
Oxidative Stress-Mediated Repression of Virulence Gene Transcription and Biofilm Formation as Antibacterial Action of Cinnamomum burmannii Essential Oil on Staphylococcus aureus

 , and
, and
Abstract
1. Introduction
2. Results
2.1. Identification of Volatile Compounds in C. burmannii Leaf Essential Oil (CBLEO)
2.2. Assay of Antibacterial Activity of CBLEO
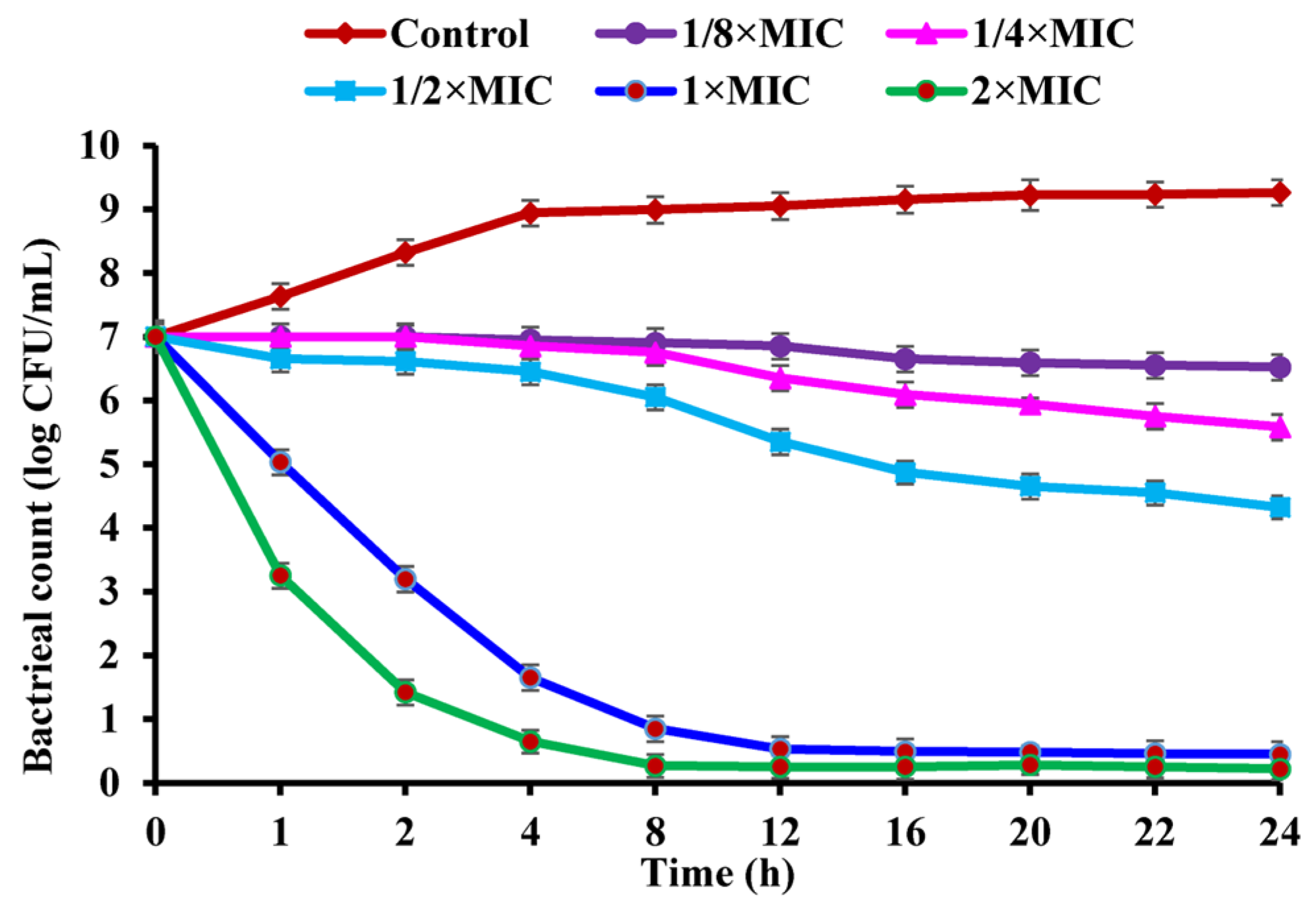
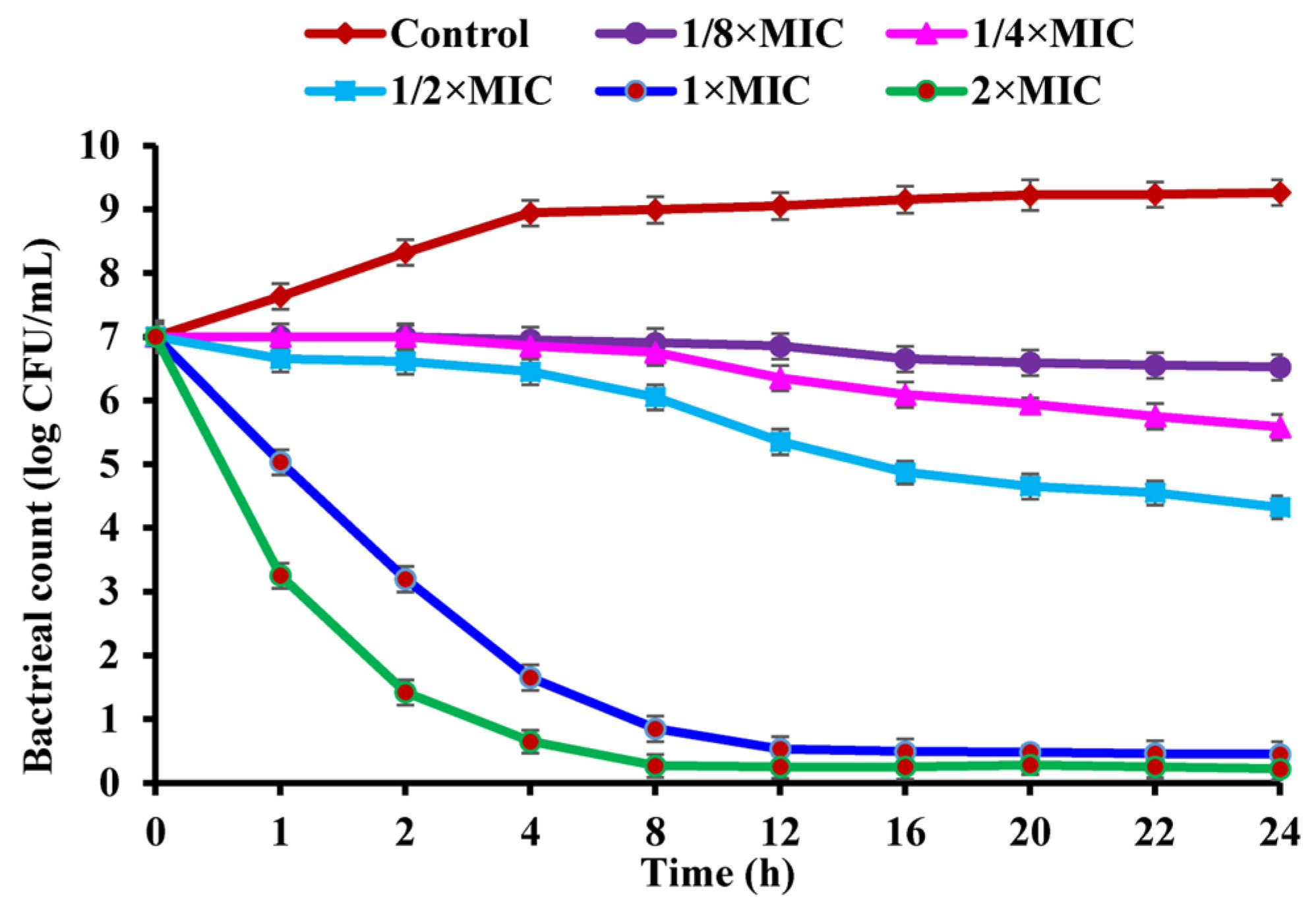
2.3. Effect of CBLEO on Bacterial Growth of S. aureus
2.4. Impact of CBLEO on Cell Structure of S. aureus
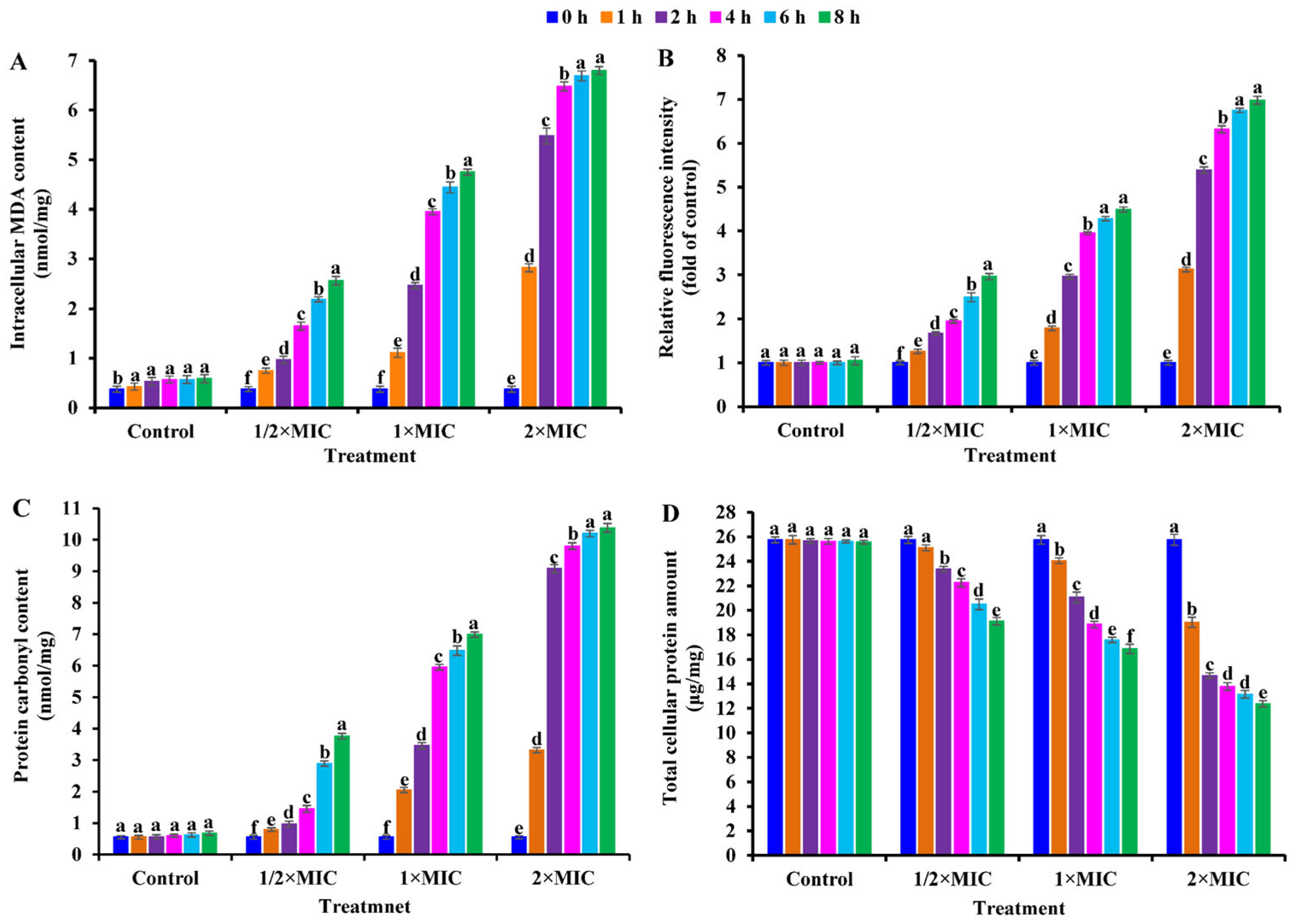
2.5. Effect of CBLEO on Cellular MDA and ROS Generation and Protein Oxidation of S. aureus
2.6. Effect of CBLEO on Cellular Total Protein Concentration of S. aureus
2.7. Effect of CBLEO on Cellular Biofilm Development and Protease Activity of S. aureus
2.8. Effect of CBLEO on Genome DNA of S. aureus
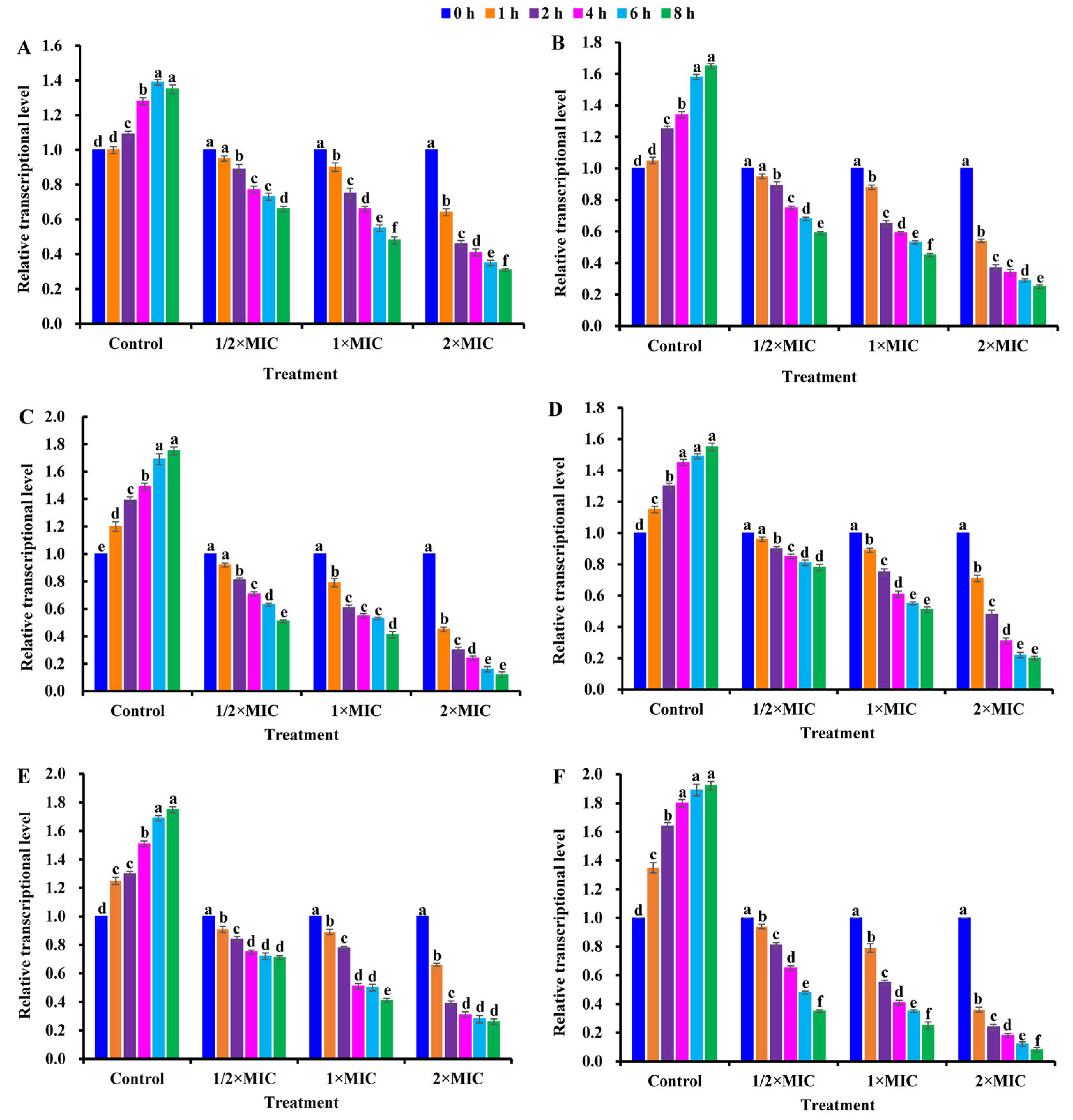
2.9. Effect of CBLEO on Transcript of Virulence-Related Genes and Regulatory Proteins in S. aureus
3. Discussion
3.1. Rich Volatile Profiling with High Borneol Amount and Good Antibacterial Activity of CBLEO
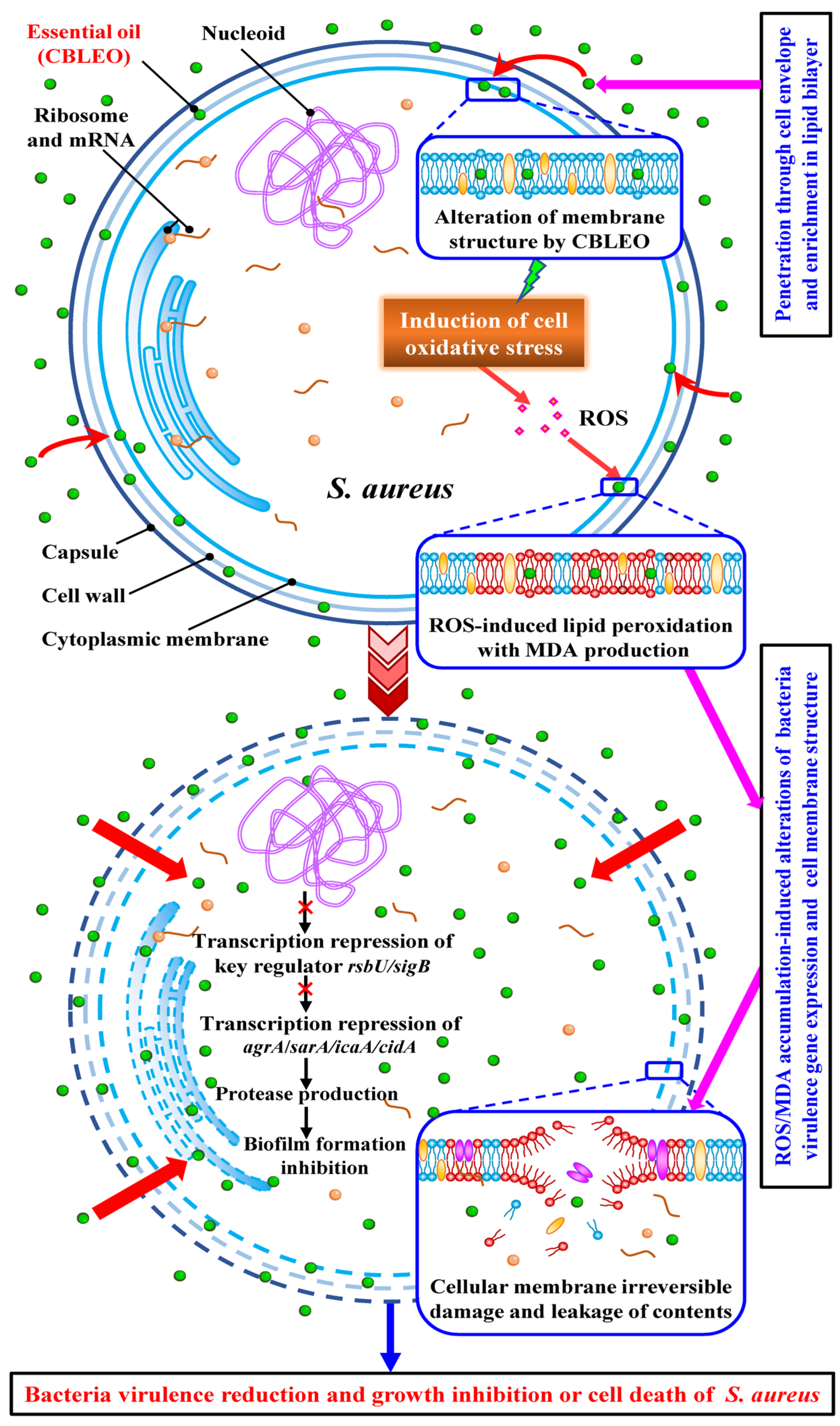
3.2. ROS-Generation-Mediated Oxidative Stress and Cell Membrane Damage Involved in Antibacterial Action of CBLEO
3.3. CBLEO-Induced Transcription Repression of Virulence-Associated Genes in S. aureus as Pivotal Antibacterial Action
4. Materials and Methods
4.1. Plant Materials
4.2. Extraction of Essential Oil and Analysis of Chemical Components of CBLEO
4.3. Bacterial Strains and Culture
4.4. Assessment of Antibacterial Activity of CBLEO
4.4.1. Detection of Diameter of Inhibition Zone (DIZ)
4.4.2. Determination of Minimum Inhibitory (MIC) and Bactericidal Concentration (MBC)
4.5. Analysis of Bacterial Growth Kinetics of S. aureus
4.6. Analysis of Antibacterial Mechanism of CBLEO on S. aureus
4.6.1. Cell Membrane Permeability
4.6.2. Integrity of Cell Membrane
4.6.3. Cell Wall Damage
4.6.4. Scanning Electron Microscope (SEM) Analysis
4.6.5. Analyses of Cellular Protein Oxidation and ROS and MDA Production
4.7. Analysis of Bacterial Total Protein
4.8. Total Protease Assay
4.9. Analysis of Virulence-Associated Gene Expression
4.10. Assay of Anti-biofilm Activity
4.11. Assay of Binding Activity of CBLEO to Bacteria Genome DNA
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gutiérrez-del-Río, I.; Fernández, J.; Lombó, F. Plant nutraceuticals as antimicrobial agents in food preservation: Terpenoids, polyphenols and thiols. Int. J. Antimicrob. Agents 2018, 52, 309–315. [Google Scholar] [CrossRef]
- Erickson, M.C.; Doyle, M.P. The challenges of eliminating or substituting antimicrobial preservatives in foods. Annu Rev. Food Sci. Technol. 2017, 8, 371–390. [Google Scholar] [CrossRef]
- Dahham, S.S.; Tabana, Y.M.; Iqbal, M.A.; Ahamed, M.B.; Ezzat, M.O.; Majid, A.S.; Majid, A.M. The anticancer, antioxidant and antimicrobial properties of the sesquiterpene β-caryophyllene from the essential oil of Aquilaria crassna. Molecules 2015, 20, 11808–11829. [Google Scholar] [CrossRef] [PubMed]
- Akarca, G. Composition and antibacterial effect on food borne pathogens of Hibiscus surrattensis L. calyces essential oil. Ind. Crops Prod. 2019, 137, 285–289. [Google Scholar] [CrossRef]
- Bouyahya, A.; Assemian, I.C.C.; Mouzount, H.; Bourais, I.; Et-Touys, A.; Fellah, H.; Benjouad, A.; Dakka, N.; Bakri, Y. Could volatile compounds from leaves and fruits of Pistacia lentiscus constitute a novel source of anticancer, antioxidant, antiparasitic and antibacterial drugs? Ind. Crops Prod. 2019, 128, 62–69. [Google Scholar] [CrossRef]
