
Biomarkers of Depression among Adolescent Girls: BDNF and Epigenetics

 , , ,
, , ,  , ,
, ,
Abstract
1. Introduction
2. Results
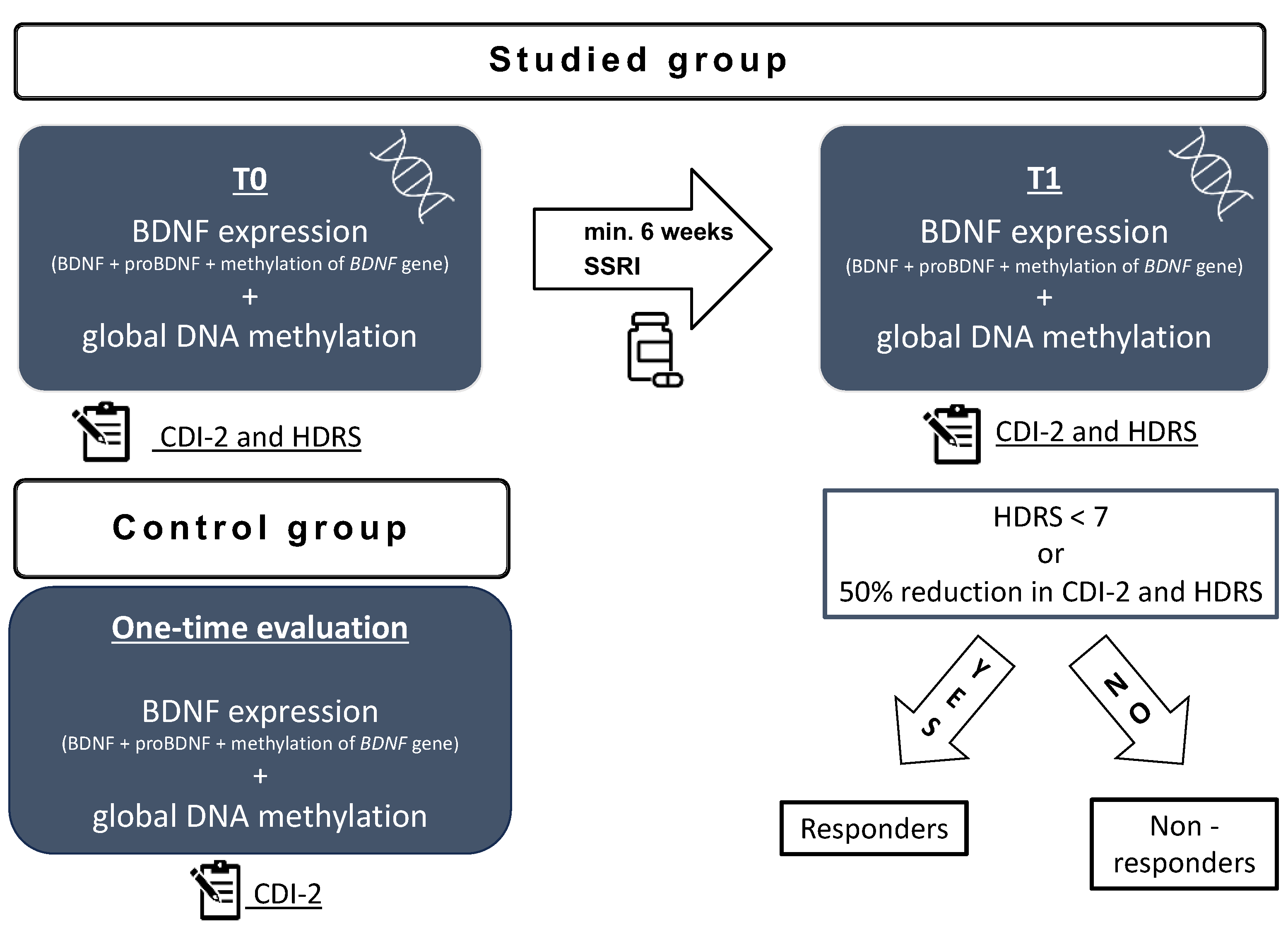
2.1. Recruitment
2.2. Clinical Characteristics
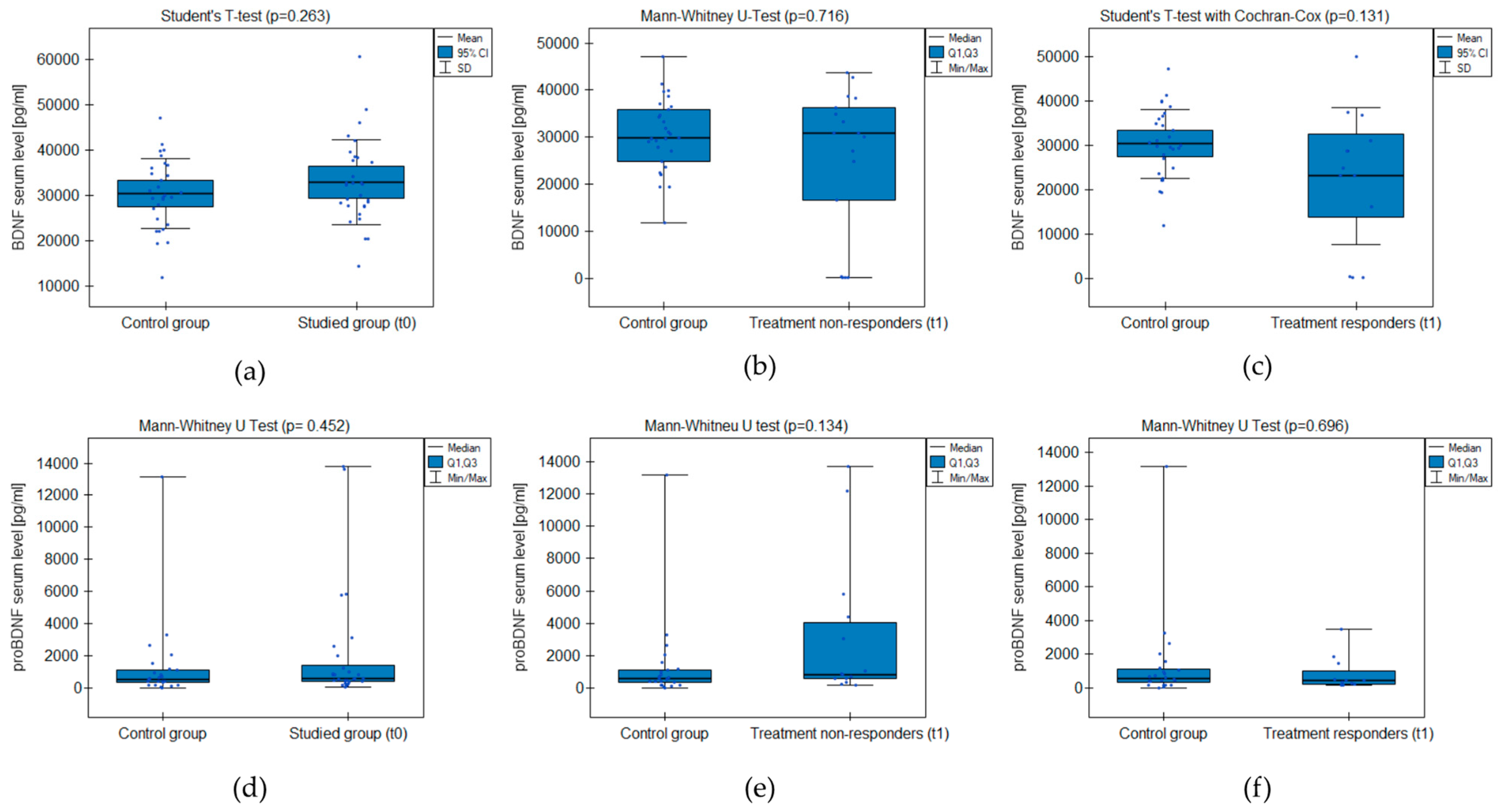
2.3. Biochemical and Molecular Results
2.3.1. BDNF and proBDNF Serum Levels
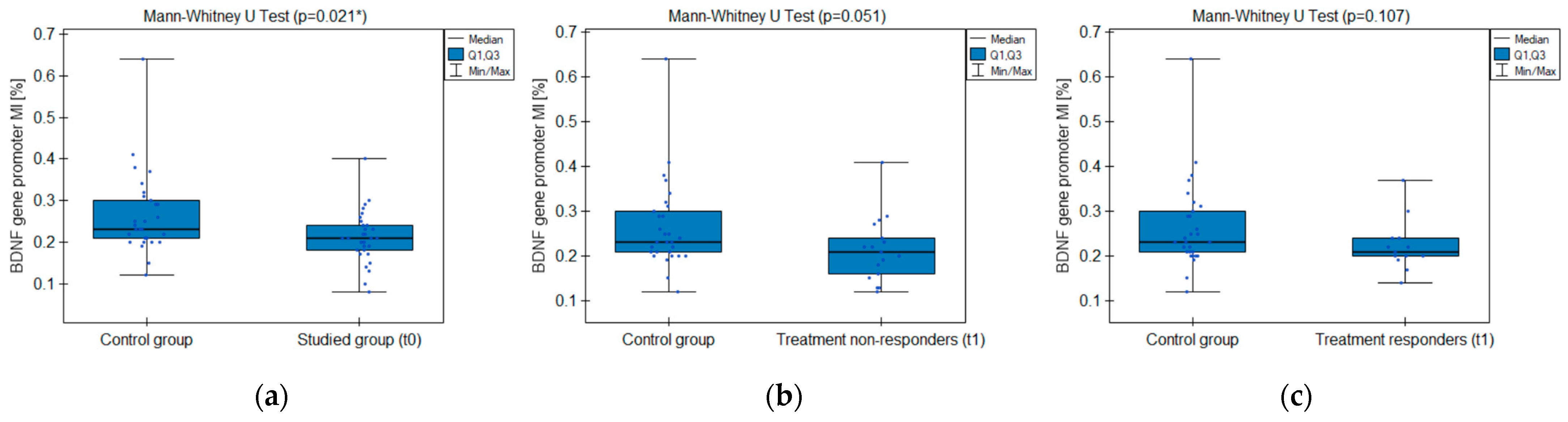
