
ITRAQ Based Proteomics Reveals the Potential Mechanism of Placental Injury Induced by Prenatal Stress
 ,
,  ,
,
Abstract

1. Introduction
2. Results
2.1. Behavioral Assessment of Pregnant Rats
2.2. Neurobehavioral and Developmental Milestones of the Pups
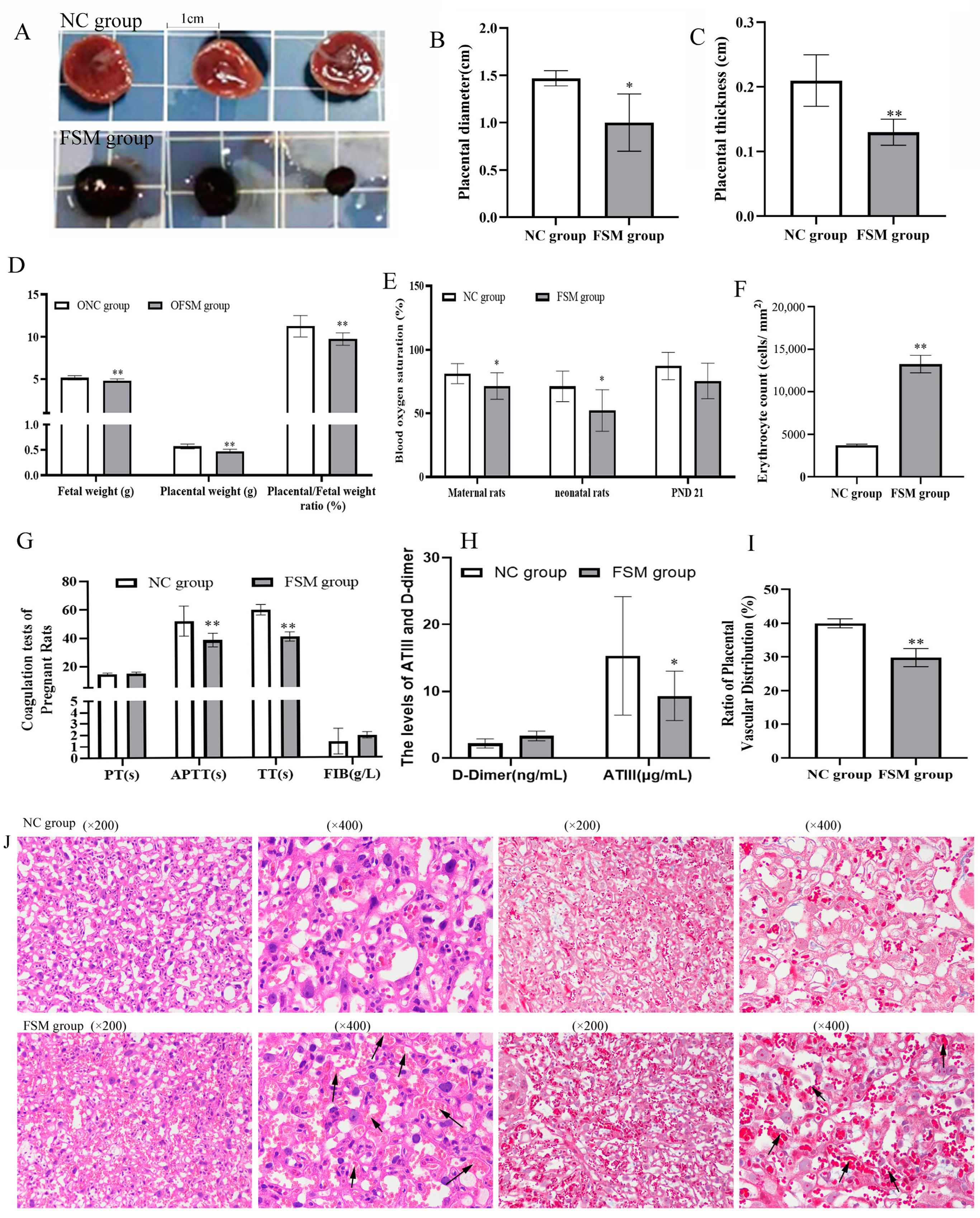
2.3. The Impact of Maternal Psychological Stress on Placental Functionality and Oxygen Transfer
2.4. Effects of Prenatal Fear Stress on Placental Histomorphology in Rats
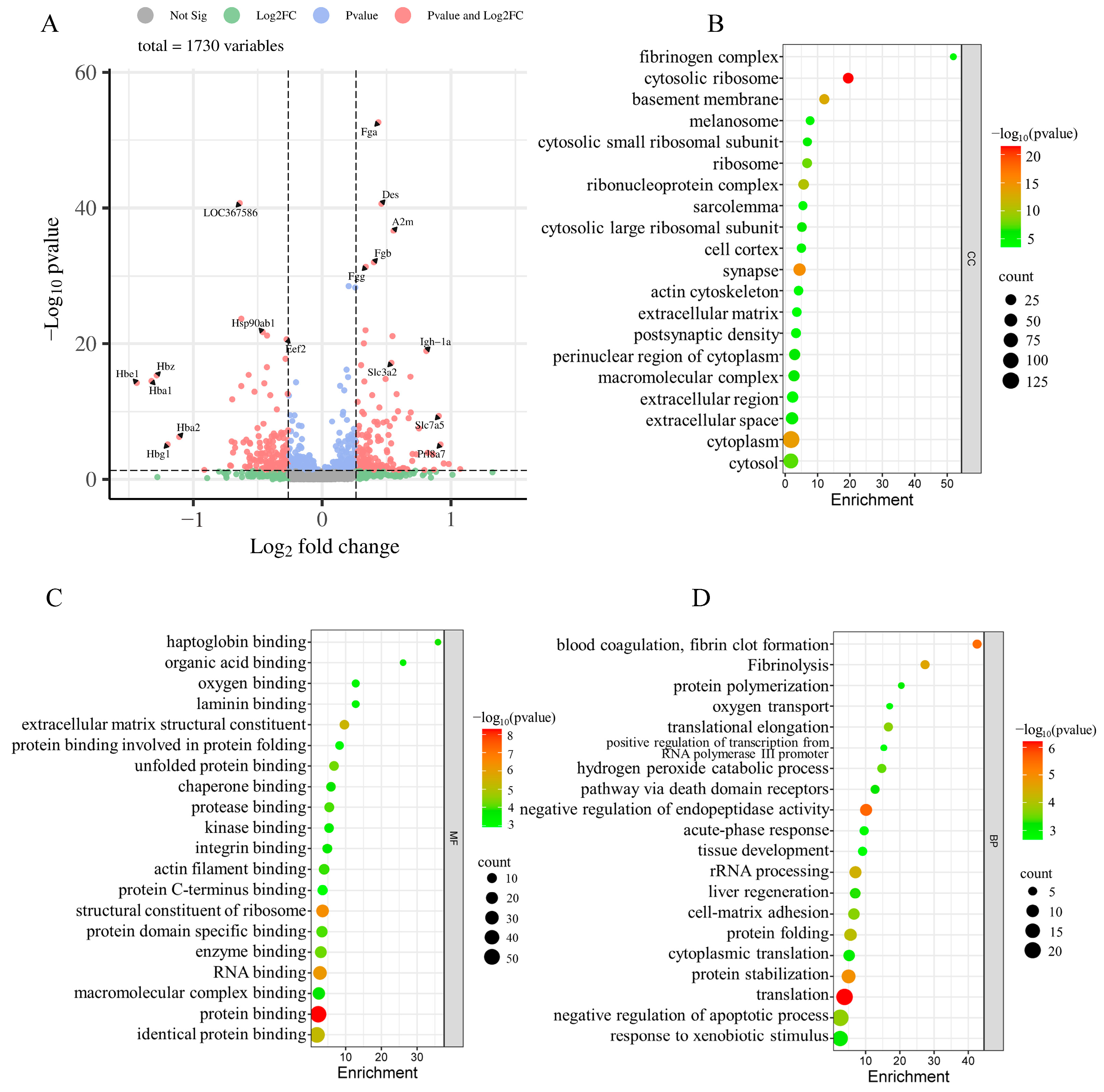
2.5. iTRAQ-Based Proteomic Analyses
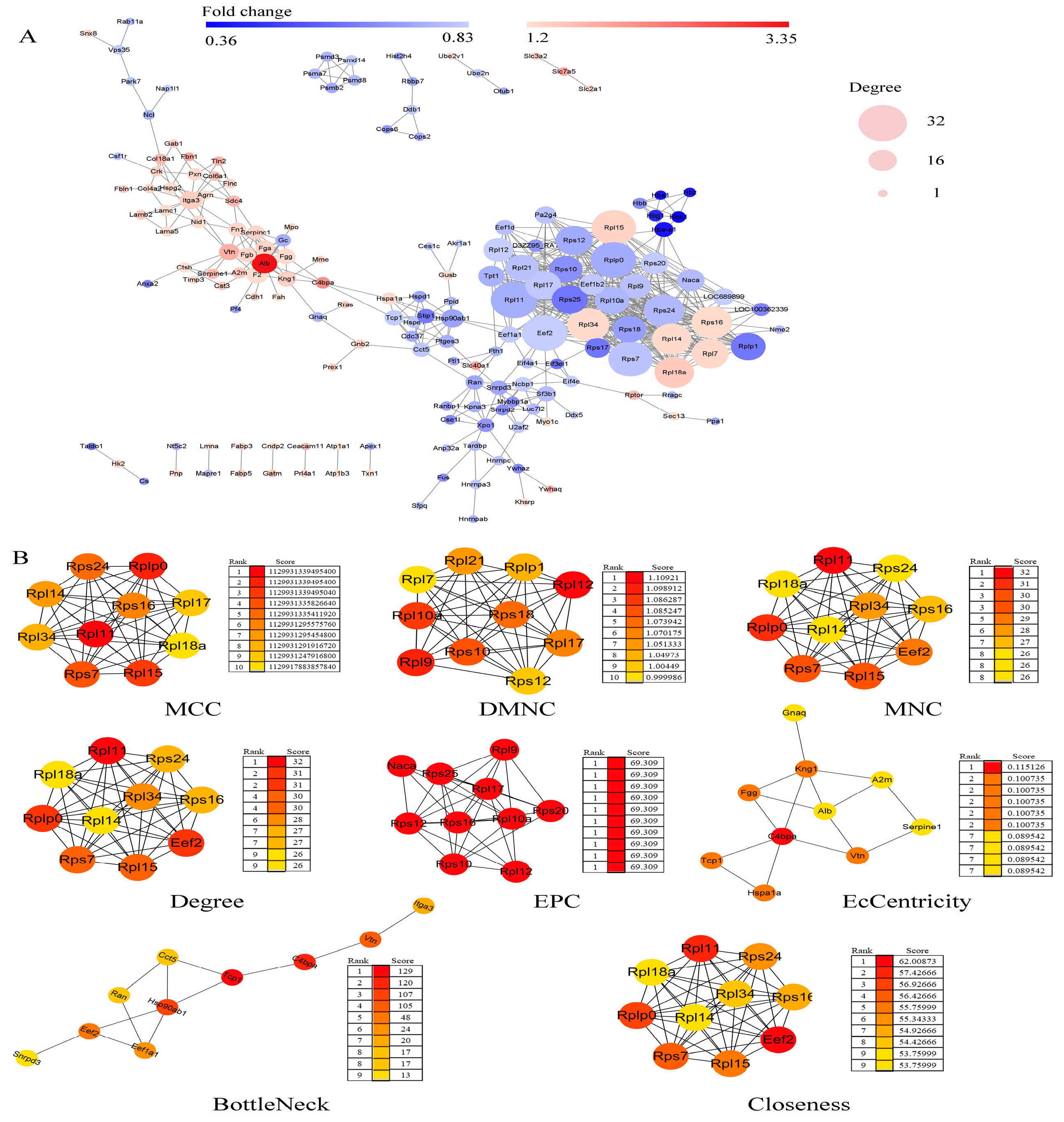
2.6. Protein–Protein Interaction (PPI) Network Construction and Hub Genes Extraction
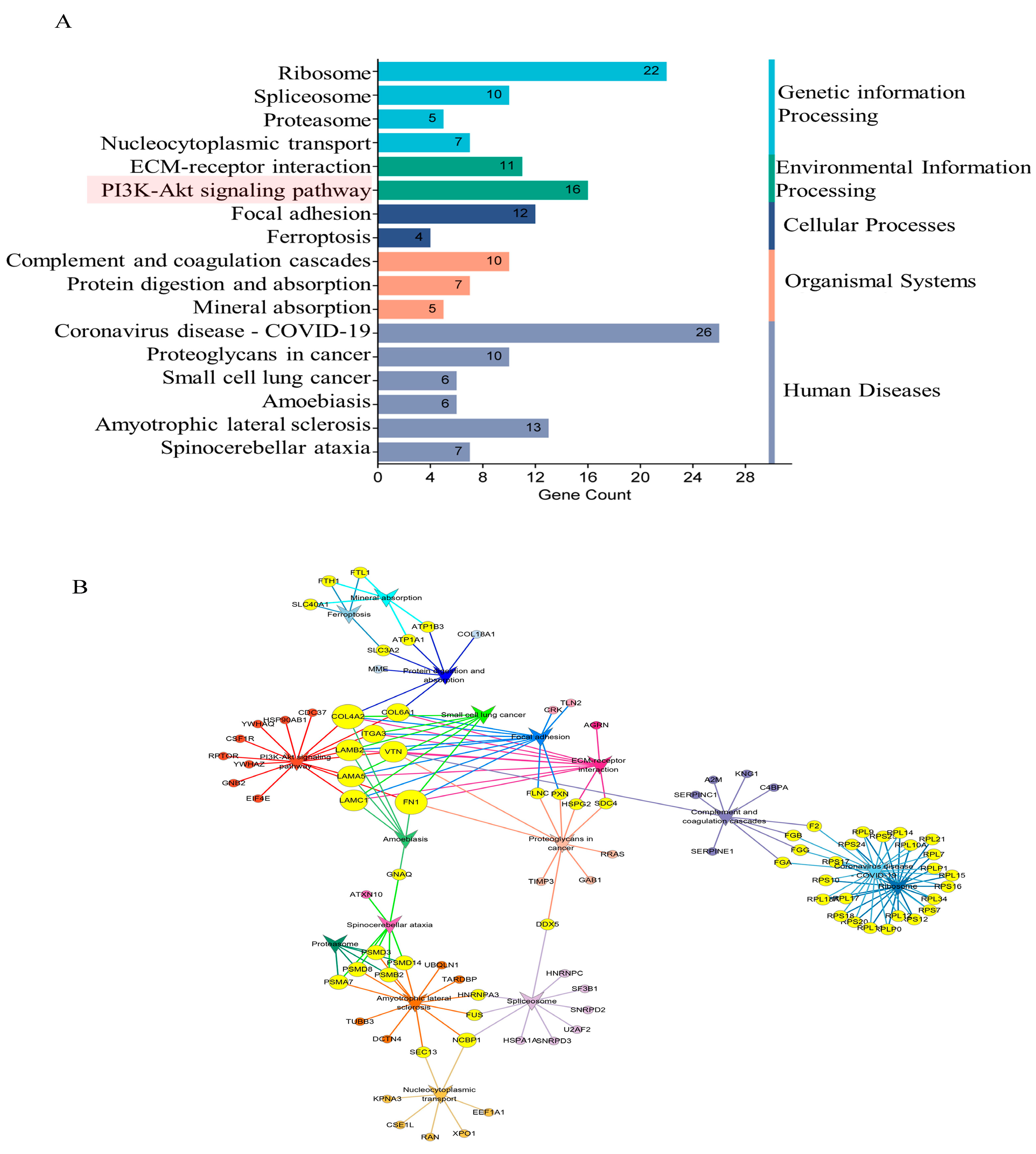
2.7. GO and KEGG Enrichment Analyses of DEPs
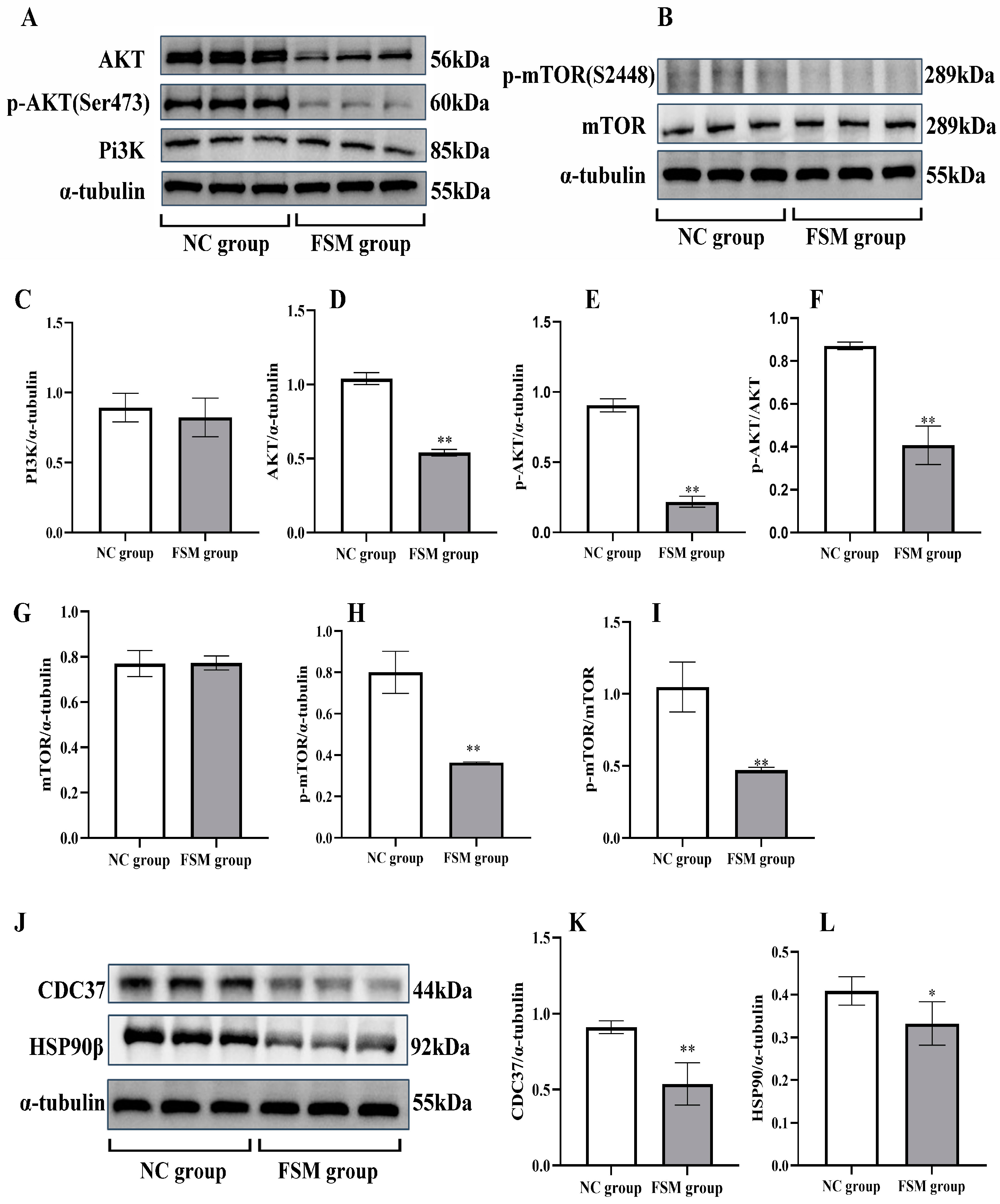
2.8. The Impact of Maternal Stress on Placental PI3K/AKT/mTOR Pathway Signaling
2.9. The Impact of Maternal Stress on Placental Cdc37 and HSP90β Protein Expression
3. Discussion
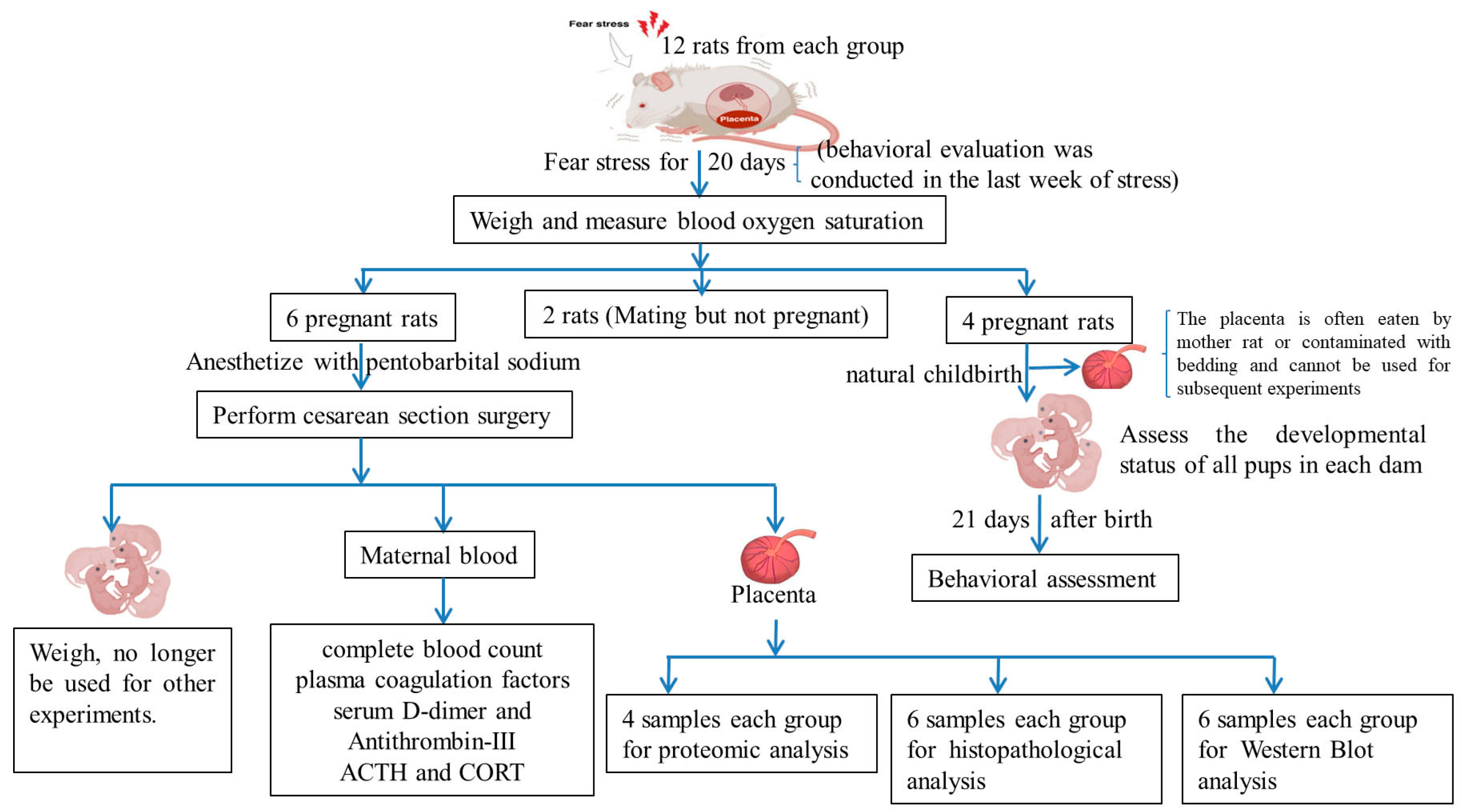
4. Materials and Methods
4.1. Animal Care and Ethics Statement
4.2. Animal Treatment
4.2.1. Prenatal Fear Stress
4.2.2. Stress State Assessment of Pregnant Rats
4.2.3. The Behavioral Assessment of the Offspring
4.3. Method of Behavioral Testing
4.3.1. Open Field-Test
4.3.2. TST
4.3.3. SPT
4.3.4. MWM
4.4. Sample Preparation
4.5. Biochemical Assay
4.6. Histological Examination
4.7. Proteomic Analysis
4.7.1. Protein Extraction
4.7.2. Protein Digestion and Peptide iTRAQ Labeling
4.7.3. LC-MS/MS Analyses
4.7.4. Protein Identification and Quantification
4.7.5. Bioinformatics Analysis
4.8. Western Blot Analysis
4.9. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession | Protein Names |
|---|---|
| P06238 | Alpha-2-macroglobulin (A2m) |
| D4A2F1 | Agrin (Agrn) |
| P02770 | Serum albumin (Alb) |
| A0A0G2JX94 | Protein Amot (Amot) |
| P36633 | Amiloride-sensitive amine oxidase [copper-containing] (Aoc1) |
| P29975 | Aquaporin-1 (Aqp1) |
| P06685 | Sodium/potassium-transporting ATPase subunit alpha-1 (Atp1a1) |
| Q63377 | Sodium/potassium-transporting ATPase subunit beta-3 (Atp1b3) |
| M0RCA6 | Advillin (Avil) |
| Q63514 | C4b-binding protein alpha chain (C4bpa) |
| Q9R0T4 | Cadherin-1 (Cdh1) |
| Q91XU8 | Phosphatidate cytidylyltransferase 2 (Cds2) |
| Q4QQU3 | Carcinoembryonic antigen-related cell adhesion molecule 11 (Ceacam11) |
| Q4V8J0 | Carcinoembryonic antigen gene family 4 (Cgm4) |
| A0A0G2JZF2 | CAP-Gly domain-containing linker protein 2 (Clip2) |
| Q6Q0N1 | Cytosolic non-specific dipeptidase (Cndp2) |
| F1LR02 | Procollagen, type XVIII, alpha 1, isoform CRA_a (Col18a1) |
| F1M6Q3 | Protein Col4a2 (Col4a2) |
| D3ZUL3 | Protein Col6a1 (Col6a1) |
| Q63768 | Adapter molecule crk (Crk) |
| P14841 | Cystatin-C (Cst3) |
| P00787 | Cathepsin B (Ctsb) |
| Q32PZ8 | Ctsq protein (Ctsql2) |
| Q9QUR2-2 | Isoform 2 of Dynactin subunit 4 (Dctn4) |
| A0A0G2K719 | Protein Ddx3x (Ddx3x) |
| Q6P725 | Desmin (Des) |
| Q9EPB1 | Dipeptidyl peptidase 2 (Dpp7) |
| D4A269 | Uncharacterized protein (ENSRNOG00000005630) |
| A0A0G2JUY4 | Uncharacterized protein (ENSRNOG00000050102) |
| D3ZF97 | Protein Erlec1 (Erlec1) |
| D3ZVK6 | Protein LOC103691893 (Erv3) |
| D3ZJ32 | Protein Esyt2 (Esyt2) |
| G3V843 | Prothrombin (F2) |
| P07483 | Fatty acid-binding protein, heart (Fabp3) |
| P55053 | Fatty acid-binding protein, epidermal (Fabp5) |
| P25093 | Fumarylacetoacetase (Fah) |
| B2GV94 | Fam134c protein (Fam134c) |
| B2GUZ9 | Fam49b protein (Fam49b) |
| D3ZQ25 | Fibulin-1 (Fbln1) |
| G3V9M6 | Fibrillin 1, isoform CRA_a (Fbn1) |
| P06399 | Fibrinogen alpha chain (Fga) |
| P14480 | Fibrinogen beta chain (Fgb) |
| P02680-2 | Isoform Gamma-A of Fibrinogen gamma chain (Fgg) |
| A0A0H2UHR7 | Filamin-C (Flnc) |
| P04937 | Fibronectin (Fn1) |
| A0A0G2K0A1 | Protein Fndc3c1 (Fndc3c2) |
