

Transcriptomic Analysis of the Molecular Mechanism Potential of Grafting—Enhancing the Ability of Oriental Melon to Tolerate Low-Nitrogen Stress

 ,
,
Abstract
1. Introduction
2. Results
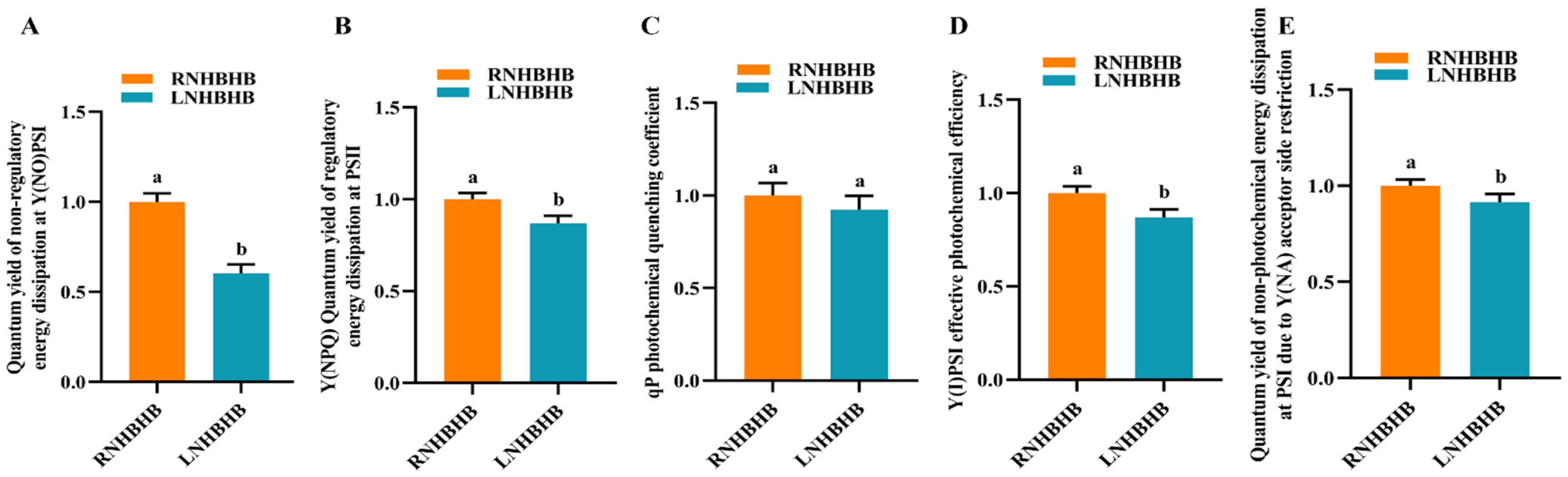
2.1. Effect of Low-Nitrogen Stress on the Fluorescence Characteristics of Self-Rooted Oriental Melon Seedlings
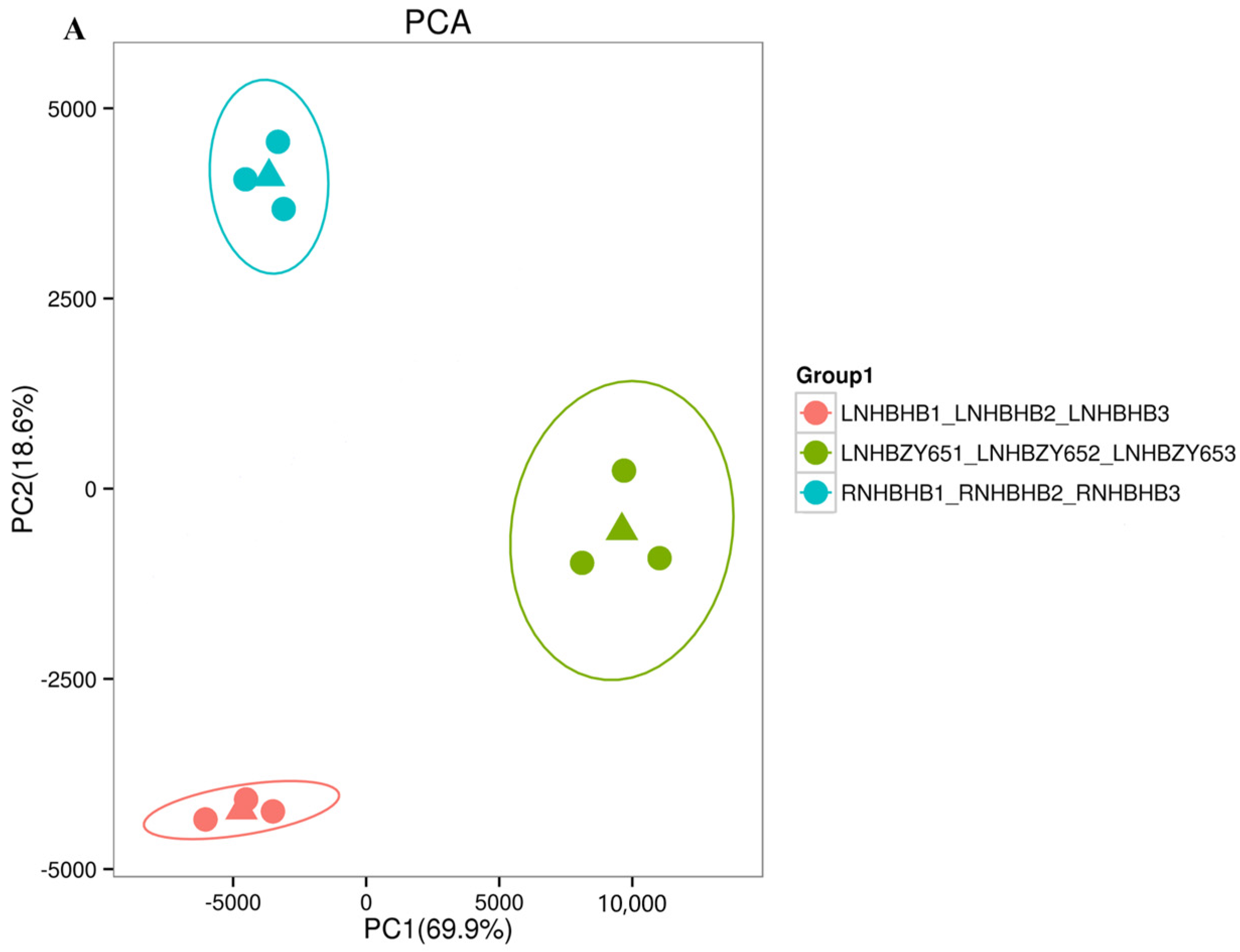
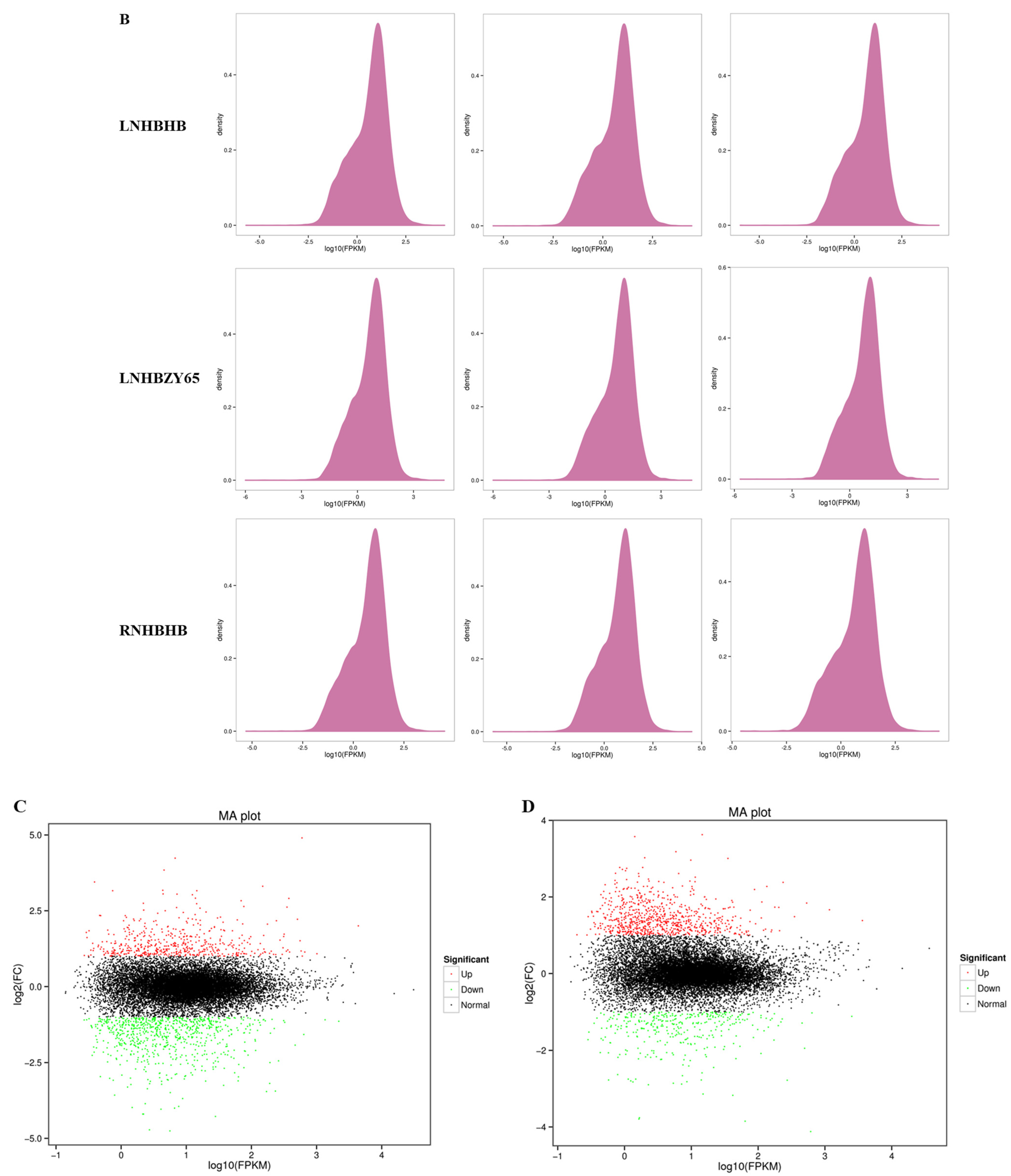
2.2. Transcriptome Data Analysis and Functional Annotation
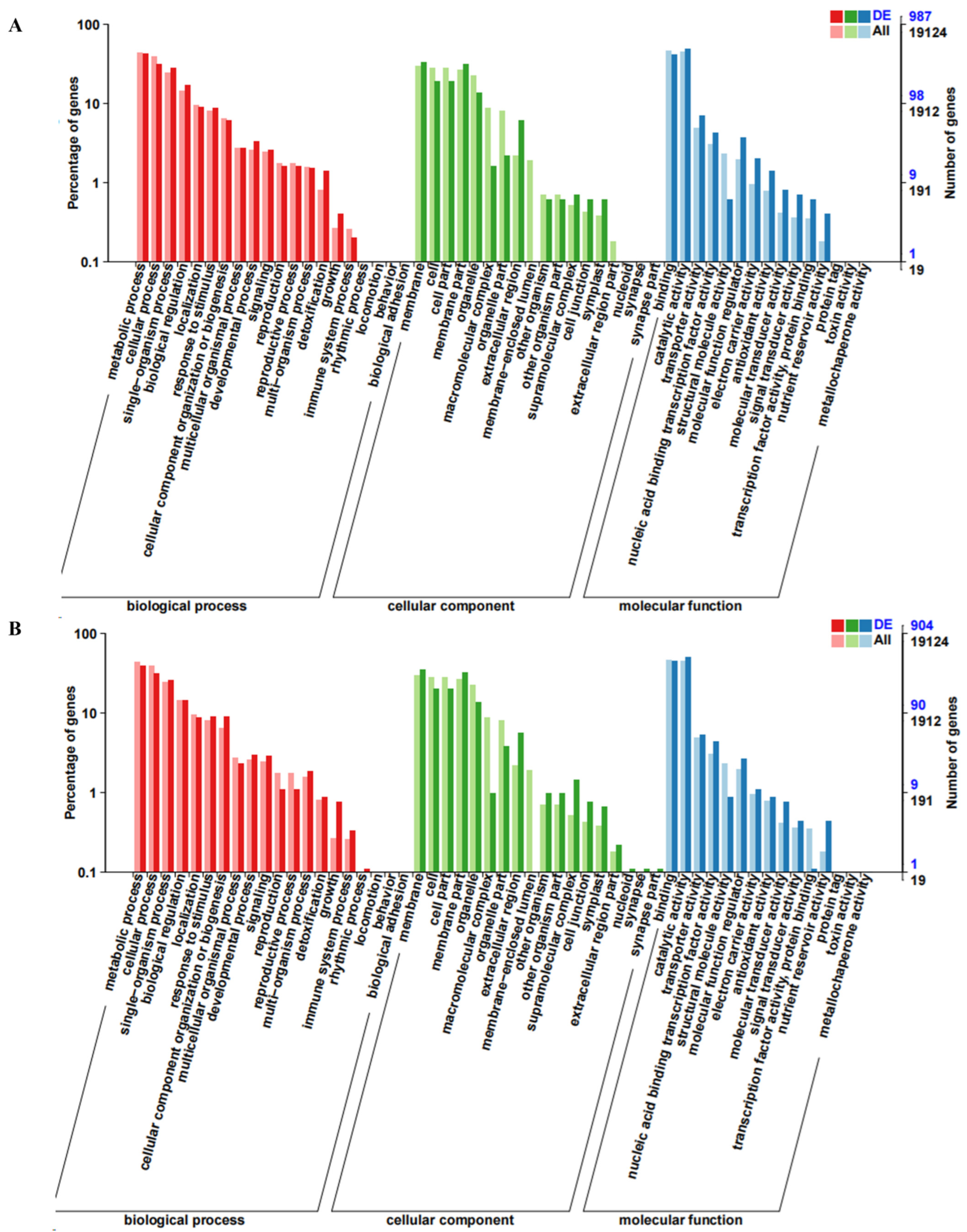
2.3. GO and KEGG Enrichment Analysis of Differentially Expressed Genes (DEGs)
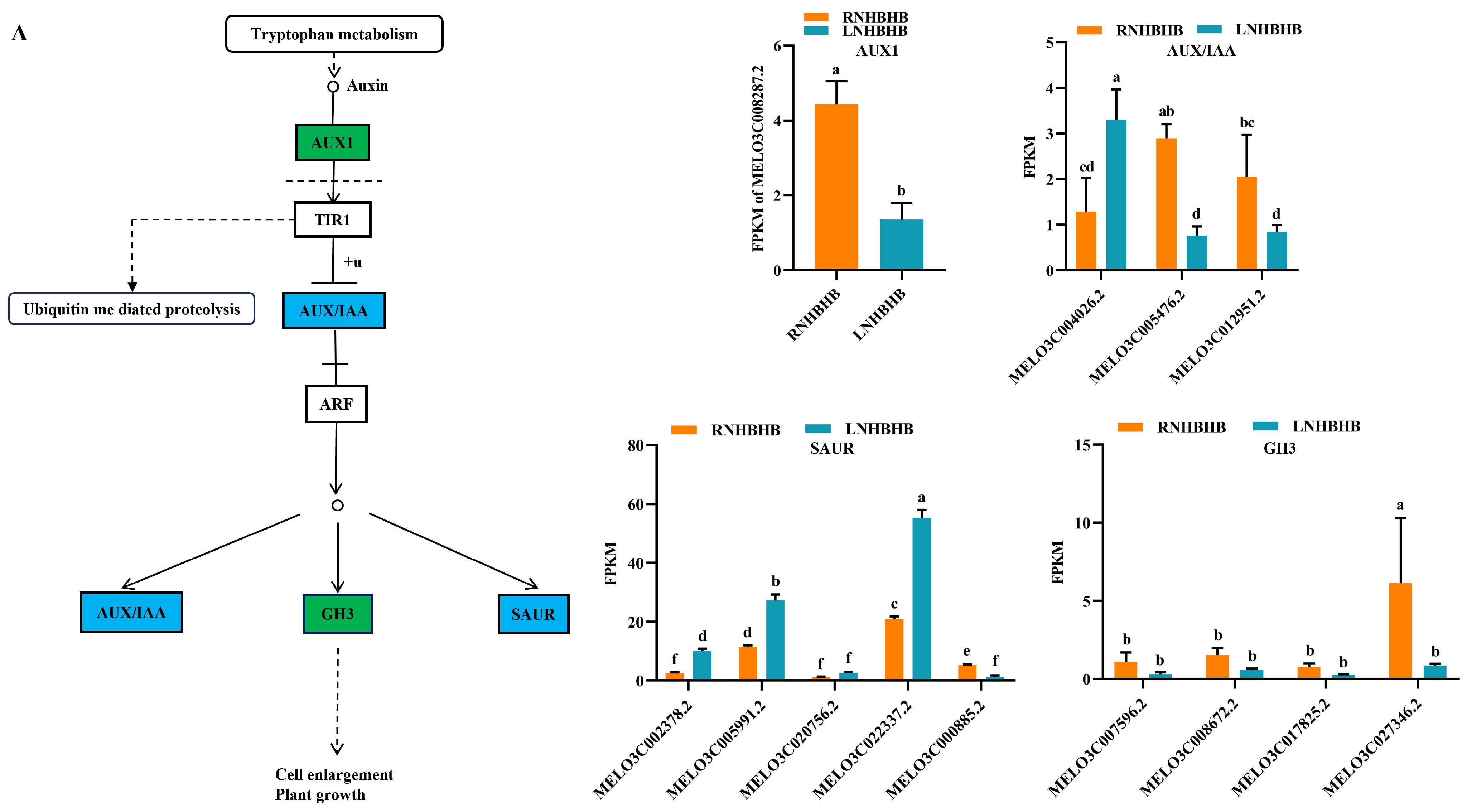
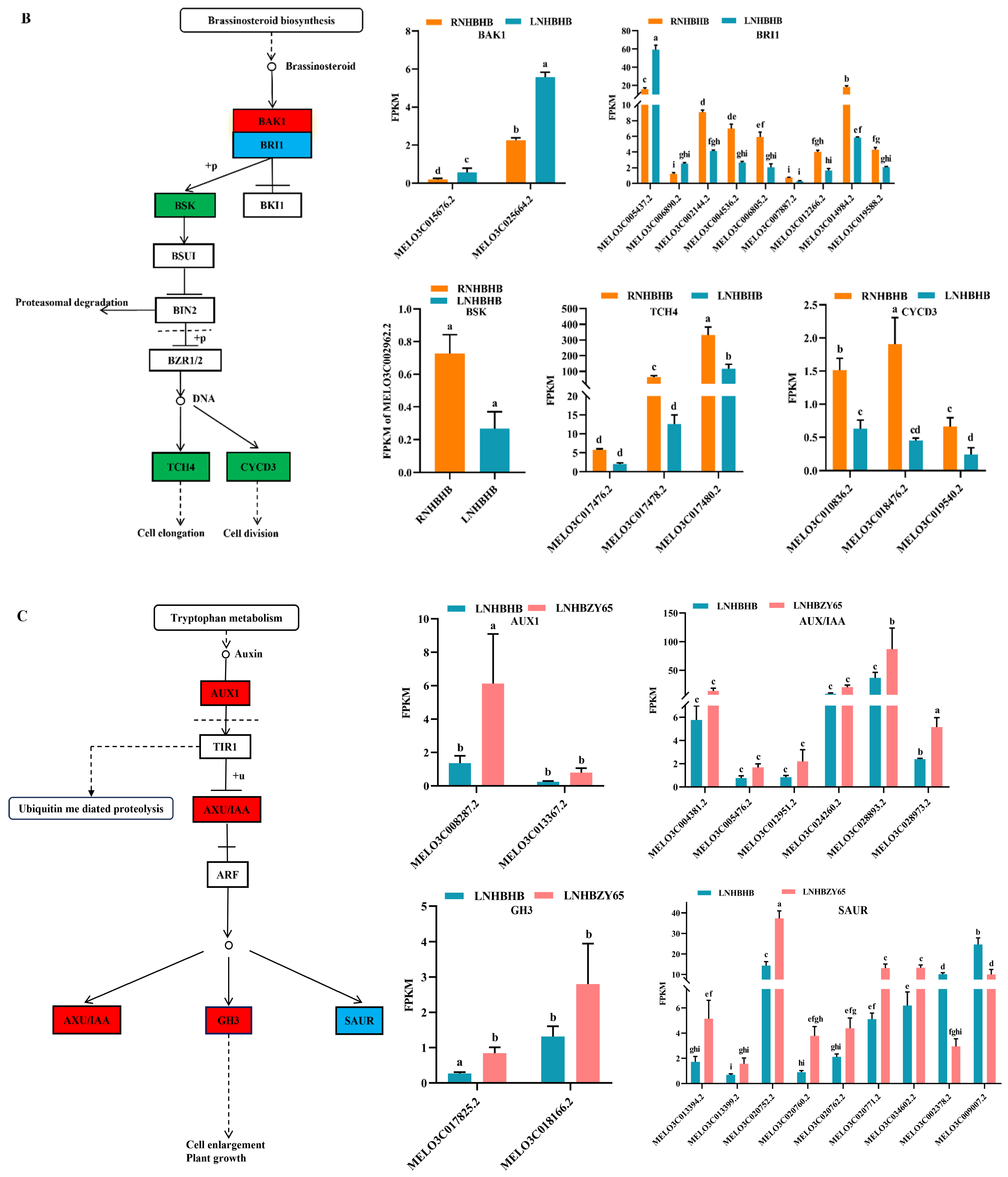
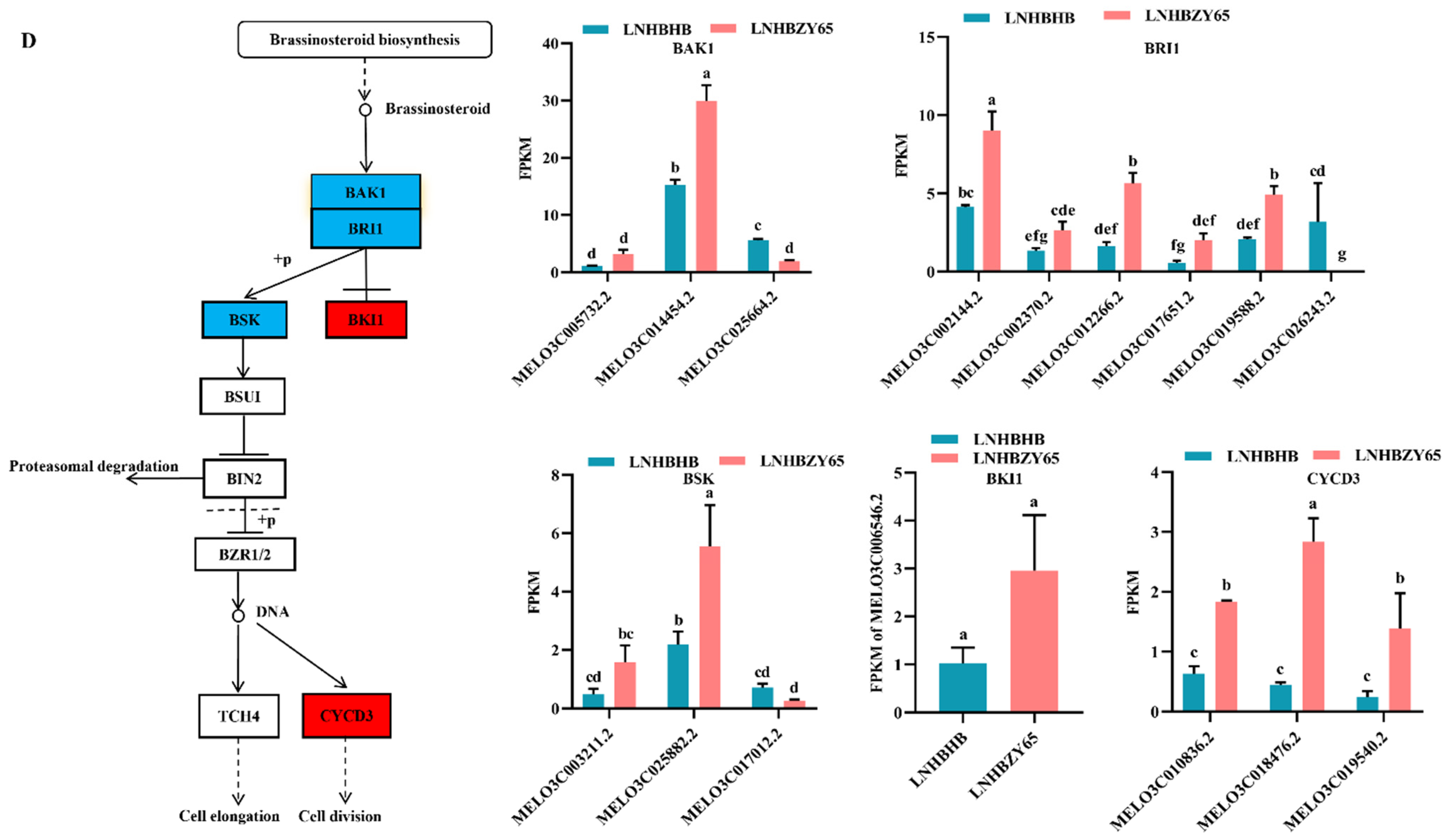
2.4. Differentially Expressed Genes (DEGs) Involved in Signal Transduction
2.5. Differentially Expressed Genes (DEGs) Involved in Oxidative Stress
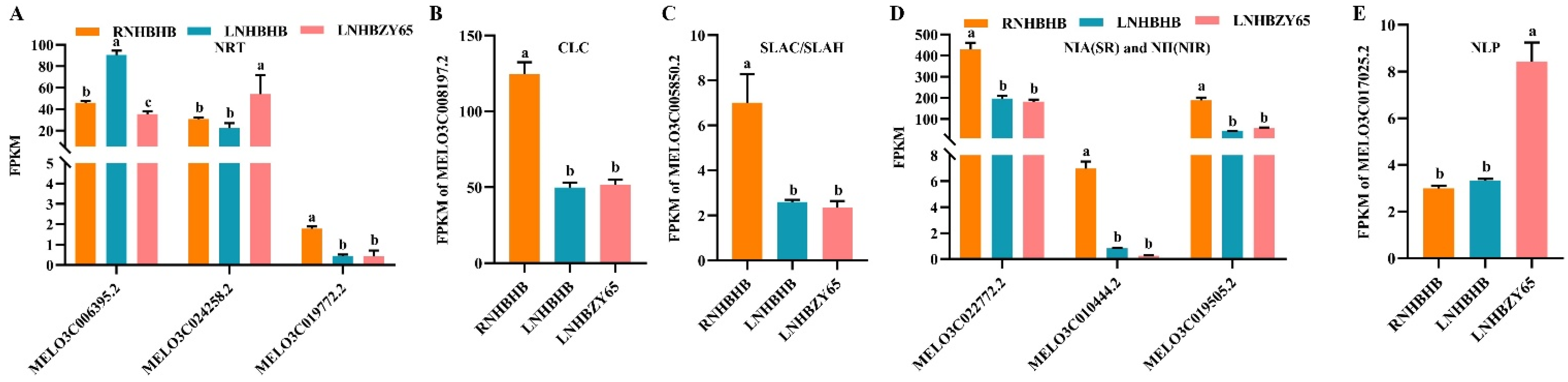
2.6. Differentially Expressed Genes (DEGs) Involved in Nitrogen Metabolism
2.7. Differentially Expressed Genes (DEGs) Involved in Carbon Metabolism
2.8. Differentially Expressed Genes (DEGs) Involved in Secondary Metabolites
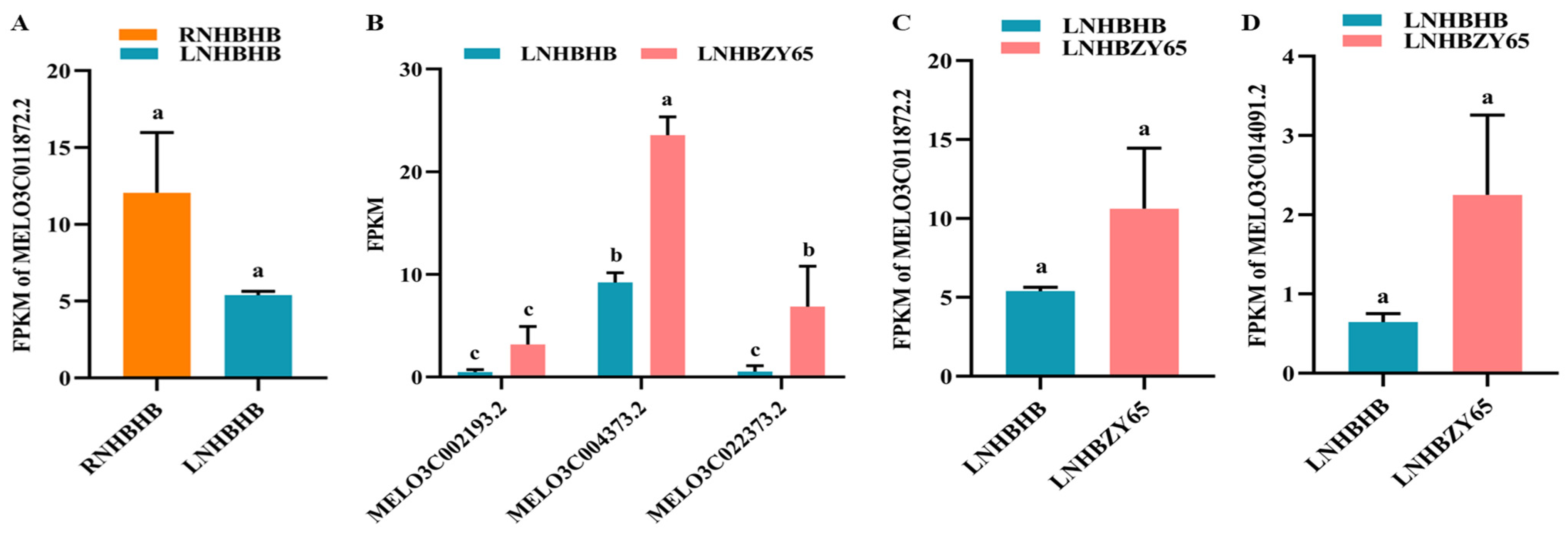
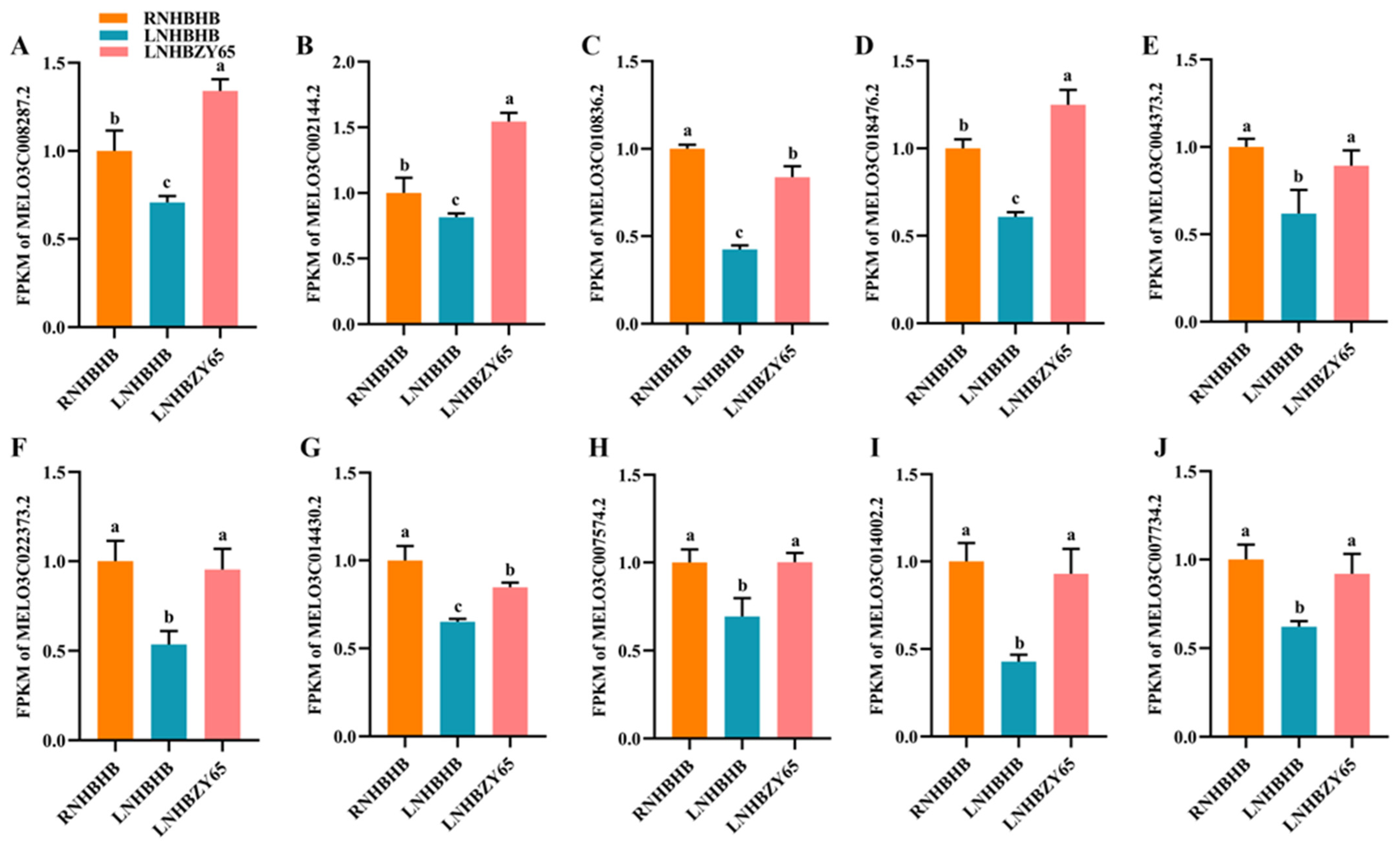
2.9. Validation of RNA-Seq Data by qRT-PCR
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Grafting Methods
4.2. Plant Culture and Nitrogen Treatment
4.3. Determination of Chlorophyll Fluorescence Characteristic Coefficient
4.4. Library Preparation for Transcriptome Sequencing
4.5. Quality Control, Mapping Reads to the Reference Genome, and Annotation
4.6. Quantification of Gene Expression Levels
4.7. Enrichment Analysis of GO Enrichment and KEGG Pathway
4.8. Quantitative Real-Time PCR (qRT-PCR)
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, R.; Tian, M.; Wu, X.; Huang, Y. Differential global gene expression changes in response to low nitrogen stress in two maize inbred lines with contrasting low nitrogen tolerance. Genes Genom. 2011, 33, 491–497. [Google Scholar] [CrossRef]
