
Targeted DNA Methylation Using Modified DNA Probes: A Potential Therapeutic Tool for Depression and Stress-Related Disorders



Abstract
1. Introduction
2. Results
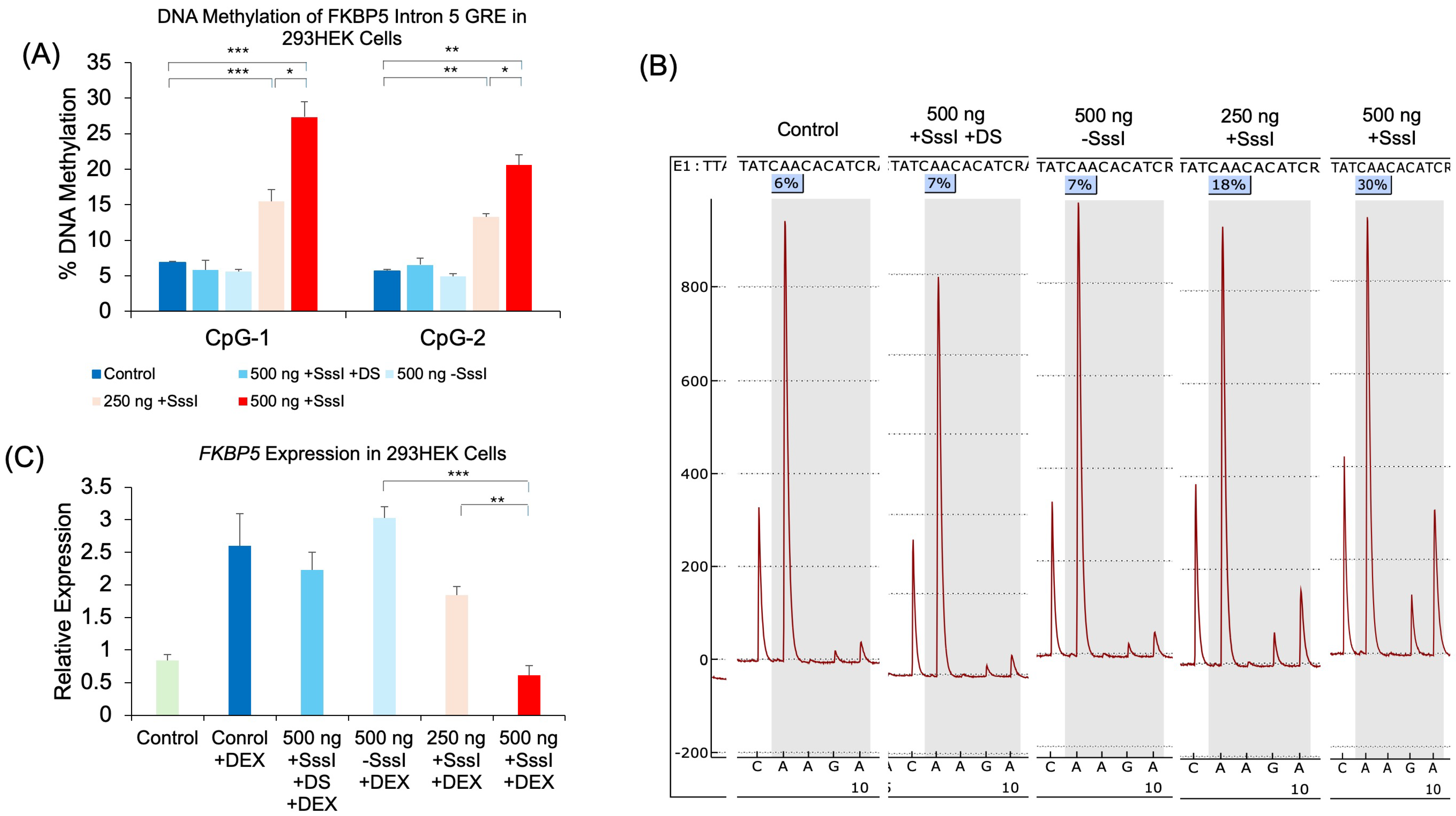
2.1. The Use of Methylated, Single-Stranded Probe to Induce Target-Specific DNAm
2.2. The Effect of Probe-Induced DNAm on Gene Expression
2.3. Persistence of Probe-Induced DNAm Patterns and Accumulation of DNAm Following Multiple Probe Transfections
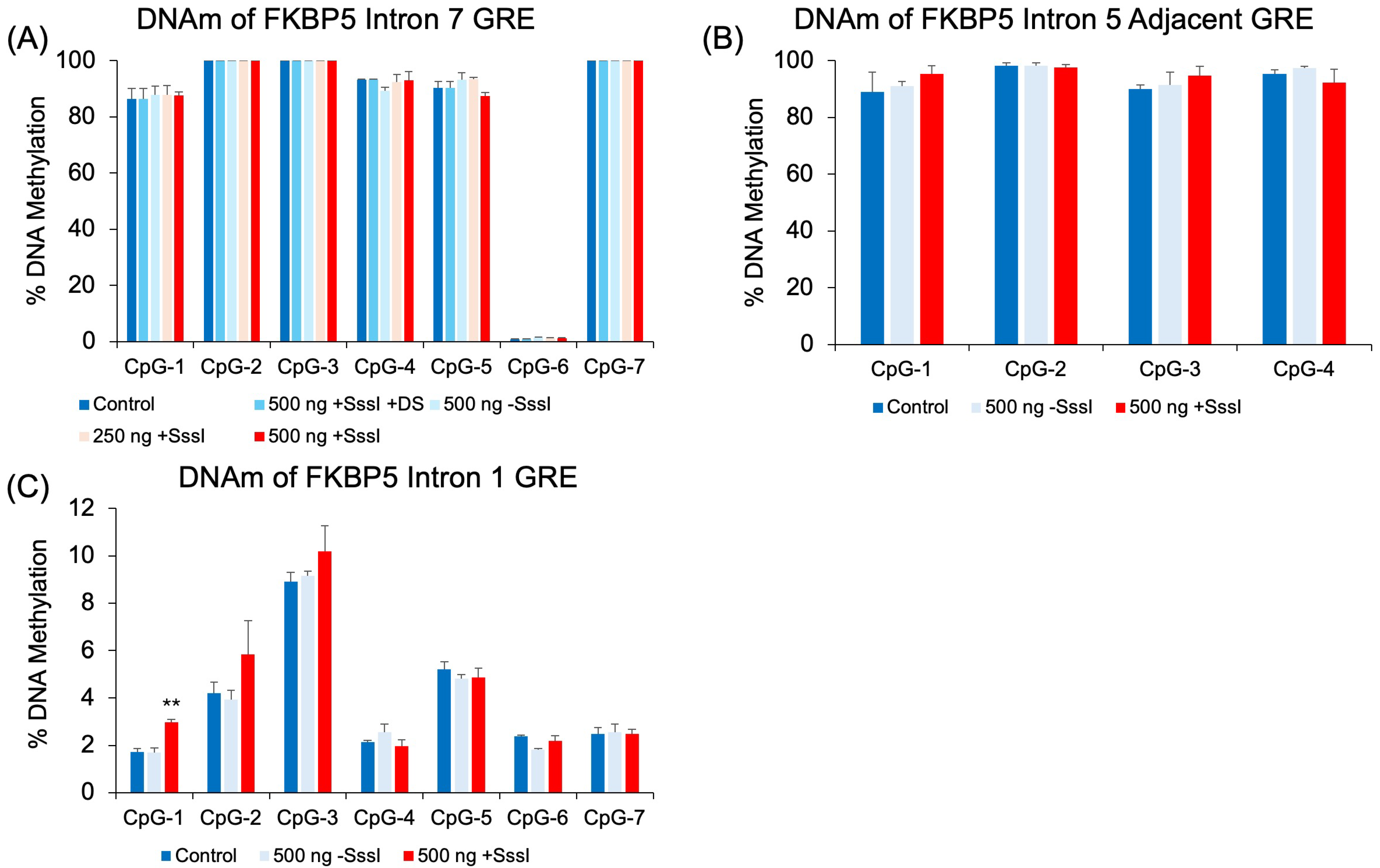
2.4. Effect of DNA Probes in Non-Targeted Regions
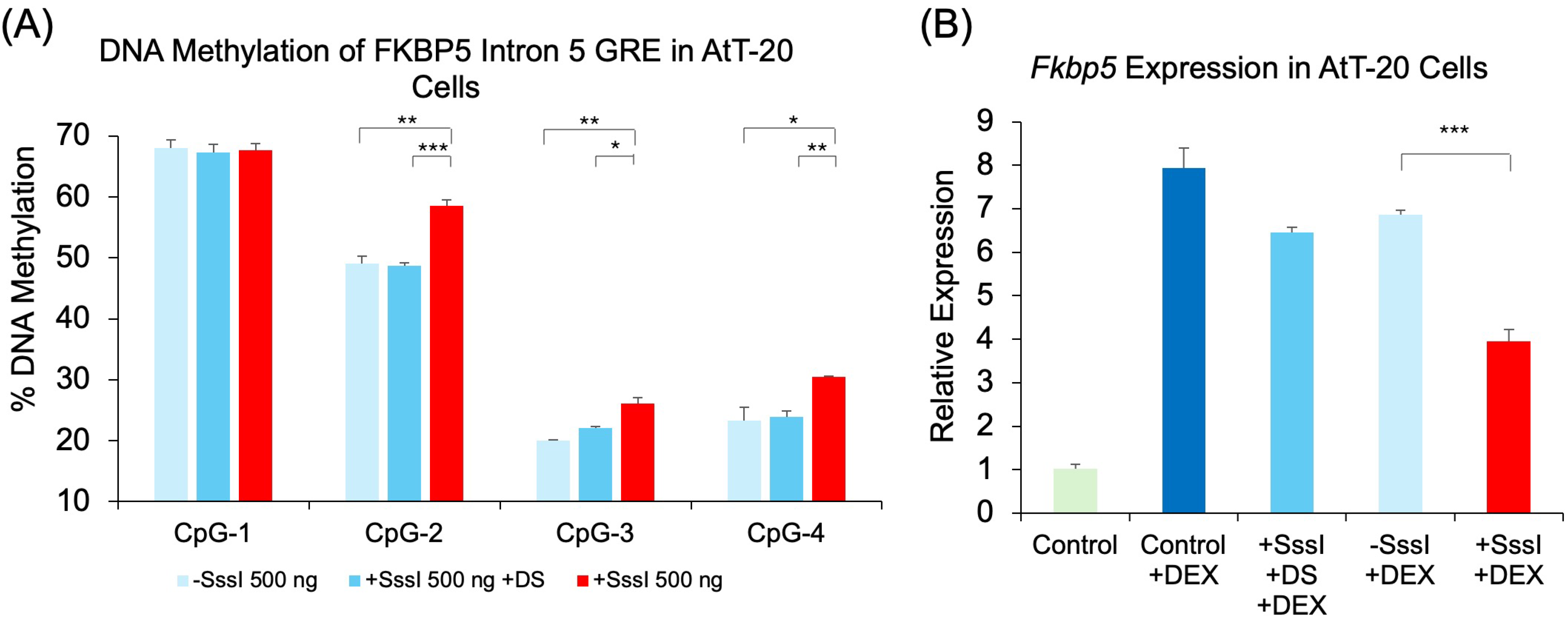
2.5. Epigenetic and Transcriptional Effects of Methylated, Single-Stranded Probe in a Mouse Pituitary Cell Line
2.6. Additional Genomic Target of DNAm Probe: MAOA
3. Discussion
4. Methods
4.1. Probe Design and Amplification
4.2. In Vitro Methylation of DNA Probes
4.3. Cell Culture and Transfection
4.4. DNA Extraction and Methylation Analysis by Bisulfite Pyrosequencing
4.5. Gene Expression Analysis
4.6. Statistics and Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lee, W.; Kim, J.; Yun, J.-M.; Ohn, T.; Gong, Q. MeCP2 regulates gene expression through recognition of H3K27me3. Nat. Commun. 2020, 11, 3140. [Google Scholar] [CrossRef]
- Nishiyama, A.; Nakanishi, M. Navigating the DNA methylation landscape of cancer. Trends Genet. 2021, 37, 1012–1027. [Google Scholar] [CrossRef]
- Shireby, G.; Dempster, E.L.; Policicchio, S.; Smith, R.G.; Pishva, E.; Chioza, B.; Davies, J.P.; Burrage, J.; Lunnon, K.; Vellame, D.S.; et al. DNA methylation signatures of Alzheimer’s disease neuropathology in the cortex are primarily driven by variation in non-neuronal cell-types. Nat. Commun. 2022, 13, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.; Yang, B.; Rothschild, G.; Mann, J.J.; Sanford, L.D.; Tang, X.; Huang, C.; Wang, C.; Zhang, W. Epigenetic regulation in major depression and other stress-related disorders: Molecular mechanisms, clinical relevance and therapeutic potential. Signal Transduct. Target. Ther. 2023, 8, 309. [Google Scholar] [CrossRef]
- Persaud, N.S.; Cates, H.M. The Epigenetics of Anxiety Pathophysiology: A DNA Methylation and Histone Modification Focused Review. Eneuro 2022, 10. [Google Scholar] [CrossRef]
