
ARTP/NTG Compound Mutagenesis Improved the Spinosad Production and the Insecticidal Virulence of Saccharopolyspora Spinosa
Abstract

1. Introduction
2. Results
2.1. ARTP/NTG Compound Mutagenesis Improved the Titer of Spinosad in S. spinosa
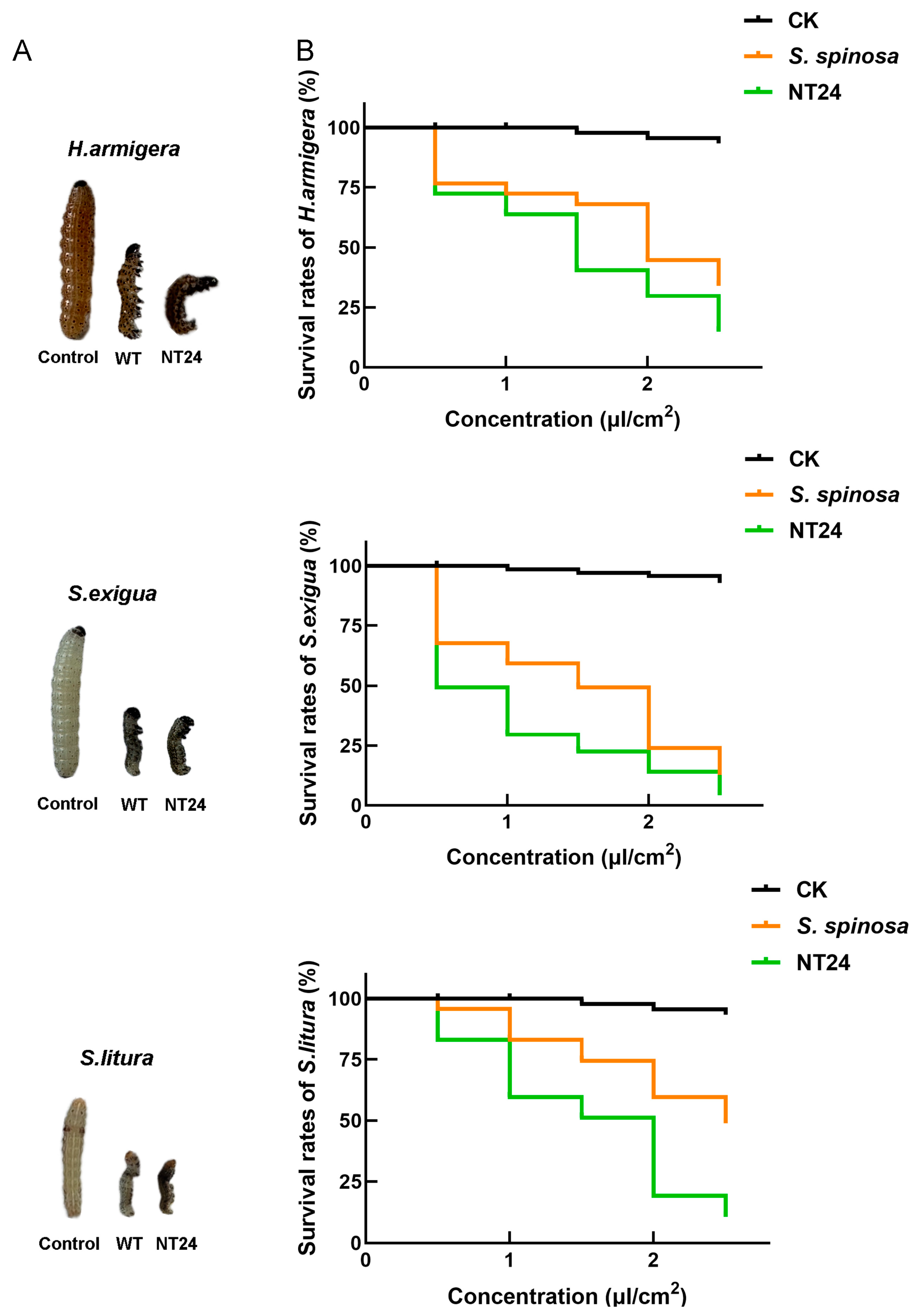
2.2. Biological Activity Assay of NT24
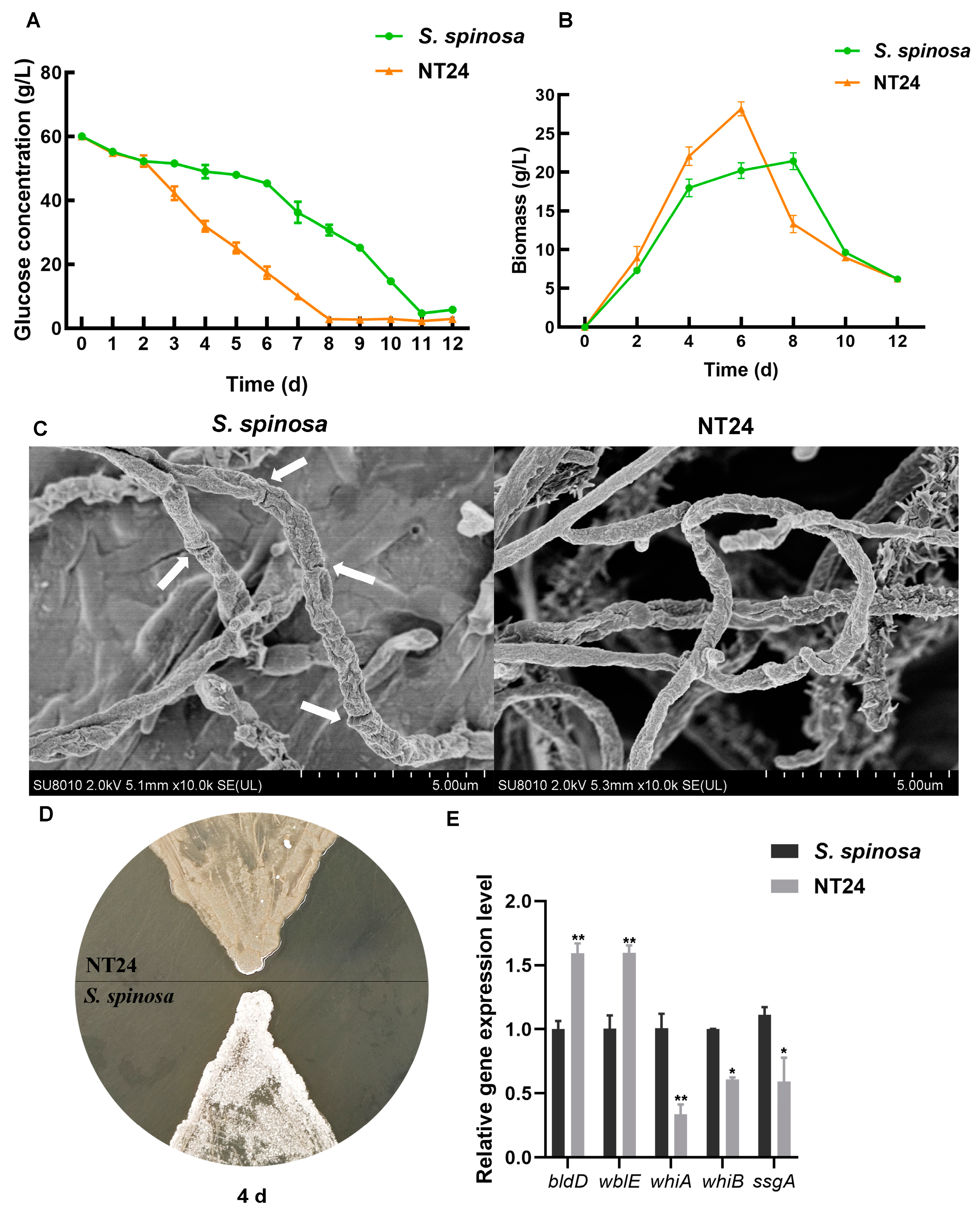
2.3. Phenotypic Differences Between WT and NT24
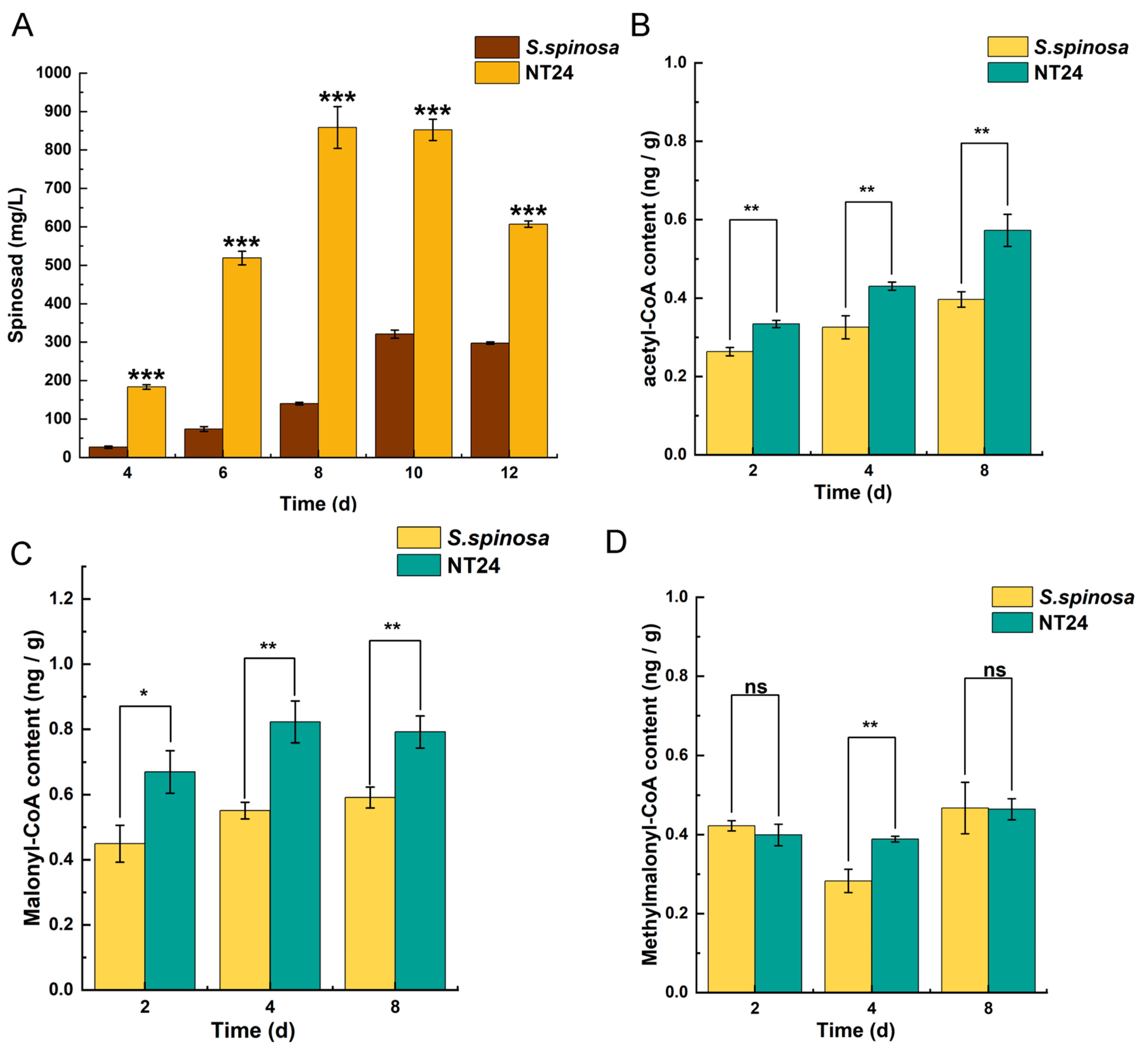
2.4. Differences of the Accumulation in Spinosad and Short-Chain Acyl-coA
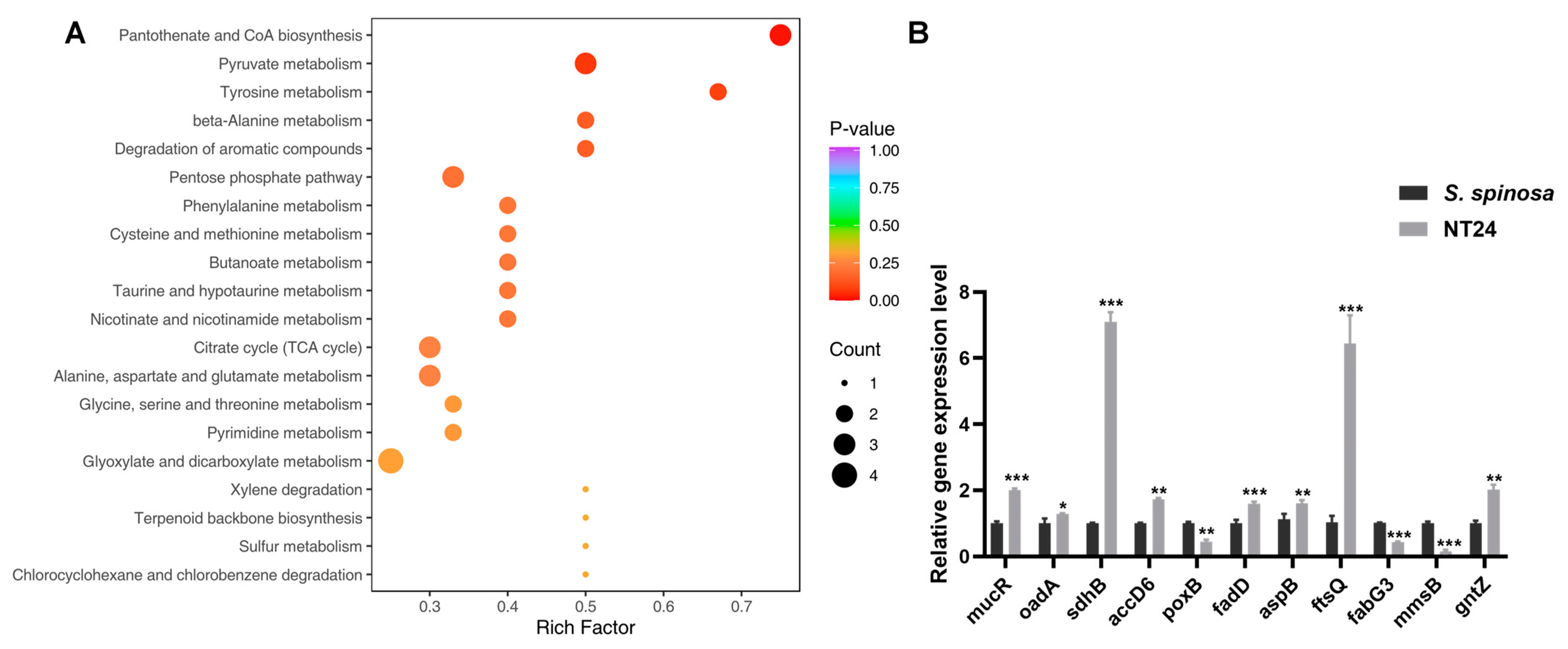
2.5. Analyses of Resequencing and Metabolomics
3. Discussion
4. Materials and Methods
4.1. Strains and Culture Conditions
4.2. Semi-Lethal Concentration Assay of WT and Mutant Strain NT24
4.3. Detection and Identification of Spinosad
4.4. ARTP/NTG Mutagenesis
4.5. Growth Curve and Glucose Consumption Determination
4.6. Morphological Observation
4.7. Determination of Short-Chain Acyl-CoA Precursors
4.8. Genome Sequencing and Assembly
4.9. Total RNA Extraction and RT-qPCR Analysis
4.10. Stability Analysis
4.11. Statistic Method
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Al-Fadhli, A.A.; Threadgill, M.D.; Mohammed, F.; Sibley, P.; Al-Ariqi, W.; Parveen, I. Macrolides from rare actinomycetes: Structures and bioactivities. Int. J. Antimicrob. Agents 2022, 59, 106523. [Google Scholar] [CrossRef] [PubMed]
- Ding, T.; Yang, L.-J.; Zhang, W.-D.; Shen, Y.-H. The secondary metabolites of rare actinomycetes: Chemistry and bioactivity. RSC Adv. 2019, 9, 21964–21988. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, K.; Gupta, R.K. Rare actinomycetes: A potential storehouse for novel antibiotics. Crit. Rev. Biotechnol. 2011, 32, 108–132. [Google Scholar] [CrossRef] [PubMed]
- Guojun, Y.; Yuping, H.; Yan, J.; Kaichun, L.; Haiyang, X. A New Medium for Improving Spinosad Production by Saccharopolyspora spinosa. Jundishapur J. Microbiol. 2016, 9, e16765. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef]