- Wang, F.; Wang, X.; Zhao, C.; Wang, J.; Li, P.; Dou, Y.; Bi, Y. Alternative pathway is involved in the tolerance of highland barley to the low-nitrogen stress by maintaining the cellular redox homeostasis. Plant Cell Rep. 2016, 35, 317–328. [Google Scholar] [CrossRef]
- Xin, M.; Wang, L.; Liu, Y.; Feng, Z.; Zhou, X.; Qin, Z. Transcriptome profiling of cucumber genome expression in response to long-term low nitrogen stress. Acta Physiol. Plant. 2017, 39, 1–11. [Google Scholar] [CrossRef]
- Rao, I.M.; Miles, J.W.; Beebe, S.E.; Horst, W.J. Root adaptations to soils with low fertility and aluminum toxicity. Ann. Bot. 2016, 118, 593–605. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; He, J. Protective role of anthocyanins in plants under low nitrogen stress. Biochem. Biophys. Res. Commun. 2018, 498, 946–953. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Wang, K.J.; Jiang, G.M.; Biswas, D.K.; Xu, H.; Li, L.F.; Li, Y.H. Effects of nitrogen deficiency on photosynthetic traits of maize hybrids released in different years. Ann. Bot. 2005, 96, 925–930. [Google Scholar] [CrossRef] [PubMed]
- Tattini, M.; Loreto, F.; Fini, A.; Guidi, L.; Brunetti, C.; Velikova, V.; Gori, A.; Ferrini, F. Isoprenoids and phenylpropanoids are part of the antioxidant defense orchestrated daily by drought-stressed P latanus × acerifolia plants during Mediterranean summers. New Phytol. 2015, 207, 613–626. [Google Scholar] [CrossRef] [PubMed]
- Quan, X.; Zeng, J.; Ye, L.; Chen, G.; Han, Z.; Shah, J.M.; Zhang, G. Transcriptome profiling analysis for two Tibetan wild barley genotypes in responses to low nitrogen. BMC Plant Biol. 2016, 16, 30. [Google Scholar] [CrossRef]
- Dong, H.; Li, W.; Eneji, A.E.; Zhang, D. Nitrogen rate and plant density effects on yield and late-season leaf senescence of cotton raised on a saline field. Field Crops Res. 2012, 126, 137–144. [Google Scholar] [CrossRef]
- Lymperopoulos, P.; Msanne, J.; Rabara, R. Phytochrome and phytohormones: Working in tandem for plant growth and development. Front. Plant Sci. 2018, 9, 1037. [Google Scholar] [CrossRef]