- Katrinli, S.; Wani, A.H.; Maihofer, A.X.; Ratanatharathorn, A.; Daskalakis, N.P.; Montalvo-Ortiz, J.; Núñez-Ríos, D.L.; Zannas, A.S.; Zhao, X.; Aiello, A.E.; et al. Epigenome-wide association studies identify novel DNA methylation sites associated with PTSD: A meta-analysis of 23 military and civilian cohorts. Genome Med. 2024, 16, 147. [Google Scholar] [CrossRef]
- Rahman, M.F.; McGowan, P.O. Cell-type-specific epigenetic effects of early life stress on the brain. Transl. Psychiatry 2022, 12, 326. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, W.; Xu, H.; Ellenbroek, B.; Dai, J.; Wang, L.; Yan, C.; Wang, W. The Changes of Histone Methylation Induced by Adolescent Social Stress Regulate the Resting-State Activity in mPFC. Research 2023, 6, 0264. [Google Scholar] [CrossRef]
- Erdmann, R.M.; Picard, C.L. RNA-directed DNA Methylation. PLoS Genet 2020, 16, e1009034. [Google Scholar] [CrossRef] [PubMed Central]
- Hu, C.; Liu, X.; Zeng, Y.; Liu, J.; Wu, F. DNA methyltransferase inhibitors combination therapy for the treatment of solid tumor: Mechanism and clinical application. Clin. Epigenetics 2021, 13, 166. [Google Scholar] [CrossRef]
- Liu, X.S.; Wu, H.; Ji, X.; Stelzer, Y.; Wu, X.; Czauderna, S.; Shu, J.; Dadon, D.; Young, R.A.; Jaenisch, R. Editing DNA Methylation in the Mammalian Genome. Cell 2016, 167, 233–247.e17. [Google Scholar] [CrossRef] [PubMed]
- Vojta, A.; Dobrinić, P.; Tadić, V.; Bočkor, L.; Korać, P.; Julg, B.; Klasić, M.; Zoldoš, V. Repurposing the CRISPR-Cas9 system for targeted DNA methylation. Nucleic Acids Res. 2016, 44, 5615–5628. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Tao, Y.; Gao, X.; Zhang, L.; Li, X.; Zou, W.; Ruan, K.; Wang, F.; Xu, G.-L.; Hu, R. A CRISPR-based approach for targeted DNA demethylation. Cell Discov. 2016, 2, 16009. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Koo, T.; Jee, H.-G.; Cho, H.-Y.; Lee, G.; Lim, D.-G.; Shin, H.S.; Kim, J.-S. CRISPR RNAs trigger innate immune responses in human cells. Genome Res. 2018, 28, 367–373. [Google Scholar] [CrossRef]
- Ewaisha, R.; Anderson, K.S. Immunogenicity of CRISPR therapeutics—Critical considerations for clinical translation. Front. Bioeng. Biotechnol. 2023, 11, 1138596. [Google Scholar] [CrossRef]
- Hakim, C.H.; Kumar, S.R.P.; Pérez-López, D.O.; Wasala, N.B.; Zhang, D.; Yue, Y.; Teixeira, J.; Pan, X.; Zhang, K.; Million, E.D.; et al. Cas9-specific immune responses compromise local and systemic AAV CRISPR therapy in multiple dystrophic canine models. Nat. Commun. 2021, 12, 6769. [Google Scholar] [CrossRef]