| D3ZAL7 | Protein Gab1 (Gab1) |
| P50442 | Glycine amidinotransferase, mitochondrial (Gatm) |
| P54313 | Guanine nucleotide-binding protein G(I)/G(S)/G(T) subunit beta-2 (Gnb2) |
| F1LQQ8 | Beta-glucuronidase (Gusb) |
| O35952-2 | Isoform 2 of Hydroxyacylglutathione hydrolase, mitochondrial (Hagh) |
| D4A3K5 | Histone H1.1 (Hist1h1a) |
| D3ZBN0 | Histone H1.5 (Hist1h1b) |
| P27881 | Hexokinase-2 (Hk2) |
| Q6P747 | Heterochromatin protein 1-binding protein 3 (Hp1bp3) |
| Q6AYS8 | Estradiol 17-beta-dehydrogenase 11 (Hsd17b11) |
| Q5I0N4 | Estradiol 17-beta-dehydrogenase 2 (Hsd17b2) |
| P0DMW0 | Heat shock 70 kDa protein 1A (Hspa1a) |
| F1LTJ5 | Uncharacterized protein (Hspg2) |
| B0BNJ2 | CEA-related cell adhesion molecule 12 (Predicted) (Igfl3) |
| P20761 | Ig gamma-2B chain C region (Igh-1a) |
| D3ZCG9 | Integrin alpha 3 variant A (Itga3) |
| M0R961 | Far upstream element-binding protein 2 (Khsrp) |
| Q70AM4 | Kinesin-like protein (Kif13b) |
| Q5PQU1 | Kininogen 1 (Kng1) |
| P08932 | T-kininogen 2 (KNT2) |
| G3V779 | Ladinin (Predicted), isoform CRA_b (Lad1) |
| F1MAN8 | Laminin, alpha 5, isoform CRA_a (Lama5) |
| M0R6K0 | Laminin subunit beta-2 (Lamb2) |
| F1MAA7 | Protein Lamc1 (Lamc1) |
| Q99MZ8 | LIM and SH3 domain protein 1 (Lasp1) |
| G3V8L3 | Lamin A, isoform CRA_b (Lmna) |
| Q4V799 | Similar to solute carrier family 7 (Cationic amino acid transporter, y+ system), member 3 (LOC292543) |
| A0A0G2JYK0 | Protein LOC299282 (LOC299282) |
| A0A0G2JU68 | Protein LOC304903 (LOC680415) |
| Q5BK33 | p55 protein (LOC691642) |
| A0A0G2JYI0 | Protein Lrba (Lrba) |
| P28494 | Alpha-mannosidase 2 (Man2a1) |
| Q5M7W5 | Microtubule-associated protein 4 (Map4) |
| F1LMW7 | Myristoylated alanine-rich C-kinase substrate (Marcks) |
| D3ZBP4 | Protein-methionine sulfoxide oxidase MICAL1 (Mical1) |
| A0A0H2UHX5 | Neprilysin (Mme) |
| D3ZGE2 | Myeloperoxidase (Mapped) (Mpo) |
| Q9Z1W6-4 | Isoform 4 of Protein LYRIC (Mtdh) |
| Q63355 | Unconventional myosin-Ic (Myo1c) |
| Q6JE36 | Protein NDRG1 (Ndrg1) |
| Q6P756 | Adaptin ear-binding coat-associated protein 2 (Necap2) |
| F1LM84 | Nidogen-1 (Nid1) |
| F1LZB6 | Protein Nrk (Nrk) |
| D3ZZ80 | Obscurin-like protein 1 (Obsl1) |
| P20717 | Protein-arginine deiminase type-2 (Padi2) |
| G3V6S1 | PRKC apoptosis WT1 regulator protein (Pawr) |
| Q62785 | 28 kDa heat- and acid-stable phosphoprotein (Pdap1) |
| Q9Z1Z9 | PDZ and LIM domain protein 7 (Pdlim7) |
| Q5I0D7 | Xaa-Pro dipeptidase (Pepd) |
| Q6P742 | Proteolipid protein 2 (Plp2) |
| P85973 | Purine nucleoside phosphorylase (Pnp) |
| D3ZS72 | Protein Prex1 (Prex1) |
| P09320 | Prolactin-4A1 (Prl4a1) |
| G3V850 | Prolactin-7A2 (Prl7a3) |
| A0A0G2JTQ7 | Prolactin-8A7 (Prl8a7) |
| Q4V883 | Protein Psg16 (Psg16) |
| Q4V8K0 | Pregnancy-specific glycoprotein 29 (Psg29) |
| P06302 | Prothymosin alpha (Ptma) |
| O88902 | Tyrosine-protein phosphatase non-receptor type 23 (Fragment) (Ptpn23) |
| G3V8L9 | Polymerase I and transcript release factor (Ptrf) |
| Q1EG89 | Myocardial ischemic preconditioning associated protein 7 (Pxn) |
| F1LSW7 | 60S ribosomal protein L14 (Rpl14) |
| P61314 | 60S ribosomal protein L15 (Rpl15) |
| P62718 | 60S ribosomal protein L18a (Rpl18a) |
| B2RZD4 | 60S ribosomal protein L34 (Rpl34) |
| B0K031 | 60S ribosomal protein L7 (Rpl7) |
| P62250 | 40S ribosomal protein S16 (Rps16) |
| D3ZDU2 | Protein Rptor (Rptor) |
| D3Z8L7 | Ras-related protein R-Ras (Rras) |
| Q9ERM8 | Secretory carrier membrane protein 3 (Fragment) (Scamp3) |
| I3PMS6 | Syndecan (Fragment) (Sdc4) |
| Q5XFW8 | Protein SEC13 homolog (Sec13) |
| Q8VIF7 | Selenium-binding protein 1 (Selenbp1) |
| A0A0H2UHI5 | Protein LOC299282 (Serpina3n) |
| Q4G075 | Leukocyte elastase inhibitor A (Serpinb1a) |
| Q5M7T5 | Protein Serpinc1 (Serpinc1) |
| P20961 | Plasminogen activator inhibitor 1 (Serpine1) |
