing response in burn wounds of rats. Nanomedicine 2020, 15, 2133–2147. [Google Scholar] [CrossRef] [PubMed]
- Adamiak, K.; Gaida, V.A.; Schäfer, J.; Bosse, L.; Diemer, C.; Reiter, R.J.; Slominski, A.T.; Steinbrink, K.; Sionkowska, A.; Kleszczyński, K. Melatonin/Sericin Wound Healing Patches: Implications for Melanoma Therapy. Int. J. Mol. Sci. 2024, 25, 4858. [Google Scholar] [CrossRef]
- Mokhtari, T.; Yue, L.P.; Hu, L. Exogenous melatonin alleviates neuropathic pain-induced affective disorders by suppressing NF-κB/ NLRP3 pathway and apoptosis. Sci. Rep. 2023, 13, 2111. [Google Scholar] [CrossRef]
- Zeng, Y.; Fang, Q.; Chen, J.; Wang, Y.; Liu, X.; Zhang, X.; Shi, Y.; Zhan, H.; Zhong, X.; Yao, M.; et al. Melatonin Improves Mitochondrial Dysfunction and Attenuates Neuropathic Pain by Regulating SIRT1 in Dorsal Root Ganglions. Neuroscience 2023, 534, 29–40. [Google Scholar] [CrossRef]
- Hsieh, M.C.; Lai, C.Y.; Lin, L.T.; Chou, D.; Yeh, C.M.; Cheng, J.K.; Wang, H.H.; Lin, K.H.; Lin, T.B.; Peng, H.Y. Melatonin Relieves Paclitaxel-Induced Neuropathic Pain by Regulating pNEK2-Dependent Epigenetic Pathways in DRG Neurons. ACS Chem. Neurosci. 2023, 14, 4227–4239. [Google Scholar] [CrossRef]
- Kuthati, Y.; Wong, C.S. The Melatonin Type 2 Receptor Agonist IIK7 Attenuates and Reverses Morphine Tolerance in Neuropathic Pain Rats Through the Suppression of Neuroinflammation in the Spinal Cord. Pharmaceuticals 2024, 17, 1638. [Google Scholar] [CrossRef]
- Chen, I.J.; Yang, C.P.; Lin, S.H.; Lai, C.M.; Wong, C.S. The Circadian Hormone Melatonin Inhibits Morphine-Induced Tolerance and Inflammation via the Activation of Antioxidative Enzymes. Antioxidants 2020, 9, 780. [Google Scholar] [CrossRef]
- Shin, S.M.; Baek, E.J.; Kim, K.H.; Kim, K.J.; Park, E.J. Polydeoxyribonucleotide exerts opposing effects on ERK activity in human skin keratinocytes and fibroblasts. Mol. Med. Rep. 2023, 28, 148. [Google Scholar] [CrossRef]
- Nguyen, T.H.; Wang, S.L.; Nguyen, V.B. Recent advances on polydeoxyribonucleotide extraction and its novel application in cosmeceuticals. Int. J. Biol. Macromol. 2024, 282, 137051. [Google Scholar] [CrossRef] [PubMed]
- Chae, D.; Oh, S.W.; Choi, Y.S.; Kang, D.J.; Park, C.W.; Lee, J.; Seo, W.S. First Report on Microbial-Derived Polydeoxyribonucleotide: A Sustainable and Enhanced Alternative to Salmon-Based Polydeoxyribonucleotide. Curr. Issues Mol. Biol. 2025, 47, 41. [Google Scholar] [CrossRef] [PubMed]
- Baek, A.; Baek, D.; Kim, S.H.; Kim, J.; Notario, G.R.; Lee, D.W.; Kim, H.J.; Cho, S.R. Polydeoxyribonucleotide ameliorates IL-1β-induced impairment of chondrogenic differentiation in human bone marrow-derived mesenchymal stem cells. Sci. Rep. 2024, 14, 26076. [Google Scholar] [CrossRef]
- Kim, J.Y.; Pak, C.S.; Park, J.H.; Jeong, J.H.; Heo, C.Y. Effects of polydeoxyribonucleotide in the treatment of pressure ulcers. J. Korean Med. Sci. 2014, 29 (Suppl. S3), S222–S227. [Google Scholar] [CrossRef]
- Kwon, T.R.; Han, S.W.; Kim, J.H.; Lee, B.C.; Kim, J.M.; Hong, J.Y.; Kim, B.J. Polydeoxyribonucleotides Improve Diabetic Wound Healing in Mouse Animal Model for Experimental Validation. Ann. Dermatol. 2019, 31, 403–413. [Google Scholar] [CrossRef]
- Kim, C.H.; Hong, J.Y.; Shin, S.; Shin, S.Y.; Chung, J.H.; Thoma, D.S.; Lim, H.C. Preclinical Investigation on the Effect of Collagen Matrix With Polydeoxyribonucleotide at Buccally Positioned Implants. Clin. Implant Dent. Relat. Res. 2025, 27, e13411. [Google Scholar] [CrossRef]
- Kim, T.Y.; Kim, Y.T.; Hwang, J.T. Clinical Updates in Polydeoxyribonucleotide Injection. J. Korean Orthop. Assoc. 2024, 59, 386–394. [Google Scholar] [CrossRef]
- Ko, I.G.; Jin, J.J.; Hwang, L.; Kim, S.H.; Kim, C.J.; Won, K.Y.; Na, Y.G.; Kim, K.H.; Kim, S.J. Adenosine A2A Receptor Agonist Polydeoxyribonucleotide Alleviates Interstitial Cystitis-Induced Voiding Dysfunction by Suppressing Inflammation and Apoptosis in Rats. J. Inflamm. Res. 2021, 14, 367–378. [Google Scholar] [CrossRef]
- Ko, I.G.; Jin, J.J.; Hwang, L.; Kim, S.H.; Kim, C.J.; Jeon, J.W.; Chung, J.Y.; Han, J.H. Adenosine A2A receptor agonist polydeoxyribonucleotide ameliorates short-term memory impairment by suppressing cerebral ischemia-induced inflammation via MAPK pathway. PLoS ONE 2021, 16, e0248689. [Google Scholar] [CrossRef]