- Guo, C.; Ma, X.; Gao, F.; Guo, Y. Off-target effects in CRISPR/Cas9 gene editing. Front. Bioeng. Biotechnol. 2023, 11, 1143157. [Google Scholar] [CrossRef]
- Höijer, I.; Emmanouilidou, A.; Östlund, R.; van Schendel, R.; Bozorgpana, S.; Tijsterman, M.; Feuk, L.; Gyllensten, U.; Hoed, M.D.; Ameur, A. CRISPR-Cas9 induces large structural variants at on-target and off-target sites in vivo that segregate across generations. Nat. Commun. 2022, 13, 627. [Google Scholar] [CrossRef]
- Frangoul, H.; Altshuler, D.; Cappellini, M.D.; Chen, Y.-S.; Domm, J.; Eustace, B.K.; Foell, J.; De La Fuente, J.; Grupp, S.; Handgretinger, R.; et al. CRISPR-Cas9 Gene Editing for Sickle Cell Disease and β-Thalassemia. N. Engl. J. Med. 2021, 384, 252–260. [Google Scholar] [CrossRef]
- Stadtmauer, E.A.; Fraietta, J.A.; Davis, M.M.; Cohen, A.D.; Weber, K.L.; Lancaster, E.; Mangan, P.A.; Kulikovskaya, I.; Gupta, M.; Chen, F.; et al. CRISPR-engineered T cells in patients with refractory cancer. Science 2020, 367, eaba7365. [Google Scholar] [CrossRef]
- Musunuru, K.; A Grandinette, S.; Wang, X.; Hudson, T.R.; Briseno, K.; Berry, A.M.; Hacker, J.L.; Hsu, A.; A Silverstein, R.; Hille, L.T.; et al. Patient-Specific In Vivo Gene Editing to Treat a Rare Genetic Disease. N. Engl. J. Med. 2025, 392, 2235–2243. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Wu, X.; Chandra, S.; Lyon, C.; Ning, B.; Jiang, L.; Fan, J.; Hu, T.Y. Extracellular vesicles: Emerging tools as therapeutic agent carriers. Acta Pharm. Sin. B 2022, 12, 3822–3842. [Google Scholar] [CrossRef]
- Seifuddin, F.; Wand, G.; Cox, O.; Pirooznia, M.; Moody, L.; Yang, X.; Tai, J.; Boersma, G.; Tamashiro, K.; Zandi, P.; et al. Genome-wide Methyl-Seq analysis of blood-brain targets of glucocorticoid exposure. Epigenetics 2017, 12, 637–652. [Google Scholar] [CrossRef]
- Scammell, J.G.; Denny, W.B.; Valentine, D.L.; Smith, D.F. Overexpression of the FK506-Binding Immunophilin FKBP51 Is the Common Cause of Glucocorticoid Resistance in Three New World Primates. Gen. Comp. Endocrinol. 2001, 124, 152–165. [Google Scholar] [CrossRef] [PubMed]
- Nelson, J.C.; Davis, J.M. DST Studies in Psychotic Depression: A Meta-Analysis. Am. J. Psychiatry 1997, 154, 1497–1503. [Google Scholar] [CrossRef]
