
Potential Antitumor Mechanism of Propolis Against Skin Squamous Cell Carcinoma A431 Cells Based on Untargeted Metabolomics


Abstract
1. Introduction
2. Results
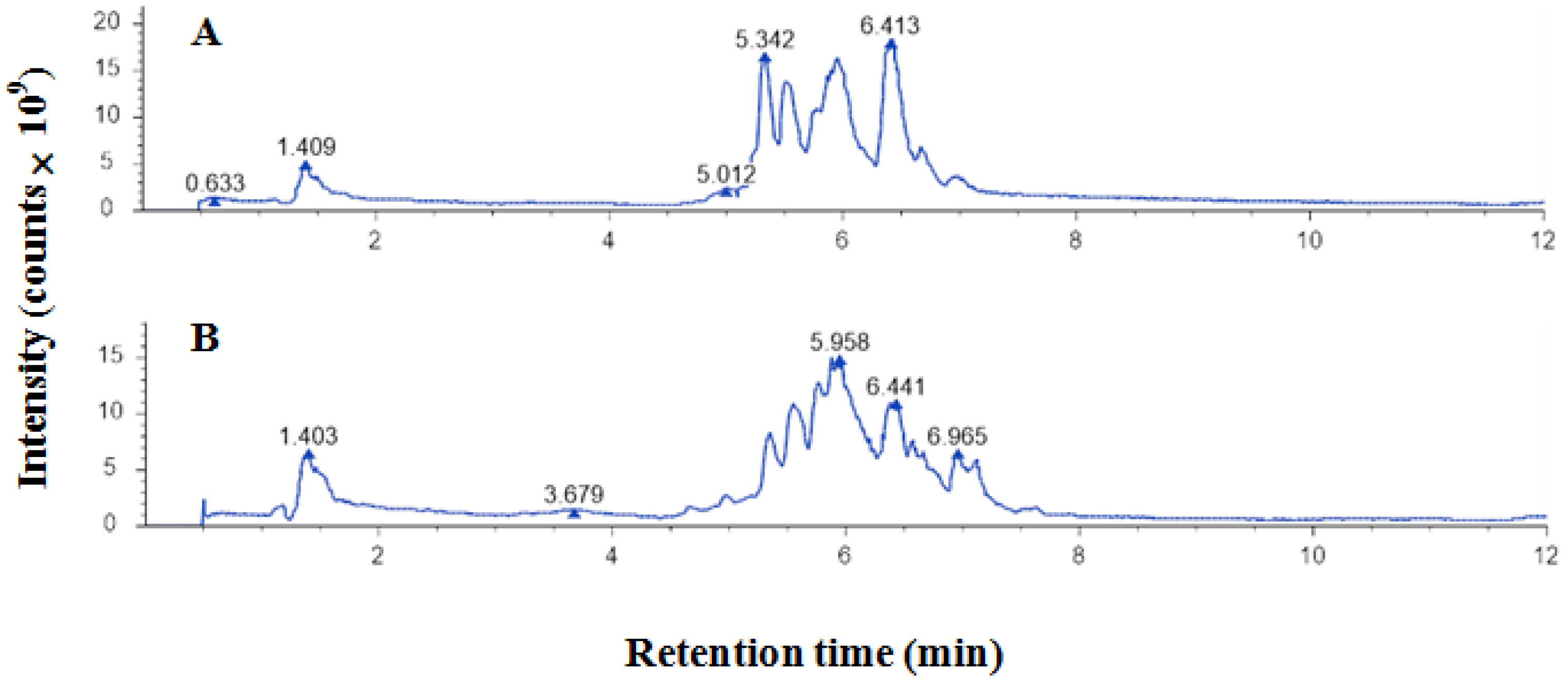
2.1. Polyphenols of EEP
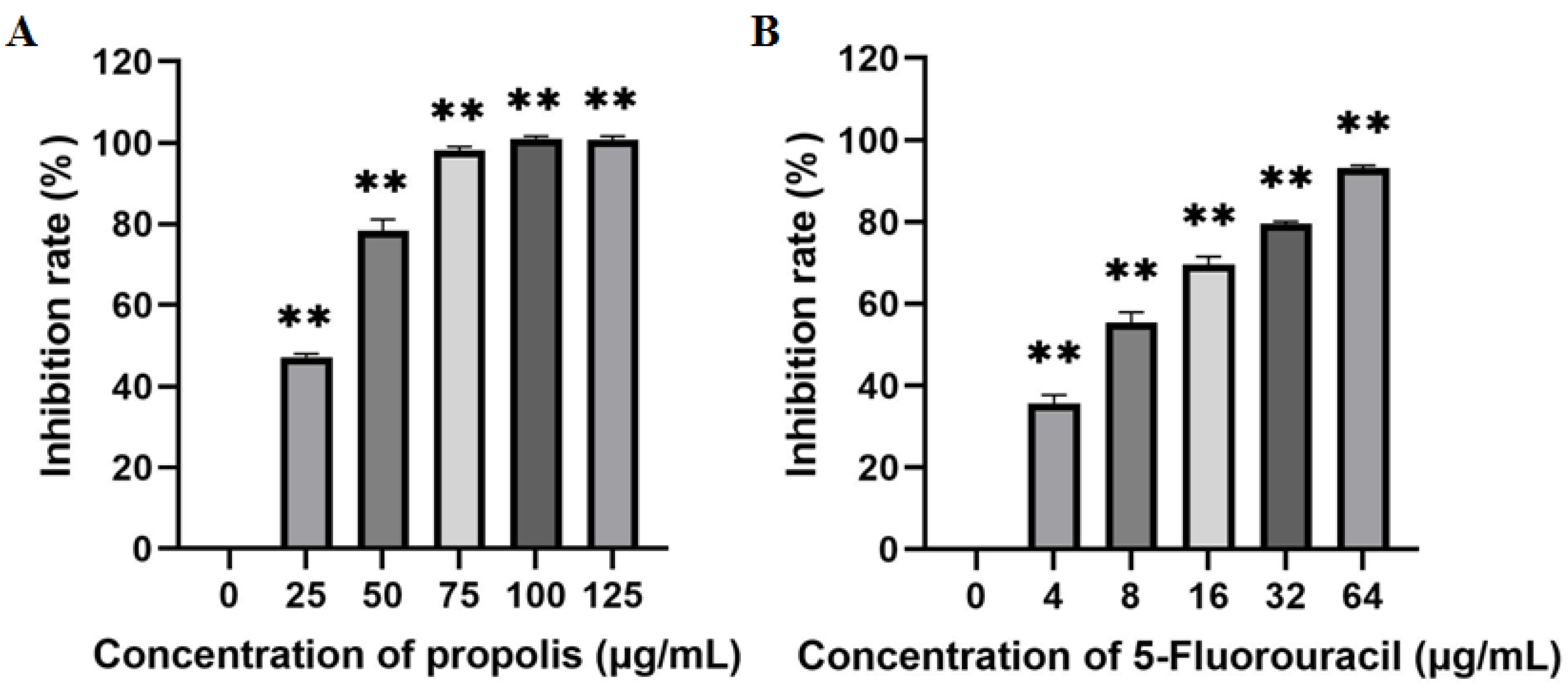
2.2. Inhibitory Effect of EEP on the Proliferation of A431 Cells
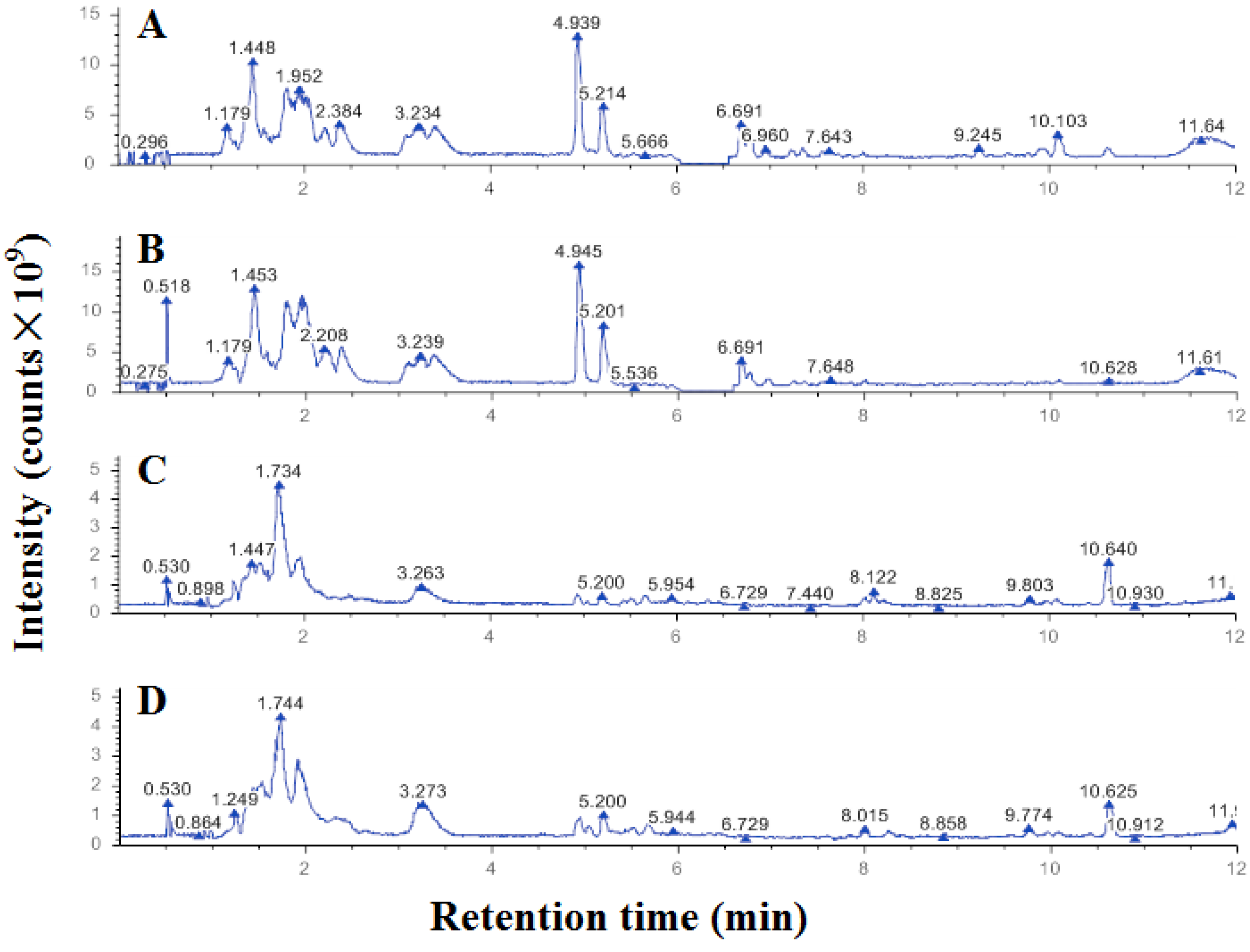
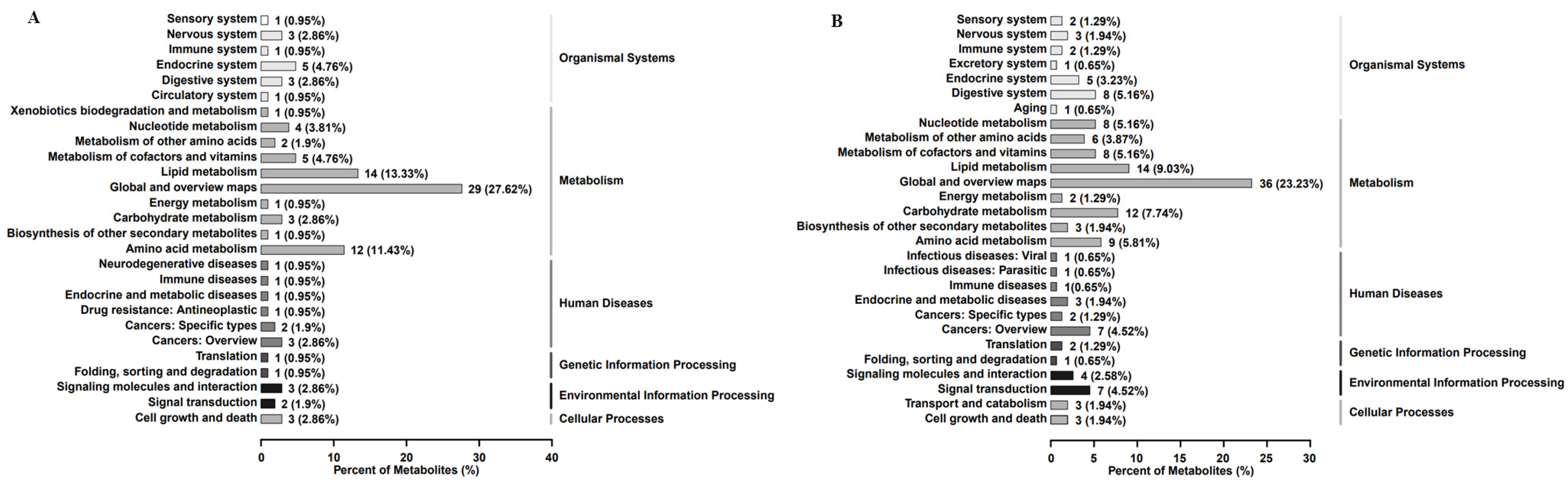
2.3. Differential Metabolites of A431 Cells Induced by EEP
3. Discussion
4. Materials and Methods
4.1. Propolis Extraction and Polyphenols Determination
4.2. Antitumor Effects of Propolis on the Proliferation of A431 Cells
4.3. Untargeted Metabolomic Effects of Propolis on the Proliferation of A431 Cells
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bobiş, O. Plants: Sources of diversity in propolis properties. Plants 2022, 11, 2298. [Google Scholar] [CrossRef] [PubMed]
- Moise, A.R.; Bobiş, O. Baccharis dracunculifolia and Dalbergia ecastophyllum, main plant sources for bioactive properties in green and red Brazilian propolis. Plants 2020, 9, 1619. [Google Scholar] [CrossRef] [PubMed]
- Pant, K.; Sharma, A.; Chopra, H.K.; Nanda, V. Impact of biodiversification on propolis composition, functionality, and application in foods as natural preservative: A review. Food Control 2024, 155, 110097. [Google Scholar] [CrossRef]
- Vagish Kumar, L.S. Propolis in dentistry and oral cancer management. N. Am. J. Med. Sci. 2014, 6, 250–259. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Yañez, N.; Ruiz-Hurtado, P.A.; Rivera-Yañez, C.R.; Arciniega-Martínez, I.M.; Yepez-Ortega, M.; Mendoza-Arroyo, B.; Rebollar-Ruíz, X.A.; Méndez-Cruz, A.R.; Reséndiz-Albor, A.A.; Nieto-Yañez, O. The role of propolis as a natural product with potential gastric cancer treatment properties: A systematic review. Foods 2023, 12, 415. [Google Scholar] [CrossRef]
- Souza, R.P.; Bonfim-Mendonça, P.S.; Damke, G.M.Z.F.; de-Assis Carvalho, A.R.B.; Ratti, B.A.; Dembogurski, D.S.O.; da-Silva, V.R.S.; Silva, S.O.; Da-Silva, D.B.; Bruschi, M.L.; et al. Artepillin C induces selective oxidative stress and inhibits migration and invasion in a comprehensive panel of human cervical cancer cell lines. Anti-Cancer Agents Med. Chem. 2018, 18, 1750–1760. [Google Scholar] [CrossRef]
- Frión-Herrera, Y.; Gabbia, D.; Scaffidi, M.; Zagni, L.; Cuesta-Rubio, O.; De Martin, S.; Carrara, M. Cuban brown propolis interferes in the crosstalk between colorectal cancer cells and M2 macrophages. Nutrients 2020, 12, 2040. [Google Scholar] [CrossRef]
- Meghalatha, T.S.; Muninathan, N. Antitumor activity of withaferin-A and propolis in benz (a) pyrene-induced breast cancer. Bioinformation 2022, 18, 841–844. [Google Scholar] [CrossRef]
- Goto, T.; Kimura, H.; Yoshino, T.; Sawada, A.; Akamatsu, S.; Kobayashi, T.; Yamasaki, T.; Tazawa, S.; Fujimoto, M.; Hidaka, Y.; et al. Efficacy and safety of Brazilian green propolis in biochemically recurrent prostate cancer after radical prostatectomy: A single-arm phase II study. Int. J. Transl. Med. 2022, 2, 618–632. [Google Scholar] [CrossRef]
- Chiu, H.; Han, Y.; Shen, Y.; Golovinskaia, O.; Venkatakrishnan, K.; Wang, C. Chemopreventive and chemotherapeutic effect of propolis and its constituents: A mini-review. J. Cancer Prev. 2020, 25, 70–78. [Google Scholar] [CrossRef]
- Simanjuntak, M.V.; Jauhar, M.M.; Syaifie, P.H.; Arda, A.G.; Mardliyati, E.; Shalannanda, W.; Hermanto, B.R.; Anshori, I. Revealing propolis potential activity on inhibiting estrogen receptor and heat shock protein 90 overexpressed in breast cancer by bioinformatics approaches. Bioinf. Biol. Insights 2024, 18, 11779322231224187. [Google Scholar] [CrossRef] [PubMed]
- Ay, E.N.; Caner, A.; Özsoy Hepokur, C.; Danışman Kalındemirtaş, F.; ÖZEN EROĞLU, G.; Kariper, I.A. Propolis nanoparticles synthesis and characterization with cytotoxic and apoptotic effects on breast cancer cells. J. Taibah Univ. Med. Sci. 2023, 17, 2249628. [Google Scholar] [CrossRef]
- Kocyigit, A.; Guler, E.M.; Durmus, E.; Yenigun, V.B.; Kanimdan, E.; Ozman, Z.; Yasar, O.; Goren, A.C.; Hekimoglu, E.R.; Oruc, H.H.; et al. Propolis enhances 5-fluorouracil mediated antitumor efficacy and reduces side effects in colorectal cancer: An in vitro and in vivo study. Chem. Biodivers. 2023, 20, e202300591. [Google Scholar] [CrossRef] [PubMed]
- Frión-Herrera, Y.; Díaz-García, A.; Ruiz-Fuentes, J.; Rodríguez-Sánchez, H.; Sforcin, J.M. The cytotoxic effects of propolis on breast cancer cells involve PI3K/Akt and ERK1/2 pathways, mitochondrial membrane potential, and reactive oxygen species generation. Inflammopharmacology 2019, 27, 1081–1089. [Google Scholar] [CrossRef]
- Xuan, H.; Li, Z.; Yan, H.; Sang, Q.; Wang, K.; He, Q.; Wang, Y.; Hu, F. Antitumor activity of chinese propolis in human breast cancer MCF-7 and MDA-MB-231 cells. Evid. Based. Complement. Alternat. Med. 2014, 2014, 280120. [Google Scholar] [CrossRef]
- Corchado-Cobos, R.; García-Sancha, N.; González-Sarmiento, R.; Pérez-Losada, J.; Cañueto, J. Cutaneous squamous cell carcinoma: From biology to therapy. Int. J. Mol. Sci. 2020, 21, 2956. [Google Scholar] [CrossRef]
- Cleavenger, J.; Johnson, S.M. Non melanoma skin cancer review. J. Ark. Med. Soc. 2014, 110, 230–234. [Google Scholar] [PubMed]
- Lomas, A.; Leonardi-Bee, J.; Bath-Hextall, F. A systematic review of worldwide incidence of nonmelanoma skin cancer. Br. J. Dermatol. 2012, 166, 1069–1080. [Google Scholar] [CrossRef]
- Xu, C.; Zhuo, Y.; Liu, Y.; Chen, H. Itraconazole inhibits the growth of cutaneous squamous cell carcinoma by targeting HMGCS1/ACSL4 Axis. Front. Pharmacol. 2022, 13, 828983. [Google Scholar] [CrossRef]
- Wang, Y.; Li, Z.; Teng, M.; Liu, J. Dihydroartemisinin inhibits activation of the AIM2 inflammasome pathway and NF-κB/HIF-1α/VEGF pathway by inducing autophagy in A431 human cutaneous squamous cell carcinoma cells. Int. J. Med. Sci. 2021, 18, 2705–2715. [Google Scholar] [CrossRef]
- Liu, W.; Liu, X. LINC00641 inhibits the development of cutaneous squamous cell carcinoma by downregulating miR-424 in A431 Cells. Cancer Biother. Radiopharm. 2021, 39, 532–540. [Google Scholar] [CrossRef] [PubMed]
- Endo-Munoz, L.; Dahler, A.; Teakle, N.; Rickwood, D.; Hazar-Rethinam, M.; Abdul-Jabbar, I.; Sommerville, S.; Dickinson, I.; Kaur, P.; Paquet-Fifield, S.; et al. E2F7 can regulate proliferation, differentiation, and apoptotic responses in human keratinocytes: Implications for cutaneous squamous cell carcinoma formation. Cancer Res. 2009, 69, 1800–1808. [Google Scholar] [CrossRef] [PubMed]
- Teh, S.W.; Elderdery, A.; Rampal, S.; Subbiah, S.K.; Mok, P.L. Clustered regularly interspaced short palindromic repeats (CRISPR)-Cas9 transfection of guide RNA targeting on MMP9 as anti-cancer therapy in human cutaneous squamous cell carcinoma cell line A431. Wspolczesna Onkol. 2023, 27, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Tian, Y.; Yang, A.; Tan, W.; Liu, X.; Yang, W. Antitumor effect of poplar propolis on human cutaneous squamous cell carcinoma A431 cells. Int. J. Mol. Sci. 2023, 24, 16753. [Google Scholar] [CrossRef]
