
The Abundant Distribution and Duplication of SARS-CoV-2 in the Cerebrum and Lungs Promote a High Mortality Rate in Transgenic hACE2-C57 Mice

 ,
, 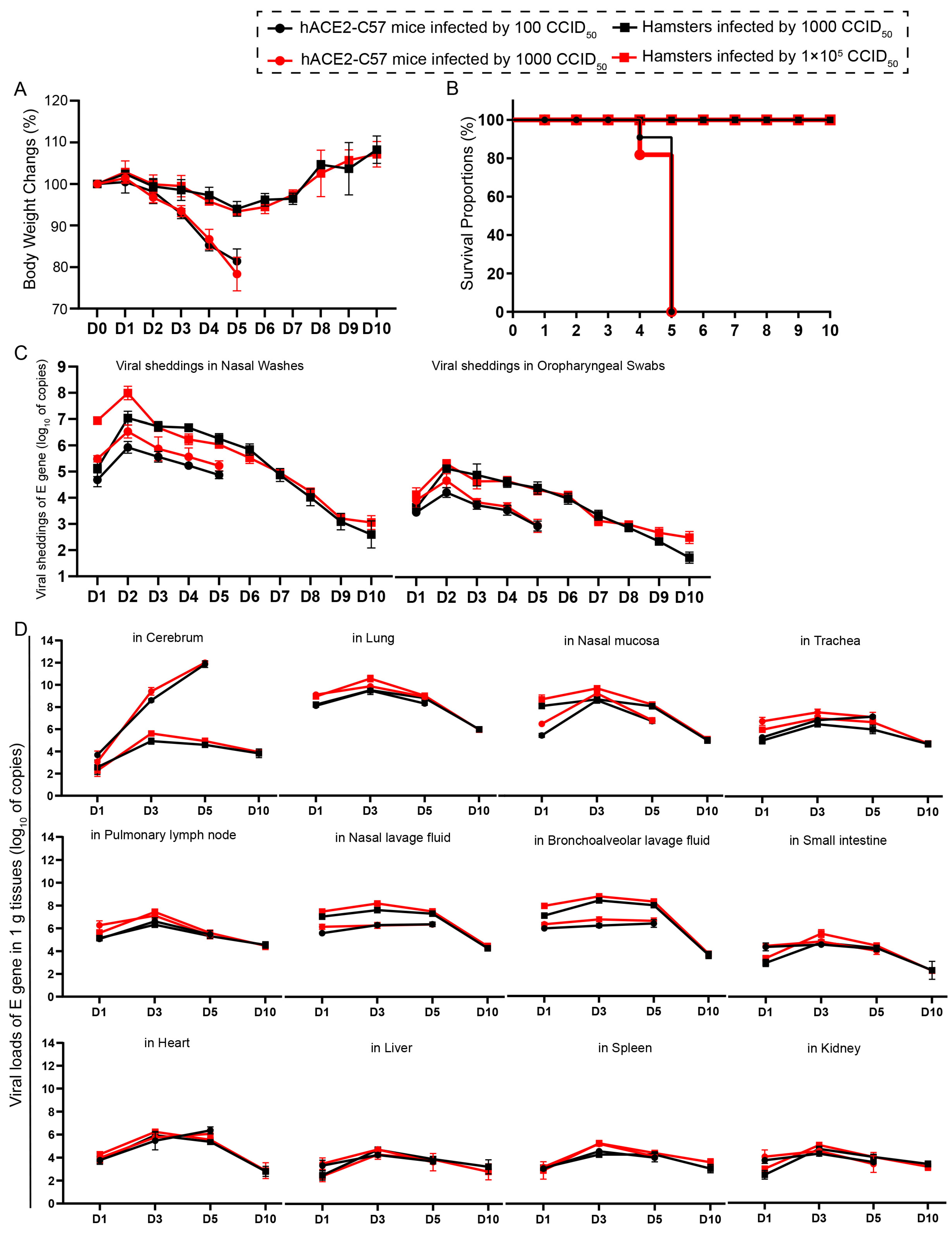{kind=link}
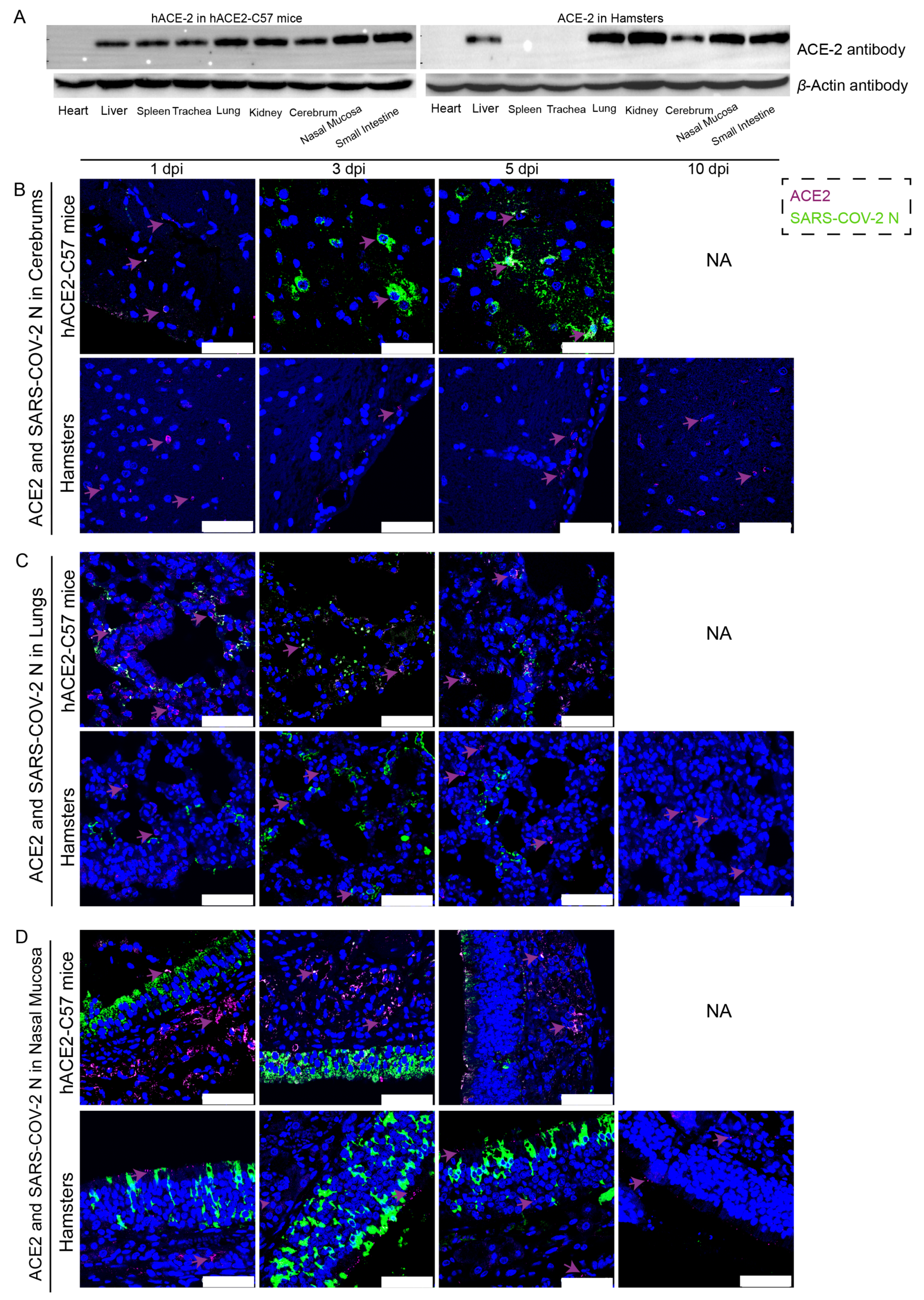{kind=link}
{kind=link}
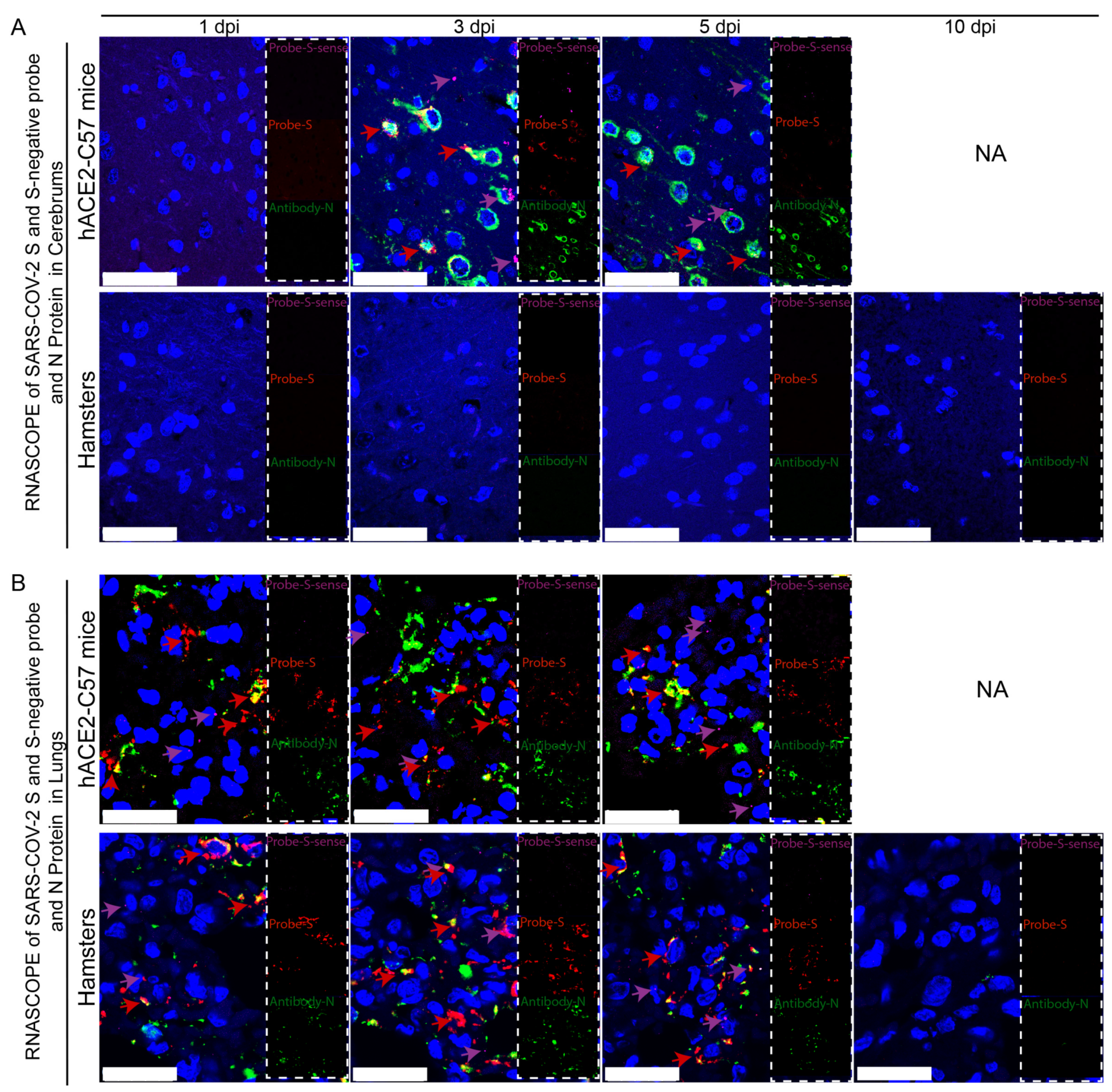{kind=link}
{kind=link}
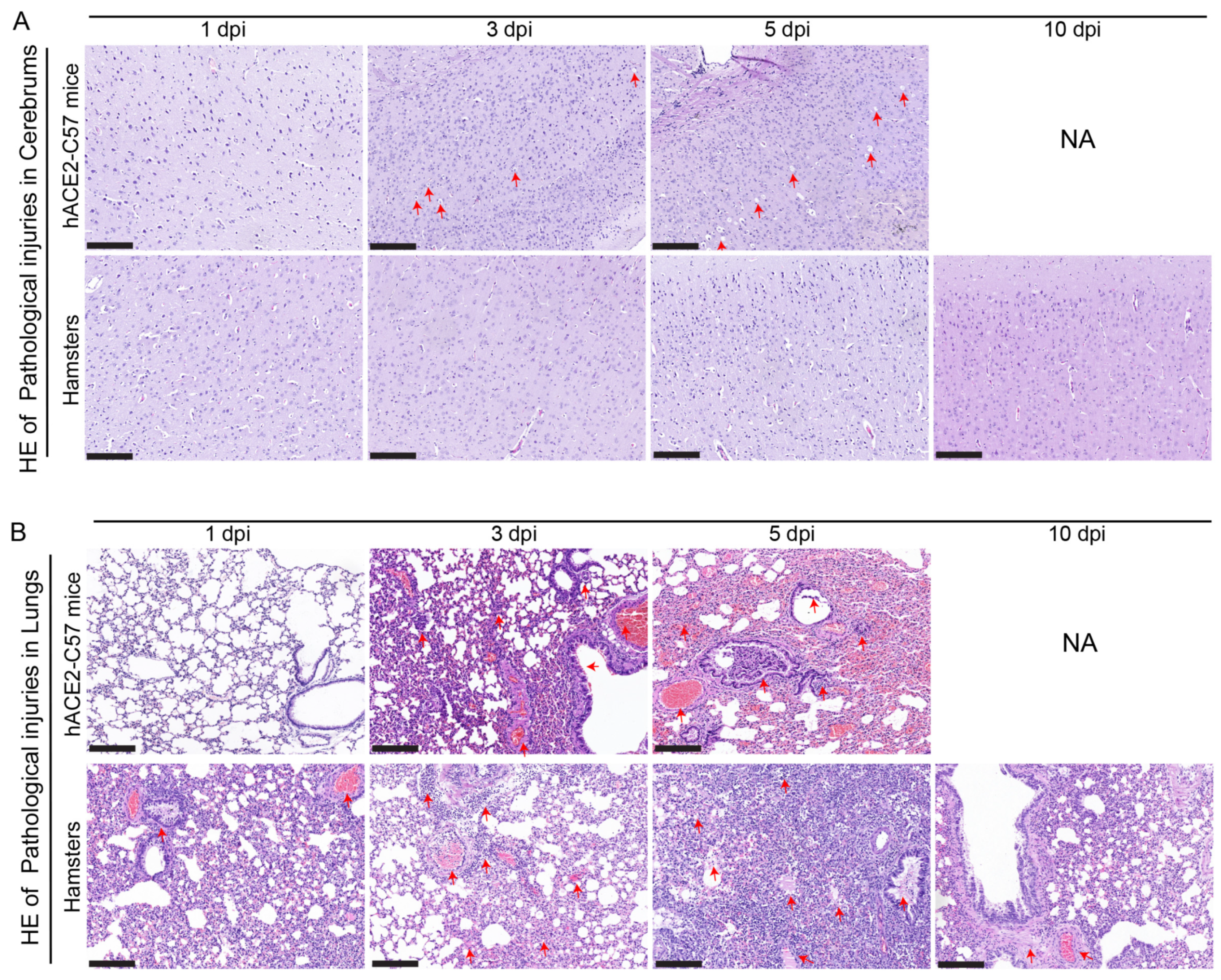{kind=link}
Abstract
1. Introduction
2. Results
2.1. Both hACE2-C57 Mice and Hamsters Were Susceptible to SARS-CoV-2 after Intranasal Infection, Which Resulted in Complete Fatality with High Viral Loads in the Cerebrums of hACE2-C57 Mice
2.2. ACE2 Was Expressed in Most Internal Organs of Both hACE2-C57 Mice and Hamsters, and the SARS-CoV-2 Viral N Protein Was Distributed in the Cerebrum of hACE2-C57 Mice but Not in That of Hamsters
2.3. SARS-CoV-2 Was Mainly Distributed in the Neurons of the Cerebral Cortex of hACE2-C57 Mice, While the Virus Was Widely Detected in Alveolar Cells in the Lungs of Both hACE2-C57 Mice and Hamsters
2.4. Significant Viral Replication and a Strong Viral Load Were Observed in Both the Cerebrums and Lungs of hACE2-C57 Mice, While Abundant Viral Replication Was Present in the Lungs but Not the Cerebrum of Hamsters
2.5. Obvious Increases in IL-1β and Minimal Changes in IFN-γ Were Observed in the Cerebrum of hACE2-C57 Mice Infected with SARS-CoV-2
2.6. Prominent Pathological Injuries Were Observed in the Cerebrum of hACE2-C57 Mice, While Both hACE2-C57 Mice and Hamsters Exhibited Severe Interstitial Pneumonia following SARS-CoV-2 Infection
3. Discussion
4. Materials and Methods
4.1. Animals and Biosafety
4.2. Viruses
4.3. Virus Load Detection
4.4. Western Blotting
4.5. Immunofluorescence (IF)
4.5.1. Determination of the Distribution and Abundance of ACE2 and SARS-CoV-2 via Immunofluorescence