| Q5EC47 | Protein Slc13a4 (Slc13a4) |
| A0A0G2K2S2 | Solute carrier family 2, facilitated glucose transporter member 1 (Slc2a1) |
| Q9EQ25 | Sodium-coupled neutral amino acid transporter 4 (Slc38a4) |
| Q794F9 | 4F2 cell-surface antigen heavy chain (Slc3a2) |
| M0R6B7 | Protein LOC100911874 (Slc40a1) |
| Q63016 | Large neutral amino acids transporter small subunit 1 (Slc7a5) |
| D3ZUC9 | Oxidative-stress responsive 1 (Predicted) (Snx8) |
| P12001 | 60S ribosomal protein L18 (Strn) |
| P48032 | Metalloproteinase inhibitor 3 (Timp3) |
| Q4V8N0 | Lipocalin 7, isoform CRA_a (Tinagl1) |
| D3ZA84 | Protein Tln2 (Tln2) |
| D3ZF26 | Protein Tnks1bp1 (Tnks1bp1) |
| Q4QRB4 | Tubulin beta-3 chain (Tubb3) |
| Q5RJR2 | Twinfilin-1 (Twf1) |
| P11232 | Thioredoxin (Txn1) |
| D3ZF39 | Protein Uap1 (Uap1) |
| D3ZFY8 | Protein LOC100362142 (Ube2v1) |
| Q3KR94 | Protein Vtn (Vtn) |
| P70490 | Lactadherin (Ywhaq) |
| F1LVT2 | Protein Zfp36l3 (Zfp36l3) |
| Q1RP74 | Protein LOC100911774 (ZH14) |
| A0A0G2JSJ3 | Solute carrier family 2 (Facilitated glucose transporter), member 3 (Slc2a3) |
| Accession | Protein Names |
|---|---|
| Q5XI64 | Monoacylglycerol lipase ABHD6 (Abhd6) |
| P45953 | Very long-chain specific acyl-CoA dehydrogenase, mitochondrial (Acadvl) |
| P51635 | Alcohol dehydrogenase [NADP(+)] (Akr1a1) |
| P23457 | 3-alpha-hydroxysteroid dehydrogenase (Akr1c14) |
| A0A0G2K7X0 | Acidic leucine-rich nuclear phosphoprotein 32 family member A (Anp32a) |
| Q07936 | Annexin A2 (Anxa2) |
| Q4FZU6 | Annexin A8 (Anxa8) |
| P43138 | DNA-(apurinic or apyrimidinic site) lyase (Apex1) |
| D4A6C5 | Protein Arhgap1 (Arhgap1) |
| Q9ER24 | Ataxin-10 (Atxn10) |
| Q6P7P5 | Basic leucine zipper and W2 domain-containing protein 1 (Bzw1) |
| Q68FQ0 | T-complex protein 1 subunit epsilon (Cct5) |
| A0A0G2K3C1 | Hsp90 co-chaperone Cdc37 (Cdc37) |
| G3V8V4 | Longevity assurance homolog 2 (S. cerevisiae) (Cers2) |
| P10959 | Carboxylesterase 1C (Ces1c) |
| D4A4T9 | Cysteine and histidine-rich domain-containing protein 1 (Chordc1) |
| P61203 | COP9 signalosome complex subunit 2 (Cops2) |
| D3ZI16 | COP9 (Constitutive photomorphogenic) homolog, subunit 6 (Arabidopsis thaliana) (Predicted), isoform CRA_a (Cops6) |
| G3V936 | Citrate synthase (Cs) |
| D3ZPR0 | Chromosome segregation 1-like (S. cerevisiae) (Predicted) (Cse1l) |
| A0A0G2KBC4 | Tyrosine-protein kinase receptor (Csf1r) |
| G3V8T4 | DNA damage-binding protein 1 (Ddb1) |
| Q5U216 | ATP-dependent RNA helicase DDX39A (Ddx39a) |
| B6DTP5 | DEAD (Asp-Glu-Ala-Asp) box polypeptide 5 (Fragment) (Ddx5) |
| Q7M0E3 | Destrin (Dstn) |
| P62630 | Elongation factor 1-alpha 1 (Eef1a1) |
| B5DEN5 | Eukaryotic translation elongation factor 1 beta 2 (Eef1b2) |
| Q68FR9 | Elongation factor 1-delta (Eef1d) |
| P05197 | Elongation factor 2 (Eef2) |
| A0A0G2K273 | Eukaryotic translation initiation factor 3 subunit E (Eif3e) |
| Q6P3V8 | Eukaryotic translation initiation factor 4A1 (Eif4a1) |
| P63074 | Eukaryotic translation initiation factor 4E (Eif4e) |
| D4A8F2 | Protein Rsu1 (ENSRNOG00000017595) |
| D4A6G6 | Protein LOC100362339 (ENSRNOG00000018471) |
| D3ZZ95 | 60S ribosomal protein L36 (ENSRNOG00000031947) |
| P19132 | Ferritin heavy chain (Fth1) |
| P02793 | Ferritin light chain 1 (Ftl1) |
| Q5PQK2 | Fusion, derived from t(1216) malignant liposarcoma (Human) (Fus) |
| P04276 | Vitamin D-binding protein (Gc) |
| D4AE68 | Guanine nucleotide binding protein, alpha q polypeptide, isoform CRA_a (Gnaq) |
| A0A0G2K0Z7 | Glycerol-3-phosphate dehydrogenase, mitochondrial (Gpd2) |
| Q00715 | Histone H2B type 1 (H2B1) |
| P01946 | Hemoglobin subunit alpha-1/2 (Hba1) |
| A0A0G2JSV6 | Protein Hba2 (Hba2) |
| A0A0G2JSW3 | Hemoglobin subunit beta-1 (Hbb) |
| P11517 | Hemoglobin subunit beta-2 (HBB2) |
| O88752 | Epsilon 1 globin (Hbe1) |