- Binder, E.B.; Salyakina, D.; Lichtner, P.; Wochnik, G.M.; Ising, M.; Pütz, B.; Papiol, S.; Seaman, S.; Lucae, S.; Kohli, M.A.; et al. Polymorphisms in FKBP5 are associated with increased recurrence of depressive episodes and rapid response to antidepressant treatment. Nat. Genet. 2004, 36, 1319–1325. [Google Scholar] [CrossRef]
- Tozzi, L.; Farrell, C.; Booij, L.; Doolin, K.; Nemoda, Z.; Szyf, M.; Pomares, F.B.; Chiarella, J.; O'Keane, V.; Frodl, T. Epigenetic Changes of FKBP5 as a Link Connecting Genetic and Environmental Risk Factors with Structural and Functional Brain Changes in Major Depression. Neuropsychopharmacology 2017, 43, 1138–1145. [Google Scholar] [CrossRef]
- Lee, R.S.; Tamashiro, K.L.K.; Yang, X.; Purcell, R.H.; Harvey, A.; Willour, V.L.; Huo, Y.; Rongione, M.; Wand, G.S.; Potash, J.B. Chronic Corticosterone Exposure Increases Expression and Decreases Deoxyribonucleic Acid Methylation of Fkbp5 in Mice. Endocrinology 2010, 151, 4332–4343. [Google Scholar] [CrossRef]
- Cox, O.H.; Song, H.Y.; Garrison-Desany, H.M.; Gadiwalla, N.; Carey, J.L.; Menzies, J.; Lee, R.S. Characterization of glucocorticoid-induced loss of DNA methylation of the stress-response gene Fkbp5 in neuronal cells. Epigenetics 2021, 16, 1377–1397. [Google Scholar] [CrossRef]
- Wochnik, G.M.; Rüegg, J.; Abel, G.A.; Schmidt, U.; Holsboer, F.; Rein, T. FK506-binding Proteins 51 and 52 Differentially Regulate Dynein Interaction and Nuclear Translocation of the Glucocorticoid Receptor in Mammalian Cells. J. Biol. Chem. 2005, 280, 4609–4616. [Google Scholar] [CrossRef]
- Miller, O.; Shakespeare-Finch, J.; Bruenig, D.; Mehta, D. DNA methylation of NR3C1 and FKBP5 is associated with posttraumatic stress disorder, posttraumatic growth, and resilience. Psychol. Trauma Theory Res. Pract. Policy 2020, 12, 750–755. [Google Scholar] [CrossRef] [PubMed]
- Kremer, T.L.; Chen, J.; Buhl, A.; Berhe, O.; Bilek, E.; Geiger, L.S.; Ma, R.; Moessnang, C.; Reichert, M.; Reinhard, I.; et al. Multimodal Associations of FKBP5 Methylation with Emotion-Regulatory Brain Circuits. Biol. Psychiatry 2024, 96, 858–867. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Wang, H.; Yan, A.; Yin, F.; Qiao, X. DNA Methylation in Alcohol Use Disorder. Int. J. Mol. Sci. 2023, 24, 10130. [Google Scholar] [CrossRef]