- Zhai, L.; Wang, X.; Tang, D.; Qi, Q.; Yer, H.; Jiang, X.; Han, Z.; McAvoy, R.; Li, W.; Li, Y. Molecular and physiological characterization of the effects of auxin-enriched rootstock on grafting. Hortic. Res. 2021, 8, 74. [Google Scholar] [CrossRef] [PubMed]
- Lv, X.; Zhang, Y.; Hu, L.; Zhang, Y.; Zhang, B.; Xia, H.; Du, W.; Fan, S.; Kong, L. Low-nitrogen stress stimulates lateral root initiation and nitrogen assimilation in wheat: Roles of phytohormone signaling. J. Plant Growth Regul. 2021, 40, 436–450. [Google Scholar] [CrossRef]
- Zheng, L.; Ma, J.; Song, C.; An, N.; Zhang, D.; Zhao, C.; Qi, S.; Han, M. Genome-wide identification and expression profiling analysis of brassinolide signal transduction genes regulating apple tree architecture. Acta Physiol. Plant. 2017, 39, 177. [Google Scholar] [CrossRef]
- Zhu, Y.; Guo, J.; Wu, F.; Yu, H.; Min, J.; Zhao, Y.; Xu, C. Genome-Wide Characteristics of GH9B Family Members in Melon and Their Expression Profiles under Exogenous Hormone and Far-Red Light Treatment during the Grafting Healing Process. Int. J. Mol. Sci. 2023, 24, 8258. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.-Y.; Wu, W.; Li, W.-W. Identifying the major pollution sources and pollution loading status of Qiputang River in Taihu Lake basin of China. Desalination Water Treat. 2013, 51, 4736–4743. [Google Scholar] [CrossRef]
- Liang, J.; Chen, X.; Guo, P.; Ren, H.; Xie, Z.; Zhang, Z.; Zhen, A. Grafting improves nitrogen-use efficiency by regulating the nitrogen uptake and metabolism under low-nitrate conditions in cucumber. Sci. Hortic. 2021, 289, 110454. [Google Scholar] [CrossRef]
- Zhang, Z.; Cao, B.; Chen, Z.; Xu, K. Grafting enhances the photosynthesis and nitrogen absorption of tomato plants under low-nitrogen stress. J. Plant Growth Regul. 2021, 41, 1714–1725. [Google Scholar] [CrossRef]
- Zhu, J.-K. Salt and drought stress signal transduction in plants. Annu. Rev. Plant Biol. 2002, 53, 247–273. [Google Scholar] [CrossRef] [PubMed]
