
A Common Polymorphism in RNASE6 Impacts Its Antimicrobial Activity toward Uropathogenic Escherichia coli
 , ,
, , 
Abstract
1. Introduction
2. Results
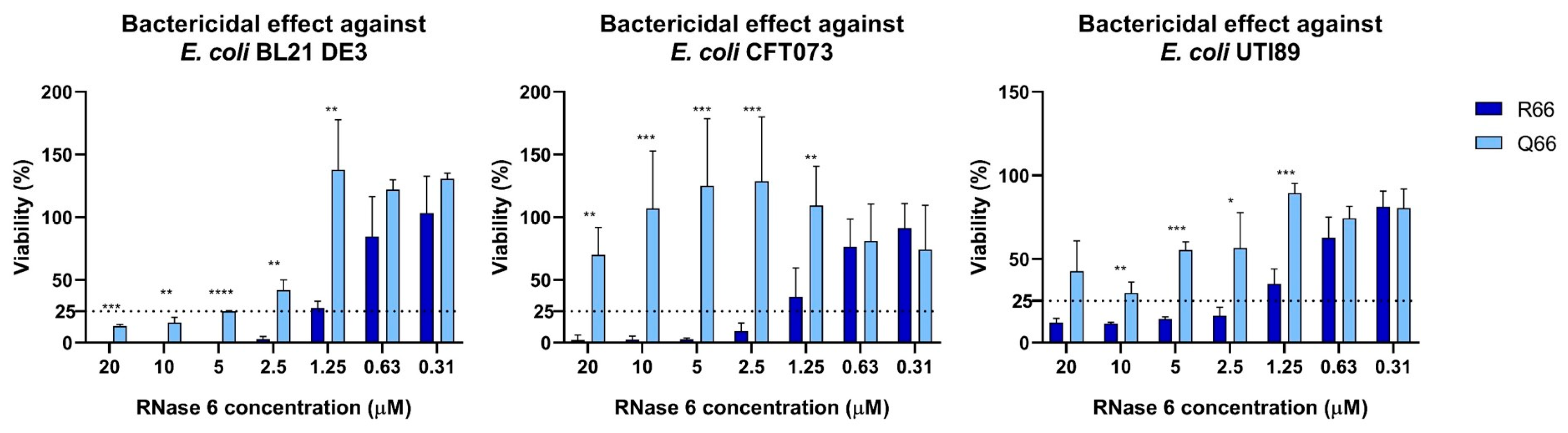
2.1. RNASE6 rs1045922 Is a Common, Non-Synonymous SNP That Alters RNase 6 Antimicrobial Activity
2.2. Both SNP RNase6-R66 and -Q66 Display the Same Catalytic Activity
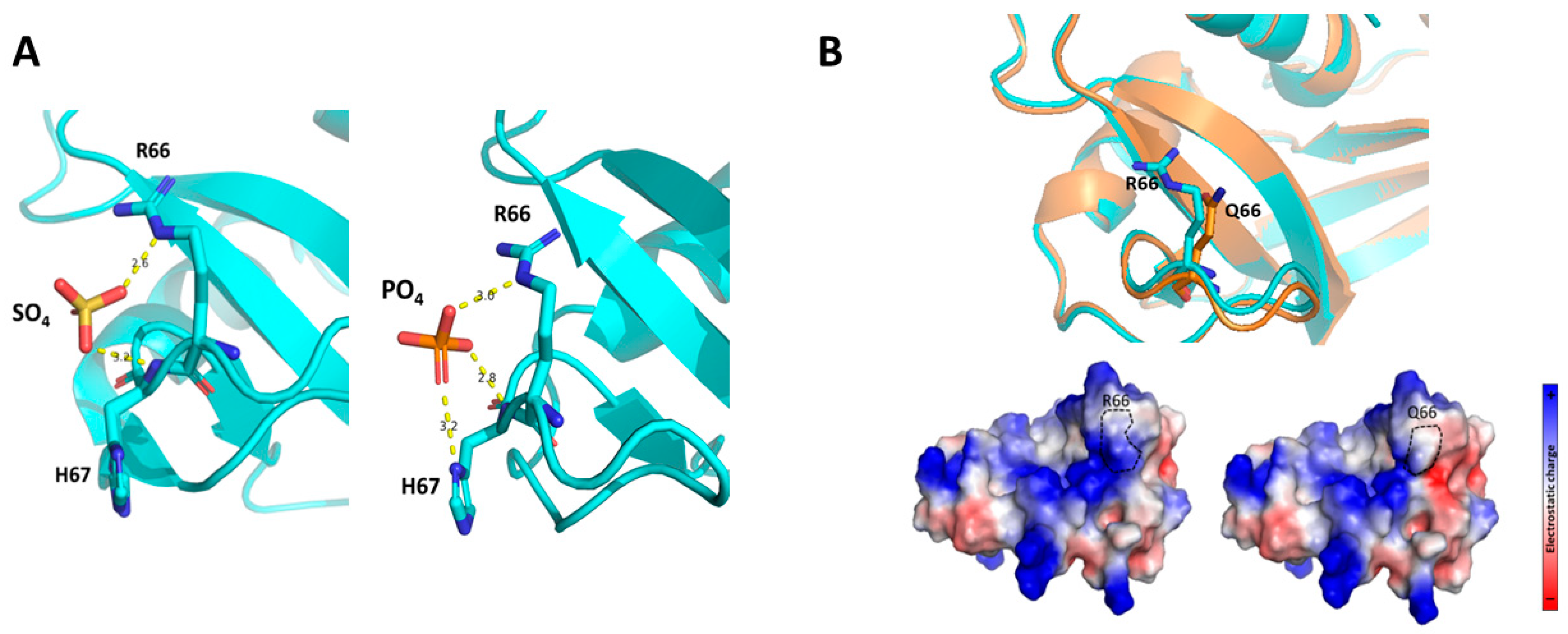
2.3. The RNase6 R66Q Substitution Significantly Reduces the Positive Electrostatic Charge at the Protein Surface
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Expression of RNase6-R66 and -Q66 Variants
4.3. Minimum Bactericidal Concentration (MBC) Determination
4.4. Lipopolysaccharide Binding Assay
4.5. Bacterial Agglutination Assay
4.6. Spectrophotometric Kinetic Assay
4.7. 3D Structure Modelling
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Flores-Mireles, A.L.; Walker, J.N.; Caparon, M.; Hultgren, S.J. Urinary tract infections: Epidemiology, mechanisms of infection and treatment options. Nat. Rev. Microbiol. 2015, 13, 269–284. [Google Scholar] [CrossRef] [PubMed]
- Medina, M.; Castillo-Pino, E. An introduction to the epidemiology and burden of urinary tract infections. Ther. Adv. Urol. 2019, 11, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Foxman, B. Urinary tract infection syndromes: Occurrence, recurrence, bacteriology, risk factors, and disease burden. Infect. Dis. Clin. North. Am. 2014, 28, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.R.; Russo, T.A. Acute Pyelonephritis in Adults. N. Engl. J. Med. 2018, 378, 48–59. [Google Scholar] [CrossRef]
- Becknell, B.; Schober, M.; Korbel, L.; Spencer, J.D. The diagnosis, evaluation and treatment of acute and recurrent pediatric urinary tract infections. Expert. Rev. Anti Infect. Ther. 2015, 13, 81–90. [Google Scholar] [CrossRef]
- Korbel, L.; Howell, M.; Spencer, J.D. The clinical diagnosis and management of urinary tract infections in children and adolescents. Paediatr. Int. Child. Health 2017, 37, 273–279. [Google Scholar] [CrossRef]
- Paul, R. State of the Globe: Rising Antimicrobial Resistance of Pathogens in Urinary Tract Infection. J. Glob. Infect. Dis. 2018, 10, 117–118. [Google Scholar] [CrossRef]
- Abbott, I.J.; Peel, T.N.; Cairns, K.A.; Stewardson, A.J. Antibiotic management of urinary tract infections in the post-antibiotic era: A narrative review highlighting diagnostic and antimicrobial stewardship. Clin. Microbiol. Infect. 2023, 29, 1254–1266. [Google Scholar] [CrossRef]
