
The Microbiota in Long COVID

Abstract
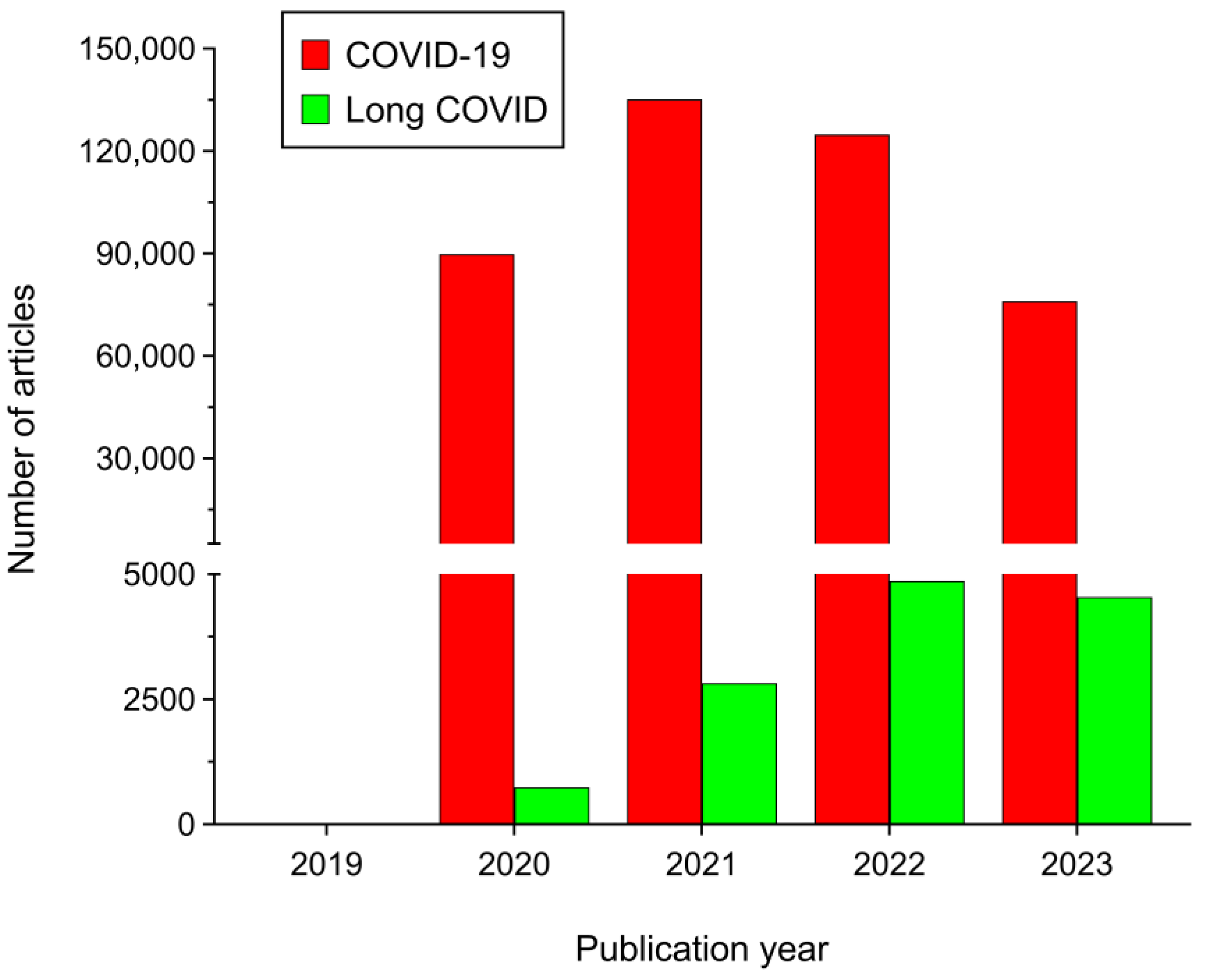
1. Introduction
2. COVID-19 and Long COVID: Two Different Sides of the Same Coin

{kind=link}
| COVID-19 | Long COVID | |
|---|---|---|
| Timelines | Symptoms usually appear between days 2 and 14 after infection by the SARS-CoV-2 virus [30] | It varies depending on the different definitions: according to the World Health Organization it occurs 3 months after the onset of COVID-19, with symptoms lasting at least 2 months and not explained by an alternative diagnosis [9]; according to the Centers for Disease Control and Prevention it occurs when a wide range of new, returning, or ongoing symptoms are present 4 weeks after SARS-CoV-2 infection [10] According to the onset of the different symptoms, parosmia appears 3 months after infection [40]; paraesthesia, hair loss, blurry vision, and swelling of the legs, hands, and feet are more common at 2 months after infection [41]; pain in joints, bones, ears, neck, and back are more common at 1 year after infection [41]; neurocognitive symptoms worsen over time and tend to persist longer [31,42]; gastrointestinal and respiratory symptoms are the most likely to resolve [31,42] |
| Diagnosis | Easy, rapid, cheap, and reliable, thanks to the availability of diagnostic tools that detect viral RNA (RT-qPCR) [24] or viral antigens [25] | Unclear, due to the lack of consensus about the definition of long COVID, the different diagnostic criteria used by distinct clinicians, and the similarity between its symptoms and those from other post-infection syndromes such as post-viral fatigue [28]. Blood tests, checking blood pressure and heart rate, chest X-ray, and measuring oxygen levels are the diagnostic tests commonly used, but long COVID usually is a diagnosis of exclusion [29] |
| Incidence | 772,166,517 cumulative cases worldwide to date (https://covid19.who.int/, accessed on 22 November 2023) | 10–30% of non-hospitalized cases [26,27] 50–70% of hospitalized cases [26,27] 10–12% of vaccinated cases [43,44] |
| Most frequent symptoms | Systemic symptoms: elevated temperature, exhaustion, and weakness Cardiovascular symptoms: palpitations, acute myocardial injury, acute pericarditis, blood pressure abnormalities, and myocarditis Ophthalmic symptoms: conjunctivitis, anterior uveitis, retinitis, and optic neuritis Musculoskeletal symptoms: body ache and arthralgia Respiratory symptoms: runny nose, cough, sore throat, and shortness of breath Gastrointestinal symptoms: diarrhoea, loss of appetite, nausea, vomiting, anorexia, and abdominal pain Olfactory and gustatory symptoms: hyposmia, anosmia, hypogeusia, and ageusia Neurological symptoms: headache, confusion, loss of speech, languidness, and malaise Dermatologic symptoms: erythematous rash, widespread urticaria, chickenpox-like vesicles, urticarial eruptions, and petechiae [30] | Systemic symptoms: fatigue, post exertional malaise, elevated temperature, chills, and skin sensations Reproductive, genitourinary, and endocrine symptoms: extreme thirst, all menstrual/period issues, abnormally irregular periods, abnormally heavy periods, and sexual dysfunction Cardiovascular symptoms: palpitations, tachycardia, pain/burning in chest, visibly inflamed/bulging veins, and bradycardia Musculoskeletal symptoms: tightness of chest, muscle aches, joint pain, stiff neck, and muscle spasms Immunologic and autoimmune symptoms: heightened reaction to old allergies, new allergies, new anaphylaxis reaction, and shingles Neurological symptoms: brain fog, headache, loss of speech, and confusion Respiratory symptoms: shortness of breath, dry cough, breathing difficulty (normal O2 level), cough with mucus production, and sneezing Gastrointestinal symptoms: diarrhoea, loss of appetite, nausea, abdominal pain, and gastroesophageal reflux Dermatologic symptoms: itchy skin, skin rashes, petechiae, COVID toe, and peeling skin [31] |