- Halliday, K.J.; Martínez-García, J.F.; Josse, E.-M. Integration of light and auxin signaling. Cold Spring Harb. Perspect. Biol. 2009, 1, a001586. [Google Scholar] [CrossRef]
- Grossman, A.; Takahashi, H. Macronutrient utilization by photosynthetic eukaryotes and the fabric of interactions. Annu. Rev. Plant Biol. 2001, 52, 163–210. [Google Scholar] [CrossRef]
- Raja, V.; Majeed, U.; Kang, H.; Andrabi, K.I.; John, R. Abiotic stress: Interplay between ROS, hormones and MAPKs. Environ. Exp. Bot. 2017, 137, 142–157. [Google Scholar] [CrossRef]
- Liu, T.; Cao, L.; Cheng, Y.; Ji, J.; Wei, Y.; Wang, C.; Duan, K. MKK4/5-MPK3/6 cascade regulates agrobacterium-mediated transformation by modulating plant immunity in Arabidopsis. Front. Plant Sci. 2021, 12, 731690. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Chai, G.; Xu, L.; Yang, K.; Zhuang, Y.; Yang, A.; Liu, S.; Kong, Y.; Zhou, G. Phosphorylation-mediated inactivation of C3H14 by MPK4 enhances bacterial-triggered immunity in Arabidopsis. Plant Physiol. 2022, 190, 1941–1959. [Google Scholar] [CrossRef] [PubMed]
- Lampard, G.R.; Lukowitz, W.; Ellis, B.E.; Bergmann, D.C. Novel and expanded roles for MAPK signaling in Arabidopsis stomatal cell fate revealed by cell type–specific manipulations. Plant Cell 2009, 21, 3506–3517. [Google Scholar] [CrossRef] [PubMed]
- Frink, C.R.; Waggoner, P.E.; Ausubel, J.H. Nitrogen fertilizer: Retrospect and prospect. Proc. Natl. Acad. Sci. USA 1999, 96, 1175–1180. [Google Scholar] [CrossRef] [PubMed]
- Krapp, A.; David, L.C.; Chardin, C.; Girin, T.; Marmagne, A.; Leprince, A.-S.; Chaillou, S.; Ferrario-Méry, S.; Meyer, C.; Daniel-Vedele, F. Nitrate transport and signalling in Arabidopsis. J. Exp. Bot. 2014, 65, 789–798. [Google Scholar] [CrossRef] [PubMed]
- Cui, G.; Zhang, Y.; Zhang, W.; Lang, D.; Zhang, X.; Li, Z.; Zhang, X. Response of carbon and nitrogen metabolism and secondary metabolites to drought stress and salt stress in plants. J. Plant Biol. 2019, 62, 387–399. [Google Scholar] [CrossRef]