| Q63066 | Beta-globin chain (Fragment) (Hbg1) |
| G3V8R3 | Protein Hbz (Hbz) |
| D3ZJ08 | Histone H3 (Hist2h3c2) |
| P62804 | Histone H4 (Hist2h4) |
| Q6URK4-2 | Isoform 2 of Heterogeneous nuclear ribonucleoprotein A3 (Hnrnpa3) |
| Q9QX80 | CArG-binding factor A (Hnrnpab) |
| G3V9R8 | Heterogeneous nuclear ribonucleoprotein C (Hnrnpc) |
| P34058 | Heat shock protein HSP 90-beta (Hsp90ab1) |
| P63039 | 60 kDa heat shock protein, mitochondrial (Hspd1) |
| P26772 | 10 kDa heat shock protein, mitochondrial (Hspe1) |
| Q56R18 | Importin subunit alpha (Kpna3) |
| Q5U1W8 | High-mobility group nucleosome binding domain 1 (LOC100911295) |
| Q63910 | Alpha globin (LOC287167) |
| Q5M7V3 | LOC367586 protein (LOC367586) |
| D4A5L9 | Protein LOC679794 (LOC679794) |
| D3ZTH8 | Protein LOC689899 (LOC689899) |
| B2RYP6 | LUC7-like 2 (S. cerevisiae) (Luc7l2) |
| Q66HR2 | Microtubule-associated protein RP/EB family member 1 (Mapre1) |
| O35821 | Myb-binding protein 1A (Mybbp1a) |
| Q63357 | Unconventional myosin-Id (Myo1d) |
| B2RYX0 | Naca protein (Naca) |
| G3V6H9 | Nucleosome assembly protein 1-like 1 (Nap1l1) |
| Q56A27 | Nuclear cap-binding protein subunit 1 (Ncbp1) |
| P13383 | Nucleolin (Ncl) |
| P19804 | Nucleoside diphosphate kinase B (Nme2) |
| Q8K1Q0 | Glycylpeptide N-tetradecanoyltransferase 1 (Nmt1) |
| D3ZMY7 | Protein Nt5c2 (Nt5c2) |
| A0A0G2KB27 | Protein Nup62cl (Nup62cl) |
| A0JPJ7 | Obg-like ATPase 1 (Ola1) |
| B2RYG6 | Ubiquitin thioesterase OTUB1 (Otub1) |
| Q6AYD3 | Proliferation-associated protein 2G4 (Pa2g4) |
| O88767 | Protein deglycase DJ-1 (Park7) |
| P06765 | Platelet factor 4 (Pf4) |
| P16446 | Phosphatidylinositol transfer protein alpha isoform (Pitpna) |
| F7EPH4 | Protein Sar1a (Ppa1) |
| Q6DGG0 | Peptidyl-prolyl cis-trans isomerase D (Ppid) |
| P09321 | Prolactin-3B1 (Prl3b1) |
| A0A0G2K0W9 | Proteasome subunit alpha type (Psma7) |
| P40307 | Proteasome subunit beta type-2 (Psmb2) |
| Q4V8E2 | Proteasome (Prosome, macropain) 26S subunit, non-ATPase, 14 (Psmd14) |
| Q5U2S7 | Proteasome (Prosome, macropain) 26S subunit, non-ATPase, 3 (Psmd3) |
| Q3B8P5 | Psmd8 protein (Fragment) (Psmd8) |
| P83868 | Prostaglandin E synthase 3 (Ptges3) |
| Q63011 | Zero beta-globin (Fragment) (Q63011) |
| P62494 | Ras-related protein Rab-11A (Rab11a) |
| P62828 | GTP-binding nuclear protein Ran (Ran) |
| D4A2G9 | Protein Ranbp1 (Ranbp1) |
| Q71UF4 | Histone-binding protein RBBP7 (Rbbp7) |
| B2RZA9 | Ube2l3 protein (rCG_36772) |
| G3V7V6 | All-trans-13,14-dihydroretinol saturase, isoform CRA_b (Retsat) |
| D4AD70 | Protein RGD1561636 (RGD1561636) |
| P62907 | 60S ribosomal protein L10a (Rpl10a) |
| Q4V8I6 | 60S ribosomal protein L11 (Rpl11) |
| P23358 | 60S ribosomal protein L12 (Rpl12) |
| P24049 | 60S ribosomal protein L17 (Rpl17) |
| D3ZPN7 | Protein LOC100360604 (Rpl21) |
| P17077 | 60S ribosomal protein L9 (Rpl9) |
| P19945 | 60S acidic ribosomal protein P0 (Rplp0) |
| P19944 | 60S acidic ribosomal protein P1 (Rplp1) |
| P63326 | 40S ribosomal protein S10 (Rps10) |
| D3ZHB3 | 40S ribosomal protein S12 (Rps12) |
| A0A0H2UHQ8 | 40S ribosomal protein S17 (Rps17) |
| A0A0H2UHT6 | 40S ribosomal protein S18 (Rps18) |
| A0A0H2UHG7 | 40S ribosomal protein S20 (Rps20) |
| P62850-2 | Isoform 2 of 40S ribosomal protein S24 (Rps24) |
| A0A0H2UHU0 | 40S ribosomal protein S25 (Rps25) |
| O55215 | Ribosomal protein S2 (Rps2-ps6) |
| P62083 | 40S ribosomal protein S7 (Rps7) |
| Q0D2L6 | Protein Rragc (Rragc) |
| Q6AXV4 | Sorting and assembly machinery component 50 homolog (Samm50) |
| Q6AY18 | Protein Sar1a (Sar1a) |
| Q7TP42 | Ab2-292 (Sec62) |
| Q63556 | Serine protease inhibitor A3M (Fragment) (Serpina3m) |
| G3V7T6 | Protein Sf3b1 (Sf3b1) |
| A0A0G2K8K0 | Protein Sfpq (Sfpq) |
| B5DES0 | Protein Snrpd2 (Snrpd2) |
| M0R907 | Protein Snrpd3 (Snrpd3) |