| Pathogenesis | Dysfunctional immune responses to the infection of the lower respiratory tract by the SARS-CoV-2 virus damage the pulmonary alveoli. This leads to an increase in epithelium and endothelium permeability, which promotes inflammation, coagulation, and the influx of immune cells that induce exacerbated inflammatory responses and immunopathology [38] | Several, potentially overlapping, hypothesis, highlighting [6]: |
| Risk factors | Higher age, male sex, post-menopausality, higher body mass index, smoking, longer waiting time to admission, and preexisting comorbidities (including hypertension, cardiovascular disease, diabetes cerebrovascular disease, tuberculosis, chronic renal disease, and chronic obstructive pulmonary disease) [36] | Female sex, older age, higher body mass index, smoking, and preexisting comorbidities (including anxiety and/or depression, asthma, COPD, diabetes, IHD, and immunosuppression). Moreover, patients who needed hospitalization or ICU care during the acute phase of COVID-19 present more than twice the risk of developing long COVID [37] |
| Prevention and treatment | Prevention: social distancing and prophylactic measures to avoid infection by SARS-CoV-2, and vaccines Treatment:
| Prevention: all the prevention and treatment strategies that reduce risk of suffering from severe COVID-19 Treatment: there are currently no effective treatments for long COVID, but the different symptoms are treated individually, including pacing to treat postexertional malaise, probiotics and/or diet changes to treat gastrointestinal symptoms, apheresis and anticoagulants to treat abnormal clotting, naltrexone to treat pain and fatigue, or intravenous immunoglobulin to treat immune dysfunction [6] Notably, nutritional supplements such as the combination of hydroxytyrosol, acetyl L-carnitine, and vitamins B, C, and D [53], the combination of L-Arginine and Vitamin C [54], Apportal® [55], or Requpero® [56], have been shown to improve long COVID symptoms Moreover, Sulodexide significantly improves long-lasting post-COVID-19 endothelial dysfunction and alleviates chest pain and palpitations [57] |
3. The Gut Microbiota in Long COVID
4. The Upper Respiratory Tract Microbiota in Long COVID
5. Final Considerations and Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R. A Novel Coronavirus from Patients with Pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Cucinotta, D.; Vanelli, M. WHO Declares COVID-19 a Pandemic. Acta Biomed. 2020, 91, 157–160. [Google Scholar] [CrossRef] [PubMed]
- Soheili, M.; Khateri, S.; Moradpour, F.; Mohammadzedeh, P.; Zareie, M.; Mortazavi, S.M.M.; Manifar, S.; Kohan, H.G.; Moradi, Y. The efficacy and effectiveness of COVID-19 vaccines around the world: A mini-review and meta-analysis. Ann. Clin. Microbiol. Antimicrob. 2023, 22, 42. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Shao, W.; Chen, X.; Zhang, B.; Wang, G.; Zhang, W. Real-world effectiveness of COVID-19 vaccines: A literature review and meta-analysis. Int. J. Infect. Dis. 2022, 114, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Wise, J. COVID-19: WHO declares end of global health emergency. BMJ 2023, 381, 1041. [Google Scholar] [CrossRef] [PubMed]
- Davis, H.E.; McCorkell, L.; Vogel, J.M.; Topol, E.J. Long COVID: Major findings, mechanisms and recommendations. Nat. Rev. Microbiol. 2023, 21, 133–146. [Google Scholar] [CrossRef] [PubMed]
- Baig, A.M. Chronic COVID syndrome: Need for an appropriate medical terminology for long-COVID and COVID long-haulers. J. Med. Virol. 2021, 93, 2555–2556. [Google Scholar] [CrossRef]
- Greenhalgh, T.; Knight, M.; A’Court, C.; Buxton, M.; Husain, L. Management of post-acute COVID-19 in primary care. BMJ 2020, 370, m3026. [Google Scholar] [CrossRef]
- Soriano, J.B.; Murthy, S.; Marshall, J.C.; Relan, P.; Diaz, J.V. A clinical case definition of post-COVID-19 condition by a Delphi consensus. Lancet Infect. Dis. 2022, 22, e102–e107. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Long COVID or Post-COVID Conditions; CDC: Atlanta, GA, USA, 2022.