- Ribbenstedt, A.; Ziarrusta, H.; Benskin, J.P. Development, characterization and comparisons of targeted and non-targeted metabolomics methods. PLoS ONE 2018, 13, e0207082. [Google Scholar] [CrossRef]
- Zhang, Y.; Ren, H.; Jiang, Y.; Gao, Y.; Liu, S. Urinary metabonomics of stomach cancer assessed by rapid resolution liquid chromatography/time-of-fight mass spectrometry. Chin. Med. J. (Engl.) 2013, 126, 1930–1933. [Google Scholar] [CrossRef]
- Yu, X.; Wu, Q.; Lu, W.; Wang, Y.; Ma, X.; Chen, Z.; Yan, C. Metabonomics study of lung cancer cells based on liquid l chromatography-mass spectrometry. SePu 2013, 31, 691–696. [Google Scholar] [CrossRef]
- Gan, C.; Huang, X.; Wu, Y.; Zhan, J.; Zhang, X.; Liu, Q.; Huang, Y. Untargeted metabolomics study and pro-apoptotic properties of B-norcholesteryl benzimidazole compounds in ovarian cancer SKOV3 cells. J. Steroid Biochem. Mol. Biol. 2020, 202, 105709. [Google Scholar] [CrossRef]
- Liu, X.; Tian, Y.; Yang, A.; Zhang, C.; Miao, X.; Yang, W. Antitumor effects of poplar propolis on DLBCL SU-DHL-2 cells. Foods 2023, 12, 283. [Google Scholar] [CrossRef]
- Zabaiou, N.; Fouache, A.; Trousson, A.; Baron, S.; Zellagui, A.; Lahouel, M.; Lobaccaro, J.A. Biological properties of propolis extracts: Something new from an ancient product. Chem. Phys. Lipids 2017, 207, 214–222. [Google Scholar] [CrossRef]
- Zheng, Y.; Deng, G.; Liang, Q.; Chen, D.; Guo, R.; Lai, R. Antioxidant activity of quercetin and its glucosides from propolis: A theoretical study. Sci. Rep. 2017, 7, 7543. [Google Scholar] [CrossRef] [PubMed]
- Zulhendri, F.; Felitti, R.; Fearnley, J.; Ravalia, M. The use of propolis in dentistry, oral health, and medicine: A review. J. Oral Biosci. 2021, 63, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Celińska-Janowicz, K.; Zaręba, I.; Lazarek, U.; Teul, J.; Tomczyk, M.; Pałka, J.; Miltyk, W. Constituents of propolis: Chrysin, caffeic acid, p-coumaric acid, and ferulic acid induce PRODH/POX-dependent apoptosis in human tongue squamous cell carcinoma cell (CAL-27). Front. Pharmacol. 2018, 9, 336. [Google Scholar] [CrossRef] [PubMed]
- Frozza, C.O.; Garcia, C.S.; Gambato, G.; de Souza, M.D.; Salvador, M.; Moura, S.; Padilha, F.F.; Seixas, F.K.; Collares, T.; Borsuk, S.; et al. Chemical characterization, antioxidant and cytotoxic activities of Brazilian red propolis. Food Chem. Toxicol. 2013, 52, 137–142. [Google Scholar] [CrossRef]
- Asgharpour, F.; Moghadamnia, A.A.; Zabihi, E.; Kazemi, S.; Ebrahimzadeh Namvar, A.; Gholinia, H.; Motallebnejad, M.; Nouri, H.R. Iranian propolis efficiently inhibits growth of oral streptococci and cancer cell lines. BMC Complement. Altern. Med. 2019, 19, 266. [Google Scholar] [CrossRef]
- Wezgowiec, J.; Wieczynska, A.; Wieckiewicz, W.; Kulbacka, J.; Saczko, J.; Pachura, N.; Wieckiewicz, M.; Gancarz, R.; Wilk, K.A. Polish propolis-chemical composition and biological effects in tongue cancer cells and macrophages. Molecules 2020, 25, 2426. [Google Scholar] [CrossRef]
- Žižić, J.B.; Vuković, N.L.; Jadranin, M.B.; Anđelković, B.D.; Tešević, V.V.; Kacaniova, M.M.; Sukdolak, S.B.; Marković, S.D. Chemical composition, cytotoxic and antioxidative activities of ethanolic extracts of propolis on HCT-116 cell line. J. Sci. Food Agric. 2013, 93, 3001–3009. [Google Scholar] [CrossRef]
- Khacha-Ananda, S.; Tragoolpua, K.; Chantawannakul, P.; Tragoolpua, Y. Propolis extracts from the northern region of Thailand suppress cancer cell growth through induction of apoptosis pathways. Invest. New Drugs 2016, 34, 707–722. [Google Scholar] [CrossRef]
- Slominski, A.; Zbytek, B.; Nikolakis, G.; Manna, P.R.; Skobowiat, C.; Zmijewski, M.; Li, W.; Janjetovic, Z.; Postlethwaite, A.; Zouboulis, C.C.; et al. Steroidogenesis in the skin: Implications for local immune functions. J. Steroid Biochem. Mol. Biol. 2013, 137, 107–123. [Google Scholar] [CrossRef]
- Huang, B.; Song, B.; Xu, C. Cholesterol metabolism in cancer: Mechanisms and therapeutic opportunities. Nat. Metab. 2020, 2, 132–141. [Google Scholar] [CrossRef]
- Bessone, F.; Razori, M.V.; Roma, M.G. Molecular pathways of nonalcoholic fatty liver disease development and progression. Cell. Mol. Life Sci. 2019, 76, 99–128. [Google Scholar] [CrossRef] [PubMed]
- Bélanger, A.; Pelletier, G.; Labrie, F.; Barbier, O.; Chouinard, S. Inactivation of androgens by UDP-glucuronosyltransferase enzymes in humans. Trends Endocrinol. Metab. 2003, 14, 473–479. [Google Scholar] [CrossRef] [PubMed]
- Gauthier-Landry, L.; Bélanger, A.; Barbier, O. Multiple roles for UDP-glucuronosyltransferase (UGT)2B15 and UGT2B17 enzymes in androgen metabolism and prostate cancer evolution. J. Steroid Biochem. Mol. Biol. 2015, 145, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Carmina, E.; Godwin, A.J.; Stanczyk, F.Z.; Lippman, J.S.; Lobo, R.A. The association of serum androsterone glucuronide with inflammatory lesions in women with adult acne. J. Endocrinol. Investig. 2002, 25, 765–768. [Google Scholar] [CrossRef]
- Zhao, H.; Wu, L.; Yan, G.; Chen, Y.; Zhou, M.; Wu, Y.; Li, Y. Inflammation and tumor progression: Signaling pathways and targeted intervention. Signal Transduct. Targeted Ther. 2021, 6, 263. [Google Scholar] [CrossRef]
- Barfeld, S.J.; Itkonen, H.M.; Urbanucci, A.; Mills, I.G. Androgen-regulated metabolism and biosynthesis in prostate cancer. Endocr.-Relat. Cancer. 2014, 21, T57–T66. [Google Scholar] [CrossRef]
- Snaterse, G.; Visser, J.A.; Arlt, W.; Hofland, J. Circulating steroid hormone variations throughout different stages of prostate cancer. Endocr.-Relat. Cancer. 2017, 24, R403–R420. [Google Scholar] [CrossRef]
- Li, Z.; Bao, H. Anti-tumor effect of Inonotus hispidus petroleum ether extract in H22 tumor-bearing mice and analysis its mechanism by untargeted metabonomic. J. Ethnopharmacol. 2022, 285, 114898. [Google Scholar] [CrossRef]
- Panin, L.E.; Khoshchenko, O.M.; Poliakov, L.M. Effect of apolipoprotein A-1 containing steroid hormones on DNA and protein biosynthesis in cells of ascitic hepatoma HA-1. Vopr. Onkol. 2007, 53, 562–565. [Google Scholar]
- Valko-Rokytovská, M.; Očenáš, P.; Salayová, A.; Kostecká, Z. Breast cancer: Targeting of steroid hormones in cancerogenesis and diagnostics. Int. J. Mol. Sci. 2021, 22, 5878. [Google Scholar] [CrossRef]
- Murphy, L.J. Growth factors and steroid hormone action in endometrial cancer. J. Steroid Biochem. Mol. Biol. 1994, 48, 419–423. [Google Scholar] [CrossRef] [PubMed]
- Saito, R.F.; Andrade, L.N.S.; Bustos, S.O.; Chammas, R. Phosphatidylcholine-derived lipid mediators: The crosstalk between cancer cells and immune cells. Front. Immunol. 2022, 13, 768606. [Google Scholar] [CrossRef] [PubMed]