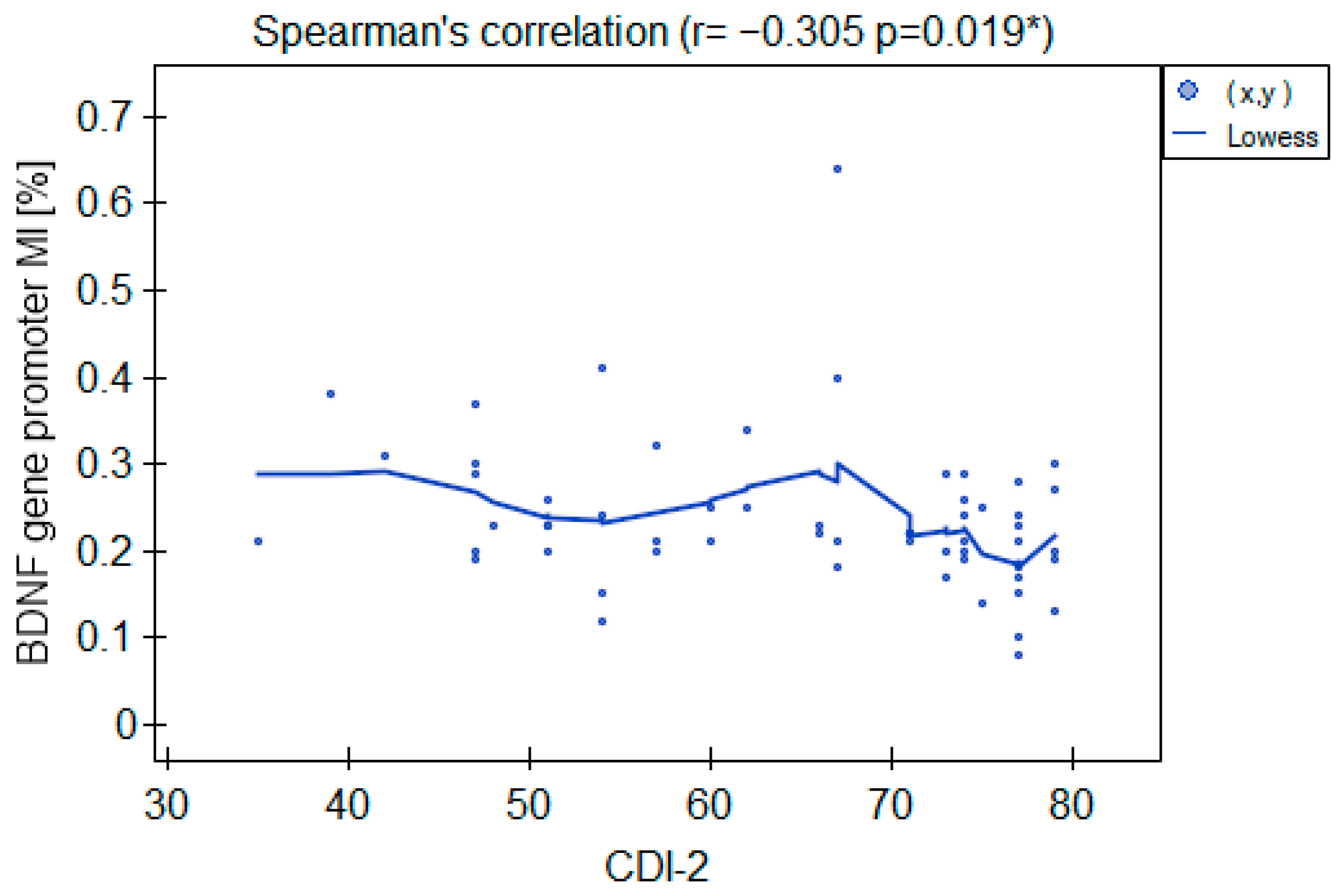
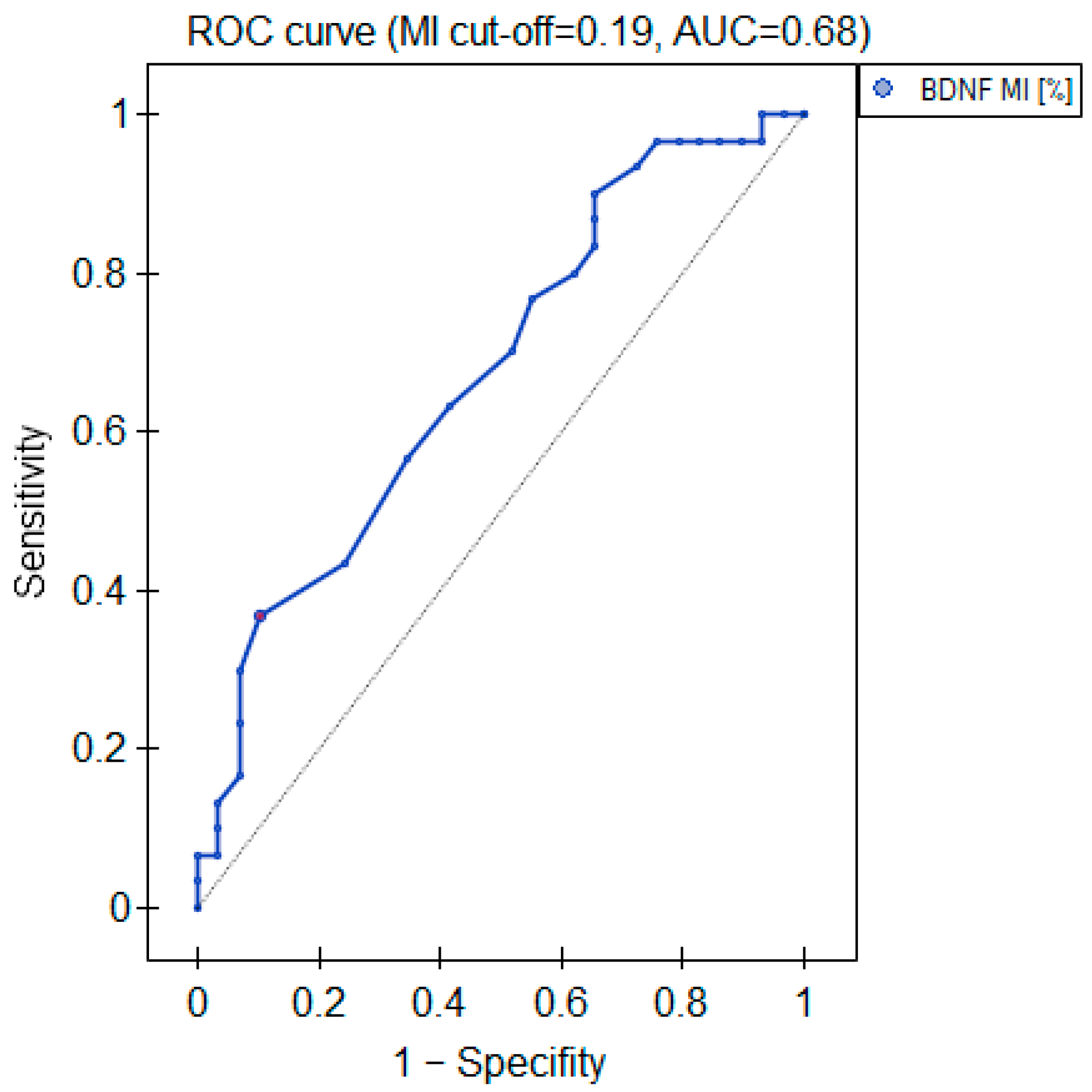
2.3.2. BDNF Gene Promoter Methylation
2.3.3. Global DNA Methylation (%-5mc)
3. Discussion
4. Materials and Methods
4.1. Ethical Declaration
4.2. Participants
4.3. Clinical Assessment
4.4. Biochemical Assessment
4.4.1. Global DNA Methylation
4.4.2. BDNF Methylation Analysis
4.4.3. BDNF and proBDNF Serum Levels
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Malhi, G.S.; Mann, J.J. Seminar Depression. Lancet 2018, 392, 2299–2312. [Google Scholar] [CrossRef]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders; American Psychiatric Association: Arlington, VA, USA, 2013. [Google Scholar] [CrossRef]
- Shorey, S.; Ng, E.D.; Wong, C.H.J. Global prevalence of depression and elevated depressive symptoms among adolescents: A systematic review and meta-analysis. Br. J. Clin. Psychol. 2022, 61, 287–305. [Google Scholar] [CrossRef]
- Mullen, S. Major depressive disorder in children and adolescents. Ment. Health Clin. 2018, 8, 275–283. [Google Scholar] [CrossRef]
- Suicide. Available online: https://www.who.int/news-room/fact-sheets/detail/suicide (accessed on 5 August 2023).