- Marchesi, J.R.; Ravel, J. The vocabulary of microbiome research: A proposal. Microbiome 2015, 3, 31. [Google Scholar] [CrossRef]
- Consortium, H.M.P. Structure, function and diversity of the healthy human microbiome. Nature 2012, 486, 207–214. [Google Scholar] [CrossRef]
- Hou, K.; Wu, Z.X.; Chen, X.Y.; Wang, J.Q.; Zhang, D.; Xiao, C.; Zhu, D.; Koya, J.B.; Wei, L.; Li, J.; et al. Microbiota in health and diseases. Signal Transduct. Target. Ther. 2022, 7, 135. [Google Scholar] [CrossRef] [PubMed]
- Lynch, S.V.; Pedersen, O. The Human Intestinal Microbiome in Health and Disease. N. Engl. J. Med. 2016, 375, 2369–2379. [Google Scholar] [CrossRef] [PubMed]
- Dubourg, G.; Edouard, S.; Raoult, D. Relationship between nasopharyngeal microbiota and patient’s susceptibility to viral infection. Expert Rev. Anti-Infect. Ther. 2019, 17, 437–447. [Google Scholar] [CrossRef] [PubMed]
- Candel, S.; Tyrkalska, S.D.; Álvarez-Santacruz, C.; Mulero, V. The nasopharyngeal microbiome in COVID-19. Emerg. Microbes Infect. 2023, 12, e2165970. [Google Scholar] [CrossRef]
- Lloyd-Price, J.; Abu-Ali, G.; Huttenhower, C. The healthy human microbiome. Genome Med. 2016, 8, 51. [Google Scholar] [CrossRef]
- Guarner, F.; Malagelada, J.R. Gut flora in health and disease. Lancet 2003, 361, 512–519. [Google Scholar] [CrossRef]
- Yeoh, Y.K.; Zuo, T.; Lui, G.C.; Zhang, F.; Liu, Q.; Li, A.Y.; Chung, A.C.; Cheung, C.P.; Tso, E.Y.; Fung, K.S.; et al. Gut microbiota composition reflects disease severity and dysfunctional immune responses in patients with COVID-19. Gut 2021, 70, 698–706. [Google Scholar] [CrossRef]
- Zuo, T.; Zhang, F.; Lui, G.C.Y.; Yeoh, Y.K.; Li, A.Y.L.; Zhan, H.; Wan, Y.; Chung, A.C.K.; Cheung, C.P.; Chen, N.; et al. Alterations in Gut Microbiota of Patients with COVID-19 during Time of Hospitalization. Gastroenterology 2020, 159, 944–955.e8. [Google Scholar] [CrossRef]
- Sahin-Yilmaz, A.; Naclerio, R.M. Anatomy and physiology of the upper airway. Proc. Am. Thorac. Soc. 2011, 8, 31–39. [Google Scholar] [CrossRef]
- Hitzenbichler, F.; Bauernfeind, S.; Salzberger, B.; Schmidt, B.; Wenzel, J.J. Comparison of Throat Washings, Nasopharyngeal Swabs and Oropharyngeal Swabs for Detection of SARS-CoV-2. Viruses 2021, 13, 653. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; O’Leary, T.J. Relative sensitivity of anterior nares and nasopharyngeal swabs for initial detection of SARS-CoV-2 in ambulatory patients: Rapid review and meta-analysis. PLoS ONE 2021, 16, e0254559. [Google Scholar] [CrossRef] [PubMed]
- Yüce, M.; Filiztekin, E.; Özkaya, K.G. COVID-19 diagnosis—A review of current methods. Biosens. Bioelectron. 2021, 172, 112752. [Google Scholar] [CrossRef] [PubMed]
- Yamayoshi, S.; Sakai-Tagawa, Y.; Koga, M.; Akasaka, O.; Nakachi, I.; Koh, H.; Maeda, K.; Adachi, E.; Saito, M.; Nagai, H.; et al. Comparison of Rapid Antigen Tests for COVID-19. Viruses 2020, 12, 1420. [Google Scholar] [CrossRef]
- Ceban, F.; Ling, S.; Lui, L.M.W.; Lee, Y.; Gill, H.; Teopiz, K.M.; Rodrigues, N.B.; Subramaniapillai, M.; Di Vincenzo, J.D.; Cao, B.; et al. Fatigue and cognitive impairment in Post-COVID-19 Syndrome: A systematic review and meta-analysis. Brain Behav. Immun. 2022, 101, 93–135. [Google Scholar] [CrossRef] [PubMed]
- Bull-Otterson, L.; Baca, S.; Saydah, S.; Boehmer, T.K.; Adjei, S.; Gray, S.; Harris, A.M. Post–COVID Conditions among Adult COVID-19 Survivors Aged 18–64 and ≥65 Years—United States, March 2020–November 2021. Morb. Mortal. Wkly. Rep. 2022, 71, 713–717. [Google Scholar] [CrossRef]
- Behan, P.O.; Behan, W.M. Postviral fatigue syndrome. Crit. Rev. Neurobiol. 1988, 4, 157–178. [Google Scholar]
- Leviner, S. Recognizing the Clinical Sequelae of COVID-19 in Adults: COVID-19 Long-Haulers. J. Nurse Pract. 2021, 17, 946–949. [Google Scholar] [CrossRef]