| P38656 | Lupus La protein homolog (Ssb) |
| O35814 | Stress-induced-phosphoprotein 1 (Stip1) |
| Q6AYB6 | Synapse associated protein 1 (Syap1) |
| Q9EQS0 | Transaldolase (Taldo1) |
| I6L9G6 | Protein Tardbp (Tardbp) |
| P28480 | T-complex protein 1 subunit alpha (Tcp1) |
| P09495 | Tropomyosin alpha-4 chain (Tpm4) |
| P63029 | Translationally-controlled tumor protein (Tpt1) |
| Q498E0 | Thioredoxin domain-containing protein 12 (Txndc12) |
| F2Z3T9 | Protein U2af2 (U2af2) |
| Q9EQX9 | Ubiquitin-conjugating enzyme E2 N (Ube2n) |
| Q9JJP9 | Ubiquilin-1 (Ubqln1) |
| B0BN55 | Uroporphyrinogen decarboxylase (Urod) |
| F1LM09 | Ubiquitin carboxyl-terminal hydrolase (Usp7) |
| G3V8A5 | Protein Vps35 (Vps35) |
| Q80U96 | Exportin-1 (Xpo1) |
| P63102 | 14-3-3 protein zeta/delta (Ywhaz) |
| Q8VIL3 | ZW10 interactor (Zwint) |
References
- Chan, J.C.; Nugent, B.M.; Bale, T.L. Parental Advisory: Maternal and Paternal Stress Can Impact Offspring Neurodevelopment. Biol. Psychiatry 2018, 83, 886–894. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.J.; Yang, L.P.; Hou, J.L.; Li, X.M.; Chen, L.; Zhu, J.H.; Wang, Q.Y.; Li, G.; Zhao, P.Y.; Liu, X.H.; et al. Prenatal Stress Impairs Postnatal Learning and Memory Development via Disturbance of the cGMP-PKG Pathway and Oxidative Phosphorylation in the Hippocampus of Rats. Front. Mol. Neurosci. 2020, 13, 158. [Google Scholar] [CrossRef] [PubMed]
- Dahlerup, B.R.; Egsmose, E.L.; Siersma, V.; Mortensen, E.L.; Hedegaard, M.; Knudsen, L.E.; Mathiesen, L. Maternal stress and placental function, a study using questionnaires and biomarkers at birth. PLoS ONE 2018, 13, e0207184. [Google Scholar] [CrossRef]
- Van den Bergh, B.R.H.; van den Heuvel, M.I.; Lahti, M.; Braeken, M.; de Rooij, S.R.; Entringer, S.; Hoyer, D.; Roseboom, T.; Räikkönen, K.; King, S.; et al. Prenatal developmental origins of behavior and mental health: The influence of maternal stress in pregnancy. Neurosci. Biobehav. Rev. 2020, 117, 26–64. [Google Scholar] [CrossRef] [PubMed]
- Mathiesen, L.; Bay-Richter, C.; Wegener, G.; Liebenberg, N.; Knudsen, L.E. Maternal stress and placental function; ex vivo placental perfusion studying cortisol, cortisone, tryptophan and serotonin. PLoS ONE 2020, 15, e0233979. [Google Scholar] [CrossRef]
- Lund, R.J.; Kyläniemi, M.; Pettersson, N.; Kaukonen, R.; Konki, M.; Scheinin, N.M.; Karlsson, L.; Karlsson, H.; Ekholm, E. Placental DNA methylation marks are associated with maternal depressive symptoms during early pregnancy. Neurobiol. Stress 2021, 15, 100374. [Google Scholar] [CrossRef]
- Wang, Q.; Pan, M.; Zhang, T.; Jiang, Y.; Zhao, P.; Liu, X.; Gao, A.; Yang, L.; Hou, J. Fear Stress During Pregnancy Affects Placental m6A-Modifying Enzyme Expression and Epigenetic Modification Levels. Front. Genet. 2022, 13, 927615. [Google Scholar] [CrossRef]
- Aushev, V.N.; Li, Q.; Deyssenroth, M.; Zhang, W.; Finik, J.; Hurd, Y.L.; Nomura, Y.; Chen, J. Placental gene network modules are associated with maternal stress during pregnancy and infant temperament. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2021, 35, e21922. [Google Scholar] [CrossRef]
- Francis, J.W.; Hausmann, S.; Ikram, S.; Yin, K.; Mealey-Farr, R.; Flores, N.M.; Trinh, A.T.; Chasan, T.; Thompson, J.; Mazur, P.K.; et al. FAM86A methylation of eEF2 links mRNA translation elongation to tumorigenesis. Mol. Cell 2024, 84, 1753–1763.e7. [Google Scholar] [CrossRef]
- Yang, S.; Chen, S.; Zhao, Y.; Wu, T.; Wang, Y.; Li, T.; Fu, L.; Ye, T.; Hu, Y.Q.; Chen, H. Identification of a coagulation-related signature correlated with immune infiltration and their prognostic implications in lung adenocarcinoma. Thorac. Cancer 2023, 14, 3295–3308. [Google Scholar] [CrossRef]