- De Almeida, W.S.; de Lima, S.G.; Barreto, H.M.; Andrade, L.M.d.S.; Fonseca, L.; Athayde Sobrinho, C.; Santos, A.R.B.; Muratori, M.C.S. Chemical composition and antimicrobial activity of the essential oil of Lippia lasiocalycina Cham. (Verbenaceae). Ind. Crops Prod. 2018, 125, 236–240. [Google Scholar] [CrossRef]
- Chen, F.; Miao, X.; Lin, Z.; Xiu, Y.; Shi, L.; Zhang, Q.; Liang, D.; Lin, S.; He, B. Disruption of metabolic function and redox homeostasis as antibacterial mechanism of Lindera glauca fruit essential oil against Shigella flexneri. Food Control 2021, 130, 108282. [Google Scholar] [CrossRef]
- Dai, J.; Li, C.; Cui, H.; Lin, L. Unraveling the anti-bacterial mechanism of Litsea cubeba essential oil against E. coli O157:H7 and its application in vegetable juices. Int. J. Food Microbiol. 2021, 338, 108989. [Google Scholar] [CrossRef]
- Pandey, A.K.; Kumar, P.; Singh, P.; Tripathi, N.N.; Bajpai, V.K. Essential oils: Sources of antimicrobials and food preservatives. Front. Microbiol. 2017, 7, 2161. [Google Scholar] [CrossRef]
- Bondi, M.; Lauková, A.; de Niederhausern, S.; Messi, P.; Papadopoulou, C. Natural preservatives to improve food quality and safety. J. Food Qual. 2017, 2017, 1090932. [Google Scholar] [CrossRef]
- Sharma, S.; Barkauskaite, S.; Jaiswal, A.K.; Jaiswal, S. Essential oils as additives in active food packaging. Food Chem. 2021, 343, 128403. [Google Scholar] [CrossRef] [PubMed]
- Ardjoum, N.; Chibani, N.; Shankar, S.; Salmieri, S.; Djidjelli, H.; Lacroix, M. Incorporation of Thymus vulgaris essential oil and ethanolic extract of propolis improved the antibacterial, barrier and mechanical properties of corn starch-based films. Int. J. Biol. Macromol. 2023, 224, 578–583. [Google Scholar] [CrossRef] [PubMed]
- Bajpai, V.K.; Sharma, A.; Baek, K.-H. Antibacterial mode of action of Cudrania tricuspidata fruit essential oil, affecting membrane permeability and surface characteristics of food-borne pathogens. Food Control 2013, 32, 582–590. [Google Scholar] [CrossRef]
- Hu, W.; Li, C.; Dai, J.; Cui, H.; Lin, L. Antibacterial activity and mechanism of Litsea cubeba essential oil against methicillin-resistant Staphylococcus aureus (MRSA). Ind. Crops Prod. 2019, 130, 34–41. [Google Scholar] [CrossRef]
- De Souza Moura, W.; de Souza, S.R.; Campos, F.S.; Sander Rodrigues Cangussu, A.; Macedo Sobrinho Santos, E.; Silva Andrade, B.; Borges Gomes, C.H.; Fernandes Viana, K.; Haddi, K.; Oliveira, E.E.; et al. Antibacterial activity of Siparuna guianensis essential oil mediated by impairment of membrane permeability and replication of pathogenic bacteria. Ind. Crops Prod. 2020, 146, 112142. [Google Scholar] [CrossRef]
- Cui, H.; Zhang, C.; Li, C.; Lin, L. Antibacterial mechanism of oregano essential oil. Ind. Crops Prod. 2019, 139, 111498. [Google Scholar] [CrossRef]
- OuYang, Q.; Tao, N.; Zhang, M. A damaged oxidative phosphorylation mechanism is involved in the antifungal activity of citral against Penicillium digitatum. Front. Microbiol. 2018, 9, 239. [Google Scholar] [CrossRef]
- Mansuri, A.; Lokhande, K.; Kore, S.; Gaikwad, S.; Nawani, N.; Swamy, K.V.; Junnarkar, M.; Pawar, S. Antioxidant, anti-quorum sensing, biofilm inhibitory activities and chemical composition of Patchouli essential oil: In vitro and in silico approach. J. Biomol. Struct. Dyn. 2020, 40, 154–165. [Google Scholar] [CrossRef]
- Wu, Y.; Bai, J.; Zhong, K.; Huang, Y.; Gao, H. A dual antibacterial mechanism involved in membrane disruption and DNA binding of 2R,3R-dihydromyricetin from pine needles of Cedrus deodara against Staphylococcus aureus. Food Chem. 2017, 218, 463–470. [Google Scholar] [CrossRef]
- Wu, S.; Duan, N.; Gu, H.; Hao, L.; Ye, H.; Gong, W.; Wang, Z. A review of the methods for detection of Staphylococcus aureus enterotoxins. Toxins 2016, 8, 176. [Google Scholar] [CrossRef]