- Baj, J.; Karakuła-Juchnowicz, H.; Teresiński, G.; Buszewicz, G.; Ciesielka, M.; Sitarz, R.; Forma, A.; Karakuła, K.; Flieger, W.; Portincasa, P.; et al. COVID-19: Specific and Non-Specific Clinical Manifestations and Symptoms: The Current State of Knowledge. J. Clin. Med. 2020, 9, 1753. [Google Scholar] [CrossRef]
- Davis, H.E.; Assaf, G.S.; McCorkell, L.; Wei, H.; Low, R.J.; Re’em, Y.; Redfield, S.; Austin, J.P.; Akrami, A. Characterizing long COVID in an international cohort: 7 months of symptoms and their impact. EClinicalMedicine 2021, 38, 101019. [Google Scholar] [CrossRef]
- Xie, Y.; Al-Aly, Z. Risks and burdens of incident diabetes in long COVID: A cohort study. Lancet Diabetes Endocrinol. 2022, 10, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Xu, E.; Bowe, B.; Al-Aly, Z. Long-term cardiovascular outcomes of COVID-19. Nat. Med. 2022, 28, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Kedor, C.; Freitag, H.; Meyer-Arndt, L.; Wittke, K.; Hanitsch, L.G.; Zoller, T.; Steinbeis, F.; Haffke, M.; Rudolf, G.; Heidecker, B.; et al. A prospective observational study of post-COVID-19 chronic fatigue syndrome following the first pandemic wave in Germany and biomarkers associated with symptom severity. Nat. Commun. 2022, 13, 5104. [Google Scholar] [CrossRef]
- Cairns, R.; Hotopf, M. A systematic review describing the prognosis of chronic fatigue syndrome. Occup. Med. 2005, 55, 20–31. [Google Scholar] [CrossRef]
- Wolff, D.; Nee, S.; Hickey, N.S.; Marschollek, M. Risk factors for COVID-19 severity and fatality: A structured literature review. Infection 2021, 49, 15–28. [Google Scholar] [CrossRef] [PubMed]
- Tsampasian, V.; Elghazaly, H.; Chattopadhyay, R.; Debski, M.; Naing, T.K.P.; Garg, P.; Clark, A.; Ntatsaki, E.; Vassiliou, V.S. Risk Factors Associated With Post-COVID-19 Condition: A Systematic Review and Meta-analysis. JAMA Intern. Med. 2023, 183, 566–580. [Google Scholar] [CrossRef]
- Lamers, M.M.; Haagmans, B.L. SARS-CoV-2 pathogenesis. Nat. Rev. Microbiol. 2022, 20, 270–284. [Google Scholar] [CrossRef]
- Panahi, Y.; Gorabi, A.M.; Talaei, S.; Beiraghdar, F.; Akbarzadeh, A.; Tarhriz, V.; Mellatyar, H. An overview on the treatments and prevention against COVID-19. Virol. J. 2023, 20, 23. [Google Scholar] [CrossRef]
- Walker, A.; Kelly, C.; Pottinger, G.; Hopkins, C. Parosmia-a common consequence of COVID-19. BMJ 2022, 377, e069860. [Google Scholar] [CrossRef]
- Tran, V.T.; Porcher, R.; Pane, I.; Ravaud, P. Course of post COVID-19 disease symptoms over time in the ComPaRe long COVID prospective e-cohort. Nat. Commun. 2022, 13, 1812. [Google Scholar] [CrossRef]
- Jason, L.A.; Islam, M.; Conroy, K.; Cotler, J.; Torres, C.; Johnson, M.; Mabie, B. COVID-19 Symptoms Over Time: Comparing Long-Haulers to ME/CFS. Fatigue 2021, 9, 59–68. [Google Scholar] [CrossRef]
- Al-Aly, Z.; Bowe, B.; Xie, Y. Long COVID after breakthrough SARS-CoV-2 infection. Nat. Med. 2022, 28, 1461–1467. [Google Scholar] [CrossRef]
- Ayoubkhani, D.; Bosworth, M.L.; King, S.; Pouwels, K.B.; Glickman, M.; Nafilyan, V.; Zaccardi, F.; Khunti, K.; Alwan, N.A.; Walker, A.S. Risk of Long COVID in People Infected With Severe Acute Respiratory Syndrome Coronavirus 2 After 2 Doses of a Coronavirus Disease 2019 Vaccine: Community-Based, Matched Cohort Study. Open Forum Infect. Dis. 2022, 9, ofac464. [Google Scholar] [CrossRef]
- Swank, Z.; Senussi, Y.; Manickas-Hill, Z.; Yu, X.G.; Li, J.Z.; Alter, G.; Walt, D.R. Persistent Circulating Severe Acute Respiratory Syndrome Coronavirus 2 Spike Is Associated With Post-acute Coronavirus Disease 2019 Sequelae. Clin. Infect. Dis. 2023, 76, e487–e490. [Google Scholar] [CrossRef]
- Glynne, P.; Tahmasebi, N.; Gant, V.; Gupta, R. Long COVID following mild SARS-CoV-2 infection: Characteristic T cell alterations and response to antihistamines. J. Investig. Med. 2022, 70, 61–67. [Google Scholar] [CrossRef]
- Phetsouphanh, C.; Darley, D.R.; Wilson, D.B.; Howe, A.; Munier, C.M.L.; Patel, S.K.; Juno, J.A.; Burrell, L.M.; Kent, S.J.; Dore, G.J.; et al. Immunological dysfunction persists for 8 months following initial mild-to-moderate SARS-CoV-2 infection. Nat. Immunol. 2022, 23, 210–216. [Google Scholar] [CrossRef]