4.5.2. Determination of the Distribution and Abundance of IFN-γ and IL-1β via Immunofluorescence
4.6. Immunohistochemistry (IHC)
4.7. RNAscope-I Codetection
4.8. Histopathology
4.9. IFN-γ and IL-1β Quantifications via RT-Real Time PCR
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ong, W.Y.; Satish, R.L.; Herr, D.R. ACE2, Circumventricular Organs and the Hypothalamus, and COVID-19. NeuroMolecular Med. 2022, 24, 363–373. [Google Scholar] [CrossRef]
- Conde, C.G.; Quintana, P.L.; Quintero, M.I.; Ramos, V.Y.; Moscote, S.L. Neurotropism of SARS-CoV-2: Mechanisms and manifestations. J. Neurol. Sci. 2020, 412, 116824. [Google Scholar] [CrossRef] [PubMed]
- Mao, L.; Jin, H.; Wang, M.; Hu, Y.; Chen, S.; He, Q.; Chang, J.; Hong, C.; Zhou, Y.; Wang, D.; et al. Neurologic Manifestations of Hospitalized Patients with Coronavirus Disease 2019 in Wuhan, China. JAMA Neurol. 2020, 77, 683–690. [Google Scholar] [CrossRef]
- Graham, E.L.; Clark, J.R.; Orban, Z.S.; Lim, P.H.; Szymanski, A.L.; Taylor, C.; DiBiase, R.M.; Jia, D.T.; Balabanov, R.; Ho, S.U.; et al. Persistent neurologic symptoms and cognitive dysfunction in non-hospitalized COVID-19 “long haulers”. Ann. Clin. Transl. Neurol. 2021, 8, 1073–1085. [Google Scholar] [CrossRef] [PubMed]
- Meinhardt, J.; Radke, J.; Dittmayer, C.; Franz, J.; Thomas, C.; Mothes, R.; Laue, M.; Schneider, J.; Brunink, S.; Greuel, S.; et al. Olfactory transmucosal SARS-CoV-2 invasion as a port of central nervous system entry in individuals with COVID-19. Nat. Neurosci. 2021, 24, 168–175. [Google Scholar] [CrossRef]
- Zheng, M. ACE2 and COVID-19 Susceptibility and Severity. Aging Dis. 2022, 13, 360–372. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M. Cellular Tropism of SARS-CoV-2 across Human Tissues and Age-related Expression of ACE2 and TMPRSS2 in Immune-inflammatory Stromal Cells. Aging Dis. 2021, 12, 718–725. [Google Scholar] [CrossRef]