- Spiteller, P.; Kern, W.; Reiner, J.; Spiteller, G. Aldehydic lipid peroxidation products derived from linoleic acid. Biochim. Biophys. Acta. 2001, 1531, 188–208. [Google Scholar] [CrossRef] [PubMed]
- Nava Lauson, C.B.; Tiberti, S.; Corsetto, P.A.; Conte, F.; Tyagi, P.; Machwirth, M.; Ebert, S.; Loffreda, A.; Scheller, L.; Sheta, D.; et al. Linoleic acid potentiates CD8+ T cell metabolic fitness and antitumor immunity. Cell Metab. 2023, 35, 633–650. [Google Scholar] [CrossRef]
- Liu, J.; Geng, W.; Sun, H.; Liu, C.; Huang, F.; Cao, J.; Xia, L.; Zhao, H.; Zhai, J.; Li, Q.; et al. Integrative metabolomic characterisation identifies altered portal vein serum metabolome contributing to human hepatocellular carcinoma. Gut 2022, 71, 1203–1213. [Google Scholar] [CrossRef]
- Qiu, J.; Zhao, Z.; Suo, H.; Paraghamian, S.E.; Hawkins, G.M.; Sun, W.; Zhang, X.; Hao, T.; Deng, B.; Shen, X.; et al. Linoleic acid exhibits anti-proliferative and anti-invasive activities in endometrial cancer cells and a transgenic model of endometrial cancer. Cancer Biol. Ther. 2024, 25, 2325130. [Google Scholar] [CrossRef]
- Wang, W.; Yu, L.; Li, Z.; Xiao, Y.; Jiang, H.; Tang, Y.L.; Chen, Y.; Xue, H. Dysregulated arginine metabolism in precursor B-cell acute lymphoblastic leukemia in children: A metabolomic study. BMC Pediatr. 2024, 24, 540. [Google Scholar] [CrossRef]
- Zhu, S.; Han, X.; Yang, R.; Tian, Y.; Zhang, Q.; Wu, Y.; Dong, S.; Zhang, B. Metabolomics study of ribavirin in the treatment of orthotopic lung cancer based on UPLC-Q-TOF/MS. Chem.-Biol. Interact. 2023, 370, 110305. [Google Scholar] [CrossRef]
- Zhao, L.; Yao, L.; Chen, R.; He, J.; Lin, T.; Qiu, S.; Chen, G.; Chen, H.; Qiu, S. Pinostrobin from plants and propolis against human coronavirus HCoV-OC43 by modulating host AHR/CYP1A1 pathway and lipid metabolism. Antivir. Res. 2023, 212, 105570. [Google Scholar] [CrossRef]
- Tsuji, P.A.; Winn, R.N.; Walle, T. Accumulation and metabolism of the anticancer flavonoid 5,7-dimethoxyflavone compared to its unmethylated analog chrysin in the Atlantic killifish. Chem. Biol. Interact. 2006, 164, 85–92. [Google Scholar] [CrossRef]
- Kumazawa, S.; Shimoi, K.; Hayashi, K.; Ishii, T.; Hamasaka, T.; Nakayama, T. Identification of metabolites in plasma and urine of Uruguayan propolis-treated rats. J. Agric. Food Chem. 2004, 52, 3083–3088. [Google Scholar] [CrossRef] [PubMed]
- Stabrauskiene, J.; Kopustinskiene, D.M.; Lazauskas, R.; Bernatoniene, J. Naringin and naringenin: Their mechanisms of action and the potential anticancer activities. Biomedicines 2022, 10, 1686. [Google Scholar] [CrossRef] [PubMed]
- Rao, H.; Liu, C.; Wang, A.; Ma, C.; Xu, Y.; Ye, T.; Su, W.; Zhou, P.; Gao, W.Q.; Li, L.; et al. SETD2 deficiency accelerates sphingomyelin accumulation and promotes the development of renal cancer. Nat. Commun. 2023, 14, 7572. [Google Scholar] [CrossRef] [PubMed]
- Tan, G.C.; Negro, G.; Pinggera, A.; Tizen Laim, N.M.S.; Mohamed Rose, I.; Ceral, J.; Ryska, A.; Chin, L.K.; Kamaruddin, N.A.; Mohd Mokhtar, N.; et al. Aldosterone-producing adenomas: Histopathology-genotype correlation and identification of a novel CACNA1D mutation. Hypertension 2017, 70, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Makowski, N.; Burckhardt, B.B. Enabling insights into the maturation of the renin-angiotensin-aldosterone system in children-Development of a low-volume LC-MS assay for the simultaneous determination of aldosterone, its precursor, and main metabolite. Steroids 2019, 148, 73–81. [Google Scholar] [CrossRef]
- Sun, Y.; Zhou, Q.; Chen, F.; Gao, X.; Yang, L.; Jin, X.; Wink, M.; Sharopov, F.S.; Sethi, G. Berberine inhibits breast carcinoma proliferation and metastasis under hypoxic microenvironment involving gut microbiota and endogenous metabolites. Pharmacol. Res. 2023, 193, 106817. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Name | Formula | Molecular Weight | RT/[min] | m/z | Relative Quantitative Value | Polarity Ion Mode |
|---|---|---|---|---|---|---|---|
| 1 | 7-Methoxyflavone | C16H12O3 | 252.0743 | 2.319 | 253.0816 | 165,691,937.9 | positive |
| 2 | 4′-Methoxyflavone | C16H12O3 | 252.0742 | 2.751 | 253.08127 | 40,927,754.05 | positive |
| 3 | Eriodictyol | C15H12O6 | 242.0573 | 5.114 | 287.05548 | 19,766,216.42 | negative |
| 4 | Taxifolin | C15H12O7 | 304.0579 | 5.17 | 305.06513 | 33,877,502.65 | positive |
| 5 | Neodiosmin | C28H32O15 | 608.1722 | 5.334 | 609.17948 | 25,006,731.57 | positive |
| 6 | Hesperetin | C16H14O6 | 302.0782 | 5.406 | 320.11203 | 98,982,625.12 | positive |
| 7 | Rutin | C27H30O16 | 610.1532 | 5.463 | 609.14589 | 60,165,204.17 | negative |
| 8 | Quercetin-3β-D-glucoside | C21H20O12 | 464.0958 | 5.497 | 463.0885 | 150,118,518.8 | negative |
| 9 | Epicatechin | C15H14O6 | 290.0793 | 5.547 | 289.07202 | 205,621,918.1 | negative |
| 10 | Nobiletin | C21H22O8 | 442.1237 | 5.555 | 425.12034 | 38,494,781.4 | positive |
| 11 | Trifolin | C21H20O11 | 448.0999 | 5.59 | 449.10716 | 129,227,583.4 | positive |
| 12 | Galangin | C15H10O5 | 270.0526 | 5.784 | 271.05985 | 6,988,985,974 | positive |
| 13 | Quercetin | C15H10O7 | 302.0424 | 5.808 | 301.03512 | 12,911,294,036 | negative |
| 14 | Kaempferol | C15H10O6 | 286.0477 | 6.025 | 285.04042 | 3,447,864,851 | negative |
| 15 | Apigenin | C15H10O5 | 270.0526 | 6.048 | 271.05989 | 7,294,339,651 | positive |
| 16 | Sakuranin | C22H24O10 | 470.1182 | 6.36 | 471.12546 | 148,046,024.5 | positive |
| 17 | Pinocembrin | C15H12O4 | 256.0733 | 6.452 | 257.0806 | 3,402,672,117 | positive |
| 18 | Chrysin | C15H10O4 | 254.0577 | 6.606 | 255.06497 | 9,396,111,669 | positive |
| 19 | Kaempferitrin | C27H30O14 | 578.159 | 6.657 | 579.16626 | 15,900,785.8 | positive |
| 20 | Wogonin | C16H12O5 | 284.0682 | 6.773 | 285.07551 | 1,690,423,123 | positive |
| 21 | Luteolin | C15H10O6 | 286.0477 | 6.95 | 285.04047 | 1,048,387,356 | negative |
| 22 | Naringenin | C15H12O5 | 272.0685 | 6.959 | 271.06118 | 6,956,970,847 | negative |
| 23 | Vitexin | C21H20O10 | 454.0871 | 6.968 | 455.09441 | 328,176,072.8 | positive |
| 24 | Myricitrin | C21H20O12 | 464.0945 | 9.333 | 463.08725 | 1,004,733.206 | negative |
| 25 | Metanephrine | C10H15NO3 | 197.105 | 1.782 | 198.11208 | 148,715,444.3 | positive |
| 26 | Epinephrinebitartrate | C13H19NO9 | 333.105 | 1.99 | 334.11243 | 51,641,235.49 | positive |
| 27 | Vanillin | C8H8O3 | 130.063 | 4.603 | 131.07061 | 12,412,803.31 | positive |
| 28 | 2-Methoxyresorcinol | C7H8O3 | 140.047 | 4.908 | 141.05468 | 49,647,059.53 | positive |
| 29 | Catechol | C6H6O2 | 110.037 | 5.035 | 109.02963 | 221,250,004.2 | negative |
| 30 | Vanillylalcohol | C8H10O3 | 154.063 | 5.129 | 153.05594 | 167,498,687.3 | negative |
| 31 | N-Acetyldopamine | C10H13NO3 | 195.09 | 5.143 | 196.09692 | 253,902,426.9 | positive |
| 32 | 3-Methoxytyramine | C9H13NO2 | 167.095 | 5.225 | 202.06389 | 3,554,901.715 | negative |