- Zhao, D.; Reddy, K.R.; Kakani, V.G.; Reddy, V. Nitrogen deficiency effects on plant growth, leaf photosynthesis, and hyperspectral reflectance properties of sorghum. Eur. J. Agron. 2006, 22, 391–403. [Google Scholar] [CrossRef]
- Quan, N.T.; Anh, L.H.; Khang, D.T.; Tuyen, P.T.; Toan, N.P.; Minh, T.N.; Minh, L.T.; Bach, D.T.; Ha, P.T.T.; Elzaawely, A.A.; et al. Involvement of secondary metabolites in response to drought stress of rice (Oryza sativa L.). Agriculture 2016, 6, 23. [Google Scholar] [CrossRef]
- Khare, S.; Singh, N.; Singh, A.; Hussain, I.; Niharika, K.; Yadav, V.; Bano, C.; Yadav, R.K.; Amist, N. Plant secondary metabolites synthesis and their regulations under biotic and abiotic constraints. J. Plant Biol. 2020, 63, 203–216. [Google Scholar] [CrossRef]
- Wang, J.; Song, K.; Sun, L.; Qin, Q.; Sun, Y.; Pan, J.; Xue, Y. Morphological and transcriptome analysis of wheat seedlings response to low nitrogen stress. Plants 2019, 8, 98. [Google Scholar] [CrossRef] [PubMed]
- Tantray, A.Y.; Bashir, S.S.; Ahmad, A. Low nitrogen stress regulates chlorophyll fluorescence in coordination with photosynthesis and Rubisco efficiency of rice. Physiol. Mol. Biol. Plants 2020, 26, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Krouk, G.; Lacombe, B.; Bielach, A.; Perrine-Walker, F.; Malinska, K.; Mounier, E.; Hoyerova, K.; Tillard, P.; Leon, S.; Ljung, K.; et al. Nitrate-regulated auxin transport by NRT1. 1 defines a mechanism for nutrient sensing in plants. Dev. Cell 2010, 18, 927–937. [Google Scholar] [CrossRef] [PubMed]
- LIANG, Z.-m.; CAO, X.-d.; Rong, G.; Nian, G.; TANG, Y.-y.; Vinay, N.; Yang, L. Brassinosteroids alleviates wheat floret degeneration under low nitrogen stress by promoting the distribution of sucrose from stems to spikes. J. Integr. Agric. 2023. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.; Song, G.; Bae, H. Suppressive effect of fraxetin on adipogenesis and reactive oxygen species production in 3T3-L1 cells by regulating MAPK signaling pathways. Antioxidants 2022, 11, 1893. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, B.; Song, Y.; Chen, Y.; Fu, J.; Liu, J.; Ma, T.; Xi, Z.; Liu, H. Genome-wide (ChIP-seq) identification of target genes regulated by WRKY33 during submergence stress in Arabidopsis. BMC Genom. Data 2021, 22, 16. [Google Scholar] [CrossRef] [PubMed]