- Rice, F.; Riglin, L.; Lomax, T.; Souter, E.; Potter, R.; Smith, D.J.; Thapar, A.K.; Thapar, A. Adolescent and adult differences in major depression symptom profiles. J. Affect. Disord. 2019, 243, 175–181. [Google Scholar] [CrossRef]
- Bylund, D.B.; Reed, A.L. Childhood and adolescent depression: Why do children and adults respond differently to antidepressant drugs? Neurochem. Int. 2007, 51, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Fava, M. Diagnosis and definition of treatment-resistant depression. Biol. Psychiatry 2003, 53, 649–659. [Google Scholar] [CrossRef]
- Rice, F. The genetics of depression in childhood and adolescence. Curr. Psychiatry Rep. 2009, 11, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Nie, Z.; Shu, H.; Kuang, Y.; Chen, X.; Cheng, J.; Yu, S.; Liu, H. The Role of BDNF on Neural Plasticity in Depression. Front. Cell. Neurosci. 2020, 14, 82. [Google Scholar] [CrossRef]
- Miyanishi, H.; Nitta, A. A role of BDNF in the depression pathogenesis and a potential target as antidepressant: The modulator of stress sensitivity ‘shati/Nat8l-BDNF system’ in the dorsal striatum. Pharmaceuticals 2021, 14, 889. [Google Scholar] [CrossRef]
- Tripp, A.; Oh, H.; Guilloux, J.-P.; Martinowich, K.; Lewis, D.A.; Sibille, E. Brain-Derived Neurotrophic Factor Signaling and Subgenual Anterior Cingulate Cortex Dysfunction in Major Depressive Disorder. Am. J. Psychiatry 2012, 169, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, Y.; Rizavi, H.S.; Conley, R.R.; Roberts, R.C.; Tamminga, C.A.; Pandey, G.N. Altered gene expression of brain-derived neurotrophic factor and receptor tyrosine kinase B in postmortem brain of suicide subjects. Arch. Gen. Psychiatry 2003, 60, 804–815. [Google Scholar] [CrossRef]
- Youssef, M.M.; Underwood, M.D.; Huang, Y.-Y.; Hsiung, S.-C.; Liu, Y.; Simpson, N.R.; Bakalian, M.J.; Rosoklija, G.B.; Dwork, A.J.; Arango, V.; et al. Association of BDNF Val66MET polymorphism and brain BDNF levels with major depression and suicide. Int. J. Neuropsychopharmacol. 2018, 21, 528–538. [Google Scholar] [CrossRef] [PubMed]
- Molendijk, M.L.; Bus, B.A.; Spinhoven, P.; Penninx, B.W.; Kenis, G.; Prickaerts, J.; Voshaar, R.C.O.; Elzinga, B.M. Serum levels of brain-derived neurotrophic factor in major depressive disorder: State-trait issues, clinical features and pharmacological treatment. Mol. Psychiatry 2011, 16, 1088–1095. [Google Scholar] [CrossRef] [PubMed]
- Polyakova, M.; Stuke, K.; Schuemberg, K.; Mueller, K.; Schoenknecht, P.; Schroeter, M.L. BDNF as a biomarker for successful treatment of mood disorders: A systematic & quantitative meta-analysis. J. Affect. Disord. 2015, 174, 432–440. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Pang, P.T.; Woo, N.H. The yin and yang of neurotrophin action. Nat. Rev. Neurosci. 2005, 6, 603–614. [Google Scholar] [CrossRef] [PubMed]
- Cohen-Cory, S.; Kidane, A.H.; Shirkey, N.J.; Marshak, S. Brain-derived neurotrophic factor and the development of structural neuronal connectivity. Dev. Neurobiol. 2010, 70, 271–288. [Google Scholar] [CrossRef] [PubMed]
- Costa, R.O.; Perestrelo, T.; Almeida, R.D. PROneurotrophins and CONSequences. Mol. Neurobiol. 2018, 55, 2934–2951. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.R.; Zhang, X.Y.; Liu, Y.; Du, J.Y.; Liang, R.; Yu, M.; Zhang, F.Q.; Mu, X.F.; Li, F.; Zhou, L.; et al. Antidepressant Drugs Correct the Imbalance Between proBDNF/p75NTR/Sortilin and Mature BDNF/TrkB in the Brain of Mice with Chronic Stress. Neurotox. Res. 2020, 37, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Xiong, J.; Lim, Y.; Ruan, Y.; Huang, C.; Zhu, Y.; Zhong, J.-H.; Xiao, Z.; Zhou, X.-F. Upregulation of blood proBDNF and its receptors in major depression. J. Affect. Disord. 2013, 150, 776–784. [Google Scholar] [CrossRef]
- Webb, L.M.; Phillips, K.E.; Ho, M.C.; Veldic, M.; Blacker, C.J. The relationship between DNA methylation and antidepressant medications: A systematic review. Int. J. Mol. Sci. 2020, 21, 826. [Google Scholar] [CrossRef]
- Pruunsild, P.; Kazantseval, A.; Aid, T.; Palm, K.; Timmusk, T. Dissecting the human BDNF locus: Bidirectional transcription, complex splicing, and multiple promoters. Genomics 2007, 90, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Aid, T.; Kazantseva, A.; Piirsoo, M.; Palm, K.; Timmusk, T. Mouse and rat BDNF gene structure and expression revisited. J. Neurosci. Res. 2007, 85, 525–535. [Google Scholar] [CrossRef]