- Klengel, T.; Mehta, D.; Anacker, C.; Rex-Haffner, M.; Pruessner, J.C.; Pariante, C.M.; Pace, T.W.W.; Mercer, K.B.; Mayberg, H.S.; Bradley, B.; et al. Allele-specific FKBP5 DNA demethylation mediates gene–childhood trauma interactions. Nat. Neurosci. 2013, 16, 33–41. [Google Scholar] [CrossRef]
- Shumay, E.; Logan, J.; Volkow, N.D.; Fowler, J.S. Evidence that the methylation state of the monoamine oxidase A (MAOA) gene predicts brain activity of MAO A enzyme in healthy men. Epigenetics 2012, 7, 1151–1160. [Google Scholar] [CrossRef]
- Xu, Z.; Shi, J.; Chen, Q.; Yang, S.; Wang, Z.; Xiao, B.; Lai, Z.; Jing, Y.; Li, Y.; Li, X. Regulation of de novo and maintenance DNA methylation by DNA methyltransferases in postimplantation embryos. J. Biol. Chem. 2024, 301, 107990. [Google Scholar] [CrossRef]
- Shi, M.-Q.; Xu, Y.; Fu, X.; Pan, D.-S.; Lu, X.-P.; Xiao, Y.; Jiang, Y.-Z. Advances in targeting histone deacetylase for treatment of solid tumors. J. Hematol. Oncol. 2024, 17, 37. [Google Scholar] [CrossRef]
- Kimura, H.; Cook, P.R. Kinetics of core histones in living human cells: Little exchange of H3 and H4 and some rapid exchange of H2B. J. Cell Biol. 2001, 153, 1341–1353. [Google Scholar] [CrossRef] [PubMed Central]
- Kassem, S.; Ferrari, P.; Hughes, A.L.; Soudet, J.; Rando, O.J.; Strubin, M. Histone exchange is associated with activator function at transcribed promoters and with repression at histone loci. Sci. Adv. 2020, 6, eabb0333. [Google Scholar] [CrossRef]
- Wang, C.; Liu, H. Factors influencing degradation kinetics of mRNAs and half-lives of microRNAs, circRNAs, lncRNAs in blood in vitro using quantitative PCR. Sci. Rep. 2022, 12, 7259. [Google Scholar] [CrossRef]
- Marzi, M.J.; Ghini, F.; Cerruti, B.; de Pretis, S.; Bonetti, P.; Giacomelli, C.; Gorski, M.M.; Kress, T.; Pelizzola, M.; Muller, H.; et al. Degradation dynamics of microRNAs revealed by a novel pulse-chase approach. Genome Res. 2016, 26, 554–565. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.; Ga, Y.J.; Kim, S.H.; Cho, Y.H.; Kim, J.W.; Kim, C.; Yeh, J.-Y. Small interfering RNA (siRNA)-based therapeutic applications against viruses: Principles, potential, and challenges. J. Biomed. Sci. 2023, 30, 88. [Google Scholar] [CrossRef] [PubMed]
- Neumeier, J.; Meister, G. siRNA Specificity: RNAi Mechanisms and Strategies to Reduce Off-Target Effects. Front. Plant Sci. 2021, 11, 526455. [Google Scholar] [CrossRef] [PubMed]