- Colla, G.; Suárez, C.M.C.; Cardarelli, M.; Rouphael, Y. Improving nitrogen use efficiency in melon by grafting. HortScience 2010, 45, 559–565. [Google Scholar] [CrossRef]
- Hou, S.A.; Zhu, Y.; Wu, X.; Xin, Y.; Guo, J.; Wu, F.; Yu, H.; Sun, Z.; Xu, C. Scion-to-rootstock mobile transcription factor CmHY5 positively modulates the nitrate uptake capacity of melon scion grafted on squash rootstock. Int. J. Mol. Sci. 2022, 24, 162. [Google Scholar] [CrossRef]
- Iqbal, A.; Dong, Q.; Wang, X.; Gui, H.; Zhang, H.; Zhang, X.; Song, M. Transcriptome analysis reveals differences in key genes and pathways regulating carbon and nitrogen metabolism in cotton genotypes under N starvation and resupply. Int. J. Mol. Sci. 2020, 21, 1500. [Google Scholar] [CrossRef]
- Diaz, C.; Saliba-Colombani, V.; Loudet, O.; Belluomo, P.; Moreau, L.; Daniel-Vedele, F.; Morot-Gaudry, J.-F.; Masclaux-Daubresse, C. Leaf yellowing and anthocyanin accumulation are two genetically independent strategies in response to nitrogen limitation in Arabidopsis thaliana. Plant Cell Physiol. 2006, 47, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.R.; Perkins-Veazie, P.; Sakata, Y.; Lopez-Galarza, S.; Maroto, J.V.; Lee, S.-G.; Huh, Y.-C.; Sun, Z.; Miguel, A.; King, S.R.; et al. Cucurbit grafting. Crit. Rev. Plant Sci. 2008, 27, 50–74. [Google Scholar] [CrossRef]
- Demmig-Adams, B.; Adams III, W.W.; Barker, D.H.; Logan, B.A.; Bowling, D.R.; Verhoeven, A.S. Using chlorophyll fluorescence to assess the fraction of absorbed light allocated to thermal dissipation of excess excitation. Physiol. Plant. 1996, 98, 253–264. [Google Scholar] [CrossRef]