- Kuhn, H.W.; Hreha, T.N.; Hunstad, D.A. Immune defenses in the urinary tract. Trends Immunol. 2023, 44, 701–711. [Google Scholar] [CrossRef]
- Ching, C.; Schwartz, L.; Spencer, J.D.; Becknell, B. Innate immunity and urinary tract infection. Pediatr. Nephrol. 2020, 35, 1183–1192. [Google Scholar] [CrossRef]
- Lacerda Mariano, L.; Ingersoll, M.A. The immune response to infection in the bladder. Nat. Rev. Urol. 2020, 17, 439–458. [Google Scholar] [CrossRef] [PubMed]
- Becknell, B.; Schwaderer, A.; Hains, D.S.; Spencer, J.D. Amplifying renal immunity: The role of antimicrobial peptides in pyelonephritis. Nat. Rev. Nephrol. 2015, 11, 642–655. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.S.; Townes, C.L.; Hall, J.; Pickard, R.S. Maintaining a sterile urinary tract: The role of antimicrobial peptides. J. Urol. 2009, 182, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Lazzaro, B.P.; Zasloff, M.; Rolff, J. Antimicrobial peptides: Application informed by evolution. Science 2020, 368, eaau5480. [Google Scholar] [CrossRef] [PubMed]
- Becknell, B.; Ching, C.; Spencer, J.D. The Responses of the Ribonuclease A Superfamily to Urinary Tract Infection. Front. Immunol. 2019, 10, 2786. [Google Scholar] [CrossRef] [PubMed]
- Becknell, B.; Eichler, T.E.; Beceiro, S.; Li, B.; Easterling, R.S.; Carpenter, A.R.; James, C.L.; McHugh, K.M.; Hains, D.S.; Partida-Sanchez, S.; et al. Ribonucleases 6 and 7 have antimicrobial function in the human and murine urinary tract. Kidney Int. 2015, 87, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Mishra, B.; Lau, K.; Lushnikova, T.; Golla, R.; Wang, X. Antimicrobial peptides in 2014. Pharmaceuticals 2015, 8, 123–150. [Google Scholar] [CrossRef]
- Chromek, M.; Slamova, Z.; Bergman, P.; Kovacs, L.; Podracka, L.; Ehren, I.; Hokfelt, T.; Gudmundsson, G.H.; Gallo, R.L.; Agerberth, B.; et al. The antimicrobial peptide cathelicidin protects the urinary tract against invasive bacterial infection. Nat. Med. 2006, 12, 636–641. [Google Scholar] [CrossRef]
- Spencer, J.D.; Schwaderer, A.L.; Wang, H.; Bartz, J.; Kline, J.; Eichler, T.; DeSouza, K.R.; Sims-Lucas, S.; Baker, P.; Hains, D.S. Ribonuclease 7, an antimicrobial peptide upregulated during infection, contributes to microbial defense of the human urinary tract. Kidney Int. 2013, 83, 615–625. [Google Scholar] [CrossRef]