- Liu, Q.; Mak, J.W.Y.; Su, Q.; Yeoh, Y.K.; Lui, G.C.; Ng, S.S.S.; Zhang, F.; Li, A.Y.L.; Lu, W.; Hui, D.S.; et al. Gut microbiota dynamics in a prospective cohort of patients with post-acute COVID-19 syndrome. Gut 2022, 71, 544–552. [Google Scholar] [CrossRef]
- Su, Y.; Yuan, D.; Chen, D.G.; Ng, R.H.; Wang, K.; Choi, J.; Li, S.; Hong, S.; Zhang, R.; Xie, J.; et al. Multiple early factors anticipate post-acute COVID-19 sequelae. Cell 2022, 185, 881–895.e20. [Google Scholar] [CrossRef]
- Arthur, J.M.; Forrest, J.C.; Boehme, K.W.; Kennedy, J.L.; Owens, S.; Herzog, C.; Liu, J.; Harville, T.O. Development of ACE2 autoantibodies after SARS-CoV-2 infection. PLoS ONE 2021, 16, e0257016. [Google Scholar] [CrossRef] [PubMed]
- Haffke, M.; Freitag, H.; Rudolf, G.; Seifert, M.; Doehner, W.; Scherbakov, N.; Hanitsch, L.; Wittke, K.; Bauer, S.; Konietschke, F.; et al. Endothelial dysfunction and altered endothelial biomarkers in patients with post-COVID-19 syndrome and chronic fatigue syndrome (ME/CFS). J. Transl. Med. 2022, 20, 138. [Google Scholar] [CrossRef]
- Spudich, S.; Nath, A. Nervous system consequences of COVID-19. Science 2022, 375, 267–269. [Google Scholar] [CrossRef]
- Naureen, Z.; Dautaj, A.; Nodari, S.; Fioretti, F.; Dhuli, K.; Anpilogov, K.; Lorusso, L.; Paolacci, S.; Michelini, S.; Guda, T.; et al. Proposal of a food supplement for the management of post-COVID syndrome. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 67–73. [Google Scholar] [CrossRef]
- Izzo, R.; Trimarco, V.; Mone, P.; Aloè, T.; Capra Marzani, M.; Diana, A.; Fazio, G.; Mallardo, M.; Maniscalco, M.; Marazzi, G.; et al. Combining L-Arginine with vitamin C improves long-COVID symptoms: The LINCOLN Survey. Pharmacol. Res. 2022, 183, 106360. [Google Scholar] [CrossRef]
- Rossato, M.S.; Brilli, E.; Ferri, N.; Giordano, G.; Tarantino, G. Observational study on the benefit of a nutritional supplement, supporting immune function and energy metabolism, on chronic fatigue associated with the SARS-CoV-2 post-infection progress. Clin. Nutr. ESPEN 2021, 46, 510–518. [Google Scholar] [CrossRef]
- Barletta, M.A.; Marino, G.; Spagnolo, B.; Bianchi, F.P.; Falappone, P.C.F.; Spagnolo, L.; Gatti, P. Coenzyme Q10 + alpha lipoic acid for chronic COVID syndrome. Clin. Exp. Med. 2023, 23, 667–678. [Google Scholar] [CrossRef]
- Charfeddine, S.; Ibnhadjamor, H.; Jdidi, J.; Torjmen, S.; Kraiem, S.; Bahloul, A.; Makni, A.; Kallel, N.; Moussa, N.; Boudaya, M.; et al. Sulodexide Significantly Improves Endothelial Dysfunction and Alleviates Chest Pain and Palpitations in Patients With Long-COVID-19: Insights From TUN-EndCOV Study. Front. Cardiovasc. Med. 2022, 9, 866113. [Google Scholar] [CrossRef]
- Tian, Y.; Sun, K.Y.; Meng, T.Q.; Ye, Z.; Guo, S.M.; Li, Z.M.; Xiong, C.L.; Yin, Y.; Li, H.G.; Zhou, L.Q. Gut Microbiota May Not Be Fully Restored in Recovered COVID-19 Patients After 3-Month Recovery. Front. Nutr. 2021, 8, 638825. [Google Scholar] [CrossRef]
- Upadhyay, V.; Suryawanshi, R.K.; Tasoff, P.; McCavitt-Malvido, M.; Kumar, R.G.; Murray, V.W.; Noecker, C.; Bisanz, J.E.; Hswen, Y.; Ha, C.W.Y.; et al. Mild SARS-CoV-2 infection results in long-lasting microbiota instability. mBio 2023, 14, e0088923. [Google Scholar] [CrossRef]
- Buttery, S.; Philip, K.E.J.; Williams, P.; Fallas, A.; West, B.; Cumella, A.; Cheung, C.; Walker, S.; Quint, J.K.; Polkey, M.I.; et al. Patient symptoms and experience following COVID-19: Results from a UK-wide survey. BMJ Open Respir. Res. 2021, 8, e001075. [Google Scholar] [CrossRef]
- Adler, L.; Gazit, S.; Pinto, Y.; Perez, G.; Mizrahi Reuveni, M.; Yehoshua, I.; Hoffman, R.; Azuri, J.; Patalon, T. Long-COVID in patients with a history of mild or asymptomatic SARS-CoV-2 infection: A Nationwide Cohort Study. Scand J. Prim. Health Care 2022, 40, 342–349. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Gu, S.; Lu, H.; Shi, D.; Guo, J.; Wu, W.R.; Yang, Y.; Li, Y.; Xu, K.J.; Ding, C.; et al. Six-month follow-up of gut microbiota richness in patients with COVID-19. Gut 2022, 71, 222–225. [Google Scholar] [CrossRef]