- Irrera, N.; Arcoraci, V.; Mannino, F.; Vermiglio, G.; Pallio, G.; Minutoli, L.; Bagnato, G.; Anastasi, G.P.; Mazzon, E.; Bramanti, P.; et al. Activation of A2A Receptor by PDRN Reduces Neuronal Damage and Stimulates WNT/β-CATENIN Driven Neurogenesis in Spinal Cord Injury. Front. Pharmacol. 2018, 9, 506. [Google Scholar] [CrossRef]
- Squadrito, F.; Micali, A.; Rinaldi, M.; Irrera, N.; Marini, H.; Puzzolo, D.; Pisani, A.; Lorenzini, C.; Valenti, A.; Laurà, R.; et al. Polydeoxyribonucleotide, an Adenosine-A2A Receptor Agonist, Preserves Blood Testis Barrier from Cadmium-Induced Injury. Front. Pharmacol. 2017, 7, 537. [Google Scholar] [CrossRef] [PubMed]
- Minutoli, L.; Arena, S.; Bonvissuto, G.; Bitto, A.; Polito, F.; Irrera, N.; Arena, F.; Fragalà, E.; Romeo, C.; Nicotina, P.A.; et al. Activation of adenosine A2A receptors by polydeoxyribonucleotide increases vascular endothelial growth factor and protects against testicular damage induced by experimental varicocele in rats. Fertil. Steril. 2011, 95, 1510–1513. [Google Scholar] [CrossRef] [PubMed]
- Ko, I.G.; Jin, J.J.; Hwang, L.; Kim, S.H.; Kim, C.J.; Han, J.H.; Lee, S.; Kim, H.I.; Shin, H.P.; Jeon, J.W. Polydeoxyribonucleotide Exerts Protective Effect Against CCl4-Induced Acute Liver Injury Through Inactivation of NF-κB/MAPK Signaling Pathway in Mice. Int. J. Mol. Sci. 2020, 21, 7894. [Google Scholar] [CrossRef]
- Castellini, C.; Belletti, S.; Govoni, P.; Guizzardi, S. Anti-Inflammatory Property of PDRN—An in Vitro Study on Cultured Macrophages. Adv. Biosci. Biotechnol. 2017, 8, 13–26. [Google Scholar] [CrossRef]
- Bitto, A.; Polito, F.; Irrera, N.; D’Ascola, A.; Avenoso, A.; Nastasi, G.; Campo, G.M.; Micali, A.; Bagnato, G.; Minutoli, L.; et al. Polydeoxyribonucleotide reduces cytokine production and the severity of collagen-induced arthritis by stimulation of adenosine A (2A) receptor. Arthritis Rheum. 2011, 63, 3364–3371. [Google Scholar] [CrossRef]
- Marques, C.; Porcello, A.; Cerrano, M.; Hadjab, F.; Chemali, M.; Lourenço, K.; Hadjab, B.; Raffoul, W.; Applegate, L.A.; Laurent, A.E. From Polydeoxyribonucleotides (PDRNs) to Polynucleotides (PNs): Bridging the Gap Between Scientific Definitions, Molecular Insights, and Clinical Applications of Multifunctional Biomolecules. Biomolecules 2025, 15, 148. [Google Scholar] [CrossRef]
- Lee, D.; Lee, J.; Seol, Y.J.; Lee, Y.M.; Koo, K.T. Effect of polydeoxyribonucleotide on early bone formation in lateral bone augmentation with immediate implant placement: An experimental in vivo study. Sci. Rep. 2023, 13, 16853. [Google Scholar] [CrossRef]
- Khan, A.; Wang, G.; Zhou, F.; Gong, L.; Zhang, J.; Qi, L.; Cui, H. Polydeoxyribonucleotide: A promising skin anti-aging agent. Chin. J. Plast. Reconstr. Surg. 2022, 4, 187–193. [Google Scholar] [CrossRef]
- Baek, A.; Kim, Y.; Lee, J.W.; Lee, S.C.; Cho, S.R. Effect of Polydeoxyribonucleotide on Angiogenesis and Wound Healing in an In Vitro Model of Osteoarthritis. Cell Transplant. 2018, 27, 1623–1633. [Google Scholar] [CrossRef]
- Jo, S.; Baek, A.; Cho, Y.; Kim, S.H.; Baek, D.; Hwang, J.; Cho, S.R.; Kim, H.J. Therapeutic effects of polydeoxyribonucleotide in an in vitro neuronal model of ischemia/reperfusion injury. Sci. Rep. 2023, 13, 6004. [Google Scholar] [CrossRef]
- Shin, D.Y.; Park, J.U.; Choi, M.H.; Kim, S.; Kim, H.E.; Jeong, S.H. Polydeoxyribonucleotide-delivering therapeutic hydrogel for diabetic wound healing. Sci. Rep. 2020, 10, 16811. [Google Scholar] [CrossRef] [PubMed]
- Squadrito, F.; Bitto, A.; Irrera, N.; Pizzino, G.; Pallio, G.; Minutoli, L.; Altavilla, D. Pharmacological Activity and Clinical Use of PDRN. Front. Pharmacol. 2017, 8, 224. [Google Scholar] [CrossRef] [PubMed]
- Colangelo, M.T.; Galli, C.; Guizzardi, S. Polydeoxyribonucleotide regulation of inflammation. Adv. Wound Care 2020, 9, 576–589. [Google Scholar] [CrossRef]
- Seo, S.J.; Lee, S.S.; Hwang, J.T.; Han, S.H.; Lee, J.R.; Kim, S. Effect of polydeoxyribonucleotide and polynucleotide on rotator cuff healing and fatty infiltration in a diabetic rat model. Sci. Rep. 2024, 14, 20623. [Google Scholar] [CrossRef]
- Noh, T.K.; Chung, B.Y.; Kim, S.Y.; Lee, M.H.; Kim, M.J.; Youn, C.S.; Lee, M.W.; Chang, S.E. Novel Anti-Melanogenesis Properties of Polydeoxyribonucleotide, a Popular Wound Healing Booster. Int. J. Mol. Sci. 2016, 17, 1448. [Google Scholar] [CrossRef]