- Forster, C.S.; Johnson, K.; Patel, V.; Wax, R.; Rodig, N.; Barasch, J.; Bachur, R.; Lee, R.S. Urinary NGAL deficiency in recurrent urinary tract infections. Pediatr. Nephrol. 2017, 32, 1077–1080. [Google Scholar] [CrossRef]
- Eichler, T.; Bender, K.; Murtha, M.J.; Schwartz, L.; Metheny, J.; Solden, L.; Jaggers, R.M.; Bailey, M.T.; Gupta, S.; Mosquera, C.; et al. Ribonuclease 7 Shields the Kidney and Bladder from Invasive Uropathogenic Escherichia coli Infection. J. Am. Soc. Nephrol. 2019, 30, 1385–1397. [Google Scholar] [CrossRef] [PubMed]
- Bender, K.; Schwartz, L.L.; Cohen, A.; Vasquez, C.M.; Murtha, M.J.; Eichler, T.; Thomas, J.P.; Jackson, A.; Spencer, J.D. Expression and function of human ribonuclease 4 in the kidney and urinary tract. Am. J. Physiol. Renal Physiol. 2021, 320, F972–F983. [Google Scholar] [CrossRef] [PubMed]
- Hains, D.S.; Polley, S.; Liang, D.; Saxena, V.; Arregui, S.; Ketz, J.; Barr-Beare, E.; Rawson, A.; Spencer, J.D.; Cohen, A.; et al. Deleted in malignant brain tumor 1 genetic variation confers urinary tract infection risk in children and mice. Clin. Transl. Med. 2021, 11, e477. [Google Scholar] [CrossRef] [PubMed]
- Schwaderer, A.L.; Wang, H.; Kim, S.; Kline, J.M.; Liang, D.; Brophy, P.D.; McHugh, K.M.; Tseng, G.C.; Saxena, V.; Barr-Beare, E.; et al. Polymorphisms in alpha-Defensin-Encoding DEFA1A3 Associate with Urinary Tract Infection Risk in Children with Vesicoureteral Reflux. J. Am. Soc. Nephrol. 2016, 27, 3175–3186. [Google Scholar] [CrossRef] [PubMed]
- Pierce, K.R.; Eichler, T.; Mosquera Vasquez, C.; Schwaderer, A.L.; Simoni, A.; Creacy, S.; Hains, D.S.; Spencer, J.D. Ribonuclease 7 polymorphism rs1263872 reduces antimicrobial activity and associates with pediatric urinary tract infections. J. Clin. Investig. 2021, 131. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, H.F.; Dyer, K.D. Molecular cloning and characterization of a novel human ribonuclease (RNase k6): Increasing diversity in the enlarging ribonuclease gene family. Nucleic Acids Res. 1996, 24, 3507–3513. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Li, J.; Moussaoui, M.; Boix, E. Immune Modulation by Human Secreted RNases at the Extracellular Space. Front. Immunol. 2018, 9, 1012. [Google Scholar] [CrossRef]