- Salgado, V.L.; Sparks, T.C.; Gilbert, L.I.; Gill, S.S. The Spinosyns: Chemistry, Biochemistry, Mode of Action, and Resistance. In Insect Control: Biological and Synthetic Agents; Academic Press: Cambridge, MA, USA, 2010; pp. 137–169. [Google Scholar]
- Madduri, K.; Waldron, C.; Merlo, D.J. Rhamnose Biosynthesis Pathway Supplies Precursors for Primary and Secondary Metabolism in Saccharopolyspora spinosa. J. Bacteriol. 2001, 183, 5632–5638. [Google Scholar] [CrossRef]
- Huang, K.-x.; Xia, L.; Zhang, Y.; Ding, X.; Zahn, J.A. Recent advances in the biochemistry of spinosyns. Appl. Microbiol. Biotechnol. 2009, 82, 13–23. [Google Scholar] [CrossRef]
- Kirst, H.A. The spinosyn family of insecticides: Realizing the potential of natural products research. J. Antibiot. 2010, 63, 101–111. [Google Scholar] [CrossRef]
- Santos, V.S.V.; Pereira, B.B. Properties, toxicity and current applications of the biolarvicide spinosad. J. Toxicol. Environ. Health B Crit. Rev. 2020, 23, 13–26. [Google Scholar] [CrossRef]
- Tamilselvan, R.; Kennedy, J.S.; Suganthi, A. Sublethal and transgenerational effects of spinetoram on the biological traits of Plutella xylostella (L.) (Lepidoptera: Plutellidae). Ecotoxicology 2021, 30, 667–677. [Google Scholar] [CrossRef]
- Nault, B.A.; Sandhi, R.K.; Harding, R.S.; Grundberg, E.A.; Rusinek, T.; Munyaneza, J. Optimizing Spinosyn Insecticide Applications for Allium Leafminer (Diptera: Agromyzidae) Management in Allium Crops. J. Econ. Entomol. 2022, 115, 618–623. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.-D.; Guo, L.-H.; Ali, A.; Desneux, N.; Zang, L.-S.; Musser, F. Synergism of Adjuvants Mixed With Spinetoram for the Management of Bean Flower Thrips,Megalurothrips usitatus (Thysanoptera: Thripidae) in Cowpeas. J. Econ. Entomol. 2022, 115, 2013–2019. [Google Scholar] [CrossRef] [PubMed]