- Park, H.J.; Byun, K.A.; Oh, S.; Kim, H.M.; Chung, M.S.; Son, K.H.; Byun, K. The Combination of Niacinamide, Vitamin C, and PDRN Mitigates Melanogenesis by Modulating Nicotinamide Nucleotide Transhydrogenase. Molecules 2022, 27, 4923. [Google Scholar] [CrossRef]
- Kim, B.R.; Kwon, S.H.; Kim, J.W.; Jeong, W.J.; Cha, W.; Jung, Y.H.; Na, J.I.; Huh, C.H.; Shin, J.W. Early Postoperative Injections of Polydeoxyribonucleotide Prevent Hypertrophic Scarring After Thyroidectomy: A Randomized Controlled Trial. Adv. Wound Care 2023, 12, 361–370. [Google Scholar] [CrossRef]
- Stazi, M.; Negro, S.; Megighian, A.; D’Este, G.; Solimena, M.; Jockers, R.; Lista, F.; Montecucco, C.; Rigoni, M. Melatonin promotes regeneration of injured motor axons via MT1 receptors. J. Pineal Res. 2021, 70, e12695. [Google Scholar] [CrossRef]
- Chang, H.M.; Liu, C.H.; Hsu, W.M.; Chen, L.Y.; Wang, H.P.; Wu, T.H.; Chen, K.Y.; Ho, W.H.; Liao, W.C. Proliferative effects of melatonin on Schwann cells: Implication for nerve regeneration following peripheral nerve injury. J. Pineal Res. 2014, 56, 322–332. [Google Scholar] [CrossRef]
- Yawoot, N.; Sengking, J.; Wicha, P.; Govitrapong, P.; Tocharus, C.; Tocharus, J. Melatonin attenuates reactive astrogliosis and glial scar formation following cerebral ischemia and reperfusion injury mediated by GSK-3β and RIP1K. J. Cell. Physiol. 2022, 237, 1818–1832. [Google Scholar] [CrossRef]
- Madhu, L.N.; Kodali, M.; Attaluri, S.; Shuai, B.; Melissari, L.; Rao, X.; Shetty, A.K. Melatonin improves brain function in a model of chronic Gulf War Illness with modulation of oxidative stress, NLRP3 inflammasomes, and BDNF-ERK-CREB pathway in the hippocampus. Redox Biol. 2021, 43, 101973. [Google Scholar] [CrossRef] [PubMed]
- Acuña-Castroviejo, D.; Escames, G.; Macías, M.; Muñóz Hoyos, A.; Molina Carballo, A.; Arauzo, M.; Montes, R. Cell protective role of melatonin in the brain. J. Pineal Res. 1995, 19, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Samec, M.; Liskova, A.; Koklesova, L.; Zhai, K.; Varghese, E.; Samuel, S.M.; Šudomová, M.; Lucansky, V.; Kassayova, M.; Pec, M.; et al. Metabolic Anti-Cancer Effects of Melatonin: Clinically Relevant Prospects. Cancers 2021, 13, 3018. [Google Scholar] [CrossRef]
- Chuffa, L.G.A.; Carvalho, R.F.; Justulin, L.A.; Cury, S.S.; Seiva, F.R.F.; Jardim-Perassi, B.V.; Zuccari, D.A.P.C.; Reiter, R.J. A meta-analysis of microRNA networks regulated by melatonin in cancer: Portrait of potential candidates for breast cancer treatment. J. Pineal Res. 2020, 69, e12693. [Google Scholar] [CrossRef]
- Bilska, B.; Schedel, F.; Piotrowska, A.; Stefan, J.; Zmijewski, M.; Pyza, E.; Reiter, R.J.; Steinbrink, K.; Slominski, A.T.; Tulic, M.K.; et al. Mitochondrial function is controlled by melatonin and its metabolites in vitro in human melanoma cells. J. Pineal Res. 2021, 70, e12728. [Google Scholar] [CrossRef]
- Joo, S.S.; Yoo, Y.M. Melatonin induces apoptotic death in LNCaP cells via p38 and JNK pathways: Therapeutic implications for prostate cancer. J. Pineal Res. 2009, 47, 8–14. [Google Scholar] [CrossRef]
- Kim, T.H.; Heo, S.Y.; Han, J.S.; Jung, W.K. Anti-inflammatory effect of polydeoxyribonucleotides (PDRN) extracted from red alga (Porphyra sp.) (Ps-PDRN) in RAW 264.7 macrophages stimulated with Escherichia coli lipopolysaccharides: A comparative study with commercial PDRN. Cell Biochem. Funct. 2023, 41, 889–897. [Google Scholar] [CrossRef]
- Baek, S.W.; Kim, D.S.; Song, D.H.; Lee, S.; Lee, J.K.; Park, S.Y.; Kim, J.H.; Kim, T.H.; Park, C.G.; Han, D.K. PLLA Composites Combined with Delivery System of Bioactive Agents for Anti-Inflammation and Re-Endothelialization. Pharmaceutics 2022, 14, 2661. [Google Scholar] [CrossRef]
- Kim, T.H.; Heo, S.Y.; Oh, G.W.; Heo, S.J.; Jung, W.K. Applications of Marine Organism-Derived Polydeoxyribonucleotide: Its Potential in Biomedical Engineering. Mar. Drugs 2021, 19, 296. [Google Scholar] [CrossRef]
- Roh, E.J.; Kim, D.S.; Kim, J.H.; Lim, C.S.; Choi, H.; Kwon, S.Y.; Park, S.Y.; Kim, J.Y.; Kim, H.M.; Hwang, D.Y.; et al. Multimodal therapy strategy based on a bioactive hydrogel for repair of spinal cord injury. Biomaterials 2023, 299, 122160. [Google Scholar] [CrossRef]
- Hwang, L.; Ko, I.G.; Jin, J.J.; Kim, S.H.; Kim, C.J.; Hwang, J.J.; Choi, C.W.; Chang, B.S. Attenuation effect of polydeoxyribonucleotide on inflammatory cytokines and apoptotic factors induced by particulate matter (PM10) damage in human bronchial cells. J. Biochem. Mol. Toxicol. 2021, 35, e22635. [Google Scholar] [CrossRef]