- Pulido, D.; Arranz-Trullen, J.; Prats-Ejarque, G.; Velazquez, D.; Torrent, M.; Moussaoui, M.; Boix, E. Insights into the Antimicrobial Mechanism of Action of Human RNase6: Structural Determinants for Bacterial Cell Agglutination and Membrane Permeation. Int. J. Mol. Sci. 2016, 17, 552. [Google Scholar] [CrossRef]
- Ruiz-Rosado, J.D. Human Ribonuclease 6 has a Protective Role During Experimental Urinary Tract Infection. J. Innate Immun. 2023, 15, 865–875. [Google Scholar] [CrossRef]
- Genomes Project, C.; Auton, A.; Brooks, L.D.; Durbin, R.M.; Garrison, E.P.; Kang, H.M.; Korbel, J.O.; Marchini, J.L.; McCarthy, S.; McVean, G.A.; et al. A global reference for human genetic variation. Nature 2015, 526, 68–74. [Google Scholar]
- Mulvey, M.A.; Schilling, J.D.; Hultgren, S.J. Establishment of a persistent Escherichia coli reservoir during the acute phase of a bladder infection. Infect. Immun. 2001, 69, 4572–4579. [Google Scholar] [CrossRef] [PubMed]
- Mobley, H.L.; Green, D.M.; Trifillis, A.L.; Johnson, D.E.; Chippendale, G.R.; Lockatell, C.V.; Jones, B.D.; Warren, J.W. Pyelonephritogenic Escherichia coli and killing of cultured human renal proximal tubular epithelial cells: Role of hemolysin in some strains. Infect. Immun. 1990, 58, 1281–1289. [Google Scholar] [CrossRef] [PubMed]
- Prats-Ejarque, G.; Arranz-Trullen, J.; Blanco, J.A.; Pulido, D.; Nogues, M.V.; Moussaoui, M.; Boix, E. The first crystal structure of human RNase 6 reveals a novel substrate-binding and cleavage site arrangement. Biochem. J. 2016, 473, 1523–1536. [Google Scholar] [CrossRef] [PubMed]
- Prats-Ejarque, G.; Blanco, J.A.; Salazar, V.A.; Nogues, V.M.; Moussaoui, M.; Boix, E. Characterization of an RNase with two catalytic centers. Human RNase6 catalytic and phosphate-binding site arrangement favors the endonuclease cleavage of polymeric substrates. Biochim. Biophys. Acta Gen. Subj. 2019, 1863, 105–117. [Google Scholar] [CrossRef]
- Bryant, P.; Pozzati, G.; Elofsson, A. Improved prediction of protein-protein interactions using AlphaFold2. Nat. Commun. 2022, 13, 1265. [Google Scholar] [CrossRef]
- Rivas-Santiago, B.; Serrano, C.J.; Enciso-Moreno, J.A. Susceptibility to infectious diseases based on antimicrobial peptide production. Infect. Immun. 2009, 77, 4690–4695. [Google Scholar] [CrossRef]
- Eriksson, J.; Reimert, C.M.; Kabatereine, N.B.; Kazibwe, F.; Ireri, E.; Kadzo, H.; Eltahir, H.B.; Mohamed, A.O.; Vennervald, B.J.; Venge, P. The 434(G>C) polymorphism within the coding sequence of Eosinophil Cationic Protein (ECP) correlates with the natural course of Schistosoma mansoni infection. Int. J. Parasitol. 2007, 37, 1359–1366. [Google Scholar] [CrossRef]