- Mayes, M.A.; Thompson, G.D.; Husband, B.; Miles, M.M. Spinosad Toxicity to Pollinators and Associated Risk. Rev. Environ. Contam. Toxicol. 2002, 179, 37–71. [Google Scholar] [CrossRef]
- Sparks, T.C.; Crouse, G.D.; Durst, G. Natural products as insecticides: The biology, biochemistry and quantitative structure–activity relationships of spinosyns and spinosoids. Pest Manag. Sci. 2001, 57, 896–905. [Google Scholar] [CrossRef]
- Perry, T.; Chen, W.; Ghazali, R.; Yang, Y.T.; Christesen, D.; Martelli, F.; Lumb, C.; Bao Luong, H.N.; Mitchell, J.; Holien, J.K.; et al. Role of nicotinic acetylcholine receptor subunits in the mode of action of neonicotinoid, sulfoximine and spinosyn insecticides in Drosophila melanogaster. Insect Biochem. Mol. Biol. 2021, 131, 103547. [Google Scholar] [CrossRef]
- Yang, M.; Wang, B.; Gao, J.; Zhang, Y.; Xu, W.; Tao, L. Spinosad induces programmed cell death involves mitochondrial dysfunction and cytochrome C release in Spodoptera frugiperda Sf9 cells. Chemosphere 2017, 169, 155–161. [Google Scholar] [CrossRef]
- Gokbulut, C.; Ozuicli, M.; Aslan, B.; Aydin, L.; Cirak, V.Y. The residue levels of spinosad and abamectin in eggs and tissues of laying hens following spray application. Avian Pathol. 2019, 48 (Suppl. S1), S44–S51. [Google Scholar] [CrossRef]
- Liu, Y.; Sun, H.; Wang, S. Dissipation and Residue of Spinosad in Zucchini Under Field Conditions. Bull. Environ. Contam. Toxicol. 2013, 91, 256–259. [Google Scholar] [CrossRef]
- Sarfraz, M.; Dosdall, L.M.; Keddie, B.A. Spinosad: A Promising Tool for Integrated Pest Management. Outlooks Pest Manag. 2005, 16, 78–84. [Google Scholar] [CrossRef]
- Tao, H.; Zhang, Y.; Deng, Z.; Liu, T. Strategies for Enhancing the Yield of the Potent Insecticide Spinosad in Actinomycetes. Biotechnol. J. 2018, 14, e1700769. [Google Scholar] [CrossRef]