- Ren, W.; Zhu, Y.; Lan, J.; Chen, H.; Wang, Y.; Shi, H.; Feng, F.; Chen, D.Y.; Close, B.; Zhao, X.; et al. Susceptibilities of Human ACE2 Genetic Variants in Coronavirus Infection. J. Virol. 2022, 96, e149221. [Google Scholar] [CrossRef]
- Viana, R.; Moyo, S.; Amoako, D.G.; Tegally, H.; Scheepers, C.; Althaus, C.L.; Anyaneji, U.J.; Bester, P.A.; Boni, M.F.; Chand, M.; et al. Rapid epidemic expansion of the SARS-CoV-2 Omicron variant in southern Africa. Nature 2022, 603, 679–686. [Google Scholar] [CrossRef]
- Yamasoba, D.; Kimura, I.; Nasser, H.; Morioka, Y.; Nao, N.; Ito, J.; Uriu, K.; Tsuda, M.; Zahradnik, J.; Shirakawa, K.; et al. Virological characteristics of the SARS-CoV-2 Omicron BA.2 spike. Cell 2022, 185, 2103–2115. [Google Scholar] [CrossRef]
- Halfmann, P.J.; Iida, S.; Iwatsuki-Horimoto, K.; Maemura, T.; Kiso, M.; Scheaffer, S.M.; Darling, T.L.; Joshi, A.; Loeber, S.; Singh, G.; et al. SARS-CoV-2 Omicron virus causes attenuated disease in mice and hamsters. Nature 2022, 603, 687–692. [Google Scholar] [CrossRef]
- Uraki, R.; Kiso, M.; Iida, S.; Imai, M.; Takashita, E.; Kuroda, M.; Halfmann, P.J.; Loeber, S.; Maemura, T.; Yamayoshi, S.; et al. Characterization and antiviral susceptibility of SARS-CoV-2 Omicron BA.2. Nature 2022, 607, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.; Liu, P.; Wang, N.; Wang, L.; Fan, K.; Zhu, Q.; Wang, K.; Chen, R.; Feng, R.; Jia, Z.; et al. Structural and functional characterizations of infectivity and immune evasion of SARS-CoV-2 Omicron. Cell 2022, 185, 860–871. [Google Scholar] [CrossRef] [PubMed]