- Petrenko, V.; Vrublevskaya, V.; Bystrova, M.; Masulis, I.; Kopylova, E.; Skarga, Y.; Zhmurina, M.; Morenkov, O. Proliferation, migration, and resistance to oxidative and thermal stresses of HT1080 cells with knocked out genes encoding Hsp90α and Hsp90β. Biochem. Biophys. Res. Commun. 2023, 674, 62–68. [Google Scholar] [CrossRef]
- Zhang, H.; Zheng, T.; Qin, C.; Zhang, X.; Lin, H.; Huang, X.; Liu, Q.; Chang, S.; Zhang, L.; Guo, J.; et al. CCT6A promotes cell proliferation in colon cancer by targeting BIRC5 associated with p53 status. Cancer Gene Ther. 2024, 31, 1151–1163. [Google Scholar] [CrossRef]
- Brannigan, R.; Cannon, M.; Tanskanen, A.; Huttunen, M.O.; Leacy, F.P.; Clarke, M.C. The association between subjective maternal stress during pregnancy and offspring clinically diagnosed psychiatric disorders. Acta Psychiatr. Scand. 2019, 139, 304–310. [Google Scholar] [CrossRef]
- Jensen Pena, C.; Monk, C.; Champagne, F.A. Epigenetic effects of prenatal stress on 11beta-hydroxysteroid dehydrogenase-2 in the placenta and fetal brain. PLoS ONE 2012, 7, e39791. [Google Scholar] [CrossRef] [PubMed]
- Lévy, F. Neuroendocrine control of maternal behavior in non-human and human mammals. Ann. D’endocrinologie 2016, 77, 114–125. [Google Scholar] [CrossRef] [PubMed]
- Hellgren, M. Hemostasis during normal pregnancy and puerperium. Semin. Thromb. Hemost. 2003, 29, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Ciurea, E.L.; Berceanu, C.; Voicu, N.L.; Pirnoiu, D.; Berceanu, S.; Stepan, A.E. Morphological Survey of Placenta in Trombophilia Related Hypoperfusion of Maternal-Fetal Blood Flow. Curr. Health Sci. J. 2018, 44, 85–91. [Google Scholar]
- Austin, A.W.; Wissmann, T.; von Kanel, R. Stress and hemostasis: An update. Semin. Thromb. Hemost. 2013, 39, 902–912. [Google Scholar] [CrossRef]
- von Känel, R. Acute mental stress and hemostasis: When physiology becomes vascular harm. Thromb. Res. 2015, 135 (Suppl. 1), S52–S55. [Google Scholar] [CrossRef]
- Simcox, L.E.; Ormesher, L.; Tower, C.; Greer, I.A. Thrombophilia and Pregnancy Complications. Int. J. Mol. Sci. 2015, 16, 28418–28428. [Google Scholar] [CrossRef]
- Kupferminc, M.J. Thrombophilia and pregnancy. Reprod. Biol. Endocrinol. RB&E 2003, 1, 111. [Google Scholar]
- Ozmen, A.; Unek, G.; Korgun, E.T. Effect of glucocorticoids on mechanisms of placental angiogenesis. Placenta 2017, 52, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Drobinskaia, A.N.; Nadeeva, A.P.; Zhukova, V.A.; Pasman, N.M.; Karpov, M.A.; Teliatnikova, N.V. Placental morphology in inherited thrombophilia. Arkhiv Patol. 2014, 76, 33–36. [Google Scholar]
- Chen, M.; Chao, B.; Xu, J.; Liu, Z.; Tao, Y.; He, J.; Wang, J.; Yang, H.; Luo, X.; Qi, H. CPT1A modulates PI3K/Akt/mTOR pathway to promote preeclampsia. Placenta 2023, 133, 23–31. [Google Scholar] [CrossRef]
- Liu, Z.; Hong, Z.; Qu, P. Proteomic Analysis of Human Endometrial Tissues Reveals the Roles of PI3K/AKT/mTOR Pathway and Tumor Angiogenesis Molecules in the Pathogenesis of Endometrial Cancer. BioMed Res. Int. 2020, 2020, 5273969. [Google Scholar] [CrossRef]
- Zhao, H.; Wong, R.J.; Stevenson, D.K. The Impact of Hypoxia in Early Pregnancy on Placental Cells. Int. J. Mol. Sci. 2021, 22, 9675. [Google Scholar] [CrossRef]
- Beetch, M.; Akhaphong, B.; Wong, A.; Clifton, B.; Jo, S.; Mohan, R.; Abrahante Llorens, J.E.; Alejandro, E.U. Impact of placental mTOR deficiency on peripheral insulin signaling in adult mice offspring. J. Mol. Endocrinol. 2023, 71, e230035. [Google Scholar] [CrossRef]
- Gupta, M.B.; Jansson, T. Novel roles of mechanistic target of rapamycin signaling in regulating fetal growth†. Biol. Reprod. 2019, 100, 872–884. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, O.R.; Fisher, H.M.; Dionelis, K.N.; Jeffreys, E.C.; Higgins, J.S.; Musial, B.; Sferruzzi-Perri, A.N.; Fowden, A.L. Corticosterone alters materno-fetal glucose partitioning and insulin signalling in pregnant mice. J. Physiol. 2015, 593, 1307–1321. [Google Scholar] [CrossRef]