- Fisher, E.L.; Otto, M.; Cheung, G.Y.C. Basis of virulence in enterotoxin-mediated Staphylococcal food poisoning. Front. Microbiol. 2018, 9, 436. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zhao, F.; Wang, J.; Zhong, N. Biofilm formation and control strategies of foodborne pathogens: Food safety perspectives. RSC Adv. 2017, 7, 36670–36683. [Google Scholar] [CrossRef]
- Sun, F.; Liang, H.; Kong, X.; Xie, S.; Cho, H.; Deng, X.; Ji, Q.; Zhang, H.; Alvarez, S.; Hicks, L.M.; et al. Quorum-sensing agr mediates bacterial oxidation response via an intramolecular disulfide redox switch in the response regulator AgrA. Proc. Natl. Acad. Sci. USA 2012, 109, 9095–9100. [Google Scholar] [CrossRef]
- Cheung, A.L.; Nishina, K.A.; Trotonda, M.P.; Tamber, S. The SarA protein family of Staphylococcus aureus. Int. J. Biochem. Cell Biol. 2008, 40, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Jenul, C.; Horswill, A.R. Regulation of Staphylococcus aureus virulence. Microbiol. Spectr. 2019, 7, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Li, D.; Huang, X.; Yang, H.; Qiu, Z.; Zou, L.; Liang, Q.; Shi, Y.; Wu, Y.; Wu, S.; et al. Study on antibacterial and quorum-sensing inhibition activities of Cinnamomum camphora leaf essential oil. Molecules 2019, 24, 3792. [Google Scholar] [CrossRef] [PubMed]
- Manoharan, R.K.; Lee, J.-H.; Kim, Y.-G.; Kim, S.-I.; Lee, J. Inhibitory effects of the essential oils α-longipinene and linalool on biofilm formation and hyphal growth of Candida albicans. Biofouling 2017, 33, 143–155. [Google Scholar] [CrossRef]
- Joshi, J.R.; Khazanov, N.; Senderowitz, H.; Burdman, S.; Lipsky, A.; Yedidia, I. Plant phenolic volatiles inhibit quorum sensing in pectobacteria and reduce their virulence by potential binding to ExpI and ExpR proteins. Sci. Rep. 2016, 6, 38126. [Google Scholar] [CrossRef]
- Alves, S.; Duarte, A.; Sousa, S.; Domingues, F.C. Study of the major essential oil compounds of Coriandrum sativum against Acinetobacter baumannii and the effect of linalool on adhesion, biofilms and quorum sensing. Biofouling 2016, 32, 155–165. [Google Scholar] [CrossRef]
- Souza, C.M.C.; Pereira Junior, S.A.; Moraes, T.d.S.; Damasceno, J.L.; Amorim Mendes, S.; Dias, H.J.; Stefani, R.; Tavares, D.C.; Martins, C.H.G.; Crotti, A.E.M.; et al. Antifungal activity of plant-derived essential oils on Candida tropicalis planktonic and biofilms cells. Med. Mycol. 2016, 54, 515–523. [Google Scholar] [CrossRef]
- Lee, K.; Lee, J.-H.; Kim, S.-I.; Cho, M.H.; Lee, J. Anti-biofilm, anti-hemolysis, and anti-virulence activities of black pepper, cananga, myrrh oils, and nerolidol against Staphylococcus aureus. Appl. Microbiol. Biotechnol. 2014, 98, 9447–9457. [Google Scholar] [CrossRef]
- Ribeiro-Santos, R.; Andrade, M.; Madella, D.; Martinazzo, A.P.; de Aquino Garcia Moura, L.; de Melo, N.R.; Sanches-Silva, A. Revisiting an ancient spice with medicinal purposes: Cinnamon. Trends Food Sci. Technol. 2017, 62, 154–169. [Google Scholar] [CrossRef]
- Vasconcelos, N.G.; Croda, J.; Simionatto, S. Antibacterial mechanisms of cinnamon and its constituents: A review. Microb. Pathog. 2018, 120, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Al-Dhubiab, B.E. Pharmaceutical applications and phytochemical profile of Cinnamomum burmannii. Pharmacogn. Rev. 2012, 6, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Shan, B.; Cai, Y.; Brooks, J.; Corke, H. Antibacterial properties and major bioactive components of cinnamon stick (Cinnamomum burmannii): Activity against foodborne pathogenic bacteria. J. Agric. Food Chem. 2007, 55, 5484–5490. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Yang, L.; Zou, Y.; Luo, S.; Wang, X.; Liang, Y.; Du, Y.; Feng, R.; Wei, Q. Antibacterial activity and mechanism of three isomeric terpineols of Cinnamomum longepaniculatum leaf oil. Folia Microbiol. 2021, 66, 59–67. [Google Scholar] [CrossRef]