- Han, P.; Li, L.; Liu, S.; Wang, Q.; Zhang, D.; Xu, Z.; Han, P.; Li, X.; Peng, Q.; Su, C.; et al. Receptor binding and complex structures of human ACE2 to spike RBD from omicron and delta SARS-CoV-2. Cell 2022, 185, 630–640. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Gomes, J. Neuropathogenesis of SARS-CoV-2 infection. eLife 2020, 9, e59136. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, Y.; Luo, W.; Huang, L.; Xiao, J.; Li, F.; Qin, S.; Song, X.; Wu, Y.; Zeng, Q.; et al. A comprehensive investigation of the mRNA and protein level of ACE2, the putative receptor of SARS-CoV-2, in human tissues and blood cells. Int. J. Med. Sci. 2020, 17, 1522–1531. [Google Scholar] [CrossRef] [PubMed]
- Li, M.Y.; Li, L.; Zhang, Y.; Wang, X.S. Expression of the SARS-CoV-2 cell receptor gene ACE2 in a wide variety of human tissues. Infect. Dis. Poverty 2020, 9, 45. [Google Scholar] [CrossRef]
- Golden, J.W.; Cline, C.R.; Zeng, X.; Garrison, A.R.; Carey, B.D.; Mucker, E.M.; White, L.E.; Shamblin, J.D.; Brocato, R.L.; Liu, J.; et al. Human angiotensin-converting enzyme 2 transgenic mice infected with SARS-CoV-2 develop severe and fatal respiratory disease. JCI Insight 2020, 5, e142032. [Google Scholar] [CrossRef]
- Jiang, R.D.; Liu, M.Q.; Chen, Y.; Shan, C.; Zhou, Y.W.; Shen, X.R.; Li, Q.; Zhang, L.; Zhu, Y.; Si, H.R.; et al. Pathogenesis of SARS-CoV-2 in Transgenic Mice Expressing Human Angiotensin-Converting Enzyme 2. Cell 2020, 182, 50–58. [Google Scholar] [CrossRef]
- Sun, S.H.; Chen, Q.; Gu, H.J.; Yang, G.; Wang, Y.X.; Huang, X.Y.; Liu, S.S.; Zhang, N.N.; Li, X.F.; Xiong, R.; et al. A Mouse Model of SARS-CoV-2 Infection and Pathogenesis. Cell Host Microbe 2020, 28, 124–133. [Google Scholar] [CrossRef]