| 33 | L-Adrenaline | C9H13NO3 | 183.089 | 5.23 | 184.09673 | 39,891,603.29 | positive |
| 34 | Hydroquinone | C6H6O2 | 110.037 | 5.26 | 111.04412 | 142,215,214.3 | positive |
| 35 | Homovanillicacid | C9H10O4 | 182.058 | 5.266 | 181.05065 | 99,031,088.98 | negative |
| 36 | 4-Methylphenol | C7H8O | 108.058 | 5.33 | 107.05028 | 64,119,942.28 | negative |
| 37 | Isoproterenol | C11H17NO3 | 211.12 | 5.55 | 210.11346 | 23,052,347.25 | negative |
| 38 | Eugenol | C10H12O2 | 164.084 | 5.598 | 163.07669 | 141,167,860.1 | negative |
| 39 | o-Cresol | C7H8O | 108.057 | 5.617 | 109.06481 | 32,449,852.97 | positive |
| 40 | 4-Nitrophenol | C6H5NO3 | 139.027 | 5.796 | 138.0198 | 83,038,733.11 | negative |
| 41 | 4-Butylresorcinol | C10H14O2 | 166.099 | 5.829 | 149.09605 | 103,300,893.4 | positive |
| 42 | Phloroglucinol | C6H6O3 | 126.032 | 6.064 | 125.02453 | 1,391,500,576 | negative |
| 43 | Pyrogallol | C6H6O3 | 126.032 | 11.989 | 127.039 | 95,430,031.51 | positive |
| ID | Name | Formula | Molecular Weight | RT/[min] | m/z | p Value | Up/Downregulated | Polarity Mode |
|---|---|---|---|---|---|---|---|---|
| 1 | (3R)-8-hydroxy-3-(4-methoxyphenyl)-3,4-dihydro-1H-2-benzopyran-1-one | C16H14O4 | 270.08975 | 6.435 | 269.08247 | 1.02 × 10−14 | up | negative |
| 2 | (+/−)12(13)-DiHOME | C18H34O4 | 296.2349 | 8.268 | 319.22416 | 1.08 × 10−11 | up | positive |
| 3 | Maslinicacid | C30H48O4 | 494.33677 | 8.303 | 495.34404 | 7.59 × 10−10 | up | positive |
| 4 | (2R)-5-hydroxy-7-methoxy-2-phenyl-3,4-dihydro-2H-1-benzopyran-4-one | C16H14O4 | 270.08863 | 7.197 | 271.0959 | 8.28 × 10−10 | up | positive |
| 5 | Genistein | C15H10O5 | 270.05336 | 6.733 | 269.04608 | 1.6 × 10−9 | up | negative |
| 6 | 5,6-dimethoxy-2-(2-methoxyphenyl)-4H-chromen-4-one | C18H16O5 | 294.08863 | 6.858 | 295.09591 | 1.86 × 10−9 | up | positive |
| 7 | Naringenin | C15H12O5 | 272.06895 | 5.481 | 271.06168 | 2.52 × 10−9 | up | negative |
| 8 | trans-10-Heptadecenoicacid | C17H32O2 | 314.24627 | 7.167 | 313.23899 | 2.67 × 10−9 | up | negative |
| 9 | 4-methoxy-6-[2-(4-methoxyphenyl)ethyl]-2H-pyran-2-one | C15H16O4 | 282.08876 | 8.337 | 283.09604 | 3.85 × 10−9 | up | positive |
| 10 | Pinocembrin | C15H12O4 | 256.07376 | 5.918 | 257.08104 | 4.47 × 10−9 | up | positive |
| 11 | Adrenosterone | C19H24O3 | 300.17302 | 8.198 | 299.16574 | 7.19 × 10−9 | up | negative |
| 12 | 12-epiLeukotrieneB4 | C20H32O4 | 318.21778 | 8.275 | 363.216 | 7.82 × 10−9 | up | negative |
| 13 | Tetrahydroaldosterone | C21H32O5 | 364.22327 | 8.263 | 363.216 | 9.32 × 10−9 | up | negative |
| 14 | N-[2-acetyl-5-(tert-butyl)-3-thienyl]-2-(propylsulfanyl)nicotinamide | C19H24N2O2S2 | 376.13179 | 6.844 | 375.12451 | 2.25 × 10−8 | up | negative |
| 15 | SM8:1;2O/12:0 | C25H51N2O6P | 506.34854 | 8.729 | 529.33785 | 2.6 × 10−8 | up | positive |
| 16 | Isorhapontigenin | C15H14O4 | 258.08978 | 6.925 | 257.08251 | 2.73 × 10−8 | up | negative |
| 17 | 4-chloro-5-morpholino-2-quinoxalin-2-ylpyridazin-3(2H)-one | C16H14ClN5O2 | 343.084 | 2.011 | 344.09127 | 4.17 × 10−8 | up | positive |
| 18 | Papaverine | C20H21NO4 | 339.14752 | 5.87 | 340.15479 | 4.44 × 10−8 | up | positive |
| 19 | N-Acetylsphingosine | C20H39NO3 | 363.27507 | 9.628 | 364.28235 | 7.49 × 10−8 | up | positive |
| 20 | L-cysteine | C3H7NO2S | 121.01995 | 1.509 | 122.02727 | 9.54 × 10−8 | up | positive |
| 21 | Chrysin | C15H10O4 | 254.05827 | 6.656 | 253.051 | 1.02 × 10−7 | up | negative |
| 22 | Dehydroepiandrosterone(DHEA) | C19H28O2 | 270.1984 | 9.61 | 271.20554 | 1.13 × 10−7 | up | positive |
| 23 | methyl3,4,5-trihydroxycyclohex-1-ene-1-carboxylate | C8H12O5 | 210.05314 | 8.341 | 211.06041 | 1.48 × 10−7 | up | positive |
| 24 | gamma-Glutamylcysteine | C8H14N2O5S | 250.06268 | 1.657 | 249.05541 | 1.71 × 10−7 | up | negative |
| 25 | 1-Methyladenosine | C11H15N5O4 | 281.11189 | 1.883 | 282.11917 | 1.72 × 10−7 | up | positive |
| 26 | Hydrocortisone | C21H30O5 | 362.20763 | 7.825 | 361.20035 | 2.4 × 10−7 | up | negative |
| 27 | Naringeninchalcone | C15H12O5 | 272.06889 | 5.493 | 273.07612 | 2.69 × 10−7 | up | positive |
| 28 | 2-oxo-2H-chromene-3-carboxylicacid | C10H6O4 | 190.0264 | 6.86 | 191.03368 | 3.02 × 10−7 | up | positive |
| 29 | H-Gly-Pro-OH | C7H12N2O3 | 172.08505 | 1.983 | 171.0777 | 3.39 × 10−7 | up | negative |
| 30 | Resveratrol | C14H12O3 | 228.07915 | 6.837 | 227.07188 | 3.39 × 10−7 | up | negative |
| 31 | N-Methylhydantoin | C4H6N2O2 | 114.04328 | 1.739 | 115.05055 | 3.59 × 10−7 | down | positive |
| 32 | 4-MethoxycinnamicAcid | C10H10O3 | 178.06217 | 7.416 | 379.11392 | 4.6 × 10−7 | up | positive |
| 33 | D-Glucosamine6-phosphate | C6H14NO8P | 259.04597 | 1.454 | 260.05325 | 6.57 × 10−7 | up | positive |
| 34 | ProstaglandinA1ethylester | C22H36O4 | 346.25129 | 9.137 | 345.24402 | 8.48 × 10−7 | up | negative |
| 35 | 5,7-dihydroxy-3-(4-hydroxyphenyl)-4H-chromen-4-one | C15H10O5 | 270.05304 | 5.746 | 271.06031 | 9.09 × 10−7 | up | positive |
| 36 | 4-[2-(2-oxo-1-imidazolidinyl)ethyl]-1lambda~6~,4-thiazinane-1,1-dione | C9H17N3O3S | 248.10518 | 6.943 | 247.0979 | 9.44 × 10−7 | up | negative |
| 37 | (+/−)-Equol | C15H14O3 | 242.09476 | 7.083 | 241.08748 | 1.02 × 10−6 | up | negative |
| 38 | Piceatannol | C14H12O4 | 244.07412 | 6.713 | 243.06685 | 1.06 × 10−6 | up | negative |
| 39 | Ursolicacid | C30H48O3 | 456.36128 | 9.427 | 455.354 | 1.28 × 10−6 | up | negative |
| 40 | Glycerol1-hexadecanoate | C19H38O4 | 330.27757 | 8.603 | 329.27033 | 1.31 × 10−6 | up | negative |
| 41 | Glycitein | C16H12O5 | 284.06896 | 6.828 | 283.06168 | 1.32 × 10−6 | up | negative |
| 42 | Mag(18:1) | C21H40O4 | 356.29292 | 10.356 | 357.30024 | 1.35 × 10−6 | down | positive |
| 43 | 5-hydroxy-6,7-dimethoxy-2-phenyl-4H-chromen-4-one | C17H14O5 | 280.0731 | 7.82 | 281.08037 | 1.93 × 10−6 | up | positive |
| 44 | Luteolin | C15H10O6 | 286.04863 | 6.035 | 285.04137 | 2.55 × 10−6 | up | negative |
| 45 | SM8:0;2O/12:0 | C25H53N2O6P | 508.36466 | 9.391 | 531.35384 | 2.69 × 10−6 | up | positive |
| 46 | Cys-Gly | C5H10N2O3S | 178.04139 | 1.703 | 177.03412 | 2.69 × 10−6 | up | negative |
| 47 | ChlorogenicAcidMethylEster | C17H20O9 | 368.11135 | 5.712 | 367.10407 | 2.95 × 10−6 | up | negative |
| 48 | Ethyloleate | C20H38O2 | 310.2871 | 9.807 | 311.2943 | 3.03 × 10−6 | up | positive |
| 49 | Thiazolidine-4-carboxylicacid | C4H7NO2S | 133.01974 | 1.501 | 134.02701 | 3.04 × 10−6 | up | positive |
| 50 | P-AminohippuricAcid | C9H10N2O3 | 194.06932 | 3.142 | 195.07649 | 3.46 × 10−6 | down | positive |
| 51 | Acetyl-L-carnitine | C9H17NO4 | 203.11608 | 1.768 | 204.12335 | 3.6 × 10−6 | down | positive |
| 52 | JWH250N-pentanoicacidmetabolite | C22H23NO4 | 401.13942 | 5.25 | 400.13214 | 3.72 × 10−6 | up | negative |
| 53 | Hexanoylcarnitine | C13H25NO4 | 259.17874 | 5.529 | 260.18602 | 3.97 × 10−6 | down | positive |
| 54 | 2-Butoxyaceticacid | C6H12O3 | 132.07877 | 5.546 | 131.07149 | 5.21 × 10−6 | down | negative |