- Muhammad, D.R.A.; Lemarcq, V.; Alderweireldt, E.; Vanoverberghe, P.; Praseptiangga, D.; Juvinal, J.G.; Dewettinck, K. Antioxidant activity and quality attributes of white chocolate incorporated with Cinnamomum burmannii Blume essential oil. J Food Sci. Technol. 2020, 57, 1731–1739. [Google Scholar] [CrossRef]
- Muhammad, D.R.A.; Tuenter, E.; Patria, G.D.; Foubert, K.; Pieters, L.; Dewettinck, K. Phytochemical composition and antioxidant activity of Cinnamomum burmannii Blume extracts and their potential application in white chocolate. Food Chem. 2021, 340, 127983. [Google Scholar] [CrossRef]
- Muhammad, D.R.A.; Saputro, A.D.; Rottiers, H.; Van de Walle, D.; Dewettinck, K. Physicochemical properties and antioxidant activities of chocolates enriched with engineered cinnamon nanoparticles. Eur. Food Res. Technol. 2018, 244, 1185–1202. [Google Scholar] [CrossRef]
- Wu, G.; Lian, H.; Zhang, C.; Li, B.; Chen, J.; He, B.; Zhang, Q.; Wang, Y. Content variation and evaluation of essential oil and its main chemical components of Cinnamomum burmannii in Guangdong province. For. Environ. Sci. 2020, 36, 88–95. [Google Scholar] [CrossRef]
- Xie, P.; He, B.; Wang, Y.; Zhang, Q. Standardization of exploration and utilization of d-borneol type Cinnamomum camphora. For. Environ. Sci. 2019, 35, 94–100. [Google Scholar] [CrossRef]
- Zielinska, A.K.; Beenken, K.E.; Mrak, L.N.; Spencer, H.J.; Post, G.R.; Skinner, R.A.; Tackett, A.J.; Horswill, A.R.; Smeltzer, M.S. sarA-mediated repression of protease production plays a key role in the pathogenesis of Staphylococcus aureus USA300 isolates. Mol. Microbiol. 2012, 86, 1183–1196. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Ren, X.; Liu, Y.; Xie, Y.; Guo, Y.; Cheng, Y.; Qian, H.; Yao, W. Extraction of Cinnamomum camphora chvar. Borneol essential oil using neutral cellulase assisted-steam distillation: Optimization of extraction, and analysis of chemical constituents. Ind. Crops Prod. 2019, 141, 111794. [Google Scholar] [CrossRef]
- Bansod, S.; Chilvery, S.; Saifi, M.A.; Das, T.J.; Tag, H.; Godugu, C. Borneol protects against cerulein-induced oxidative stress and inflammation in acute pancreatitis mice model. Environ. Toxicol. 2021, 36, 530–539. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Wen, J.; Wang, Z.; Wang, J. Multiple regulation and targeting effects of borneol in the neurovascular unit in neurodegenerative diseases. Basic Clin. Pharmacol. Toxicol. 2022, 130, 5–19. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Long, Y.; Yu, S.; Zhang, D.; Yang, Q.; Ci, Z.; Cui, M.; Zhang, Y.; Wan, J.; Li, D.; et al. Borneol in cardio-cerebrovascular diseases: Pharmacological actions, mechanisms, and therapeutics. Pharmacol. Res. 2021, 169, 105627. [Google Scholar] [CrossRef]
- Chebet, J.J.; Ehiri, J.E.; McClelland, D.J.; Taren, D.; Hakim, I.A. Effect of d-limonene and its derivatives on breast cancer in human trials: A scoping review and narrative synthesis. BMC Cancer 2021, 21, 902. [Google Scholar] [CrossRef] [PubMed]
- Mosbah, H.; Louati, H.; Boujbiha, M.A.; Chahdoura, H.; Snoussi, M.; Flamini, G.; Ascrizzi, R.; Bouslema, A.; Achour, L.; Selmi, B. Phytochemical characterization, antioxidant, antimicrobial and pharmacological activities of Feijoa sellowiana leaves growing in Tunisia. Ind. Crops Prod. 2018, 112, 521–531. [Google Scholar] [CrossRef]
- Salehi, B.; Upadhyay, S.; Erdogan Orhan, I.; Kumar Jugran, A.; Jayaweera, S.L.D.; Dias, D.A.; Sharopov, F.; Taheri, Y.; Martins, N.; Baghalpour, N.; et al. Therapeutic potential of α-and β-Pinene: A miracle gift of nature. Biomolecules 2019, 9, 738. [Google Scholar] [CrossRef]