- Bridget, A.F.; Nguyen, C.T.; Magar, R.T.; Sohng, J.K. Increasing production of spinosad in Saccharopolyspora spinosa by metabolic engineering. Biotechnol. Appl. Biochem. 2022, 70, 1035–1043. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhu, Z.; Tang, J.; He, H.; Wan, Q.; Luo, Y.; Huang, W.; Yu, Z.; Hu, Y.; Ding, X.; et al. RNA-Seq-Based Transcriptomic Analysis of Saccharopolyspora spinosa Revealed the Critical Function of PEP Phosphonomutase in the Replenishment Pathway. J. Agric. Food Chem. 2020, 68, 14660–14669. [Google Scholar] [CrossRef] [PubMed]
- Jha, A.K.; Pokhrel, A.R.; Chaudhary, A.K.; Park, S.-W.; Cho, W.J.; Sohng, J.K. Metabolic Engineering of Rational Screened Saccharopolyspora spinosa for the Enhancement of Spinosyns A and D Production. Mol. Cells 2014, 37, 727–733. [Google Scholar] [CrossRef]
- Ottenheim, C.; Nawrath, M.; Wu, J.C. Microbial mutagenesis by atmospheric and room-temperature plasma (ARTP): The latest development. Bioresour. Bioprocess. 2018, 5, 12. [Google Scholar] [CrossRef]
- Buyuklyan, J.A.; Zakalyukina, Y.V.; Osterman, I.A.; Biryukov, M.V. Modern Approaches to the Genome Editing of Antibiotic Biosynthetic Clusters in Actinomycetes. Acta Naturae 2023, 15, 4–16. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, G.; Chen, Y.; Lu, Y. Challenges and Advances in Genome Editing Technologies in Streptomyces. Biomolecules 2020, 10, 734. [Google Scholar] [CrossRef]
- Twardowski, T.; Małyska, A. Uninformed and disinformed society and the GMO market. Trends Biotechnol. 2015, 33, 1–3. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, C.; Zhou, Q.Q.; Zhang, X.F.; Wang, L.Y.; Chang, H.B.; Li, H.P.; Oda, Y.; Xing, X.H. Quantitative evaluation of DNA damage and mutation rate by atmospheric and room-temperature plasma (ARTP) and conventional mutagenesis. Appl. Microbiol. Biotechnol. 2015, 99, 5639–5646. [Google Scholar] [CrossRef]