- Martinowich, K.; Hattori, D.; Wu, H.; Fouse, S.; He, F.; Hu, Y.; Fan, G.; Sun, Y.E. DNA Methylation-Related Chromatin Remodeling in Activity-Dependent Bdnf Gene Regulation. Science 2003, 302, 890–893. [Google Scholar] [CrossRef]
- Januar, V.; Ancelin, M.L.; Ritchie, K.; Saffery, R.; Ryan, J. BDNF promoter methylation and genetic variation in late-life depression. Transl. Psychiatry 2015, 5, e619. [Google Scholar] [CrossRef]
- Lin, P.-Y.; Tseng, P.T.; Lee, Y.; Hung, C.-F.; Lung, F.-W.; Chen, C.-S.; Chong, M.-Y. Age-associated decrease in global DNA methylation in patients with major depression. Neuropsychiatr. Dis. Treat. 2014, 10, 2105–2114. [Google Scholar] [CrossRef][Green Version]
- Reszka, E.; Jabłońska, E.; Lesicka, M.; Wieczorek, E.; Kapelski, P.; Szczepankiewicz, A.; Pawlak, J.; Dmitrzak-Węglarz, M. An altered global DNA methylation status in women with depression. J. Psychiatr. Res. 2021, 137, 283–289. [Google Scholar] [CrossRef]
- Essex, M.J.; Boyce, W.T.; Hertzman, C.; Lam, L.L.; Armstrong, J.M.; Neumann, S.M.A.; Kobor, M.S. Epigenetic Vestiges of Early Developmental Adversity: Childhood Stress Exposure and DNA Methylation in Adolescence. Child Dev. 2013, 84, 58–75. [Google Scholar] [CrossRef]
- Zwolińska, W.; Dmitrzak-Węglarz, M.; Słopień, A. Biomarkers in Child and Adolescent Depression. Child Psychiatry Hum. Dev. 2023, 54, 266–281. [Google Scholar] [CrossRef] [PubMed]
- Dincheva, I.; Lynch, N.B.; Lee, F.S. The Role of BDNF in the Development of Fear Learning. Depress. Anxiety 2016, 33, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Pallavi, P.; Sagar, R.; Mehta, M.; Sharma, S.; Subramanium, A.; Shamshi, F.; Sengupta, U.; Qadri, R.; Pandey, R.M.; Mukhopadhyay, A.K. Serum neurotrophic factors in adolescent depression: Gender difference and correlation with clinical severity. J. Affect. Disord. 2013, 150, 415–423. [Google Scholar] [CrossRef]
- Bilgiç, A.; Sadıç, Ç.Ç.; Kılınç, İ.; Akça, Ö.F. Exploring the association between depression, suicidality and serum neurotrophin levels in adolescents. Int. J. Psychiatry Clin. Pract. 2020, 24, 143–150. [Google Scholar] [CrossRef]
- Wrigglesworth, J.; Ryan, J.; Vijayakumar, N.; Whittle, S. Brain-derived neurotrophic factor DNA methylation mediates the association between neighborhood disadvantage and adolescent brain structure. Psychiatry Res. Neuroimaging 2019, 285, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Braithwaite, E.C.; Kundakovic, M.; Ramchandani, P.G.; Murphy, S.E.; Champagne, F.A. Maternal prenatal depressive symptoms predict infant NR3C1 1F and BDNF IV DNA methylation. Epigenetics 2015, 10, 408–417. [Google Scholar] [CrossRef] [PubMed]
- Fuchikami, M.; Morinobu, S.; Segawa, M.; Okamoto, Y.; Yamawaki, S.; Ozaki, N.; Inoue, T.; Kusumi, I.; Koyama, T.; Tsuchiyama, K.; et al. DNA methylation profiles of the brain-derived neurotrophic factor (BDNF) gene as a potent diagnostic biomarker in major depression. PLoS ONE 2011, 6, e23881. [Google Scholar] [CrossRef]
- Bakusic, J.; Vrieze, E.; Ghosh, M.; Pizzagalli, D.A.; Bekaert, B.; Claes, S.; Godderis, L. Interplay of Val66Met and BDNF methylation: Effect on reward learning and cognitive performance in major depression. Clin. Epigenetics 2021, 13, 149. [Google Scholar] [CrossRef]
- Carlberg, L.; Scheibelreiter, J.; Hassler, M.R.; Schloegelhofer, M.; Schmoeger, M.; Ludwig, B.; Kasper, S.; Aschauer, H.; Egger, G.; Schosser, A. Brain-derived neurotrophic factor (BDNF)—Epigenetic regulation in unipolar and bipolar affective disorder. J. Affect. Disord. 2014, 168, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Dennis, K.E.; Levitt, P. Regional expression of brain derived neurotrophic factor (BDNF) is correlated with dynamic patterns of promoter methylation in the developing mouse forebrain. Mol. Brain Res. 2005, 140, 1–9. [Google Scholar] [CrossRef]
- Suri, D.; Veenit, V.; Sarkar, A.; Thiagarajan, D.; Kumar, A.; Nestler, E.J.; Galande, S.; Vaidya, V.A. Early stress evokes age-dependent biphasic changes in hippocampal neurogenesis, BDNF expression, and cognition. Biol. Psychiatry 2013, 73, 658–666. [Google Scholar] [CrossRef]