- Galeano, M.; Pallio, G.; Irrera, N.; Mannino, F.; Bitto, A.; Altavilla, D.; Vaccaro, M.; Squadrito, G.; Arcoraci, V.; Colonna, M.R.; et al. Polydeoxyribonucleotide: A Promising Biological Platform to Accelerate Impaired Skin Wound Healing. Pharmaceuticals 2021, 14, 1103. [Google Scholar] [CrossRef] [PubMed]
- Ko, K.W.; Park, S.Y.; Lee, E.H.; Yoo, Y.I.; Kim, D.S.; Kim, J.Y.; Kwon, T.G.; Han, D.K. Integrated Bioactive Scaffold with Polydeoxyribonucleotide and Stem-Cell-Derived Extracellular Vesicles for Kidney Regeneration. ACS Nano 2021, 15, 7575–7585. [Google Scholar] [CrossRef]
- Colangelo, M.T.; Galli, C.; Guizzardi, S. The effects of polydeoxyribonucleotide on wound healing and tissue regeneration: A systematic review of the literature. Regen. Med. 2020, 15, 1801–1821. [Google Scholar] [CrossRef]
- Edirisinghe, S.L.; Nikapitiya, C.; Dananjaya, S.H.S.; Park, J.; Kim, D.; Choi, D.; De Zoysa, M. Effect of Polydeoxyribonucleotide (PDRN) Treatment on Corneal Wound Healing in Zebrafish (Danio rerio). Int. J. Mol. Sci. 2022, 23, 13525. [Google Scholar] [CrossRef]
- Veronesi, F.; Dallari, D.; Sabbioni, G.; Carubbi, C.; Martini, L.; Fini, M. Polydeoxyribonucleotides (PDRNs) From Skin to Musculoskeletal Tissue Regeneration via Adenosine A2A Receptor Involvement. J. Cell. Physiol. 2017, 232, 2299–2307. [Google Scholar] [CrossRef]
- Dananjaya, S.H.S.; Madushani, K.G.P.; Dilrukshi, J.; De Silva, N.D.; Sandamalika, W.M.G.; Kim, D.; Choi, D.; De Zoysa, M.; Attanayake, A.P. Development and characterization of polydeoxyribonucleotide (PDRN) loaded chitosan polyplex: In vitro and in vivo evaluation of wound healing activity. Int. J. Biol. Macromol. 2023, 253 Pt 3, 126729. [Google Scholar] [CrossRef]
- Picciolo, G.; Mannino, F.; Irrera, N.; Altavilla, D.; Minutoli, L.; Vaccaro, M.; Arcoraci, V.; Squadrito, V.; Picciolo, G.; Squadrito, F.; et al. PDRN, a natural bioactive compound, blunts inflammation and positively reprograms healing genes in an “in vitro” model of oral mucositis. Biomed. Pharmacother. 2021, 138, 111538. [Google Scholar] [CrossRef]
- Pallio, G.; Bitto, A.; Pizzino, G.; Galfo, F.; Irrera, N.; Squadrito, F.; Squadrito, G.; Pallio, S.; Anastasi, G.P.; Cutroneo, G.; et al. Adenosine Receptor Stimulation by Polydeoxyribonucleotide Improves Tissue Repair and Symptomology in Experimental Colitis. Front. Pharmacol. 2016, 7, 273. [Google Scholar] [CrossRef]
- Li, X.; Qu, S.; Ouyang, Q.; Qin, F.; Guo, J.; Qin, M.; Zhang, J. A multifunctional composite nanoparticle with antibacterial activities, anti-inflammatory, and angiogenesis for diabetic wound healing. Int. J. Biol. Macromol. 2024, 260 Pt 2, 129531. [Google Scholar] [CrossRef]
- Galeano, M.; Bitto, A.; Altavilla, D.; Minutoli, L.; Polito, F.; Calò, M.; Lo Cascio, P.; Stagno d’Alcontres, F.; Squadrito, F. Polydeoxyribonucleotide stimulates angiogenesis and wound healing in the genetically diabetic mouse. Wound Repair. Regen. 2008, 16, 208–217. [Google Scholar] [CrossRef] [PubMed]
- Dananjaya, S.H.S.; Bandara, N.; Molagoda, I.M.N.; Sandamalika, W.M.G.; Kim, D.; Ganepola, N.; Attanayake, A.P.; Choi, D. Multifunctional alginate/polydeoxyribonucleotide hydrogels for promoting diabetic wound healing. Int. J. Biol. Macromol. 2024, 257 Pt 1, 128367. [Google Scholar] [CrossRef] [PubMed]
- Tarocco, A.; Caroccia, N.; Morciano, G.; Wieckowski, M.R.; Ancora, G.; Garani, G.; Pinton, P. Melatonin as a master regulator of cell death and inflammation: Molecular mechanisms and clinical implications for newborn care. Cell Death Dis. 2019, 10, 317. [Google Scholar] [CrossRef]
- Esposito, E.; Cuzzocrea, S. Antiinflammatory activity of melatonin in central nervous system. Curr. Neuropharmacol. 2010, 8, 228–242. [Google Scholar] [CrossRef]
- Wang, J.; Gao, S.; Lenahan, C.; Gu, Y.; Wang, X.; Fang, Y.; Xu, W.; Wu, H.; Pan, Y.; Shao, A.; et al. Melatonin as an Antioxidant Agent in Stroke: An Updated Review. Aging Dis. 2022, 13, 1823–1844. [Google Scholar] [CrossRef]
- Lempesis, I.G.; Georgakopoulou, V.E.; Reiter, R.J.; Spandidos, D.A. A mid-pandemic night’s dream: Melatonin, from harbinger of anti-inflammation to mitochondrial savior in acute and long COVID-19 (Review). Int. J. Mol. Med. 2024, 53, 28. [Google Scholar] [CrossRef]