- Jonsson, U.B.; Bystrom, J.; Stalenheim, G.; Venge, P. Polymorphism of the eosinophil cationic protein-gene is related to the expression of allergic symptoms. Clin. Exp. Allergy 2002, 32, 1092–1095. [Google Scholar] [CrossRef]
- McLaughlin, R.L.; Phukan, J.; McCormack, W.; Lynch, D.S.; Greenway, M.; Cronin, S.; Saunders, J.; Slowik, A.; Tomik, B.; Andersen, P.M.; et al. Angiogenin levels and ANG genotypes: Dysregulation in amyotrophic lateral sclerosis. PLoS ONE 2010, 5, e15402. [Google Scholar] [CrossRef]
- Boix, E.; Salazar, V.A.; Torrent, M.; Pulido, D.; Nogues, M.V.; Moussaoui, M. Structural determinants of the eosinophil cationic protein antimicrobial activity. Biol. Chem. 2012, 393, 801–815. [Google Scholar] [CrossRef]
- Pulido, D.; Garcia-Mayoral, M.F.; Moussaoui, M.; Velazquez, D.; Torrent, M.; Bruix, M.; Boix, E. Structural basis for endotoxin neutralization by the eosinophil cationic protein. FEBS J. 2016, 283, 4176–4191. [Google Scholar] [CrossRef] [PubMed]
- Pulido, D.; Moussaoui, M.; Andreu, D.; Nogues, M.V.; Torrent, M.; Boix, E. Antimicrobial action and cell agglutination by the eosinophil cationic protein are modulated by the cell wall lipopolysaccharide structure. Antimicrob. Agents Chemother. 2012, 56, 2378–2385. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, C.; Bernard, D.N.; Letourneau, M.; Gagnon, J.; Gagne, D.; Bafna, K.; Calmettes, C.; Couture, J.F.; Agarwal, P.K.; Doucet, N. Insights into Structural and Dynamical Changes Experienced by Human RNase 6 upon Ligand Binding. Biochemistry 2020, 59, 755–765. [Google Scholar] [CrossRef] [PubMed]
- Prats-Ejarque, G.; Lu, L.; Salazar, V.A.; Moussaoui, M.; Boix, E. Evolutionary Trends in RNA Base Selectivity Within the RNase A Superfamily. Front. Pharmacol. 2019, 10, 1170. [Google Scholar] [CrossRef] [PubMed]
- Deming, M.S.; Dyer, K.D.; Bankier, A.T.; Piper, M.B.; Dear, P.H.; Rosenberg, H.F. Ribonuclease k6: Chromosomal mapping and divergent rates of evolution within the RNase A gene superfamily. Genome Res. 1998, 8, 599–607. [Google Scholar] [CrossRef] [PubMed]
- Ashkenazy, H.; Abadi, S.; Martz, E.; Chay, O.; Mayrose, I.; Pupko, T.; Ben-Tal, N. ConSurf 2016: An improved methodology to estimate and visualize evolutionary conservation in macromolecules. Nucleic Acids Res 2016, 44, W344–W350. [Google Scholar] [CrossRef]