- Miguez, M.J.; Bueno, D.; Espinoza, L.; Chan, W.; Perez, C. Among Adolescents, BDNF and Pro-BDNF Lasting Changes with Alcohol Use Are Stage Specific. Neural Plast. 2020, 2020, 3937627. [Google Scholar] [CrossRef]
- Södersten, K.; Pålsson, E.; Ishima, T.; Funa, K.; Landén, M.; Hashimoto, K.; Ågren, H. Abnormality in serum levels of mature brain-derived neurotrophic factor (BDNF) and its precursor proBDNF in mood-stabilized patients with bipolar disorder: A study of two independent cohorts. J. Affect. Disord. 2014, 160, 1–9. [Google Scholar] [CrossRef]
- Yoshida, T.; Ishikawa, M.; Iyo, M.; Hashimoto, K. Serum Levels of Mature Brain-Derived Neurotrophic Factor (BDNF) and Its Precursor proBDNF in Healthy Subjects. Open Clin. Chem. J. 2012, 5, 7–12. [Google Scholar] [CrossRef]
- Lee, J.; Lee, K.H.; Kim, S.H.; Han, J.Y.; Hong, S.-B.; Cho, S.-C.; Kim, J.-W.; Brent, D. Early changes of serum BDNF and SSRI response in adolescents with major depressive disorder. J. Affect. Disord. 2020, 265, 325–332. [Google Scholar] [CrossRef]
- Cubero-Millán, I.; Ruiz-Ramos, M.-J.; Molina-Carballo, A.; Martínez-Serrano, S.; Fernández-López, L.; Machado-Casas, I.; Tortosa-Pinto, P.; Ruiz-López, A.; Luna-Del-Castillo, J.-D.; Uberos, J.; et al. BDNF concentrations and daily fluctuations differ among ADHD children and respond differently to methylphenidate with no relationship with depressive symptomatology. Psychopharmacology 2017, 234, 267–279. [Google Scholar] [CrossRef]
- Simsek, S.; Uysal, C.; Kaplan, I.; Yuksel, T.; Aktas, H. BDNF and cortisol levels in children with or without post-traumatic stress disorder after sustaining sexual abuse. Psychoneuroendocrinology 2015, 56, 45–51. [Google Scholar] [CrossRef]
- Lee, B.; Shin, E.; Song, I.; Chang, B. Depression in Adolescence and Brain-Derived Neurotrophic Factor. Front. Mol. Neurosci. 2022, 15, 947192. [Google Scholar] [CrossRef]
- Sasaki, T.; Niitsu, T.; Hashimoto, T.; Kanahara, N.; Shiina, A.; Hasegawa, T.; Kimura, H.; Ishikawa, M.; Tone, J.; Yamauchi, A.; et al. Decreased levels of serum brain-derived neurotrophic factor in male pediatric patients with depression. Open Clin. Chem. J. 2011, 4, 28–33. [Google Scholar] [CrossRef]
- Tsuchimine, S.; Saito, M.; Kaneko, S.; Yasui-Furukori, N. Decreased serum levels of polyunsaturated fatty acids and folate, but not brain-derived neurotrophic factor, in childhood and adolescent females with depression. Psychiatry Res. 2015, 225, 187–190. [Google Scholar] [CrossRef] [PubMed]
- Bath, K.G.; Schilit, A.; Lee, F.S. Stress effects on BDNF expression: Effects of age, sex, and form of stress. Neuroscience 2013, 239, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Carbone, D.L.; Handa, R.J. Sex and stress hormone influences on the expression and activity of brain-derived neurotrophic factor. Neuroscience 2013, 239, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Arosio, B.; Guerini, F.R.; Voshaar, R.C.O.; Aprahamian, I. Blood Brain-Derived Neurotrophic Factor (BDNF) and Major Depression: Do We Have a Translational Perspective? Front. Behav. Neurosci. 2021, 15, 626906. [Google Scholar] [CrossRef] [PubMed]
- Martin, E.M.; Fry, R.C. Environmental Influences on the Epigenome: Exposure-Associated DNA Methylation in Human Populations. Annu. Rev. Public Health 2018, 39, 309–333. [Google Scholar] [CrossRef]
- Cavalli, G.; Heard, E. Advances in epigenetics link genetics to the environment and disease. Nature 2019, 571, 489–499. [Google Scholar] [CrossRef]
- Mitchell, C.; Schneper, L.M.; Notterman, D.A. DNA methylation, early life environment, and health outcomes. Pediatr. Res. 2016, 79, 212–219. [Google Scholar] [CrossRef]
- Faravelli, C.; Scarpato, M.A.; Castellini, G.; Sauro, C.L. Gender differences in depression and anxiety: The role of age. Psychiatry Res. 2013, 210, 1301–1303. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. The ICD-10 Classification of Mental and Behavioural Disorders: Clnical Descriptions and Diagnostic Guidelines; World Health Organization: Geneva, Switzerland, 1992; pp. 119–128.
- Depression in Children and Young People: Identification and Management NICE Guideline. Available online: www.nice.org.uk/guidance/ng134 (accessed on 19 November 2023).