- Gilron, I.; Tu, D.; Holden, R.R.; Moulin, D.E.; Duggan, S.; Milev, R. Melatonin for Neuropathic Pain: Protocol for a Double-blind, Randomized Controlled Trial. JMIR Res. Protoc. 2022, 11, e40025. [Google Scholar] [CrossRef]
- Yu, L.; Chen, Z.; Zhou, X.; Teng, F.; Bai, Q.R.; Li, L.; Li, Y.; Liu, Y.; Zeng, Q.; Wang, Y.; et al. KARS Mutations Impair Brain Myelination by Inducing Oligodendrocyte Deficiency: One Potential Mechanism and Improvement by Melatonin. J. Pineal Res. 2024, 76, e12998. [Google Scholar] [CrossRef]
- Galano, A.; Tan, D.X.; Reiter, R.J. On the free radical scavenging activities of melatonin’s metabolites, AFMK and AMK. J. Pineal Res. 2013, 54, 245–257. [Google Scholar] [CrossRef]
- Yong, W.; Ma, H.; Na, M.; Gao, T.; Zhang, Y.; Hao, L.; Yu, H.; Yang, H.; Deng, X. Roles of melatonin in the field of reproductive medicine. Biomed. Pharmacother. 2021, 144, 112001. [Google Scholar] [CrossRef]
- Poeggeler, B.; Reiter, R.J.; Tan, D.X.; Chen, L.D.; Manchester, L.C. Melatonin, hydroxyl radical-mediated oxidative damage, and aging: A hypothesis. J. Pineal Res. 1993, 14, 151–168. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Tang, Q.; Zeng, Y. Melatonin: Potential avenue for treating iron overload disorders. Ageing Res. Rev. 2022, 81, 101717. [Google Scholar] [CrossRef] [PubMed]
- Sevilla, A.; Chéret, J.; Slominski, R.M.; Slominski, A.T.; Paus, R. Revisiting the role of melatonin in human melanocyte physiology: A skin context perspective. J. Pineal Res. 2022, 72, e12790. [Google Scholar] [CrossRef]
- Reiter, R.J.; Sharma, R.; Cucielo, M.S.; Tan, D.X.; Rosales-Corral, S.; Gancitano, G.; de Almeida Chuffa, L.G. Brain washing and neural health: Role of age, sleep, and the cerebrospinal fluid melatonin rhythm. Cell. Mol. Life Sci. 2023, 80, 88. [Google Scholar] [CrossRef]
- Reiter, R.J.; Sharma, R.; Rosales-Corral, S.; de Mange, J.; Phillips, W.T.; Tan, D.X.; Bitar, R.D. Melatonin in ventricular and subarachnoid cerebrospinal fluid: Its function in the neural glymphatic network and biological significance for neurocognitive health. Biochem. Biophys. Res. Commun. 2022, 605, 70–81. [Google Scholar] [CrossRef]
- Korf, H.W.; Bruce, J.A.; Vistica, B.; Rollag, M.; Stein, B.M.; Klein, D.C. Immunoreactive S-antigen in cerebrospinal fluid: A marker of pineal parenchymal tumors? J. Neurosurg. 1989, 70, 682–687. [Google Scholar] [CrossRef]
- Squadrito, F.; Bitto, A.; Altavilla, D.; Arcoraci, V.; De Caridi, G.; De Feo, M.E.; Corrao, S.; Pallio, G.; Sterrantino, C.; Minutoli, L.; et al. The effect of PDRN, an adenosine receptor A2A agonist, on the healing of chronic diabetic foot ulcers: Results of a clinical trial. J. Clin. Endocrinol. Metab. 2014, 99, E746–E753. [Google Scholar] [CrossRef]
- Irrera, N.; Bitto, A.; Vaccaro, M.; Mannino, F.; Squadrito, V.; Pallio, G.; Arcoraci, V.; Minutoli, L.; Ieni, A.; Lentini, M.; et al. PDRN, a Bioactive Natural Compound, Ameliorates Imiquimod-Induced Psoriasis through NF-κB Pathway Inhibition and Wnt/β-Catenin Signaling Modulation. Int. J. Mol. Sci. 2020, 21, 1215. [Google Scholar] [CrossRef]
- Kim, S.E.; Ko, I.G.; Jin, J.J.; Hwang, L.; Kim, C.J.; Kim, S.H.; Han, J.H.; Jeon, J.W. Polydeoxyribonucleotide Exerts Therapeutic Effect by Increasing VEGF and Inhibiting Inflammatory Cytokines in Ischemic Colitis Rats. BioMed Res. Int. 2020, 2020, 2169083. [Google Scholar] [CrossRef]
- Spinelli, S.; Remigante, A.; Liuni, R.; Mantegna, G.; Legname, G.; Marino, A.; Morabito, R.; Dossena, S. Oxidative stress-related cellular aging causes dysfunction of the Kv3.1/KCNC1 channel reverted by melatonin. Aging Cell 2024, 23, e14185. [Google Scholar] [CrossRef]
- Scribante, A. Melatonin regulation and the function of the periodontal ligament: Future perspective and challenges. World J. Stem Cells 2025, 17, 101347. [Google Scholar] [CrossRef] [PubMed]
- Okatani, Y.; Wakatsuki, A.; Reiter, R.J.; Miyahara, Y. Acutely administered melatonin restores hepatic mitochondrial physiology in old mice. Int. J. Biochem. Cell Biol. 2003, 35, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Hao, B.; Zhang, M.; Reiter, R.J.; Lin, S.; Zheng, T.; Chen, X.; Ren, Y.; Yue, L.; Abay, B.; et al. Melatonin enhances radiofrequency-induced NK antitumor immunity, causing cancer metabolism reprogramming and inhibition of multiple pulmonary tumor development. Signal Transduct. Target. Ther. 2021, 6, 330. [Google Scholar] [CrossRef] [PubMed]