- Murphy, D.E.; de Jong, O.G.; Brouwer, M.; Wood, M.J.; Lavieu, G.; Schiffelers, R.M.; Vader, P. Extracellular vesicle-based therapeutics: Natural versus engineered targeting and trafficking. Exp. Mol. Med. 2019, 51, 1–12. [Google Scholar] [CrossRef]
- Koh, E.; Lee, E.J.; Nam, G.-H.; Hong, Y.; Cho, E.; Yang, Y.; Kim, I.-S. Exosome-SIRPα, a CD47 blockade increases cancer cell phagocytosis. Biomaterials 2017, 121, 121–129. [Google Scholar] [CrossRef]
- Alvarez-Erviti, L.; Seow, Y.; Yin, H.; Betts, C.; Lakhal, S.; Wood, M.J.A. Delivery of siRNA to the mouse brain by systemic injection of targeted exosomes. Nat. Biotechnol. 2011, 29, 341–345. [Google Scholar] [CrossRef]
- Xitong, D.; Xiaorong, Z. Targeted therapeutic delivery using engineered exosomes and its applications in cardiovascular diseases. Gene 2016, 575, 377–384. [Google Scholar] [CrossRef]
- Haraszti, R.A.; Didiot, M.; Sapp, E.; Leszyk, J.; Shaffer, S.A.; Rockwell, H.E.; Gao, F.; Narain, N.R.; DiFiglia, M.; Kiebish, M.A.; et al. High-resolution proteomic and lipidomic analysis of exosomes and microvesicles from different cell sources. J. Extracell. Vesicles 2016, 5, 32570. [Google Scholar] [CrossRef]
- Lin, Y.; Lu, Y.; Li, X. Biological characteristics of exosomes and genetically engineered exosomes for the targeted delivery of therapeutic agents. J. Drug Target. 2019, 28, 129–141. [Google Scholar] [CrossRef]
- Sato, Y.T.; Umezaki, K.; Sawada, S.; Mukai, S.-A.; Sasaki, Y.; Harada, N.; Shiku, H.; Akiyoshi, K. Engineering hybrid exosomes by membrane fusion with liposomes. Sci. Rep. 2016, 6, 21933. [Google Scholar] [CrossRef]
- Lu, Y.; Chen, T.; Lin, H.; Chen, Y.; Lin, Y.; Le, D.; Huang, Y.; Wang, A.H.; Lee, C.; Ling, T. Small Extracellular Vesicles Engineered Using Click Chemistry to Express Chimeric Antigen Receptors Show Enhanced Efficacy in Acute Liver Failure. J. Extracell. Vesicles 2025, 14, e70044. [Google Scholar] [CrossRef] [PubMed]
- Ciferri, M.C.; Bruno, S.; Rosenwasser, N.; Gorgun, C.; Reverberi, D.; Gagliani, M.C.; Cortese, K.; Grange, C.; Bussolati, B.; Quarto, R.; et al. Standardized Method to Functionalize Plasma-Extracellular Vesicles via Copper-Free Click Chemistry for Targeted Drug Delivery Strategies. ACS Appl. Bio Mater. 2024, 7, 827–838. [Google Scholar] [CrossRef] [PubMed]