| 55 | LPGO-20:1 | C26H53O8P | 524.34837 | 9.656 | 523.34109 | 5.42 × 10−6 | up | negative |
| 56 | (+/−)9-HpODE | C18H32O4 | 312.23068 | 7.109 | 311.2234 | 6.15 × 10−6 | up | negative |
| 57 | Betulin | C30H50O2 | 442.38096 | 10.323 | 443.38802 | 7.56 × 10−6 | up | positive |
| 58 | gamma-Glutamylleucine | C11H20N2O5 | 260.13759 | 5.363 | 259.13031 | 8.38 × 10−6 | up | negative |
| 59 | Phenylglyoxylicacid | C8H6O3 | 150.03189 | 5.84 | 301.07104 | 9.12 × 10−6 | up | positive |
| 60 | Formononetin | C16H12O4 | 268.07286 | 7.422 | 269.08017 | 9.19 × 10−6 | up | positive |
| 61 | Wogonin | C16H12O5 | 284.06782 | 6.827 | 285.07514 | 9.35 × 10−6 | up | positive |
| 62 | Meclocycline | C22H21ClN2O8 | 476.09669 | 6.087 | 475.08941 | 9.69 × 10−6 | up | negative |
| 63 | Hematoxylin | C16H14O6 | 302.07953 | 5.493 | 301.07225 | 1.03 × 10−5 | up | negative |
| 64 | 5,7-dihydroxy-6-methoxy-2-phenyl-3,4-dihydro-2H-1-benzopyran-4-one | C16H14O5 | 124.06416 | 5.93 | 287.09149 | 1.06 × 10−5 | up | positive |
| 65 | 4-Hydroxy-3-methoxyphenylglycolsulfate | C9H12O7S | 264.03101 | 4.801 | 263.02373 | 1.06 × 10−5 | up | negative |
| 66 | RKK | C18H38N8O4 | 408.32427 | 10.656 | 431.31357 | 1.13 × 10−5 | down | positive |
| 67 | Υ-Glutamylcysteine | C8H14N2O5S | 250.06246 | 1.994 | 251.06973 | 1.19 × 10−5 | up | positive |
| 68 | gamma-Glutamyltyrosine | C14H18N2O6 | 310.1163 | 4.941 | 311.12358 | 1.32 × 10−5 | up | positive |
| 69 | SPB15:0;2O | C15H33NO2 | 259.25039 | 6.784 | 260.25765 | 1.58 × 10−5 | down | positive |
| 70 | (6E,10E)-3,7,11,15-tetramethylhexadeca-1,6,10,14-tetraene-3,5,9-triol | C20H34O3 | 339.27689 | 8.98 | 362.26611 | 1.75 × 10−5 | up | positive |
| 71 | Benzyl6-O-beta-D-glucopyranosyl-beta-D-glucopyranoside | C19H28O11 | 478.16356 | 7.12 | 477.15628 | 1.8 × 10−5 | up | negative |
| 72 | 1-(4-hydroxyphenyl)propane-1,2-diol | C9H12O3 | 208.07353 | 8.381 | 191.07051 | 1.9 × 10−5 | up | positive |
| 73 | 23-Nordeoxycholicacid | C23H38O4 | 378.27712 | 9.48 | 401.26638 | 2.04 × 10−5 | down | positive |
| 74 | 18-β-Glycyrrhetinicacid | C30H46O4 | 470.3403 | 9.021 | 469.33302 | 2.1 × 10−5 | up | negative |
| 75 | Acetylcysteine | C5H9NO3S | 163.03053 | 1.535 | 146.02726 | 2.17 × 10−5 | up | positive |
| 76 | KKK | C18H38N6O4 | 384.28518 | 10.175 | 367.28192 | 2.2 × 10−5 | down | positive |
| 77 | Kynurenicacid | C10H7NO3 | 189.0429 | 2.602 | 190.05017 | 2.26 × 10−5 | down | positive |
| 78 | Dibutylsebacate | C18H34O4 | 314.24488 | 7.159 | 337.23407 | 2.46 × 10−5 | up | positive |
| 79 | 4-Oxoproline | C5H7NO3 | 129.04281 | 2.212 | 128.03553 | 2.66 × 10−5 | up | negative |
| 80 | Nonadecanoicacid | C19H38O2 | 298.28674 | 8.904 | 321.27596 | 2.69 × 10−5 | up | positive |
| 81 | (2R,3S,4S,5R,6R)-2-(hydroxymethyl)-6-(propan-2-yloxy)oxane-3,4,5-triol | C9H18O6 | 244.0882 | 5.68 | 245.09547 | 2.78 × 10−5 | up | positive |
| 82 | Tetranor-12(S)-HETE | C16H26O3 | 288.16699 | 7.738 | 289.17427 | 2.85 × 10−5 | down | positive |
| 83 | cyclohexyl{4-[4-nitro-2-(1H-pyrrol-1-yl)phenyl]piperazino}methanone | C21H26N4O3 | 404.18011 | 6.99 | 405.18737 | 2.98 × 10−5 | down | positive |
| 84 | N-Acetyl-L-cysteine | C5H9NO3S | 163.0306 | 3.649 | 164.03786 | 3.22 × 10−5 | up | positive |
| 85 | 1-Palmitoylglycerol | C19H38O4 | 330.27714 | 9.892 | 353.26637 | 3.46 × 10−5 | down | positive |
| 86 | Normorphine | C16H17NO3 | 271.12447 | 5.687 | 272.13175 | 3.69 × 10−5 | up | positive |
| 87 | DL-Lysine | C6H14N2O2 | 146.10571 | 1.213 | 147.11295 | 3.76 × 10−5 | down | positive |
| 88 | 23-Norcholicacid | C23H38O5 | 394.27029 | 10.275 | 393.26302 | 3.88 × 10−5 | up | negative |
| 89 | 2,3-dihydroxypropyl12-methyltridecanoate | C17H34O4 | 284.235 | 9.022 | 285.24228 | 4.19 × 10−5 | down | positive |
| 90 | Ethyl-β-D-glucuronide | C8H14O7 | 222.07419 | 2.183 | 221.06691 | 4.2 × 10−5 | down | negative |
| 91 | N-Acetylasparticacid | C6H9NO5 | 175.04819 | 1.64 | 174.04091 | 4.39 × 10−5 | up | negative |
| 92 | 6-(3-hydroxybutan-2-yl)-5-(hydroxymethyl)-4-methoxy-2H-pyran-2-one | C11H16O5 | 266.05726 | 6.879 | 267.06454 | 4.66 × 10−5 | up | positive |
| 93 | DL-4-Hydroxyphenyllacticacid | C9H10O4 | 200.06866 | 5.193 | 181.05081 | 4.73 × 10−5 | up | negative |
| 94 | Metanephrine | C10H15NO3 | 197.10607 | 5.366 | 198.11334 | 5.41 × 10−5 | up | positive |
| 95 | TPH | C15H23N5O5 | 335.15945 | 5.853 | 336.16667 | 5.66 × 10−5 | up | positive |
| 96 | 4-(2-((4-Cyanophenyl)amino)oxazol-5-yl)benzonitrile | C17H10N4O | 286.08461 | 5.45 | 285.07733 | 5.73 × 10−5 | up | negative |
| 97 | 3-Hydroxydecanoicacid | C10H20O3 | 188.14149 | 6.734 | 187.13421 | 5.85 × 10−5 | down | negative |
| 98 | Indole-3-lacticacid | C11H11NO3 | 205.07406 | 5.613 | 204.06678 | 6.04 × 10−5 | up | negative |
| 99 | Phenylpyruvicacid | C9H8O3 | 164.04762 | 5.193 | 163.04035 | 6.06 × 10−5 | up | negative |
| 100 | 1-Methylguanine | C6H7N5O | 165.06557 | 4.74 | 166.07284 | 6.59 × 10−5 | up | positive |
| 101 | (±)5(6)-DiHET | C20H34O4 | 320.23286 | 9.761 | 321.24006 | 7.25 × 10−5 | down | positive |
| 102 | Monoolein | C21H40O4 | 356.29294 | 10.106 | 379.28212 | 7.32 × 10−5 | down | positive |
| 103 | SM12:2;2O/8:0 | C25H49N2O6P | 550.33985 | 8.586 | 549.33257 | 8.46 × 10−5 | up | negative |
| 104 | L-Palmitoylcarnitine | C23H45NO4 | 399.33528 | 10.351 | 400.34256 | 9.52 × 10−5 | down | positive |
| 105 | Ureidosuccinicacid | C5H8N2O5 | 176.04359 | 1.49 | 175.03634 | 9.63 × 10−5 | up | negative |
| 106 | (+/−)11(12)-EET | C20H32O3 | 320.23546 | 8.216 | 319.22818 | 9.93 × 10−5 | up | negative |
| 107 | 1-[(3S)-3-(1,3-Benzoxazol-2-yl)-1-pyrrolidinyl]-3-methoxy-1-propanone | C15H18N2O3 | 274.13575 | 5.362 | 275.14301 | 0.000102 | up | positive |
| 108 | Estrone | C18H22O2 | 270.16277 | 4.84 | 271.17004 | 0.000105 | down | positive |
| 109 | L-(-)-Malicacid | C4H6O5 | 134.02165 | 1.651 | 133.01437 | 0.00011 | down | negative |
| 110 | 1,4-dihydroxyheptadec-16-en-2-ylacetate | C19H36O4 | 328.26148 | 9.25 | 351.25069 | 0.000113 | down | positive |
| 111 | 4-amino-6-chloro-3-cinnolinecarboxamide | C9H7ClN4O | 111.01374 | 1.589 | 223.03451 | 0.000117 | down | positive |
| 112 | Indoxylsulfuricacid | C8H7NO4S | 213.00984 | 5.195 | 212.00256 | 0.000122 | up | negative |
| 113 | WNH | C21H25N7O5 | 455.20205 | 4.721 | 456.20933 | 0.000128 | down | positive |
| 114 | 2-Arachidonoylglycerol | C23H38O4 | 378.27477 | 10.621 | 379.28204 | 0.000134 | down | positive |
| 115 | 12-Hydroxydodecanoicacid | C12H24O3 | 216.17279 | 6.982 | 215.16551 | 0.000136 | down | negative |
| 116 | Oleanolicacid | C30H48O3 | 478.34248 | 9.008 | 479.34976 | 0.000142 | up | positive |
| 117 | ProstaglandinF1β | C20H36O5 | 392.23359 | 11.278 | 391.22632 | 0.000156 | up | negative |
| 118 | DLK | C16H30N4O6 | 374.21719 | 2.92 | 188.11596 | 0.000158 | down | positive |
| 119 | N1-methyl-5-methoxy-2-({2-[(methylamino)carbonyl]phenyl}thio)benzamide | C17H18N2O3S | 366.08343 | 5.018 | 365.07616 | 0.000178 | down | negative |