- Zhang, D.; Zhou, Y.; Ma, Y.; Chen, P.; Tang, J.; Yang, B.; Li, H.; Liang, M.; Xue, Y.; Liu, Y.; et al. Gut Microbiota Dysbiosis Correlates With Long COVID-19 at One-Year After Discharge. J. Korean Med. Sci. 2023, 38, e120. [Google Scholar] [CrossRef]
- Dewhirst, F.E.; Chen, T.; Izard, J.; Paster, B.J.; Tanner, A.C.; Yu, W.H.; Lakshmanan, A.; Wade, W.G. The human oral microbiome. J. Bacteriol. 2010, 192, 5002–5017. [Google Scholar] [CrossRef]
- Awano, S.; Ansai, T.; Takata, Y.; Soh, I.; Akifusa, S.; Hamasaki, T.; Yoshida, A.; Sonoki, K.; Fujisawa, K.; Takehara, T. Oral health and mortality risk from pneumonia in the elderly. J. Dent. Res. 2008, 87, 334–339. [Google Scholar] [CrossRef]
- Scannapieco, F.A. Role of oral bacteria in respiratory infection. J. Periodontol. 1999, 70, 793–802. [Google Scholar] [CrossRef]
- Seymour, G.J.; Ford, P.J.; Cullinan, M.P.; Leishman, S.; Yamazaki, K. Relationship between periodontal infections and systemic disease. Clin. Microbiol. Infect. 2007, 13 (Suppl. S4), 3–10. [Google Scholar] [CrossRef]
- Georges, F.M.; Do, N.T.; Seleem, D. Oral dysbiosis and systemic diseases. Front. Dent. Med. 2022, 3, 995423. [Google Scholar] [CrossRef]
- Haran, J.P.; Bradley, E.; Zeamer, A.L.; Cincotta, L.; Salive, M.C.; Dutta, P.; Mutaawe, S.; Anya, O.; Meza-Segura, M.; Moormann, A.M.; et al. Inflammation-type dysbiosis of the oral microbiome associates with the duration of COVID-19 symptoms and long COVID. JCI Insight 2021, 6, e152346. [Google Scholar] [CrossRef]
- Kageyama, S.; Takeshita, T.; Furuta, M.; Tomioka, M.; Asakawa, M.; Suma, S.; Takeuchi, K.; Shibata, Y.; Iwasa, Y.; Yamashita, Y. Relationships of Variations in the Tongue Microbiota and Pneumonia Mortality in Nursing Home Residents. J. Gerontol. A Biol. Sci. Med. Sci. 2018, 73, 1097–1102. [Google Scholar] [CrossRef] [PubMed]
- Mammen, M.J.; Scannapieco, F.A.; Sethi, S. Oral-lung microbiome interactions in lung diseases. Periodontol 2000 2020, 83, 234–241. [Google Scholar] [CrossRef] [PubMed]
- Larsen, J.M. The immune response to Prevotella bacteria in chronic inflammatory disease. Immunology 2017, 151, 363–374. [Google Scholar] [CrossRef]
- Segal, L.N.; Clemente, J.C.; Tsay, J.C.; Koralov, S.B.; Keller, B.C.; Wu, B.G.; Li, Y.; Shen, N.; Ghedin, E.; Morris, A.; et al. Enrichment of the lung microbiome with oral taxa is associated with lung inflammation of a Th17 phenotype. Nat. Microbiol. 2016, 1, 16031. [Google Scholar] [CrossRef]
- Segal, L.N.; Alekseyenko, A.V.; Clemente, J.C.; Kulkarni, R.; Wu, B.; Gao, Z.; Chen, H.; Berger, K.I.; Goldring, R.M.; Rom, W.N.; et al. Enrichment of lung microbiome with supraglottic taxa is associated with increased pulmonary inflammation. Microbiome 2013, 1, 19. [Google Scholar] [CrossRef] [PubMed]
- van den Bogert, B.; Meijerink, M.; Zoetendal, E.G.; Wells, J.M.; Kleerebezem, M. Immunomodulatory properties of Streptococcus and Veillonella isolates from the human small intestine microbiota. PLoS ONE 2014, 9, e114277. [Google Scholar] [CrossRef] [PubMed]
- Delwiche, E.A.; Pestka, J.J.; Tortorello, M.L. The veillonellae: Gram-negative cocci with a unique physiology. Annu. Rev. Microbiol. 1985, 39, 175–193. [Google Scholar] [CrossRef] [PubMed]
- Ren, Z.; Wang, H.; Cui, G.; Lu, H.; Wang, L.; Luo, H.; Chen, X.; Ren, H.; Sun, R.; Liu, W.; et al. Alterations in the human oral and gut microbiomes and lipidomics in COVID-19. Gut 2021, 70, 1253–1265. [Google Scholar] [CrossRef] [PubMed]
- Shetty, S.A.; Marathe, N.P.; Lanjekar, V.; Ranade, D.; Shouche, Y.S. Comparative genome analysis of Megasphaera sp. reveals niche specialization and its potential role in the human gut. PLoS ONE 2013, 8, e79353. [Google Scholar] [CrossRef]