- Chan, J.F.; Zhang, A.J.; Yuan, S.; Poon, V.K.; Chan, C.C.; Lee, A.C.; Chan, W.M.; Fan, Z.; Tsoi, H.W.; Wen, L.; et al. Simulation of the Clinical and Pathological Manifestations of Coronavirus Disease 2019 (COVID-19) in a Golden Syrian Hamster Model: Implications for Disease Pathogenesis and Transmissibility. Clin. Infect. Dis. 2020, 71, 2428–2446. [Google Scholar] [CrossRef] [PubMed]
- Damas, J.; Hughes, G.M.; Keough, K.C.; Painter, C.A.; Persky, N.S.; Corbo, M.; Hiller, M.; Koepfli, K.P.; Pfenning, A.R.; Zhao, H.; et al. Broad host range of SARS-CoV-2 predicted by comparative and structural analysis of ACE2 in vertebrates. Proc. Natl. Acad. Sci. USA 2020, 117, 22311–22322. [Google Scholar] [CrossRef] [PubMed]
- Suresh, V.; Parida, D.; Minz, A.P.; Sethi, M.; Sahoo, B.S.; Senapati, S. Tissue Distribution of ACE2 Protein in Syrian Golden Hamster (Mesocricetus auratus) and Its Possible Implications in SARS-CoV-2 Related Studies. Front. Pharmacol. 2020, 11, 579330. [Google Scholar] [CrossRef] [PubMed]
- Sia, S.F.; Yan, L.M.; Chin, A.; Fung, K.; Choy, K.T.; Wong, A.; Kaewpreedee, P.; Perera, R.; Poon, L.; Nicholls, J.M.; et al. Pathogenesis and transmission of SARS-CoV-2 in golden hamsters. Nature 2020, 583, 834–838. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhao, X.; Li, J.; Zheng, H.; Zhao, Y.; Yang, J.; Zhou, J.; Yang, F.; Chen, Y.; Zuo, Y.; et al. Nasal Mucosa Exploited by SARS-CoV-2 for Replicating and Shedding during Reinfection. Viruses 2022, 14, 1608. [Google Scholar] [CrossRef]
- Hong, N.; Yu, W.; Xia, J.; Shen, Y.; Yap, M.; Han, W. Evaluation of ocular symptoms and tropism of SARS-CoV-2 in patients confirmed with COVID-19. Acta Ophthalmol. 2020, 98, e649–e655. [Google Scholar] [CrossRef]
- Jin, X.; Lian, J.S.; Hu, J.H.; Gao, J.; Zheng, L.; Zhang, Y.M.; Hao, S.R.; Jia, H.Y.; Cai, H.; Zhang, X.L.; et al. Epidemiological, clinical and virological characteristics of 74 cases of coronavirus-infected disease 2019 (COVID-19) with gastrointestinal symptoms. Gut 2020, 69, 1002–1009. [Google Scholar] [CrossRef]