- Basso, A.D.; Solit, D.B.; Chiosis, G.; Giri, B.; Tsichlis, P.; Rosen, N. Akt forms an intracellular complex with heat shock protein 90 (Hsp90) and Cdc37 and is destabilized by inhibitors of Hsp90 function. J. Biol. Chem. 2002, 277, 39858–39866. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, S.; Ding, D.; Yu, Z.; Sun, W.; Wang, Y. Up-Regulation of Cdc37 Contributes to Schwann Cell Proliferation and Migration After Sciatic Nerve Crush. Neurochem. Res. 2018, 43, 1182–1190. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Wu, F.; Hu, J.; Xu, Y.; Zhang, J.; Li, Y.; Lin, Y.; Liu, X. LDHA deficiency inhibits trophoblast proliferation via the PI3K/AKT/FOXO1/CyclinD1 signaling pathway in unexplained recurrent spontaneous abortion. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2023, 37, e22744. [Google Scholar] [CrossRef] [PubMed]
- Zohn, I.E. Hsp90 and complex birth defects: A plausible mechanism for the interaction of genes and environment. Neurosci. Lett. 2020, 716, 134680. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.; Tang, X.; Woodley, D.T.; Chen, M.; Li, W. The Distinct Assignments for Hsp90α and Hsp90β: More Than Skin Deep. Cells 2023, 12, 277. [Google Scholar] [CrossRef]
- Xinmin, L.; Liping, Y.; Haijiao, W.; Jianghui, Z.; Junlin, H.; Xianghong, Z. Correlation between cognitive development and levels of dopamine and 3, 4-dihydroxyphenylacetic acid in the hippocampus in 80-day-old neonatal rats born of fear-impaired pregnant rats. Chin. J. Comp. Med. 2017, 11, 10–14. [Google Scholar]
- Liping, Y.; Gai, L.; Haijiao, W.; Jianghui, Z.; Xinmin, L.; Junlin, H.; Xianghong, Z. Effects of terror stress of the pregnant rats on emotional changes of 21-day-old neonatal offspring. CJTCMP 2018, 01, 62–64. [Google Scholar]
- Geng, S.; Yang, L.; Cheng, F.; Zhang, Z.; Li, J.; Liu, W.; Li, Y.; Chen, Y.; Bao, Y.; Chen, L.; et al. Gut Microbiota Are Associated with Psychological Stress-Induced Defections in Intestinal and Blood-Brain Barriers. Front. Microbiol. 2019, 10, 3067. [Google Scholar] [CrossRef]
- Takeuchi, T.; Iwanaga, M.; Harada, E. Possible regulatory mechanism of DHA-induced anti-stress reaction in rats. Brain Res. 2003, 964, 136–143. [Google Scholar] [CrossRef]
- Smart, J.L.; Dobbing, J. Vulnerability of developing brain. II. Effects of early nutritional deprivation on reflex ontogeny and development of behaviour in the rat. Brain Res. 1971, 28, 85–95. [Google Scholar] [CrossRef]
- Wu, S.; Gao, Q.; Zhao, P.; Gao, Y.; Xi, Y.; Wang, X.; Liang, Y.; Shi, H.; Ma, Y. Sulforaphane produces antidepressant- and anxiolytic-like effects in adult mice. Behav. Brain Res. 2016, 301, 55–62. [Google Scholar] [CrossRef]
- Wang, T.; Chen, H.; Lv, K.; Ji, G.; Zhang, Y.; Wang, Y.; Li, Y.; Qu, L. iTRAQ-based proteomics analysis of hippocampus in spatial memory deficiency rats induced by simulated microgravity. J. Proteom. 2017, 160, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P.; et al. The STRING database in 2021: Customizable protein-protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2021, 49, D605–D612. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Shi, J.; Min, Z.; Zhu, H.; Sun, W. A Network Pharmacology Approach to Uncover the Molecular Mechanisms of Herbal Formula Kang-Bai-Ling for Treatment of Vitiligo. Evid. Based Complement. Alternat. Med. 2019, 2019, 3053458. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Lü, Z.; Tian, C.; Ouyang, W.; Xiong, Y.; You, Y.; Chen, L.; Deng, Y.; Zhao, X.; Sun, X. Mechanism of Shenbing decoction III in the treatment of proteinuria in chronic kidney disease: A network pharmacology-based study. Nan Fang Yi Ke Da Xue Xue Bao 2019, 39, 227–234. [Google Scholar]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.H.; Pagès, F.; Trajanoski, Z.; Galon, J. ClueGO: A Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef]