- Doykov, I.; Hällqvist, J.; Gilmour, K.C.; Grandjean, L.; Mills, K.; Heywood, W.E. ‘The long tail of COVID-19’—The detection of a prolonged inflammatory response after a SARS-CoV-2 infection in asymptomatic and mildly affected patients. F1000Research 2020, 9, 1349. [Google Scholar] [CrossRef]
- Inciardi, R.M.; Solomon, S.D.; Ridker, P.M.; Metra, M. Coronavirus 2019 Disease (COVID-19), Systemic Inflammation, and Cardiovascular Disease. J. Am. Heart Assoc. 2020, 9, e017756. [Google Scholar] [CrossRef]
- Du, M.; Ma, Y.; Deng, J.; Liu, M.; Liu, J. Comparison of Long COVID-19 Caused by Different SARS-CoV-2 Strains: A Systematic Review and Meta-Analysis. Int. J. Environ. Res. Public Health 2022, 19, 16010. [Google Scholar] [CrossRef]
- Popkov, V.A.; Silachev, D.N.; Zalevsky, A.O.; Zorov, D.B.; Plotnikov, E.Y. Mitochondria as a Source and a Target for Uremic Toxins. Int. J. Mol. Sci. 2019, 20, 3094. [Google Scholar] [CrossRef] [PubMed]
- Mendes de Almeida, V.; Engel, D.F.; Ricci, M.F.; Cruz, C.S.; Lopes, Í.; Alves, D.A.; d’ Auriol, M.; Magalhães, J.; Machado, E.C.; Rocha, V.M.; et al. Gut microbiota from patients with COVID-19 cause alterations in mice that resemble post-COVID symptoms. Gut Microbes 2023, 15, 2249146. [Google Scholar] [CrossRef] [PubMed]
- Wong, M.C.S.; Zhang, L.; Ching, J.Y.L.; Mak, J.W.Y.; Huang, J.; Wang, S.; Mok, C.K.P.; Wong, A.; Chiu, O.L.; Fung, Y.T.; et al. Effects of Gut Microbiome Modulation on Reducing Adverse Health Outcomes among Elderly and Diabetes Patients during the COVID-19 Pandemic: A Randomised, Double-Blind, Placebo-Controlled Trial (IMPACT Study). Nutrients 2023, 15, 1982. [Google Scholar] [CrossRef]
- Zhang, L.; Xu, Z.; Mak, J.W.Y.; Chow, K.M.; Lui, G.; Li, T.C.M.; Wong, C.K.; Chan, P.K.S.; Ching, J.Y.L.; Fujiwara, Y.; et al. Gut microbiota-derived synbiotic formula (SIM01) as a novel adjuvant therapy for COVID-19: An open-label pilot study. J. Gastroenterol. Hepatol. 2022, 37, 823–831. [Google Scholar] [CrossRef]
| Reference | Publication Date | Sample Size and Relevant Features | Methodology | Timing of Analysis | Main Findings |
|---|---|---|---|---|---|
| [20] | September 2020 | N = 36 (15 COVID-19 hospitalized patients, 6 subjects with community acquired pneumonia, and 15 uninfected controls) Relevant features: COVID-19 patients were stratified by disease severity (mild, moderate, severe, or critical) | Metagenomic sequencing Sequencing platform: Illumina NextSeq 550 (Illumina, San Diego, CA, USA) | Stool samples were collected two or three times per week from time of hospitalization until discharge | Opportunistic pathogens were overrepresented and beneficial commensals were underrepresented in COVID-19 patients at time of hospitalization and at all timepoints during hospitalization Gut dysbiosis persisted even after the clearance of SARS-CoV-2 and the resolution of the respiratory symptoms While the abundance of Coprobacillus, Clostridium ramosum, and Clostridium hathewayi correlated with COVID-19 severity, the correlation between the abundance of Faecalibacterium prausnitzii (an anti-inflammatory bacterium) and disease severity was inverse |
| [58] | May 2021 | N = 14 (7 COVID-19-recovered patients and 7 uninfected controls) Relevant features: all the enrolled subjects were males | 16S rRNA gene sequencing (V3–V4) Sequencing platform: Illumina MiSeq (Illumina, San Diego, CA, USA) | An average of 3 months after discharge | Alpha and beta diversity values were significantly different in COVID-19-recovered patients compared to uninfected controls at 3 months after discharge |