- Nafis, A.; Kasrati, A.; Jamali, C.A.; Mezrioui, N.; Setzer, W.; Abbad, A.; Hassani, L. Antioxidant activity and evidence for synergism of Cannabis sativa (L.) essential oil with antimicrobial standards. Ind. Crops Prod. 2019, 137, 396–400. [Google Scholar] [CrossRef]
- Huang, H.; Lian, H.; He, B.; Wang, Y.; Chen, G.; Liang, D.; Chen, X.; Luo, W.; Lin, S.; Fengqing, L. Study on the dynamic change of essential oil content and chemical constituents in the leaves of Cinnamomum burmannii chvar. borneol. For. Environ. Sci. 2019, 35, 22–26. [Google Scholar] [CrossRef]
- Wang, R.; Wang, R.; Yang, B. Extraction of essential oils from five cinnamon leaves and identification of their volatile compound compositions. Innov. Food Sci. Emerg. 2009, 10, 289–292. [Google Scholar] [CrossRef]
- Zhou, F.; Zhao, M.; Su, G.; Cui, C.; Sun, W. Gelation of salted myofibrillar protein under malondialdehyde-induced oxidative stress. Food Hydrocoll. 2014, 40, 153–162. [Google Scholar] [CrossRef]
- Hsu, K.-C.; Hsu, P.-F.; Chen, Y.-C.; Lin, H.-C.; Hung, C.-C.; Chen, P.-C.; Huang, Y.-L. Oxidative stress during bacterial growth characterized through microdialysis sampling coupled with HPLC/fluorescence detection of malondialdehyde. J. Chromatogr. B 2016, 1019, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.A.; Moon, S.-H.; Lee, J.-Y.; Kim, K.-T.; Park, Y.-S.; Paik, H.-D. Antibacterial activity of a novel flavonoid, 7-O-butyl naringenin, against methicillin-resistant Staphylococcus aureus (MRSA). Food Sci. Biotechnol. 2013, 22, 1725–1728. [Google Scholar] [CrossRef]
- Chen, F.; Wang, H.; Lin, Z.; Hu, J.; Wu, Y.; Shi, L.; Wang, J.; Xiu, Y.; Lin, S. Enzymatic and non-enzymatic bioactive compounds, and antioxidant and antimicrobial activities of the extract from one selected wild berry (Rubus coreanus) as novel natural agent for food preservation. LWT-Food Sci. Technol. 2022, 171, 114133. [Google Scholar] [CrossRef]
- Fathima, A.; Rao, J.R. Selective toxicity of Catechin-a natural flavonoid towards bacteria. Appl. Microbiol. Biotechnol. 2016, 100, 6395–6402. [Google Scholar] [CrossRef]
- Yan, L.; Li, M.; Cao, Y.; Gao, P.; Cao, Y.; Wang, Y.; Jiang, Y. The alternative oxidase of Candida albicans causes reduced fluconazole susceptibility. J. Antimicrob. Chemother. 2009, 64, 764–773. [Google Scholar] [CrossRef]
- Miura, T.; Muraoka, S.; Fujimoto, Y. Lipid peroxidation induced by indomethacin with horseradish peroxidase and hydrogen peroxide: Involvement of indomethacin radicals. Biochem. Pharmacol. 2002, 63, 2069–2074. [Google Scholar] [CrossRef]
- Du, W.; Sun, C.; Liang, Z.; Han, Y.; Yu, J. Antibacterial activity of hypocrellin A against Staphylococcus aureus. World J. Microbiol. Biotechnol. 2012, 28, 3151–3157. [Google Scholar] [CrossRef]
- Ayer, A.; Gourlay, C.W.; Dawes, I.W. Cellular redox homeostasis, reactive oxygen species and replicative ageing in Saccharomyces cerevisiae. FEMS Yeast Res. 2014, 14, 60–72. [Google Scholar] [CrossRef]
- Lu, J.; Holmgren, A. The thioredoxin antioxidant system. Free Radic. Biol. Med. 2014, 66, 75–87. [Google Scholar] [CrossRef]
- Karpenko, I.L.; Valuev-Elliston, V.T.; Ivanova, O.N.; Smirnova, O.A.; Ivanov, A.V. Peroxiredoxins—The underrated actors during virus-induced oxidative stress. Antioxidants 2021, 10, 977. [Google Scholar] [CrossRef] [PubMed]
- Chai, Y.-C.; Mieyal, J.J. Glutathione and glutaredoxin-Key players in cellular Redox homeostasis and signaling. Antioxidants 2023, 12, 1553. [Google Scholar] [CrossRef] [PubMed]
- Le, D.T.; Lee, B.C.; Marino, S.M.; Zhang, Y.; Fomenko, D.E.; Kaya, A.; Hacioglu, E.; Kwak, G.-H.; Koc, A.; Kim, H.-Y.; et al. Functional analysis of free methionine-R-sulfoxide reductase from Saccharomyces cerevisiae. J. Biol. Chem. 2009, 284, 4354–4364. [Google Scholar] [CrossRef]