- Mafi, A.; Rezaee, M.; Hedayati, N.; Hogan, S.D.; Reiter, R.J.; Aarabi, M.H.; Asemi, Z. Melatonin and 5-fluorouracil combination chemotherapy: Opportunities and efficacy in cancer therapy. Cell Commun. Signal. 2023, 21, 33. [Google Scholar] [CrossRef]
- Liu, D.; Shi, K.; Fu, M.; Chen, F. Melatonin indirectly decreases gastric cancer cell proliferation and invasion via effects on cancer-associated fibroblasts. Life Sci. 2021, 277, 119497. [Google Scholar] [CrossRef]
- Shin, Y.Y.; Seo, Y.; Oh, S.J.; Ahn, J.S.; Song, M.H.; Kang, M.J.; Oh, J.M.; Lee, D.; Kim, Y.H.; Sung, E.S.; et al. Melatonin and verteporfin synergistically suppress the growth and stemness of head and neck squamous cell carcinoma through the regulation of mitochondrial dynamics. J. Pineal Res. 2022, 72, e12779. [Google Scholar] [CrossRef]
- Pugazhenthi, K.; Kapoor, M.; Clarkson, A.N.; Hall, I.; Appleton, I. Melatonin accelerates the process of wound repair in full-thickness incisional wounds. J. Pineal Res. 2008, 44, 387–396. [Google Scholar] [CrossRef]
- Lin, J.J.; Lin, Y.; Zhao, T.Z.; Zhang, C.K.; Zhang, T.; Chen, X.L.; Ding, J.Q.; Chang, T.; Zhang, Z.; Sun, C.; et al. Melatonin Suppresses Neuropathic Pain via MT2-Dependent and -Independent Pathways in Dorsal Root Ganglia Neurons of Mice. Theranostics 2017, 7, 2015–2032. [Google Scholar] [CrossRef]
- Galley, H.F.; McCormick, B.; Wilson, K.L.; Lowes, D.A.; Colvin, L.; Torsney, C. Melatonin limits paclitaxel-induced mitochondrial dysfunction in vitro and protects against paclitaxel-induced neuropathic pain in the rat. J. Pineal Res. 2017, 63, e12444. [Google Scholar] [CrossRef]
- Kuthati, Y.; Goutham Davuluri, V.N.; Yang, C.P.; Chang, H.C.; Chang, C.P.; Wong, C.S. Melatonin MT2 receptor agonist IIK-7 produces antinociception by modulation of ROS and suppression of spinal microglial activation in neuropathic pain rats. J. Pain Res. 2019, 12, 2473–2485. [Google Scholar] [CrossRef]
- Hsieh, M.C.; Ho, Y.C.; Lai, C.Y.; Chou, D.; Wang, H.H.; Chen, G.D.; Lin, T.B.; Peng, H.Y. Melatonin impedes Tet1-dependent mGluR5 promoter demethylation to relieve pain. J. Pineal Res. 2017, 63, e12436. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Jiang, H.; Liu, X.; Hu, S.; Li, H.; Feng, Y.; Ke, J.; Long, X. Melatonin Abates TMJOA Chronic Pain by MT2R in Trigeminal Ganglion Neurons. J. Dent. Res. 2022, 101, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Polito, F.; Bitto, A.; Galeano, M.; Irrera, N.; Marini, H.; Calò, M.; Squadrito, F.; Altavilla, D. Polydeoxyribonucleotide restores blood flow in an experimental model of ischemic skin flaps. J. Vasc. Surg. 2012, 55, 479–488. [Google Scholar] [CrossRef]
- Thellung, S.; Florio, T.; Maragliano, A.; Cattarini, G.; Schettini, G. Polydeoxyribonucleotides enhance the proliferation of human skin fibroblasts: Involvement of A2 purinergic receptor subtypes. Life Sci. 1999, 64, 1661–1674. [Google Scholar] [CrossRef]
- Kang, S.H.; Choi, M.S.; Kim, H.K.; Kim, W.S.; Bae, T.H.; Kim, M.K.; Chang, S.H. Polydeoxyribonucleotide improves tendon healing following achilles tendon injury in rats. J. Orthop. Res. 2018, 36, 1767–1776. [Google Scholar] [CrossRef]
- Jeong, W.; Yang, C.E.; Roh, T.S.; Kim, J.H.; Lee, J.H.; Lee, W.J. Scar Prevention and Enhanced Wound Healing Induced by Polydeoxyribonucleotide in a Rat Incisional Wound-Healing Model. Int. J. Mol. Sci. 2017, 18, 1698. [Google Scholar] [CrossRef]
- Moretti, L.; Bizzoca, D.; Geronimo, A.; Abbaticchio, A.M.; Moretti, F.L.; Carlet, A.; Fischetti, F.; Moretti, B. Targeting Adenosine Signalling in Knee Chondropathy: The Combined Action of Polydeoxyribonucleotide and Pulsed Electromagnetic Fields: A Current Concept Review. Int. J. Mol. Sci. 2023, 24, 10090. [Google Scholar] [CrossRef]
- Joo, Y.C.; Chung, J.Y.; Kwon, S.O.; Han, J.H. Adenosine A2A Receptor Agonist, Polydeoxyribonucleotide Treatment Improves Locomotor Function and Thermal Hyperalgesia Following Neuropathic Pain in Rats. Int. Neurourol. J. 2023, 27, 243–251. [Google Scholar] [CrossRef]
- Sun, Y.; Li, Y.; Ding, X.; Xu, P.; Jing, X.; Cong, H.; Hu, H.; Yu, B.; Xu, F.J. An NIR-responsive hydrogel loaded with polydeoxyribonucleotide nano-vectors for enhanced chronic wound healing. Biomaterials 2025, 314, 122789. [Google Scholar] [CrossRef]