- Gu, C.; Wang, G.; Mai, S.; Wu, P.; Wu, J.; Wang, G.; Liu, H.; Zhang, J. ARTP mutation and genome shuffling of ABE fermentation symbiotic system for improvement of butanol production. Appl. Microbiol. Biot. 2017, 101, 2189–2199. [Google Scholar] [CrossRef]
- Maslowska, K.H.; Makiela-Dzbenska, K.; Fijalkowska, I.J. The SOS system: A complex and tightly regulated response to DNA damage. Environ. Mol. Mutagen. 2019, 60, 368–384. [Google Scholar] [CrossRef]
- Wang, L.; Zhao, H.; He, D.; Wu, Y.; Jin, L.; Li, G.; Su, N.; Li, H.; Xing, X.H. Insights into the molecular-level effects of atmospheric and room-temperature plasma on mononucleotides and single-stranded homo- and hetero-oligonucleotides. Sci. Rep. 2020, 10, 14298. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Miao, R.; Feng, R.; Yan, J.; Wang, T.; Gan, Y.; Zhao, J.; Lin, J.; Gan, B. Application of Atmospheric and Room-Temperature Plasma (ARTP) to Microbial Breeding. Curr. Issues Mol. Biol. 2023, 45, 6466–6484. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, X.F.; Li, H.P.; Wang, L.Y.; Zhang, C.; Xing, X.H.; Bao, C.Y. Atmospheric and room temperature plasma (ARTP) as a new powerful mutagenesis tool. Appl. Microbiol. Biotechnol. 2014, 98, 5387–5396. [Google Scholar] [CrossRef]
- Xu, Y.; Jing, Y.; Zhang, Q.; Xiu, J.; Tian, M.; Cui, Q.; Ma, Y.; Yi, L.; Han, L.; Qian, Y.; et al. Improving rhamnolipids biosynthesis in Pseudomonas sp. L01 through atmospheric and room-temperature plasma (ARTP) mutagenesis. Microorganisms 2023, 11, 1182. [Google Scholar] [CrossRef]
- Yu, L.; Li, F.; Ni, J.; Qin, X.; Lai, J.; Su, X.; Li, Z.; Zhang, M. UV-ARTP compound mutagenesis breeding improves macrolactins production of Bacillus siamensis and reveals metabolism changes by proteomic. J. Biotechnol. 2024, 381, 36–48. [Google Scholar] [CrossRef]