| 120 | Stearamide | C18H37NO | 283.28694 | 7.68 | 284.29419 | 0.000179 | down | positive |
| 121 | NicotinuricAcid | C8H8N2O3 | 180.05369 | 1.69 | 179.04642 | 0.00018 | up | negative |
| 122 | o-Veratraldehyde | C9H10O3 | 166.0636 | 5.049 | 184.09743 | 0.000183 | up | positive |
| 123 | Epinephrine | C9H13NO3 | 183.09019 | 5.049 | 184.09747 | 0.000183 | up | positive |
| 124 | Tetracycline | C22H24N2O8 | 444.15789 | 8.14 | 443.15061 | 0.000201 | up | negative |
| 125 | CAR16:1 | C23H44NO4 | 397.31839 | 7.664 | 398.32567 | 0.000204 | down | positive |
| 126 | LPI14:0 | C23H45O12P | 544.26608 | 8.813 | 543.2588 | 0.000216 | up | negative |
| 127 | 2-Aminoadipicacid | C6H11NO4 | 161.06904 | 1.545 | 162.0763 | 0.000228 | down | positive |
| 128 | Histamine | C5H9N3 | 111.07998 | 1.515 | 112.08726 | 0.000244 | down | positive |
| 129 | 2,4-dihydroxyheptadec-16-en-1-ylacetate | C19H36O4 | 350.24308 | 9.094 | 351.25036 | 0.000253 | down | positive |
| 130 | S-Lactoyglutathione | C13H21N3O8S | 379.10574 | 4.814 | 380.11301 | 0.000256 | up | positive |
| 131 | Propionylcarnitine | C10H19NO4 | 217.13169 | 3.356 | 218.13897 | 0.000261 | down | positive |
| 132 | Octadecanamine | C18H39N | 269.30801 | 8.64 | 270.31528 | 0.000286 | up | positive |
| 133 | ProstaglandinA3 | C20H28O4 | 332.19935 | 6.942 | 331.19207 | 0.000287 | down | negative |
| 134 | Gamma-Glu-Leu | C11H20N2O5 | 260.13613 | 7.471 | 261.14341 | 0.00033 | down | positive |
| 135 | Dithranol | C14H10O3 | 226.06402 | 5.616 | 227.07091 | 0.000339 | up | positive |
| 136 | Lithocholicacid | C24H40O3 | 422.30162 | 11.084 | 421.29434 | 0.000342 | up | negative |
| 137 | N4-(4-chloro-2,5-dimethoxyphenyl)morpholine-4-carbothioamide | C13H17ClN2O3S | 354.01766 | 1.364 | 377.00681 | 0.000344 | down | positive |
| 138 | methyl3-(6-methylpyridin-2-yl)-2,2-diphenylpropanoate | C22H21NO2 | 331.15717 | 5.452 | 332.16445 | 0.000374 | up | positive |
| 139 | LPI16:1 | C25H47O12P | 570.28161 | 9.137 | 569.27433 | 0.000389 | up | negative |
| 140 | cis-Aconiticacid | C6H6O6 | 174.0166 | 1.963 | 173.00932 | 0.000393 | down | negative |
| 141 | 2-Methylbutyroylcarnitine | C12H23NO4 | 245.16328 | 5.709 | 244.15601 | 0.000404 | down | negative |
| 142 | EMK | C16H30N4O6S | 406.18623 | 5.072 | 407.19351 | 0.000408 | down | positive |
| 143 | 6-Deoxy-D-glucose | C6H12O5 | 164.06843 | 1.556 | 199.03783 | 0.000505 | down | negative |
| 144 | 2-hydroxy-3,6-diphenylcyclohexylacetate | C20H22O3 | 332.14082 | 5.747 | 333.14809 | 0.000512 | up | positive |
| 145 | All-Trans-13,14-Dihydroretinol | C20H32O | 288.24559 | 9.872 | 289.25277 | 0.000528 | down | positive |
| 146 | 5-Sulfosalicylicacid | C7H6O6S | 217.98878 | 4.813 | 216.98151 | 0.000533 | down | negative |
| 147 | N-Acetyl-1-aspartylglutamicacid | C11H16N2O8 | 304.09117 | 2.148 | 303.08399 | 0.000537 | up | negative |
| 148 | Androsterone | C19H30O2 | 272.21401 | 9.952 | 255.2108 | 0.000595 | up | positive |
| 149 | N-Acetylcysteine | C5H9NO3S | 163.0306 | 3.487 | 162.02332 | 0.000632 | up | negative |
| 150 | 2-Aminobenzenesulfonicacid | C6H7NO3S | 173.01534 | 1.708 | 191.0488 | 0.000672 | up | positive |
| 151 | N8-Acetylspermidine | C9H21N3O | 170.14228 | 1.359 | 188.17613 | 0.000734 | down | positive |
| 152 | Uridinemonophosphate(UMP) | C9H13N2O9P | 324.03615 | 1.734 | 323.02888 | 0.000761 | down | negative |
| 153 | Tetrahydrocorticosterone | C21H34O4 | 350.24428 | 8.29 | 349.237 | 0.000795 | down | negative |
| 154 | GLK | C14H28N4O4 | 316.21178 | 2.271 | 159.1132 | 0.000843 | down | positive |
| 155 | MAG(18:3) | C21H36O4 | 352.26118 | 9.05 | 375.25045 | 0.000877 | down | positive |
| 156 | Pantothenicacid | C9H17NO5 | 219.11078 | 5.05 | 218.10349 | 0.000979 | up | negative |
| 157 | 5-Phenyl-N-(4-(trifluoromethyl)phenyl)oxazol-2-amine | C16H11F3N2O | 304.07988 | 5.086 | 303.0726 | 0.001002 | down | negative |
| 158 | PB-22 N-(4-Hydroxypentyl)-3-carboxyindolemetabolite | C14H17NO3 | 229.11413 | 5.64 | 230.12141 | 0.001009 | up | positive |
| 159 | Abametapir | C12H12N2 | 184.10031 | 5.399 | 183.09304 | 0.00106 | up | negative |
| 160 | VitaminA | C20H30O | 286.22989 | 9.341 | 287.23712 | 0.001146 | down | positive |
| 161 | 3-Hydroxy-3-methylglutaricacid | C6H10O5 | 162.05305 | 3.244 | 161.04577 | 0.001209 | up | negative |
| 162 | 4-Hydroxyisoleucine | C6H13NO3 | 147.08976 | 5.048 | 146.08249 | 0.001217 | up | negative |
| 163 | 3-Oxo-7alpha,12alpha-hydroxy-5beta-cholanoicacid | C24H38O5 | 406.27176 | 8.655 | 429.26102 | 0.00127 | up | positive |
| 164 | Ozagrel | C13H12N2O2 | 228.09032 | 5.398 | 227.08311 | 0.001283 | up | negative |
| 165 | 13Z,16Z-DocosadienoicAcid | C22H40O2 | 336.30336 | 11.759 | 335.29608 | 0.001338 | down | negative |
| 166 | Daidzein | C15H10O4 | 254.05843 | 5.322 | 253.05115 | 0.001368 | up | negative |
| 167 | L-Asparticacid | C4H7NO4 | 133.03775 | 1.439 | 132.03047 | 0.001621 | down | negative |
| 168 | UDP-N-acetylglucosamine | C17H27N3O17P2 | 607.08328 | 2.08 | 606.07601 | 0.00163 | up | negative |
| 169 | CAR18:2 | C25H46NO4 | 423.33385 | 7.927 | 424.34113 | 0.001801 | down | positive |
| 170 | Docosanamide | C22H45NO | 339.34967 | 8.113 | 340.35695 | 0.001868 | up | positive |
| 171 | 3-hydroxy-3-methylpentanedioicacid | C6H10O5 | 179.08096 | 4.901 | 180.08824 | 0.001953 | down | positive |
| 172 | L-Malate | C4H6O5 | 134.0217 | 2.222 | 133.01442 | 0.00199 | down | negative |
| 173 | D-Fructose6-phosphate | C6H13O9P | 260.02923 | 5.552 | 259.02196 | 0.002423 | up | negative |
| 174 | Lysops22:6 | C28H44NO9P | 569.27426 | 8.875 | 568.26699 | 0.002662 | up | negative |
| 175 | CAR18:1 | C25H48NO4 | 425.35002 | 8.295 | 426.35731 | 0.002684 | down | positive |
| 176 | ProstaglandinE2 | C20H32O5 | 352.2256 | 6.444 | 351.21833 | 0.002936 | down | negative |
| 177 | DL-Malicacid | C4H6O5 | 134.02166 | 1.535 | 133.01437 | 0.002944 | down | negative |
| 178 | 15-OxoEDE | C20H34O3 | 322.25119 | 8.7 | 321.24391 | 0.003066 | down | negative |
| 179 | Ergosterol | C28H44O | 396.34013 | 9.972 | 397.3474 | 0.00308 | up | positive |
| 180 | 7-Methylguanosine | C11H15N5O5 | 297.10783 | 4.736 | 298.1151 | 0.003244 | up | positive |
| 181 | 5,8-dihydroxy-10-methyl-5,8,9,10-tetrahydro-2H-oxecin-2-one | C10H14O4 | 220.06773 | 5.938 | 463.12414 | 0.003308 | up | positive |
| 182 | 3-{[(4-fluorophenyl)sulfonyl]amino}-5-phenylthiophene-2-carboxamide | C17H13FN2O3S2 | 398.0202 | 5.67 | 399.02747 | 0.00339 | down | positive |
| 183 | Uridine5′-monophosphate | C9H13N2O9P | 324.03576 | 1.506 | 325.04304 | 0.003449 | down | positive |
| 184 | Ferulicacid | C10H10O4 | 194.05835 | 5.749 | 193.05107 | 0.003609 | up | negative |
| 185 | Palmitoylsphingomyelin | C39H79N2O6P | 702.56812 | 11.165 | 703.57542 | 0.003775 | down | positive |
| 186 | Tomatidine | C27H45NO2 | 415.34254 | 10.716 | 416.34982 | 0.003785 | down | positive |