- Renbaum, P.; Abrahamove, D.; Fainsod, A.; Wilson, G.G.; Rottem, S.; Razin, A. Cloning, characterization, and expression in Escherichia coli of the gene coding for the CpG DNA methylase from Spiroplasma sp. strain MQ1(M.SssI). Nucleic Acids. Res. 1990, 18, 1145–1152. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Duis, J.; Cox, O.H.; Ji, Y.; Seifuddin, F.; Lee, R.S.; Wang, X. Effect of Genotype and Maternal Affective Disorder on Intronic Methylation of FK506 Binding Protein 5 in Cord Blood DNA. Front. Genet. 2018, 9, 648. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Probe Primers | Sequence | Size |
|---|---|---|
| Human FKBP5 Probe-Forward | 5′-AAAGTCAAACCAAACCAAATTACC -3′ | 256 bp |
| Human FKBP5 Probe-Reverse | 5′-TTTGTTACTGCTGTGCACTCTCT -3′ | |
| Human MAOA Probe-Forward | 5′-TCGACGTAGTCGTGATCGG -3′ | 170 bp |
| Human MAOA Probe-Reverse | 5′-GCAGGATATGGGGCCAAG -3′ | |
| Mouse Fkbp5 Probe-Forward | 5′-CAGACACCAGCTACTATAATTAG -3′ | 239 bp |
| Mouse Fkbp5 Probe-Reverse | 5′-GCACATGAACTCGATGTGCTGACA -3′ | |
| Pyrosequencing Primers | Sequence | Size |
| Human FKBP5 Intron 5 Outside-A | GGTAGAGAAAGAAATAAATAAGTTA | 286 bp |
| Human FKBP5 Intron 5 Outside-B | TTCTTACATTTCATTTTTATTACTACTA | |
| Human FKBP5 Intron 5 Inside-A * | AAGATTATGTAATTTAAAGGGGGAGGG | |
| Human FKBP5 Intron 5 Inside-B | CTCTCTTTCCTTTTTTCCCCCCTAT | |
| Human FKBP5 Intron 5 Pyro 1 | TCTTTCCTTTTTTCCCCCCTATT | |
| Human FKBP5 Intron 5 Pyro 2 | CAATTTAAATAATATTTTACAACT | |
| Human FKBP5 Intron 5_2 Out-A | ATTTAATTGGTTTGGGTGTTAGAA | 406 bp |
| Human FKBP5 Intron 5_2 Out-B | CCTCTCAATACTTTCAACCACA | |
| Human FKBP5 Intron 5_2 In-A * | GAGAATTATTGTATTGGAGGTT | |
| Human FKBP5 Intron 5_2 In-B | ATTCTACAAATTCCAATTATTAAC | |
| Human FKBP5 Intron 5_2 Pyro 1 | GTATTGGAGGTTTATTGGTT | |
| Human FKBP5 Intron 5_2 Pyro 2 | TAGATGATTATGAGTTTGGAGTT | |
| Human FKBP5 Intron 5_2 Pyro 3 | GTTTAAGTTTTTTTTATATTTGTT | |
| Human FKBP5 Intron 5_2 Pyro 4 | GATTTGGAGAGGGAAAGGAGGT | |
| Human FKBP5 Intron 1 Out-A | AGTTTAAATTGTTTTATGTAGAATTTATTGA | 350 bp |
| Human FKBP5 Intron 1 Out-B | TCACTCCCAAACCATACC | |
| Human FKBP5 Intron 1 Inside-A | GTTTTGAATTATATTGAAGGGTATTT | |
| Human FKBP5 Intron 1 Inside-B * | CAAAACTCCTTATACTCTTCTATTCTAA | |
| Human FKBP5 Intron 1 Pyro 1 | GTAGAATTYGATTTTAGAGA | |
| Human FKBP5 Intron 7 Outside-A | AGAGTGAAATTGAGATGGAAATATGT | 503 bp |
| Human FKBP5 Intron 7 Outside-B | AATTTCTTCTCCATCCACTTCCTATA | |