- Hwang, K.H.; Kim, J.H.; Park, E.Y.; Cha, S.K. An effective range of polydeoxyribonucleotides is critical for wound healing quality. Mol. Med. Rep. 2018, 18, 5166–5172. [Google Scholar] [CrossRef]
- Lee, K.S.; Lee, S.; Wang, H.; Lee, G.; Kim, S.; Ryu, Y.H.; Chang, N.H.; Kang, Y.W. Analysis of Skin Regeneration and Barrier-Improvement Efficacy of Polydeoxyribonucleotide Isolated from Panax Ginseng (C.A. Mey.). Adventitious Root. Mol. 2023, 28, 7240. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Lee, H.J.; Jang, H.J.; Park, H.; Kim, G.J. Human Placenta MSC-Derived DNA Fragments Exert Therapeutic Effects in a Skin Wound Model via the A2A Receptor. Int. J. Mol. Sci. 2025, 26, 1769. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Choi, S.; Kim, S.Y.; Jang, S.J.; Lee, S.; Kim, H.; Jang, J.H.; Seo, H.H.; Lee, J.H.; Choi, S.S.; et al. Wound healing effect of polydeoxyribonucleotide derived from Hibiscus sabdariffa callus via Nrf2 signaling in human keratinocytes. Biochem. Biophys. Res. Commun. 2024, 728, 150335. [Google Scholar] [CrossRef]
- Yu, M.; Lee, J.Y. Polydeoxyribonucleotide improves wound healing of fractional laser resurfacing in rat model. J. Cosmet. Laser Ther. 2017, 19, 43–48. [Google Scholar] [CrossRef]
- Kim, H.M.; Byun, K.A.; Oh, S.; Yang, J.Y.; Park, H.J.; Chung, M.S.; Son, K.H.; Byun, K. A Mixture of Topical Forms of Polydeoxyribonucleotide, Vitamin C, and Niacinamide Attenuated Skin Pigmentation and Increased Skin Elasticity by Modulating Nuclear Factor Erythroid 2-like 2. Molecules 2022, 27, 1276. [Google Scholar] [CrossRef]
- Gwak, D.W.; Hwang, J.M.; Kim, A.R.; Park, D. Does polydeoxyribonucleotide has an effect on patients with tendon or ligament pain?: A PRISMA-compliant meta-analysis. Medicine 2021, 100, e25792. [Google Scholar] [CrossRef]
- Kim, Y.N.; Baek, J.; Kim, Y.H.; Hwang, J.; Ko, Y.R.; Lee, M.S.; Kim, Y.C.; Park, H.J. Full recovery of near complete tear of anterior cruciate ligament without surgery: A case report. Medicine 2020, 99, e19899. [Google Scholar] [CrossRef]
- Lee, Y.J.; Moon, Y.S.; Kwon, D.R.; Cho, S.C.; Kim, E.H. Polydeoxyribonucleotide and Shock Wave Therapy Sequence Efficacy in Regenerating Immobilized Rabbit Calf Muscles. Int. J. Mol. Sci. 2023, 24, 12820. [Google Scholar] [CrossRef]
- Hwang, J.T.; Lee, S.S.; Han, S.H.; Sherchan, B.; Panakkal, J.J. Polydeoxyribonucleotide and Polynucleotide Improve Tendon Healing and Decrease Fatty Degeneration in a Rat Cuff Repair Model. Tissue Eng. Regen. Med. 2021, 18, 1009–1020. [Google Scholar] [CrossRef]
- Rho, J.H.; Ko, I.G.; Jin, J.J.; Hwang, L.; Kim, S.H.; Chung, J.Y.; Hwang, T.J.; Han, J.H. Polydeoxyribonucleotide Ameliorates Inflammation and Apoptosis in Achilles Tendon-Injury Rats. Int. Neurourol. J. 2020, 24 (Suppl. S2), 79–87. [Google Scholar] [CrossRef]
- Park, S.; Son, Y.; Park, J.; Lee, S.; Kim, N.H.; Jang, S.N.; Kang, T.W.; Song, J.E.; Khang, G. Polydeoxynucleotide-Loaded Visible Light Photo-Crosslinked Gelatin Methacrylate Hydrogel: Approach to Accelerating Cartilage Regeneration. Gels 2025, 11, 42. [Google Scholar] [CrossRef] [PubMed]
- Avantaggiato, A.; Palmieri, A.; Carinci, F.; Trapella, G.; Sollazzo, V.; Lauritano, D. Effects of glucosamine and nucleotide association on fibroblast: Extracellular matrix gene expression. Int. J. Immunopathol. Pharmacol. 2014, 27, 689–693. [Google Scholar] [CrossRef] [PubMed]
- Ha, Y.; Kim, J.H.; Kim, J.; Kwon, H. Non-surgical treatment of diabetic foot ulcers on the dorsum of the foot with polydeoxyribonucleotide injection: Two case reports. World J. Clin. Cases 2024, 12, 4446–4451. [Google Scholar] [CrossRef] [PubMed]
- Bizzoca, D.; Brunetti, G.; Moretti, L.; Piazzolla, A.; Vicenti, G.; Moretti, F.L.; Solarino, G.; Moretti, B. Polydeoxyribonucleotide in the Treatment of Tendon Disorders, from Basic Science to Clinical Practice: A Systematic Review. Int. J. Mol. Sci. 2023, 24, 4582. [Google Scholar] [CrossRef]
- Lee, D.O.; Yoo, J.H.; Cho, H.I.; Cho, S.; Cho, H.R. Comparing effectiveness of polydeoxyribonucleotide injection and corticosteroid injection in plantar fasciitis treatment: A prospective randomized clinical study. Foot Ankle Surg. 2020, 26, 657–661. [Google Scholar] [CrossRef]