- Zhu, Z.; Chen, W.; Zhou, H.; Cheng, H.; Luo, S.; Zhou, K.; Zhou, P.; Xia, L.; Ding, X. ARTP and NTG compound mutations improved Cry protein production and virulence of Bacillus thuringiensis X023. Appl. Microbiol. Biotechnol. 2022, 106, 4211–4221. [Google Scholar] [CrossRef]
- Li, D.; Shen, J.; Ding, Q.; Wu, J.; Chen, X. Recent progress of atmospheric and room-temperature plasma as a new and promising mutagenesis technology. Cell Biochem. Funct. 2024, 42, e3991. [Google Scholar] [CrossRef]
- Phan, T.T.; Nguyen, H.D.; Schumann, W. Development of a strong intracellular expression system for Bacillus subtilis by optimizing promoter elements. J. Biotechnol. 2012, 157, 167–172. [Google Scholar] [CrossRef]
- Liu, G.; Chater, K.F.; Chandra, G.; Niu, G.; Tan, H. Molecular Regulation of Antibiotic Biosynthesis in Streptomyces. Microbiol. Mol. Biol. Rev. 2013, 77, 112–143. [Google Scholar] [CrossRef]
- Jain, P.; Malakar, B.; Khan, M.Z.; Lochab, S.; Singh, A.; Nandicoori, V.K. Delineating FtsQ-mediated regulation of cell division in Mycobacterium tuberculosis. J. Biol. Chem. 2018, 293, 12331–12349. [Google Scholar] [CrossRef]
- Li, J.; Wang, N.; Tang, Y.; Cai, X.; Xu, Y.; Liu, R.; Wu, H.; Zhang, B. Developmental regulator BldD directly regulates lincomycin biosynthesis in Streptomyces lincolnensis. Biochem. Biophys. Res. Commun. 2019, 518, 548–553. [Google Scholar] [CrossRef] [PubMed]
- Mouri, Y.; Konishi, K.; Fujita, A.; Tezuka, T.; Ohnishi, Y.; DiRita, V.J. Regulation of Sporangium Formation by BldD in the Rare Actinomycete Actinoplanes missouriensis. J. Bacteriol. 2017, 199, e00840-16. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Lu, X.; Sun, D.; Zhuang, S.; Chen, Q.; Chen, Z.; Li, J.; Wen, Y. BldD, a master developmental repressor, activates antibiotic production in two Streptomyces species. Mol. Microbiol. 2019, 113, 123–142. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Ye, B.; Cheng, S.; Zhao, L.; Liu, Y.; Jiang, J.; Yan, X. Promoter engineering enables overproduction of foreign proteins from a single copy expression cassette in Bacillus subtilis. Microb. Cell Fact. 2019, 18, 111. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Dong, Y.-Q.; Shen, J.-L.; Yin, B.-C.; Ye, B.-C.; You, D. A meet-up of acetyl phosphate and c-di-GMP modulates BldD activity for development and antibiotic production. Nucleic Acids Res. 2023, 51, 6870–6882. [Google Scholar] [CrossRef]
- Bechthold, A.; Zechel, D.L.; Paululat, T.; Nuzzo, D.; Tsypik, O.; Makitrynskyy, R. Secondary nucleotide messenger c-di-GMP exerts a global control on natural product biosynthesis in Streptomycetes. Nucleic Acids Res. 2020, 48, 1583–1598. [Google Scholar] [CrossRef]
- Nuzzo, D.; Makitrynskyy, R.; Tsypik, O.; Bechthold, A. Identification and Characterization of Four c-di-GMP-Metabolizing Enzymes from Streptomyces ghanaensis ATCC14672 Involved in the Regulation of Morphogenesis and Moenomycin A Biosynthesis. Microorganisms 2021, 9, 284. [Google Scholar] [CrossRef]
- Craney, A.; Ahmed, S.; Nodwell, J. Towards a new science of secondary metabolism. J. Antibiot. 2013, 66, 387–400. [Google Scholar] [CrossRef]
- Jin, X.-M.; Chang, Y.-K.; Lee, J.H.; Hong, S.-K. Effects of Increased NADPH Concentration by Metabolic Engineering of the Pentose Phosphate Pathway on Antibiotic Production and Sporulation in Streptomyces lividans TK24. J. Microbiol. Biotechnol. 2017, 27, 1867–1876. [Google Scholar] [CrossRef]
- Swiercz, J.P.; Nanji, T.; Gloyd, M.; Guarné, A.; Elliot, M.A. A novel nucleoid-associated protein specific to the actinobacteria. Nucleic Acids Res. 2013, 41, 4171–4184. [Google Scholar] [CrossRef]
- Zhao, C.; Huang, Y.; Guo, C.; Yang, B.L.; Zhang, Y.; Lan, Z.; Guan, X.; Song, Y.; Zhang, X.L. Heterologous expression of spinosyn biosynthetic gene cluster in species is dependent on the expression of rhamnose biosynthesis genes. J. Mol. Microb. Biotech. 2017, 27, 190–198. [Google Scholar] [CrossRef]