| Human FKBP5 Intron 7 Inside-A | AGGAGGTATGTTGTTTTTGGAATTTAAG | |
| Human FKBP5 Intron 7 Inside-B * | AATTTATCTCTTACCTCCAACACT | |
| Human FKBP5 Intron 7 Pyro 1 | GGAGAAGTATAAAAAAAAAATGG | |
| Human FKBP5 Intron 7 Pyro 2 | GTTATAGAGTTTAGTGGTTT | |
| Human FKBP5 Intron 7 Pyro 3 | GGAGTTATAGTGTAGGTTTT | |
| Human FKBP5 Intron 7 Pyro 4 | TTAAGGAGTTATTTGGTAGA | |
| Human FKBP5 Intron 7 Pyro 5 | TGATATATAGGAATAAAATAAGAAT | |
| Human MAOA Outside-A | GATTTAGGAGYGTGTTAGTTAAAGT | 278 bp |
| Human MAOA Outside-B | TTATTATATCTACCTCCCCCAATC | |
| Human MAOA Inside-A | AGTTAAAGTATGGAGAATTAAG | |
| Human MAOA Inside-B * | ATCTACCTCCCCCAATCACACCACCAAC | |
| Human MAOA Pyro 1 | AAAGTATGGAGAATTAAGAGAAGG | |
| Human MAOA Pyro 2 | GAGTATYGYGGGTTATATG | |
| Human MAOA Pyro 3 | AGGTGGTATTTTAGGTTAGTGTGGA | |
| Mouse Fkbp5 Intron 5 Outside-A | GATGATTAGTTTTTTTTAGTAGTGATGT | 308 bp |
| Mouse Fkbp5 Intron 5 Outside-B | CTTATTATTCTCTTACTACCCTAA | |
| Mouse Fkbp5 Intron 5 Inside-A | TAGTTTTTGGGGAAGAGTGTAGAGTTAT | |
| Mouse Fkbp5 Intron 5 Inside-B * | ATTTTAAAAAACACAAAACACCCTATT | |
| Mouse Fkbp5 Intron 5 Pyro 1 | AGAAAAGGGAAAGTAGG | |
| Mouse Fkbp5 Intron 5 Pyro 2 | TAGTTTTTGTTATTGTTGTATG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Modi, N.; Guo, J.; Lee, R.A.; Greenstein, A.; Lee, R.S. Targeted DNA Methylation Using Modified DNA Probes: A Potential Therapeutic Tool for Depression and Stress-Related Disorders. Int. J. Mol. Sci. 2025, 26, 5643. https://doi.org/10.3390/ijms26125643
Modi N, Guo J, Lee RA, Greenstein A, Lee RS. Targeted DNA Methylation Using Modified DNA Probes: A Potential Therapeutic Tool for Depression and Stress-Related Disorders. International Journal of Molecular Sciences. 2025; 26(12):5643. https://doi.org/10.3390/ijms26125643
Chicago/Turabian StyleModi, Nishtaa, Jeffrey Guo, Ryan A. Lee, Alisha Greenstein, and Richard S. Lee. 2025. "Targeted DNA Methylation Using Modified DNA Probes: A Potential Therapeutic Tool for Depression and Stress-Related Disorders" International Journal of Molecular Sciences 26, no. 12: 5643. https://doi.org/10.3390/ijms26125643
APA StyleModi, N., Guo, J., Lee, R. A., Greenstein, A., & Lee, R. S. (2025). Targeted DNA Methylation Using Modified DNA Probes: A Potential Therapeutic Tool for Depression and Stress-Related Disorders. International Journal of Molecular Sciences, 26(12), 5643. https://doi.org/10.3390/ijms26125643