- Shim, B.J.; Seo, E.M.; Hwang, J.T.; Kim, D.Y.; Yang, J.S.; Seo, S.J.; Hong, M.S. Comparison of the effectiveness of extensor muscle strengthening exercise by itself, exercise with polydeoxyribonucleotide injection, and exercise with extracorporeal shockwave therapy in lateral epicondylitis: A randomized controlled trial. Clin. Shoulder Elb. 2021, 24, 231–238. [Google Scholar] [CrossRef]
- Lee, G.J.; Park, D. Usefulness of polydeoxyribonucleotide as an alternative to corticosteroids in patients with lateral epicondyitis: A case series. Medicine 2018, 97, e10809. [Google Scholar] [CrossRef]
- Chang, Y.S.; Chiang, B.L. Melatonin-Induced Nocturnal Vasodilatation Contributes to Skin Regeneration-Reply. JAMA Pediatr. 2016, 170, 622. [Google Scholar] [CrossRef]
- Mekjavic, I.B.; Macdonald, I.A. Melatonin-Induced Nocturnal Vasodilatation Contributes to Skin Regeneration. JAMA Pediatr. 2016, 170, 621–622. [Google Scholar] [CrossRef]
- Mustafa, F.E.A.; Abdel-Maksoud, F.M.; Hassan, A.H.S.; Mokhtar, D.M. Melatonin induces a stimulatory action on the scrotal skin components of Soay ram in the non-breeding season. Sci. Rep. 2020, 10, 10154. [Google Scholar] [CrossRef]
- Samra, T.; Gomez-Gomez, T.; Linowiecka, K.; Akhundlu, A.; Lopez de Mendoza, G.; Gompels, M.; Lee, W.W.; Gherardini, J.; Chéret, J.; Paus, R. Melatonin Exerts Prominent, Differential Epidermal and Dermal Anti-Aging Properties in Aged Human Eyelid Skin Ex Vivo. Int. J. Mol. Sci. 2023, 24, 15963. [Google Scholar] [CrossRef] [PubMed]
- Meyer, N.; Harvey, A.G.; Lockley, S.W.; Dijk, D.J. Circadian rhythms and disorders of the timing of sleep. Lancet 2022, 400, 1061–1078. [Google Scholar] [CrossRef] [PubMed]
- Mogavero, M.P.; DelRosso, L.M.; Fanfulla, F.; Bruni, O.; Ferri, R. Sleep disorders and cancer: State of the art and future perspectives. Sleep Med. Rev. 2021, 56, 101409. [Google Scholar] [CrossRef] [PubMed]
- Duffy, J.F.; Wang, W.; Ronda, J.M.; Czeisler, C.A. High dose melatonin increases sleep duration during nighttime and daytime sleep episodes in older adults. J. Pineal Res. 2022, 73, e12801. [Google Scholar] [CrossRef]
- Auld, F.; Maschauer, E.L.; Morrison, I.; Skene, D.J.; Riha, R.L. Evidence for the efficacy of melatonin in the treatment of primary adult sleep disorders. Sleep Med. Rev. 2017, 34, 10–22. [Google Scholar] [CrossRef]
- Nogueira, H.A.; de Castro, C.T.; da Silva, D.C.G.; Pereira, M. Melatonin for sleep disorders in people with autism: Systematic review and meta-analysis. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2023, 123, 110695. [Google Scholar] [CrossRef]
- Esposito, S.; Laino, D.; D’Alonzo, R.; Mencarelli, A.; Di Genova, L.; Fattorusso, A.; Argentiero, A.; Mencaroni, E. Pediatric sleep disturbances and treatment with melatonin. J. Transl. Med. 2019, 17, 77. [Google Scholar] [CrossRef]
- Kim, D.S.; Jeong, D.S.; Park, S.Y.; Jung, J.W.; Lee, J.E.; Lee, J.K.; Baek, S.W.; Lee, D.R.; Han, D.K. Ovarian Function Restoration with Biomimetic Scaffold Incorporating Angiogenic Molecules and Antioxidant in Chemotherapy-Induced Perimenopausal Model. Adv. Healthc. Mater. 2025, 14, e2403944. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jang, S.K.; Choi, J.; Lim, H.W.; Kim, H.-G.; Yoo, Y.-M. Comparative Analysis of Melatonin and Polydeoxyribonucleotide: Possible Benefits of Co-Treatment Effects and Potential Synergistic Applicability. Int. J. Mol. Sci. 2025, 26, 5703. https://doi.org/10.3390/ijms26125703
Jang SK, Choi J, Lim HW, Kim H-G, Yoo Y-M. Comparative Analysis of Melatonin and Polydeoxyribonucleotide: Possible Benefits of Co-Treatment Effects and Potential Synergistic Applicability. International Journal of Molecular Sciences. 2025; 26(12):5703. https://doi.org/10.3390/ijms26125703
Chicago/Turabian StyleJang, Su Kil, Jaeseok Choi, Hye Won Lim, Hong-Gyum Kim, and Yeong-Min Yoo. 2025. "Comparative Analysis of Melatonin and Polydeoxyribonucleotide: Possible Benefits of Co-Treatment Effects and Potential Synergistic Applicability" International Journal of Molecular Sciences 26, no. 12: 5703. https://doi.org/10.3390/ijms26125703
APA StyleJang, S. K., Choi, J., Lim, H. W., Kim, H.-G., & Yoo, Y.-M. (2025). Comparative Analysis of Melatonin and Polydeoxyribonucleotide: Possible Benefits of Co-Treatment Effects and Potential Synergistic Applicability. International Journal of Molecular Sciences, 26(12), 5703. https://doi.org/10.3390/ijms26125703