- Prats-Ejarque, G.; Lorente, H.; Villalba, C.; Anguita, R.; Lu, L.; Vazquez-Monteagudo, S.; Fernandez-Millan, P.; Boix, E. Structure-Based Design of an RNase Chimera for Antimicrobial Therapy. Int. J. Mol. Sci. 2021, 23, 95. [Google Scholar] [CrossRef]
- Torrent, M.; Navarro, S.; Moussaoui, M.; Nogues, M.V.; Boix, E. Eosinophil cationic protein high-affinity binding to bacteria-wall lipopolysaccharides and peptidoglycans. Biochemistry 2008, 47, 3544–3555. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Zidek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| E. coli BL21 | E. coli CF073 | E. coli UTI89 | ||||
|---|---|---|---|---|---|---|
| MBC100 | MBC75 | MBC100 | MBC75 | MBC100 | MBC75 | |
| RNase 6-R66 | 8.33 (±2.9) | 2.08 (±0.72) | >20 | 1.87 (±0.68) | >20 | 3.33 (±1.44) |
| RNase 6-Q66 | >20 | 8.22 (±2.9) | >20 | >20 | >20 | >20 |
| LPS Binding | MAC | ||
|---|---|---|---|
| EC50 (μM) | Max (%) | (µM) | |
| RNase 6-R66 | 2.64 ± 0.23 | 75.90 ± 4.41 | 0.22 ± 0.05 |
| RNase 6-Q66 | 5.15 ± 2.8 | 29.71 ± 10.5 | 1.38 ± 0.24 |
| UpA | UpG | CpA | |
|---|---|---|---|
| RNase 6-R66 | 100 | ND | 100 |
| RNase 6-Q66 | 87.5 | ND | 92 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anguita, R.; Prats-Ejarque, G.; Moussaoui, M.; Becknell, B.; Boix, E. A Common Polymorphism in RNASE6 Impacts Its Antimicrobial Activity toward Uropathogenic Escherichia coli. Int. J. Mol. Sci. 2024, 25, 604. https://doi.org/10.3390/ijms25010604
Anguita R, Prats-Ejarque G, Moussaoui M, Becknell B, Boix E. A Common Polymorphism in RNASE6 Impacts Its Antimicrobial Activity toward Uropathogenic Escherichia coli. International Journal of Molecular Sciences. 2024; 25(1):604. https://doi.org/10.3390/ijms25010604
Chicago/Turabian StyleAnguita, Raul, Guillem Prats-Ejarque, Mohammed Moussaoui, Brian Becknell, and Ester Boix. 2024. "A Common Polymorphism in RNASE6 Impacts Its Antimicrobial Activity toward Uropathogenic Escherichia coli" International Journal of Molecular Sciences 25, no. 1: 604. https://doi.org/10.3390/ijms25010604
APA StyleAnguita, R., Prats-Ejarque, G., Moussaoui, M., Becknell, B., & Boix, E. (2024). A Common Polymorphism in RNASE6 Impacts Its Antimicrobial Activity toward Uropathogenic Escherichia coli. International Journal of Molecular Sciences, 25(1), 604. https://doi.org/10.3390/ijms25010604