- Aksu, S.; Unlu, G.; Kardesler, A.C.; Cakaloz, B.; Aybek, H. Altered levels of brain-derived neurotrophic factor, proBDNF and tissue plasminogen activator in children with posttraumatic stress disorder. Psychiatry Res. 2018, 268, 478–483. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, M. A rating scale for depression. J. Neurol. Neurosurg. Psychiatry 1960, 23, 56–62. [Google Scholar] [CrossRef]
- Kovacs, M. Children’s Depression Inventory (CDI and CDI 2). In The Encyclopedia of Clinical Psychology; Cautin, R.L., Lilienfeld, S.O., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2015; pp. 1–5. [Google Scholar] [CrossRef]
- Fava, G.A.; Kellner, R.; Munari, F.; Pavan, L. The Hamilton Depression Rating Scale in normals and depressives. Acta Psychiatr. Scand. 1982, 66, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Houghton, S.; Marais, I.; Kyron, M.; Lawrence, D.; Page, A.C.; Gunasekera, S.; Glasgow, K.; Macqueen, L. Screening for depressive symptoms in adolescence: A Rasch analysis of the short-form childhood depression inventory-2 (CDI 2:SR[S]). J. Affect. Disord. 2022, 311, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Bobo, W.V.; Angleró, G.C.; Jenkins, G.; Hall-Flavin, D.K.; Weinshilboum, R.; Biernacka, J.M. Validation of the 17-item Hamilton Depression Rating Scale definition of response for adults with major depressive disorder using equipercentile linking to Clinical Global Impression scale ratings: Analysis of Pharmacogenomic Research Network Antidepressant Medication Pharmacogenomic Study (PGRN-AMPS) data. Hum. Psychopharmacol. 2016, 31, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Miller, S.A.; Dykes, D.D.; Polesky, H.F. A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Res. 1988, 16, 1215. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Studied Group (t0) | Responders (t1) | Non-Responders (t1) | Controls | |
|---|---|---|---|---|
| n | 30 | 13 | 17 | 30 |
| Age [years] | 13.07 (±1.26) | 12.85 (±0.99) | 13.24 (±1.44) | 13.43 (±1.85) |
| Sex | females | females | females | females |
| BMI [kg/m2] | 20.89 (±3.47) | 21.51 (±2.44) | 20.41 (±4.10) | 20.46 (±4.38) |
| HDRS | 20 (13–30) | 4 (1–9) | 12 (8–24) | − |
| CDI-2 | 74.5 (60–79) | 54 (47–68) | 74 (60–79) | 54 (35–79) |
| Time of treatment [weeks] | 7.27 (±1.41) | 7.46 (±1.39) | 7.12 (±1.45) | − |
| BDNF [pg/mL] | 32,890 (±9352) | 23,114 (±15,416) | 25,247 (±15,712) | 30,355 (±7763) |
| proBDNF [pg/mL] | 2068 (±3608) | 856 (±1027) | 3180 (±4470) | 1338 (±2649) |
| BDNF promoter MI [%] * | 0.212 (±0.063) | 0.223 (±0.058) | 0.214 (±0.073) | 0.264 (±0.099) |
| Global DNA methylation [%] | 0.597 (±0.290) | 0.479 (±0.365) | 0.613 (±0.464) | 0.520 (±0.231) |
| Studied Group (t0) vs. Control | Responders (t1) vs. Control | Non-Responders (t1) vs. Control | ||
|---|---|---|---|---|
| BDNF | p-value | 0.263 | 0.131 | 0.716 |
| test power | 95% | 95% | 39% | |
| proBDNF | p-value | 0.452 | 0.696 | 0.134 |
| test power | 20% | 17% | 43% | |
| BDNF promoter MI (exon IV) | p-value | 0.021 * | 0.107 | 0.051 |
| test power | 77% | 44% | 57% | |
| Global DNA methylation | p-value | 0.277 | 0.669 | 0.515 |
| test power | 29% | 10% | 20% |
| Primer Version | Forward Methylated DNA (MF) | Reverse Methylated DNA (MR) | Forward Unmethylated DNA (UF) | Reverse Unmethylated DNA (UR) |
|---|---|---|---|---|
| 1 | AGCGAGAGTAGTTTTTTTCGC | CATATAACAACGCACGTCAAA | GGTAGTGAGAGTAGTTTTTTTTGT | TCATATAACAACACACATCAAAAC |
| 2 | TGTATGGCGGAGGTAATATTC | AAACTACTCTCGCTACCGCT | ATTGTATGGTGGAGGTAATATTT | AAAAACTACTCTCACTACCACT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zwolińska, W.; Bilska, K.; Tarhonska, K.; Reszka, E.; Skibińska, M.; Pytlińska, N.; Słopień, A.; Dmitrzak-Węglarz, M. Biomarkers of Depression among Adolescent Girls: BDNF and Epigenetics. Int. J. Mol. Sci. 2024, 25, 3281. https://doi.org/10.3390/ijms25063281
Zwolińska W, Bilska K, Tarhonska K, Reszka E, Skibińska M, Pytlińska N, Słopień A, Dmitrzak-Węglarz M. Biomarkers of Depression among Adolescent Girls: BDNF and Epigenetics. International Journal of Molecular Sciences. 2024; 25(6):3281. https://doi.org/10.3390/ijms25063281
Chicago/Turabian StyleZwolińska, Weronika, Karolina Bilska, Kateryna Tarhonska, Edyta Reszka, Maria Skibińska, Natalia Pytlińska, Agnieszka Słopień, and Monika Dmitrzak-Węglarz. 2024. "Biomarkers of Depression among Adolescent Girls: BDNF and Epigenetics" International Journal of Molecular Sciences 25, no. 6: 3281. https://doi.org/10.3390/ijms25063281
APA StyleZwolińska, W., Bilska, K., Tarhonska, K., Reszka, E., Skibińska, M., Pytlińska, N., Słopień, A., & Dmitrzak-Węglarz, M. (2024). Biomarkers of Depression among Adolescent Girls: BDNF and Epigenetics. International Journal of Molecular Sciences, 25(6), 3281. https://doi.org/10.3390/ijms25063281