- Xu, C.; Zhang, Y.; Zhao, M.; Liu, Y.; Xu, X.; Li, T. Transcriptomic analysis of melon/squash graft junction reveals molecular mechanisms potentially underlying the graft union development. PeerJ 2021, 9, e12569. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.; Binns, D.; Chang, H.; Fraser, M.; Li, W.; McAnulla, C.; McWilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G.; et al. InterProScan 5: Genome-scale protein function classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef] [PubMed]
- Chou, C.-H.; Huang, H.-Y.; Huang, W.-C.; Hsu, S.-D.; Hsiao, C.-D.; Liu, C.-Y.; Chen, Y.-H.; Liu, Y.-C.; Lee, M.-L.; Chen, Y.-C.; et al. The aquatic animals’ transcriptome resource for comparative functional analysis. BMC Genom. 2018, 19, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The protein families database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef] [PubMed]
- Potter, S.C.; Luciani, A.; Eddy, S.R.; Park, Y.; López, R.; Finn, R.D. HMMER web server: 2018 update. Nucleic Acids Res. 2018, 46, W200–W204. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Zhang, A.; Han, D.; Wang, Y.; Mu, H.; Zhang, T.; Yan, X.; Pang, Q. Transcriptomic and proteomic feature of salt stress-regulated network in Jerusalem artichoke (Helianthus tuberosus L.) root based on de novo assembly sequencing analysis. Planta 2018, 247, 715–732. [Google Scholar] [PubMed]
- Kanehisa, M.; Goto, S.; Kawashima, S.; Okuno, Y.; Hattori, M. The KEGG resource for deciphering the genome. Nucleic Acids Res. 2004, 32, D277–D280. [Google Scholar] [CrossRef] [PubMed]
- Pattabiraman, S.; Warnow, T. Profile hidden markov models are not identifiable. IEEE/ACM Trans. Comput. Biol. Bioinform. 2021, 18, 162–172. [Google Scholar] [CrossRef] [PubMed]
- Tak Leung, R.W.; Jiang, X.; Chu, K.H.; Qin, J. ENPD-A database of eukaryotic nucleic acid binding proteins: Linking gene regulations to proteins. Nucleic Acids Res. 2019, 47, D322–D329. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| #Sample ID | Obtained Reads | Obtained Base (bp) | Q20 (%) | Q30 (%) | GC (%) |
|---|---|---|---|---|---|
| LNHBHB 1 | 38697041 | 11534617118 | 99.07 | 94.04 | 44.89 |
| LNHBHB 2 | 37251629 | 11094248198 | 99.05 | 93.93 | 45.00 |
| LNHBHB 3 | 36228540 | 10801550260 | 99.09 | 94.15 | 44.85 |