- Cleary, S.J.; Pitchford, S.C.; Amison, R.T.; Carrington, R.; Robaina, C.C.; Magnen, M.; Looney, M.R.; Gray, E.; Page, C.P. Animal models of mechanisms of SARS-CoV-2 infection and COVID-19 pathology. Br. J. Pharmacol. 2020, 177, 4851–4865. [Google Scholar] [CrossRef]
- Jiang, W.; Shi, L.; Cai, L.; Wang, X.; Li, J.; Li, H.; Liang, J.; Gu, Q.; Ji, G.; Li, J.; et al. A two-adjuvant multiantigen candidate vaccine induces superior protective immune responses against SARS-CoV-2 challenge. Cell Rep. 2021, 37, 110112. [Google Scholar] [CrossRef]
- Yang, M.; Li, J.; Huang, Z.; Li, H.; Wang, Y.; Wang, X.; Kang, S.; Huang, X.; Wu, C.; Liu, T.; et al. Structural Basis of a Human Neutralizing Antibody Specific to the SARS-CoV-2 Spike Protein Receptor-Binding Domain. Microbiol. Spectr. 2021, 9, e135221. [Google Scholar] [CrossRef]
- Rogers, T.F.; Zhao, F.; Huang, D.; Beutler, N.; Burns, A.; He, W.T.; Limbo, O.; Smith, C.; Song, G.; Woehl, J.; et al. Isolation of potent SARS-CoV-2 neutralizing antibodies and protection from disease in a small animal model. Science 2020, 369, 956–963. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.I.; Kim, S.G.; Kim, S.M.; Kim, E.H.; Park, S.J.; Yu, K.M.; Chang, J.H.; Kim, E.J.; Lee, S.; Casel, M.; et al. Infection and Rapid Transmission of SARS-CoV-2 in Ferrets. Cell Host Microbe 2020, 27, 704–709. [Google Scholar] [CrossRef]
- Bao, L.; Deng, W.; Huang, B.; Gao, H.; Liu, J.; Ren, L.; Wei, Q.; Yu, P.; Xu, Y.; Qi, F.; et al. The pathogenicity of SARS-CoV-2 in hACE2 transgenic mice. Nature 2020, 583, 830–833. [Google Scholar] [CrossRef]