- Grant, C.M. Role of the glutathione/glutaredoxin and thioredoxin systems in yeast growth and response to stress conditions. Mol. Microbiol. 2001, 39, 533–541. [Google Scholar] [CrossRef]
- Inoue, Y.; Matsuda, T.; Sugiyama, K.-I.; Izawa, S.; Kimura, A. Genetic analysis of glutathione peroxidase in oxidative stress response of Saccharomyces cerevisiae. J. Biol. Chem. 1999, 274, 27002–27009. [Google Scholar] [CrossRef]
- Rodríguez-Manzaneque, M.T.; Ros, J.; Cabiscol, E.; Sorribas, A.; Herrero, E. Grx5 glutaredoxin plays a central role in protection against protein oxidative damage in Saccharomyces cerevisiae. Mol. Cell. Biol. 1999, 19, 8180–8190. [Google Scholar] [CrossRef]
- King, K.; Horenstein, J.; Caparon, M. Aerotolerance and peroxide resistance in peroxidase and PerR mutants of Streptococcus pyogenes. J. Bacteriol. 2000, 182, 5290–5299. [Google Scholar] [CrossRef] [PubMed]
- Uziel, O.; Borovok, I.; Schreiber, R.; Cohen, G.; Aharonowitz, Y. Transcriptional regulation of the Staphylococcus aureus thioredoxin and thioredoxin reductase genes in response to oxygen and disulfide stress. J. Bacteriol. 2004, 186, 326–334. [Google Scholar] [CrossRef]
- Qian, J.; Wang, C.; Zhuang, H.; Zhang, J.; Yan, W. Oxidative stress responses of pathogen bacteria in poultry to plasma-activated lactic acid solutions. Food Control 2020, 118, 107355. [Google Scholar] [CrossRef]
- Thoendel, M.; Kavanaugh, J.S.; Flack, C.E.; Horswill, A.R. Peptide signaling in the staphylococci. Chem. Rev. 2011, 111, 117–151. [Google Scholar] [CrossRef] [PubMed]
- Le, K.Y.; Otto, M. Quorum-sensing regulation in staphylococci—An overview. Front. Microbiol. 2015, 6, 1174. [Google Scholar] [CrossRef] [PubMed]
- Queck, S.Y.; Jameson-Lee, M.; Villaruz, A.E.; Bach, T.-H.L.; Khan, B.A.; Sturdevant, D.E.; Ricklefs, S.M.; Li, M.; Otto, M. RNAIII-independent target gene control by the agr quorum-sensing system: Insight into the evolution of virulence regulation in Staphylococcus aureus. Mol. Cell. 2008, 32, 150–158. [Google Scholar] [CrossRef]
- Bischoff, M.; Dunman, P.; Kormanec, J.; Macapagal, D.; Murphy, E.; Mounts, W.; Berger-Bächi, B.; Projan, S. Microarray-based analysis of the Staphylococcus aureus σB regulon. J. Bacteriol. 2004, 186, 4085–4099. [Google Scholar] [CrossRef] [PubMed]
- Guldimann, C.; Boor, K.J.; Wiedmann, M.; Guariglia-Oropeza, V. Resilience in the face of uncertainty: Sigma factor B fine-tunes gene expression to support homeostasis in gram-positive bacteria. Appl. Environ. Microbiol. 2016, 82, 4456–4469. [Google Scholar] [CrossRef]
- Atwood, D.N.; Loughran, A.J.; Courtney, A.P.; Anthony, A.C.; Meeker, D.G.; Spencer, H.J.; Gupta, R.K.; Lee, C.Y.; Beenken, K.E.; Smeltzer, M.S. Comparative impact of diverse regulatory loci on Staphylococcus aureus biofilm formation. MicrobiologyOpen 2015, 4, 436–451. [Google Scholar] [CrossRef]
- Harapanahalli, A.K.; Chen, Y.; Li, J.; Busscher, H.J.; van der Mei, H.C. Influence of adhesion force on icaA and cidA gene expression and production of matrix components in Staphylococcus aureus biofilms. Appl. Environ. Microbiol. 2015, 81, 3369–3378. [Google Scholar] [CrossRef]
- Savage, V.J.; Chopra, I.; O’Neill, A.J. Population diversification in Staphylococcus aureus biofilms may promote dissemination and persistence. PLoS ONE 2013, 8, e62513. [Google Scholar] [CrossRef]
- Lauderdale, K.J.; Boles, B.R.; Cheung, A.L.; Horswill, A.R. Interconnections between sigma B, agr, and proteolytic activity in Staphylococcus aureus biofilm maturation. Infect. Immun. 2009, 77, 1623–1635. [Google Scholar] [CrossRef]
- Rice, K.C.; Mann, E.E.; Endres, J.L.; Weiss, E.C.; Cassat, J.E.; Smeltzer, M.S.; Bayles, K.W. The cidA murein hydrolase regulator contributes to DNA release and biofilm development in Staphylococcus aureus. Proc. Natl. Acad. Sci. USA 2007, 104, 8113–8118. [Google Scholar] [CrossRef]