| Index | NC Group | FSM Group |
|---|---|---|
| White blood cells (×109/L) | 7.20 ± 2.92 | 3.48 ± 1.45 * |
| Lymphocyte (×109/L) | 3.87 ± 1.02 | 2.12 ± 0.79 ** |
| Monocyte (×109/L) | 0.22 ± 0.12 | 0.17 ± 0.10 |
| Neutrophil (×109/L) | 3.12 ± 1.82 | 1.20 ± 0.59 * |
| Percentage of lymphocytes (%) | 55.70 ± 7.86 | 62.23 ± 6.54 |
| Percentage of monocytes (%) | 3.42 ± 0.45 | 4.55 ± 1.22 |
| Percentage of neutrophils (%) | 40.88 ± 7.67 | 33.22 ± 5.49 |
| Red blood cell (×1012/L) | 5.48 ± 0.27 | 5.70 ± 0.77 |
| Hemoglobin (g/L) | 107.00 ± 5.51 | 111.00 ± 13.67 |
| Hematocrit (%) | 33.33 ± 1.84 | 32.22 ± 3.62 |
| Mean corpuscular volume (fL) | 60.97 ± 1.44 | 56.80 ± 2.27 ** |
| Mean corpuscular hemoglobin (pg) | 19.50 ± 0.44 | 19.47 ± 0.46 |
| Red cell distribution width (%) | 12.32 ± 0.71 | 13.08 ± 1.41 |
| Platelet (×109/L) | 940.50 ± 128.04 | 1421.00 ± 275.97 ** |
| Mean platelet volume (fL) | 5.93 ± 0.24 | 8.07 ± 0.68 ** |
| Gene Names | Protein Names | Fold Change | p-Value |
|---|---|---|---|
| Alb | Serum albumin | 3.344481605 | 0.00107535 |
| C4bpa | C4b-binding protein alpha chain | 2.100840336 | 0.028742183 |
| Plp2 | Proteolipid protein 2 | 1.976284585 | 0.00518908 |
| Slc40a1 | Protein LOC100911874 | 1.923076923 | 0.004230614 |
| Prl8a7 | Prolactin-8A7 | 1.890359168 | 7.2348 × 10−6 |
| Slc7a5 | Large neutral amino acids transporter small subunit 1 | 1.872659176 | 4.26129 × 10−10 |
| Fbn1 | Fibrillin 1 | 1.824817518 | 0.035886107 |
| Aqp1 | Aquaporin-1 | 1.814882033 | 0.000190706 |
| Vtn | Protein Vtn | 1.76366843 | 0.000118123 |
| Igh-1a | Ig gamma-2B chain C region | 1.751313485 | 1.1581 × 10−19 |
| Rplp1 | 60S acidic ribosomal protein P1 | 0.619195046 | 0.000108538 |
| Rps17 | 40S ribosomal protein S17 | 0.616522811 | 1.57927 × 10−12 |
| Rps25 | 40S ribosomal protein S25 | 0.61462815 | 2.51193 × 10−6 |
| Eif3e | Eukaryotic translation initiation factor 3 subunit E | 0.612369871 | 0.010033231 |
| Taldo1 | Transaldolase | 0.609756098 | 0.008266795 |
| Hba2 | Protein Hba2 | 0.463606861 | 5.45252 × 10−7 |
| Hbg1 | Beta-globin chain (Fragment) | 0.43554007 | 7.10019 × 10−6 |
| Hbz | Protein Hbz | 0.410677618 | 4.77399 × 10−16 |
| Hba1 | Hemoglobin subunit alpha-1/2 | 0.399361022 | 3.2938 × 10−15 |
| Sample Name | Labels Information | |
|---|---|---|
| NC group | N1 | 113 |
| N2 | 114 | |
| N3 | 115 | |
| N4 | 116 | |
| FSM group | M1 | 117 |
| M2 | 118 | |
| M3 | 119 | |
| M4 | 121 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Hou, J.; Yang, L.; Zhang, T.; Jiang, Y.; Du, Z.; Ma, H.; Li, G.; Zhu, J.; Chen, P. ITRAQ Based Proteomics Reveals the Potential Mechanism of Placental Injury Induced by Prenatal Stress. Int. J. Mol. Sci. 2024, 25, 9978. https://doi.org/10.3390/ijms25189978
Li Y, Hou J, Yang L, Zhang T, Jiang Y, Du Z, Ma H, Li G, Zhu J, Chen P. ITRAQ Based Proteomics Reveals the Potential Mechanism of Placental Injury Induced by Prenatal Stress. International Journal of Molecular Sciences. 2024; 25(18):9978. https://doi.org/10.3390/ijms25189978
Chicago/Turabian StyleLi, Yujie, Junlin Hou, Liping Yang, Tong Zhang, Yu Jiang, Zhixing Du, Huizi Ma, Gai Li, Jianghui Zhu, and Ping Chen. 2024. "ITRAQ Based Proteomics Reveals the Potential Mechanism of Placental Injury Induced by Prenatal Stress" International Journal of Molecular Sciences 25, no. 18: 9978. https://doi.org/10.3390/ijms25189978
APA StyleLi, Y., Hou, J., Yang, L., Zhang, T., Jiang, Y., Du, Z., Ma, H., Li, G., Zhu, J., & Chen, P. (2024). ITRAQ Based Proteomics Reveals the Potential Mechanism of Placental Injury Induced by Prenatal Stress. International Journal of Molecular Sciences, 25(18), 9978. https://doi.org/10.3390/ijms25189978