- Carossino, M.; Kenney, D.; O’Connell, A.K.; Montanaro, P.; Tseng, A.E.; Gertje, H.P.; Grosz, K.A.; Ericsson, M.; Huber, B.R.; Kurnick, S.A.; et al. Fatal Neurodissemination and SARS-CoV-2 Tropism in K18-hACE2 Mice Is Only Partially Dependent on hACE2 Expression. Viruses 2022, 14, 535. [Google Scholar] [CrossRef]
- He, B.; Zhang, Y.; Pan, Y.; Xu, Y.; Gu, L.; Chen, L.; Wang, S. Interleukin 1 beta (IL1B) promoter polymorphism and cancer risk: Evidence from 47 published studies. Mutagenesis 2011, 26, 637–642. [Google Scholar] [CrossRef] [PubMed]
- McColl, K.E.; El-Omar, E. How does H. pylori infection cause gastric cancer? Keio J. Med. 2002, 51 (Suppl. 2), 53–56. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Han, H.; Ma, Q.; Li, C.; Liu, R.; Zhao, L.; Wang, W.; Zhang, P.; Liu, X.; Gao, G.; Liu, F.; et al. Profiling serum cytokines in COVID-19 patients reveals IL-6 and IL-10 are disease severity predictors. Emerg. Microbes Infect. 2020, 9, 1123–1130. [Google Scholar] [CrossRef]
- Zhao, Y.; Qin, L.; Zhang, P.; Li, K.; Liang, L.; Sun, J.; Xu, B.; Dai, Y.; Li, X.; Zhang, C.; et al. Longitudinal COVID-19 profiling associates IL-1RA and IL-10 with disease severity and RANTES with mild disease. JCI Insight 2020, 5, e139834. [Google Scholar] [CrossRef]
- Li, C.; Song, W.; Chan, J.F.W.; Chen, Y.; Liu, F.; Ye, Z.; Lam, A.H.C.; Cai, J.; Lee, A.C.-Y.; Wong, B.H.Y.; et al. Intranasal infection by SARS-CoV-2 Omicron variants can induce inflammatory brain damage in newly weaned hamsters. Emerg. Microbes Infect. 2023, 12, 2207678. [Google Scholar] [CrossRef]
- Rathnasinghe, R.; Strohmeier, S.; Amanat, F.; Gillespie, V.L.; Krammer, F.; Garcia-Sastre, A.; Coughlan, L.; Schotsaert, M.; Uccellini, M.B. Comparison of transgenic and adenovirus hACE2 mouse models for SARS-CoV-2 infection. Emerg. Microbes Infect. 2020, 9, 2433–2445. [Google Scholar] [CrossRef]