- Tsang, L.H.; Cassat, J.E.; Shaw, L.N.; Beenken, K.E.; Smeltzer, M.S. Factors contributing to the biofilm-deficient phenotype of Staphylococcus aureus sarA mutants. PLoS ONE 2008, 3, e3361. [Google Scholar] [CrossRef]
- Beenken, K.E.; Blevins, J.S.; Smeltzer, M.S. Mutation of sarA in Staphylococcus aureus limits biofilm formation. Infect. Immun. 2003, 71, 4206–4211. [Google Scholar] [CrossRef]
- Loughran, A.J.; Atwood, D.N.; Anthony, A.C.; Harik, N.S.; Spencer, H.J.; Beenken, K.E.; Smeltzer, M.S. Impact of individual extracellular proteases on Staphylococcus aureus biofilm formation in diverse clinical isolates and their isogenic sarA mutants. MicrobiologyOpen 2014, 3, 897–909. [Google Scholar] [CrossRef]
- Mootz, J.M.; Malone, C.L.; Shaw, L.N.; Horswill, A.R. Staphopains modulate Staphylococcus aureus biofilm integrity. Infect. Immun. 2013, 81, 3227–3238. [Google Scholar] [CrossRef]
- Palma, M.; Cheung, A.L. σB activity in Staphylococcus aureus is controlled by RsbU and an additional factor (s) during bacterial growth. Infect. Immun. 2001, 69, 7858–7865. [Google Scholar] [CrossRef]
- Knobloch Johannes, K.M.; Jäger, S.; Horstkotte Matthias, A.; Rohde, H.; Mack, D. RsbU-dependent regulation of Staphylococcus epidermidis biofilm formation is mediated via the alternative sigma factor σB by repression of the negative regulator gene icaR. Infect. Immun. 2004, 72, 3838–3848. [Google Scholar] [CrossRef] [PubMed]
- Conlon, K.M.; Humphreys, H.; O’Gara, J.P. Inactivations of rsbU and sarA by IS256 represent novel mechanisms of biofilm phenotypic variation in Staphylococcus epidermidis. J. Bacteriol. 2004, 186, 6208–6219. [Google Scholar] [CrossRef] [PubMed]
- Kong, C.; Chee, C.-F.; Richter, K.; Thomas, N.; Abd Rahman, N.; Nathan, S. Suppression of Staphylococcus aureus biofilm formation and virulence by a benzimidazole derivative, UM-C162. Sci. Rep. 2018, 8, 2758. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, L.; Kayama, S.; Sasaki, M.; Kato, F.; Hisatsune, J.; Tsuruda, K.; Koizumi, K.; Tatsukawa, N.; Yu, L.; Takeda, K. Inhibitory effects of antibiofilm compound 1 against Staphylococcus aureus biofilms. Microbiol. Immunol. 2016, 60, 148–159. [Google Scholar] [CrossRef]
- Zhu, B.; Hou, X.; Niu, J.; Li, P.; Fang, C.; Qiu, L.; Ha, D.; Zhang, Z.; Sun, J.; Li, Y. Volatile constituents from the fruits of Lindera glauca (Sieb. et Zucc.) with different maturities. J. Essent. Oil-Bear. Plants 2016, 19, 926–935. [Google Scholar] [CrossRef]
- Zhao, M.; Bai, J.; Bu, X.; Tang, Y.; Han, W.; Li, D.; Wang, L.; Yang, Y.; Xu, Y. Microwave-assisted aqueous two-phase extraction of phenolic compounds from Ribes nigrum L. and its antibacterial effect on foodborne pathogens. Food Control 2021, 119, 107449. [Google Scholar] [CrossRef]
- Pan, Y.; Zheng, L.-b.; Mao, Y.; Wang, J.; Lin, L.-s.; Su, Y.-q.; Li, Y. The antibacterial activity and mechanism analysis of piscidin 5 like from Larimichthys crocea. Dev. Comp. Immunol. 2019, 92, 43–49. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Compound | Formula | RI A | Percentage (%) B |
|---|---|---|---|---|
| Monoterpene hydrocarbons (14) | α-Thujene | C10H16 | 902 | 0.56 ± 0.03 e |
| α-Pinene | C10H16 | 948 | 3.96 ± 0.17 bc | |
| (−)-Camphene | C10H16 | 943 | 2.03 ± 0.12 c | |
| Sabinene | C10H16 | 897 | 2.53 ± 0.11 c | |
| β-Pinene | C10H16 | 943 | 2.38 ± 0.10 c | |
| β-Myrcene | C10H16 | 958 | 2.41 ± 0.10 c | |
| α-Phellandrene | C10H16 | 969 | 2.67 ± 0.12 c | |
| 3-Carene | C10H16 | 948 | 0.32 ± 0.02 e | |
| 4-Carene | C10H16 | 919 | 0.19 ± 0.01 f | |
| β-Cymene | C10H14 | 1042 | 3.96 ± 0.19 bc | |
| D-Limonene | C10H16 | 1018 | 7.44 ± 0.25 b | |
| β-cis-Ocimene | C10H16 | 976 | 0.43 ± 0.02 e | |
| γ-Terpinene | C10H16 | 998 | 0.50 ± 0.03 e | |
| α-Terpinolen | C10H16 | 1052 | 0.96 ± 0.06 e | |