| 187 | LPG18:3 | C24H43O9P | 506.26537 | 8.712 | 505.25809 | 0.003816 | up | negative |
| 188 | Fumaricacid | C4H4O4 | 116.01116 | 1.53 | 115.00388 | 0.003882 | down | negative |
| 189 | 5-OxoETE | C20H30O3 | 318.21584 | 8.573 | 319.22311 | 0.004151 | up | positive |
| 190 | PC32:1 | C40H78NO8P | 753.52888 | 11.487 | 754.53615 | 0.004193 | down | positive |
| 191 | LPH | C17H27N5O4 | 365.20767 | 4.864 | 183.61111 | 0.004262 | down | positive |
| 192 | N2-tetrahydrofuran-2-ylmethyl-4-(4-fluorophenyl)-1,3-thiazol-2-amine | C14H15FN2OS | 278.0942 | 4.887 | 279.10148 | 0.004346 | up | positive |
| 193 | 2,3-Dinor-8-epi-prostaglandinF2α | C18H30O5 | 326.20994 | 6.702 | 325.20266 | 0.004383 | down | negative |
| 194 | Xanthosine | C10H12N4O6 | 284.07609 | 4.58 | 283.06881 | 0.004546 | up | negative |
| 195 | Androsteroneglucuronide | C25H38O8 | 466.26151 | 11.762 | 465.25423 | 0.004653 | down | negative |
| 196 | ProstaglandinB2 | C20H30O4 | 334.21499 | 6.446 | 333.20775 | 0.00472 | down | negative |
| 197 | Pyridoxamine | C8H12N2O2 | 168.09007 | 1.709 | 167.0828 | 0.004866 | up | negative |
| 198 | LysoPC12:1 | C20H36NO7P | 433.22331 | 10.361 | 434.23059 | 0.005142 | up | positive |
| 199 | D-Sphingosine | C18H37NO2 | 281.27149 | 7.738 | 282.27877 | 0.005404 | down | positive |
| 200 | SPB19:1;2O | C19H39NO2 | 313.29764 | 7.958 | 296.29454 | 0.005472 | down | positive |
| 201 | 25-hydroxycholecalciferol | C27H44O2 | 400.33418 | 9.891 | 401.34147 | 0.005683 | up | positive |
| 202 | Tretinoin | C20H28O2 | 300.20928 | 9.096 | 299.20201 | 0.005714 | down | negative |
| 203 | SPB16:1;2O | C16H33NO2 | 271.25058 | 7.139 | 254.24738 | 0.00605 | down | positive |
| 204 | (±)7(8)-DiHDPA | C22H34O4 | 362.24388 | 10.536 | 361.23661 | 0.006437 | down | negative |
| 205 | LPE22:1 | C27H54NO7P | 535.36437 | 11.178 | 534.35709 | 0.006455 | down | negative |
| 206 | Palmitoylethanolamide | C18H37NO2 | 299.28202 | 7.513 | 300.28935 | 0.006565 | down | positive |
| 207 | L-Ascorbate | C6H8O6 | 176.03236 | 2.671 | 175.02506 | 0.006667 | up | negative |
| 208 | AICAribonucleotide | C9H15N4O8P | 300.10741 | 1.504 | 337.05604 | 0.006702 | up | negative |
| 209 | dTMP | C10H15N2O8P | 322.05653 | 2.247 | 323.06429 | 0.006993 | down | positive |
| 210 | Nervonicacid | C24H46O2 | 366.3503 | 11.682 | 365.34302 | 0.007003 | down | negative |
| 211 | XMP | C10H13N4O9P | 364.04263 | 1.528 | 363.03535 | 0.007068 | up | negative |
| 212 | Tauroursodeoxycholicacid | C26H45NO6S | 499.29183 | 8.037 | 498.28455 | 0.007212 | up | negative |
| 213 | Guanine | C5H5N5O | 151.04966 | 1.669 | 152.05694 | 0.007274 | down | positive |
| 214 | Creatinephosphate | C4H10N3O5P | 211.03577 | 1.486 | 212.04289 | 0.007724 | down | positive |
| 215 | 13,14-Dihydro-15-ketoProstaglandinE2 | C20H32O5 | 352.22891 | 9.338 | 351.22163 | 0.007811 | down | negative |
| 216 | UDP-galactose | C15H24N2O17P2 | 566.05562 | 1.835 | 565.04834 | 0.008 | down | negative |
| 217 | 2-phenyl-2,4,6,7-tetrahydrothiino[4,3-c]pyrazol-3-ol | C12H12N2OS | 232.06915 | 5.845 | 233.07651 | 0.008108 | down | positive |
| 218 | 4-Methyl-5-thiazoleethanol | C6H9NOS | 143.04103 | 4.82 | 144.04831 | 0.008803 | up | positive |
| 219 | LPC18:3 | C26H48NO7P | 563.32363 | 8.55 | 562.31635 | 0.008812 | up | negative |
| 220 | Phosphoethanolamine | C2H8NO4P | 141.01938 | 3.489 | 140.0121 | 0.008994 | down | negative |
| 221 | Deoxyribose5-Phosphate | C5H11O7P | 214.02458 | 1.465 | 259.02277 | 0.009015 | down | negative |
| 222 | LPI18:1 | C27H51O12P | 598.31322 | 10.219 | 597.30594 | 0.009165 | up | negative |
| 223 | LPK | C17H32N4O4 | 356.24351 | 4.797 | 179.12907 | 0.009173 | down | positive |
| 224 | LPE22:5 | C27H46NO7P | 527.30214 | 9.02 | 526.29487 | 0.009643 | down | negative |
| 225 | Palmitoylcarnitine | C23H45NO4 | 399.33451 | 8.128 | 400.3418 | 0.009796 | down | positive |
| 226 | 2-chloro-6-(1,4-thiazinan-4-yl)benzonitrile | C11H11ClN2S | 260.0147 | 5.936 | 261.02197 | 0.009826 | down | positive |
| 227 | dCDP | C9H15N3O10P2 | 387.02491 | 1.804 | 386.01764 | 0.010204 | down | negative |
| 228 | S-Lactoylglutathione | C13H21N3O8S | 379.10423 | 3.572 | 380.11151 | 0.010466 | down | positive |
| 229 | LPI17:1 | C26H49O12P | 584.29753 | 9.838 | 583.29026 | 0.010797 | up | negative |
| 230 | Cholesterol | C27H46O | 386.3545 | 11.396 | 387.36161 | 0.011521 | down | positive |
| 231 | Gly-Tyr | C11H14N2O4 | 238.09573 | 4.39 | 239.10301 | 0.012216 | down | positive |
| 232 | Cholest-4-en-3-one | C27H44O | 384.33957 | 11.308 | 385.34707 | 0.01228 | down | positive |
| 233 | D-Glucose6-phosphate | C6H13O9P | 260.03008 | 1.24 | 259.0228 | 0.012542 | down | negative |
| 234 | morphine-d3 | C17H16[2]H3NO3 | 271.12449 | 5.532 | 272.132 | 0.012548 | up | positive |
| 235 | Dl-Glyceraldehyde3-phosphate | C3H7O6P | 169.99808 | 1.412 | 192.98732 | 0.012554 | down | positive |
| 236 | 16(R)-HETE | C20H32O3 | 342.21464 | 9.446 | 343.22191 | 0.013386 | up | positive |
| 237 | SM20:2;2O | C25H49N2O6P | 526.31483 | 8.601 | 527.32211 | 0.013592 | up | positive |
| 238 | 3-{[(3S)-3-(1,3-Benzothiazol-2-yl)-1-pyrrolidinyl]methyl}benzonitrile | C19H17N3S | 319.11752 | 5.325 | 320.12478 | 0.013719 | down | positive |
| 239 | Mupirocin | C26H44O9 | 1044.56464 | 5.758 | 523.2896 | 0.013802 | down | positive |
| 240 | S-(Methyl)Glutathione | C11H19N3O6S | 321.09974 | 2.685 | 322.10702 | 0.013887 | up | positive |
| 241 | L-Arabinitol | C5H12O5 | 152.06881 | 1.406 | 151.0615 | 0.014314 | up | negative |
| 242 | gamma-Glutamylmethionine | C10H18N2O5S | 278.09426 | 4.877 | 277.08699 | 0.014572 | up | negative |
| 243 | PC16:0_16:1 | C40H78NO8P | 731.54681 | 11.524 | 732.55404 | 0.014578 | down | positive |
| 244 | 10-Nitrolinoleate | C18H31NO4 | 325.22584 | 6.354 | 324.21857 | 0.014608 | down | negative |
| 245 | Uridine5′-diphosphoglucuronicacid | C15H22N2O18P2 | 580.03593 | 0.936 | 579.02866 | 0.014748 | up | negative |
| 246 | 13-HPODE | C18H32O4 | 312.22892 | 7.104 | 313.236 | 0.015141 | up | positive |
| 247 | PE16:1_18:1 | C39H74NO8P | 715.51665 | 8.794 | 714.50937 | 0.016297 | down | negative |
| 248 | 4-(Diethylamino)salicylaldehyde | C11H15NO2 | 193.10845 | 5.563 | 194.11573 | 0.016762 | down | positive |
| 249 | Docosapentaenoicacid | C22H34O2 | 330.25621 | 10.186 | 329.24893 | 0.017709 | down | negative |
| 250 | Aminomalonicacid | C3H5NO4 | 119.02213 | 2.263 | 118.01486 | 0.018361 | up | negative |
| 251 | WMH | C22H28N6O4S | 472.19233 | 5.707 | 473.19961 | 0.018551 | down | positive |
| 252 | 3-(2,3-dihydro-1H-indol-1-yl)-2-[(2-furylmethyl)sulfonyl]acrylonitrile | C16H14N2O3S | 314.07222 | 1.401 | 315.0795 | 0.018716 | down | positive |
| 253 | 2-Phenylacetamide | C8H9NO | 135.06561 | 5.367 | 309.09378 | 0.019606 | up | positive |
| 254 | Pizotifen | C19H21NS | 295.1459 | 5.854 | 294.13862 | 0.02024 | down | negative |