- Liu, Z.; Xiao, J.; Tang, J.; Liu, Y.; Shuai, L.; Cao, L.; Xia, Z.; Ding, X.; Rang, J.; Xia, L. Effects of acuC on the growth development and spinosad biosynthesis of Saccharopolyspora spinosa. Microb. Cell Fact. 2021, 20, 141. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | LC50 (μL/cm2) | 95% Confidence Interval | ||||
|---|---|---|---|---|---|---|
| Pest | ||||||
| H. armigera | S. exigua | S. litura | H. armigera | S. exigua | S. litura | |
| S. spinosa | 1.890 | 1.262 | 2.478 | 1.631–2.259 | 1.057–1.439 | 2.176–3.028 |
| NT24 | 1.305 | 0.511 | 1.372 | 1.053–1.521 | 0.092–0.768 | 1.195–1.539 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, Z.; Chen, W.; Cao, L.; Xia, Z.; Rang, J.; Hu, S.; Xia, L. ARTP/NTG Compound Mutagenesis Improved the Spinosad Production and the Insecticidal Virulence of Saccharopolyspora Spinosa. Int. J. Mol. Sci. 2024, 25, 12308. https://doi.org/10.3390/ijms252212308
Zhu Z, Chen W, Cao L, Xia Z, Rang J, Hu S, Xia L. ARTP/NTG Compound Mutagenesis Improved the Spinosad Production and the Insecticidal Virulence of Saccharopolyspora Spinosa. International Journal of Molecular Sciences. 2024; 25(22):12308. https://doi.org/10.3390/ijms252212308
Chicago/Turabian StyleZhu, Zirong, Wangqiong Chen, Li Cao, Ziyuan Xia, Jie Rang, Shengbiao Hu, and Liqiu Xia. 2024. "ARTP/NTG Compound Mutagenesis Improved the Spinosad Production and the Insecticidal Virulence of Saccharopolyspora Spinosa" International Journal of Molecular Sciences 25, no. 22: 12308. https://doi.org/10.3390/ijms252212308
APA StyleZhu, Z., Chen, W., Cao, L., Xia, Z., Rang, J., Hu, S., & Xia, L. (2024). ARTP/NTG Compound Mutagenesis Improved the Spinosad Production and the Insecticidal Virulence of Saccharopolyspora Spinosa. International Journal of Molecular Sciences, 25(22), 12308. https://doi.org/10.3390/ijms252212308