| [62] | January 2022 | N = 60 (30 COVID-19 patients and 30 uninfected controls) Relevant features: Samples from COVID-19 patients with different disease severity were analysed at different timepoints: (1) acute phase, from illness onset to viral clearance; (2) convalescence, from viral clearance to 2 weeks after hospital discharge; and (3) postconvalescence, 6 months after hospital discharge | 16S rRNA gene sequencing (V3–V4) Sequencing platform: Illumina MiSeq (Illumina, CA, USA) | Stool samples were analysed at three different timepoints, according to the groups of patients established, up to 6 months after discharge | Alpha diversity significantly decreased in acute phase, convalescence, and postconvalescence patients to the same levels compared to uninfected controls |
| [48] | March 2022 | N = 106 (COVID-19 patients with different disease severity) Relevant features: 68 (76%) of the patients had developed long COVID at 6 months, whereas the remaining 38 (24%) patients had not | Metagenomic sequencing Sequencing platform: Illumina NextSeq 550 (Illumina, CA, USA) | Follow-up of patients since the diagnosis of COVID-19 up to 6 months | Gut microbiota composition at admission was associated with the occurrence of long COVID The gut microbiota of COVID-19 patients who did not develop long COVID was totally recovered and indistinguishable from that of uninfected controls at 6 months Patients who developed long COVID presented significant compositional alterations of gut microbiota at the same time after infection Their analyses revealed significant correlations between the abundance of certain bacterial taxa and the presence of some of the most characteristic symptoms of long COVID, such as the high abundance of nosocomial gut pathogens, including Clostridium innocuum, in patients with fatigue and neuropsychiatric symptoms, or the high abundance of opportunistic gut pathogens in patients with persistent respiratory symptoms |
| [63] | April 2023 | N = 219 (187 COVID-19-recovered patients and 32 uninfected controls) Relevant features: 84 (44.9%) of the long COVID-19-recovered patients developed long COVID at 1 year after discharge | 16S rRNA gene sequencing (V3–V4) Sequencing platform: Illumina MiSeq PE300 platform/NovaSeq PE250 platform (Illumina, CA, USA) | Stool samples were collected at 1 year after discharge | Alpha diversity decreased in long COVID patients compared to COVID-19-recovered patients who did not develop long COVID and uninfected controls The compositional structure of the gut microbiota was altered in COVID-19 patients who developed long COVID, as the relative abundance of the bacterial genera Eubacterium, Agathobacter, Subdoligranulum, Ruminococcus, and Veillonella was significantly different in these subjects compared to COVID-19 patients who did not develop long COVID and uninfected controls |
| [59] | June 2023 | N = 18 (14 SARS-CoV-2+ cases and 4 uninfected controls) Relevant features: all the infected subjects enrolled in this study had mild COVID-19 disease and remained outpatients, whereas uninfected individuals were household controls | 16S rRNA gene sequencing (V4) Sequencing platform: Illumina NextSeq 550 (Illumina, CA, USA) | Highly heterogeneous. Maximum of 154 days after SARS-CoV-2 initial infection | SARS-CoV-2 infection resulted in a long-lasting disruption and instability of the gut microbial ecology (with the Firmicutes and Actinobacteriota phyla and the Rothia genus as the most variable bacterial taxa following SARS-CoV-2 infection) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Álvarez-Santacruz, C.; Tyrkalska, S.D.; Candel, S. The Microbiota in Long COVID. Int. J. Mol. Sci. 2024, 25, 1330. https://doi.org/10.3390/ijms25021330
Álvarez-Santacruz C, Tyrkalska SD, Candel S. The Microbiota in Long COVID. International Journal of Molecular Sciences. 2024; 25(2):1330. https://doi.org/10.3390/ijms25021330
Chicago/Turabian StyleÁlvarez-Santacruz, Carmen, Sylwia D. Tyrkalska, and Sergio Candel. 2024. "The Microbiota in Long COVID" International Journal of Molecular Sciences 25, no. 2: 1330. https://doi.org/10.3390/ijms25021330
APA StyleÁlvarez-Santacruz, C., Tyrkalska, S. D., & Candel, S. (2024). The Microbiota in Long COVID. International Journal of Molecular Sciences, 25(2), 1330. https://doi.org/10.3390/ijms25021330