- Lee, C.Y.; Lowen, A.C. Animal models for SARS-CoV-2. Curr. Opin. Virol. 2021, 48, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Sutton, T.C.; Subbarao, K. Development of animal models against emerging coronaviruses: From SARS to MERS coronavirus. Virology 2015, 479–480, 247–258. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.; Zhao, X.; Peng, S.; Li, Y.; Li, J.; Zheng, H.; Zhang, Y.; Zhao, Y.; Tian, Y.; Yang, J.; et al. The Abundant Distribution and Duplication of SARS-CoV-2 in the Cerebrum and Lungs Promote a High Mortality Rate in Transgenic hACE2-C57 Mice. Int. J. Mol. Sci. 2024, 25, 997. https://doi.org/10.3390/ijms25020997
Li H, Zhao X, Peng S, Li Y, Li J, Zheng H, Zhang Y, Zhao Y, Tian Y, Yang J, et al. The Abundant Distribution and Duplication of SARS-CoV-2 in the Cerebrum and Lungs Promote a High Mortality Rate in Transgenic hACE2-C57 Mice. International Journal of Molecular Sciences. 2024; 25(2):997. https://doi.org/10.3390/ijms25020997
Chicago/Turabian StyleLi, Heng, Xin Zhao, Shasha Peng, Yingyan Li, Jing Li, Huiwen Zheng, Yifan Zhang, Yurong Zhao, Yuan Tian, Jinling Yang, and et al. 2024. "The Abundant Distribution and Duplication of SARS-CoV-2 in the Cerebrum and Lungs Promote a High Mortality Rate in Transgenic hACE2-C57 Mice" International Journal of Molecular Sciences 25, no. 2: 997. https://doi.org/10.3390/ijms25020997
APA StyleLi, H., Zhao, X., Peng, S., Li, Y., Li, J., Zheng, H., Zhang, Y., Zhao, Y., Tian, Y., Yang, J., Wang, Y., Zhang, X., & Liu, L. (2024). The Abundant Distribution and Duplication of SARS-CoV-2 in the Cerebrum and Lungs Promote a High Mortality Rate in Transgenic hACE2-C57 Mice. International Journal of Molecular Sciences, 25(2), 997. https://doi.org/10.3390/ijms25020997