| RNHBHB 1 | 27329828 | 8144594918 | 98.92 | 93.14 | 45.04 |
| RNHBHB 2 | 21551946 | 6410892762 | 99.11 | 94.28 | 44.94 |
| RNHBHB 3 | 34739882 | 10352846224 | 99.05 | 93.95 | 45.10 |
| LNHBZY65 1 | 32237175 | 9538969616 | 99.24 | 95.04 | 44.91 |
| LNHBZY65 2 | 37489331 | 11158639468 | 99.05 | 93.94 | 45.39 |
| LNHBZY65 3 | 20393405 | 6074389982 | 99.16 | 94.59 | 45.01 |
| Treatment | Ca(NO3)2·4H2O | KNO3 | NH4H2PO4 | K2SO4 | MgSO4·7H2O |
|---|---|---|---|---|---|
| RN | 944.00 | 0 | 114.00 | 522.00 | 492.00 |
| LN | 47.20 | 0 | 114.00 | 522.00 | 492.00 |
| Chemical Name | Grams of Compounds per Liter of Water |
|---|---|
| EDTA·Na2 | 18.60 |
| H3BO3 | 2.86 |
| MnSO4·H2O | 2.13 |
| ZnSO4·7H2O | 0.22 |
| CuSO4·5H2O | 0.08 |
| (NH4)6Mo7O24·4H2O | 0.02 |
| FeSO4·7H2O | 13.90 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, Y.; Sun, Z.; Wu, H.; Cui, C.; Meng, S.; Xu, C. Transcriptomic Analysis of the Molecular Mechanism Potential of Grafting—Enhancing the Ability of Oriental Melon to Tolerate Low-Nitrogen Stress. Int. J. Mol. Sci. 2024, 25, 8227. https://doi.org/10.3390/ijms25158227
Zhu Y, Sun Z, Wu H, Cui C, Meng S, Xu C. Transcriptomic Analysis of the Molecular Mechanism Potential of Grafting—Enhancing the Ability of Oriental Melon to Tolerate Low-Nitrogen Stress. International Journal of Molecular Sciences. 2024; 25(15):8227. https://doi.org/10.3390/ijms25158227
Chicago/Turabian StyleZhu, Yulei, Ziqing Sun, Hongxi Wu, Caifeng Cui, Sida Meng, and Chuanqiang Xu. 2024. "Transcriptomic Analysis of the Molecular Mechanism Potential of Grafting—Enhancing the Ability of Oriental Melon to Tolerate Low-Nitrogen Stress" International Journal of Molecular Sciences 25, no. 15: 8227. https://doi.org/10.3390/ijms25158227
APA StyleZhu, Y., Sun, Z., Wu, H., Cui, C., Meng, S., & Xu, C. (2024). Transcriptomic Analysis of the Molecular Mechanism Potential of Grafting—Enhancing the Ability of Oriental Melon to Tolerate Low-Nitrogen Stress. International Journal of Molecular Sciences, 25(15), 8227. https://doi.org/10.3390/ijms25158227