| 255 | 4-acetyl-4-(ethoxycarbonyl)heptanedioicacid | C12H18O7 | 296.08372 | 5.655 | 297.09104 | 0.020248 | down | positive |
| 256 | LPEO-18:1 | C23H48NO6P | 465.32294 | 10.063 | 464.31566 | 0.020855 | up | negative |
| 257 | LPC14:1 | C22H44NO7P | 511.29184 | 7.885 | 510.28456 | 0.021171 | up | negative |
| 258 | methyl6-{[4-(trifluoromethyl)anilino]carbonyl}nicotinate | C15H11F3N2O3 | 306.05724 | 4.518 | 307.06451 | 0.021742 | up | positive |
| 259 | Cortodoxone | C21H30O4 | 346.21299 | 9.662 | 345.20571 | 0.023107 | down | negative |
| 260 | β-Nicotinamidemononucleotide | C11H15N2O8P | 334.05636 | 1.47 | 335.06363 | 0.023125 | down | positive |
| 261 | N-Acetyl-L-phenylalanine | C11H13NO3 | 207.08986 | 5.653 | 206.08258 | 0.025062 | up | negative |
| 262 | Rosuvastatin | C22H28FN3O6S | 481.1694 | 4.899 | 480.16212 | 0.025277 | down | negative |
| 263 | 5-(6-hydroxy-6-methyloctyl)-2,5-dihydrofuran-2-one | C13H22O3 | 248.13969 | 6.019 | 249.14696 | 0.025292 | down | positive |
| 264 | Pyrogallol | C6H6O3 | 126.03191 | 0.422 | 127.03918 | 0.025859 | down | positive |
| 265 | 2-[5-(2-hydroxypropyl)oxolan-2-yl]propanoicacid | C10H18O4 | 240.07218 | 2.532 | 241.07956 | 0.025902 | down | positive |
| 266 | Pantetheine | C11H22N2O4S | 278.13077 | 5.181 | 277.12306 | 0.026244 | down | negative |
| 267 | GTP | C10H16N5O14P3 | 522.99019 | 1.585 | 521.98291 | 0.026991 | down | negative |
| 268 | LPS16:1 | C22H42NO9P | 495.26066 | 8.855 | 494.25338 | 0.027007 | up | negative |
| 269 | N-Acetyl-L-leucine | C8H15NO3 | 173.10543 | 2.098 | 174.11271 | 0.027241 | down | positive |
| 270 | LPE20:5 | C25H42NO7P | 499.27084 | 8.511 | 498.26356 | 0.028403 | down | negative |
| 271 | D-Mannose6-phosphate | C6H13O9P | 278.04068 | 1.454 | 259.02284 | 0.028568 | down | negative |
| 272 | (±)9-HpODE | C18H32O4 | 312.23009 | 10.662 | 311.22281 | 0.028898 | down | negative |
| 273 | 2,3-DinorprostaglandinE1 | C18H30O5 | 308.19694 | 7.269 | 307.18967 | 0.029444 | up | negative |
| 274 | CAR18:0 | C25H50NO4 | 427.36593 | 8.758 | 428.3732 | 0.029497 | down | positive |
| 275 | (2E,4E)-N-(2-methylpropyl)dodeca-2,4-dienamide | C16H29NO | 251.22467 | 8.565 | 252.23213 | 0.030263 | down | positive |
| 276 | (5-L-Glutamyl)-L-AminoAcid | C8H14N2O5 | 218.0905 | 1.757 | 217.08322 | 0.030331 | down | negative |
| 277 | Lysopc16:1 | C24H48NO7P | 493.31792 | 9.154 | 492.31064 | 0.030434 | up | negative |
| 278 | D-Homocysteine | C4H9NO2S | 135.0356 | 1.539 | 136.04286 | 0.030734 | up | positive |
| 279 | UMP | C9H13N2O9P | 324.03662 | 1.633 | 347.02603 | 0.031106 | down | positive |
| 280 | Corticosterone | C21H30O4 | 346.21876 | 10.906 | 347.22594 | 0.032826 | down | positive |
| 281 | 2-(3,4-dimethoxyphenyl)ethanamine | C10H15NO2 | 181.1107 | 5.7 | 226.1089 | 0.033212 | up | negative |
| 282 | CAR20:1 | C27H52NO4 | 453.38151 | 8.903 | 454.38877 | 0.033615 | down | positive |
| 283 | 2-hydroxy-6-[(8Z,11Z)-pentadeca-8,11,14-trien-1-yl]benzoicacid | C22H30O3 | 324.2042 | 10.077 | 325.21148 | 0.034219 | down | positive |
| 284 | N-[1-(4-methoxy-2-oxo-2H-pyran-6-yl)-2-methylbutyl]acetamide | C13H19NO4 | 270.15711 | 8.886 | 271.16438 | 0.035764 | down | positive |
| 285 | 5-Methyl-2′-deoxycytidine | C10H15N3O4 | 241.10658 | 4.966 | 240.0993 | 0.036121 | down | negative |
| 286 | 2-Mercaptobenzothiazole | C7H5NS2 | 166.98653 | 6.003 | 165.97926 | 0.036641 | down | negative |
| 287 | 11(Z),14(Z),17(Z)-Eicosatrienoicacid | C20H34O2 | 306.2558 | 10.384 | 307.26317 | 0.0368 | down | positive |
| 288 | N-Formylkynurenine | C11H12N2O4 | 236.08032 | 4.893 | 237.08759 | 0.037249 | down | positive |
| 289 | ThromoboxaneB1 | C20H36O6 | 408.22837 | 8.876 | 407.22109 | 0.038568 | up | negative |
| 290 | diethyl2-{[2-(2-thienylcarbonyl)hydrazino]methylidene}malonate | C13H16N2O5S | 350.03433 | 4.698 | 351.0416 | 0.040171 | up | positive |
| 291 | LPG20:2 | C26H49O9P | 536.3124 | 11.26 | 535.30513 | 0.040667 | down | negative |
| 292 | PEO-16:1_20:4 | C41H74NO7P | 723.52229 | 10.984 | 722.51501 | 0.042265 | down | negative |
| 293 | α-Aspartylphenylalanine | C13H16N2O5 | 280.10644 | 5.298 | 281.11372 | 0.042952 | down | positive |
| 294 | PEO-16:1_16:1 | C37H72NO7P | 673.50663 | 10.413 | 672.49935 | 0.04305 | down | negative |
| 295 | 2,4-Dinitrophenol | C6H4N2O5 | 184.01216 | 5.911 | 183.00488 | 0.043494 | down | negative |
| 296 | Caffeine | C8H10N4O2 | 194.08059 | 5.35 | 195.08787 | 0.04419 | down | positive |
| 297 | LPE20:1 | C25H50NO7P | 507.3332 | 10.382 | 506.32592 | 0.045014 | down | negative |
| 298 | Adrenicacid | C22H36O2 | 332.27176 | 10.665 | 331.26449 | 0.045049 | down | negative |
| 299 | 1a,1b-DihomoprostaglandinF2α | C22H38O5 | 418.24932 | 11.807 | 417.24204 | 0.045094 | up | negative |
| 300 | Adenosinetriphosphate(ATP) | C10H16N5O13P3 | 506.99653 | 1.555 | 505.98926 | 0.045309 | down | negative |
| 301 | PC15:0_15:1 | C38H74NO8P | 703.51539 | 11.764 | 704.52275 | 0.047494 | down | positive |
| 302 | 6-(2-furyl)-2-hydroxy-4-(2-thienyl)nicotinonitrile | C14H8N2O2S | 268.02827 | 5.044 | 267.02097 | 0.047838 | up | negative |
| 303 | Lysopa18:0 | C21H43O7P | 438.27514 | 11.05 | 437.26786 | 0.048005 | down | negative |
| Pathway | p Value | Differential Metabolites | Polarity Ion Mode |
|---|---|---|---|
| Steroid hormone bio-synthesis | 0.002248 | Tetrahydrocorticosterone; Cortodoxone; Hydrocortisone; Andros-terone glucuronide; Adrenosterone | Negative |
| Steroid hormone bio-synthesis | 0.038973 | Cholesterol; Androsterone; Estrone; Dehydroepiandrosterone (DHEA); Corticosterone | Positive |
| Linoleic acid metabo-lism | 0.044694 | PC 32:1; (+/−)12(13)-DiHOME; 13-HPODE | Positive |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Cheng, L.; Li, J.; Wang, Y.; Chen, S.; Wang, Z.; Yang, W. Potential Antitumor Mechanism of Propolis Against Skin Squamous Cell Carcinoma A431 Cells Based on Untargeted Metabolomics. Int. J. Mol. Sci. 2024, 25, 11265. https://doi.org/10.3390/ijms252011265
Wang J, Cheng L, Li J, Wang Y, Chen S, Wang Z, Yang W. Potential Antitumor Mechanism of Propolis Against Skin Squamous Cell Carcinoma A431 Cells Based on Untargeted Metabolomics. International Journal of Molecular Sciences. 2024; 25(20):11265. https://doi.org/10.3390/ijms252011265
Chicago/Turabian StyleWang, Jie, Liyuan Cheng, Jingjing Li, Yicong Wang, Siyuan Chen, Zhongdan Wang, and Wenchao Yang. 2024. "Potential Antitumor Mechanism of Propolis Against Skin Squamous Cell Carcinoma A431 Cells Based on Untargeted Metabolomics" International Journal of Molecular Sciences 25, no. 20: 11265. https://doi.org/10.3390/ijms252011265
APA StyleWang, J., Cheng, L., Li, J., Wang, Y., Chen, S., Wang, Z., & Yang, W. (2024). Potential Antitumor Mechanism of Propolis Against Skin Squamous Cell Carcinoma A431 Cells Based on Untargeted Metabolomics. International Journal of Molecular Sciences, 25(20), 11265. https://doi.org/10.3390/ijms252011265