| Monoterpene alcohol (6) | Eucalyptol | C10H18O | 1059 | 9.22 ± 0.29 b |
| Linalool | C10H18O | 1082 | 0.63 ± 0.03 e | |
| Borneol | C10H18O | 1138 | 28.40 ± 0.62 a | |
| Terpinen-4-ol | C10H18O | 1137 | 1.50 ± 0.09 cd | |
| α-Terpineol | C10H18O | 1143 | 3.15 ± 0.18 bc | |
| Guaniol | C10H18O | 1228 | 0.23 ± 0.01 f | |
| Monoterpene ketone (1) | Camphor | C10H16O | 1121 | 2.14 ± 0.10 bc |
| Monoterpene aldehyde (1) | α-Citral | C10H16O | 1174 | 0.19 ± 0.01 f |
| Monoterpene ester (2) | Bornyl acetate | C12H20O2 | 1277 | 9.33 ± 0.24 b |
| Geranyl acetate | C12H20O2 | 1352 | 0.25 ± 0.01 f | |
| Sesquiterpene hydrocarbons (5) | β-Caryophyllene | C15H24 | 1494 | 3.71 ± 0.19 bc |
| α-Caryophyllene | C15H24 | 1579 | 0.99 ± 0.05 e | |
| Germacrene D | C15H24 | 1515 | 0.58 ± 0.03 e | |
| α-Guaiene | C15H24 | 1469 | 0.19 ± 0.01 f | |
| Germacrene B | C15H24 | 1603 | 1.74 ± 0.10 cd | |
| Sesquiterpene alcohol (6) | Elemol | C15H26O | 1522 | 0.42 ± 0.02 e |
| trans-Nerolidol | C15H26O | 1564 | 1.36 ± 0.07 cd | |
| Spathulenol | C15H24O | 1536 | 1.72 ± 0.08 cd | |
| Guaiol | C15H26O | 1614 | 1.28 ± 0.08 cd | |
| (−)-Spathulenol | C15H24O | 1536 | 0.51 ± 0.03 e | |
| Bulnesol | C15H26O | 1614 | 0.44 ± 0.02 e | |
| Sesquiterpene ester (1) | Caryophyllene oxide | C15H24O | 1507 | 1.00 ± 0.08 cd |
| Others (1) | Cinnamyl acetate | C11H12O2 | 1367 | 0.24 ± 0.01 f |
| Total | 99.56 ± 0.21 |
| Microorganisms | CBLEO | Antibiotic A | ||||
|---|---|---|---|---|---|---|
| DIZ B (mm) | MIC B (μg/mL) | MBC B (μg/mL) | DIZ B (mm) | MIC B (μg/mL) | MBC B (μg/mL) | |
| Gram-positive bacteria | ||||||
| Bacillus subtilis | 21.31 ± 0.65 b | 2.0 | 4.0 | 40.70 ± 0.71 a | 0.125 | 0.25 |
| Listeria monocytogenes | 13.33 ± 0.45 bc | 4.0 | 8.0 | 35.31 ± 0.56 b | 0.125 | 0.25 |
| Staphylococcus aureus | 28.72 ± 0.72 a | 1.0 | 2.0 | 30.05 ± 0.56 b | 0.5 | 1.0 |
| Gram-negative bacteria | ||||||
| Escherichia coli | 9.71 ± 0.61 bc | 8.0 | 16.0 | 37.52 ± 0.61 a | 0.25 | 0.5 |
| Pseudomonas aeruginosa | 7.51 ± 0.48 bc | 16.0 | 32.0 | 37.31 ± 0.57 a | 0.25 | 0.5 |
| Enterobacter aerogenes | 10.21 ± 0.51 bc | 8.0 | 16.0 | 33.71 ± 0.61 b | 0.25 | 0.5 |
| Salmonella | 9.02 ± 0.43 bc | 8.0 | 16.0 | 39.51 ± 0.45 a | 0.125 | 0.25 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, L.; Lin, W.; Cai, Y.; Chen, F.; Zhang, Q.; Liang, D.; Xiu, Y.; Lin, S.; He, B. Oxidative Stress-Mediated Repression of Virulence Gene Transcription and Biofilm Formation as Antibacterial Action of Cinnamomum burmannii Essential Oil on Staphylococcus aureus. Int. J. Mol. Sci. 2024, 25, 3078. https://doi.org/10.3390/ijms25053078
Shi L, Lin W, Cai Y, Chen F, Zhang Q, Liang D, Xiu Y, Lin S, He B. Oxidative Stress-Mediated Repression of Virulence Gene Transcription and Biofilm Formation as Antibacterial Action of Cinnamomum burmannii Essential Oil on Staphylococcus aureus. International Journal of Molecular Sciences. 2024; 25(5):3078. https://doi.org/10.3390/ijms25053078
Chicago/Turabian StyleShi, Lingling, Wei Lin, Yanling Cai, Feng Chen, Qian Zhang, Dongcheng Liang, Yu Xiu, Shanzhi Lin, and Boxiang He. 2024. "Oxidative Stress-Mediated Repression of Virulence Gene Transcription and Biofilm Formation as Antibacterial Action of Cinnamomum burmannii Essential Oil on Staphylococcus aureus" International Journal of Molecular Sciences 25, no. 5: 3078. https://doi.org/10.3390/ijms25053078
APA StyleShi, L., Lin, W., Cai, Y., Chen, F., Zhang, Q., Liang, D., Xiu, Y., Lin, S., & He, B. (2024). Oxidative Stress-Mediated Repression of Virulence Gene Transcription and Biofilm Formation as Antibacterial Action of Cinnamomum burmannii Essential Oil on Staphylococcus aureus. International Journal of Molecular Sciences, 25(5), 3078. https://doi.org/10.3390/ijms25053078