
Parasite DNA and Markers of Decreased Immune Activation Associate Prospectively with Cardiac Functional Decline over 10 Years among Trypanosoma cruzi Seropositive Individuals in Brazil
 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Results
3. Discussion
4. Materials and Methods
4.1. Study Design
4.2. Blood Bank Screening Procedures
4.3. Parasite DNA Detection
4.4. Serum Biomarkers
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Frade, A.F.; Pissetti, C.W.; Ianni, B.M.; Saba, B.; Lin-Wang, H.T.; Nogueira, L.G.; Borges, A.d.M.; Buck, P.; Dias, F.; Baron, M.; et al. Genetic susceptibility to Chagas disease cardiomyopathy: Involvement of several genes of the innate immunity and chemokine-dependent migration pathways. BMC Infect. Dis. 2013, 13, 587. [Google Scholar] [CrossRef] [PubMed]
- Abel, L.C.; Rizzo, L.V.; Ianni, B.; Albuquerque, F.; Bacal, F.; Carrara, D.; Edimar, A.B.; Henrique, C.T.; Charles, M.; Jorge, K.; et al. Chronic Chagas’ disease cardiomyopathy patients display an increased IFN-gamma response to Trypanosoma cruzi infection. J. Autoimmun. 2001, 17, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Gomes, J.; Bahia-Oliveira, L.; Rocha, M.; Martins-Filho, O.; Gazzinelli, G.; Correa-Oliveira, R. Evidence that development of severe cardiomyopathy in human Chagas’ disease is due to a Th1-specific immune response. Infect. Immun. 2003, 71, 1185–1193. [Google Scholar] [CrossRef] [PubMed]
- Vitelli-Avelar, D.M.; Sathler-Avelar, R.; Teixeira-Carvalho, A.; Dias, J.C.P.; Gontijo, E.D.; Faria, A.M.; Elói-Santos, S.M.; Martins-Filho, O.A. Strategy to Assess the Overall Cytokine Profile of Circulating Leukocytes and its Association with Distinct Clinical Forms of Human Chagas Disease. Scand. J. Immunol. 2008, 68, 516–525. [Google Scholar] [CrossRef] [PubMed]
- Pinho, R.T.; Waghabi, M.C.; Cardillo, F.; Mengel, J.; Antas, P.R.Z. Scrutinizing the Biomarkers for the Neglected Chagas Disease: How Remarkable! Front. Immunol. 2016, 7, 306. [Google Scholar] [CrossRef] [PubMed]
- Sabino, E.C.; Ribeiro, A.L.; Lee, T.H.; Oliveira, C.L.; Carneiro-Proietti, A.B.; Antunes, A.P.; Menezes, M.M.; Ianni, B.M.; Salemi, V.M.; Nastari, L.; et al. Detection of Trypanosoma cruzi DNA in blood by PCR is associated with Chagas cardiomyopathy and disease severity. Eur. J. Heart Fail. 2015, 17, 416–423. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, M.T.; Schmidt, A.; da Silva, M.C.; Donadi, E.A.; da Silva, J.S.; Marin-Neto, J.A. Parasitic Load Correlates With Left Ventricular Dysfunction in Patients With Chronic Chagas Cardiomyopathy. Front. Cardiovasc. Med. 2021, 8, 741347. [Google Scholar] [CrossRef]
- Keating, S.M.; Deng, X.; Fernandes, F.; Cunha-Neto, E.; Ribeiro, A.L.; Adesina, B.; Beyer, A.; Contestable, P.; Custer, B.; Busch, M.; et al. Inflammatory and cardiac biomarkers are differentially expressed in clinical stages of Chagas disease. Int. J. Cardiol. 2015, 199, 451–459. [Google Scholar] [CrossRef]
- Kierszenbaum, F. Immunologic deficiency during experimental Chagas’ disease (Trypanosoma cruzi infection): Role of adherent, nonspecific esterase-positive splenic cells. J. Immunol. 1982, 129, 2202–2205. [Google Scholar] [CrossRef]
- Cetron, M.S.; Basilio, F.P.; Paes, J.N.; Sousa, A.Q.; Van Voorhis, W.C.; Kahn, S.J.; Wener, M.H.; Moraes, A.P. Humoral and cellular immune response of adults from northeastern Brazil with chronic Trypanosoma cruzi infection: Depressed cellular immune response to T. cruzi antigen among Chagas’ disease patients with symptomatic versus indeterminate infection. Am. J. Trop. Med. Hyg. 1993, 49, 370–382. [Google Scholar] [CrossRef]
- Rodrigues, V., Jr.; Agrelli, G.S.; Leon, S.C.; Silva Teixeira, D.N.; Tostes, S., Jr.; Rocha-Rodrigues, D.B. Fas/Fas-L expression, apoptosis and low proliferative response are associated with heart failure in patients with chronic Chagas’ disease. Microbes Infect. 2008, 10, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Antón, E.; Egui, A.; Thomas, M.C.; Carrilero, B.; Simón, M.; López-Ruz, M.; Segovia, M.; López, M.C. A proportion of CD4+ T cells from patients with chronic Chagas disease undergo a dysfunctional process, which is partially reversed by benznidazole treatment. PLoS Negl. Trop. Dis. 2021, 15, e0009059. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Antón, E.; Egui, A.; Thomas, M.C.; Simón, M.; Segovia, M.; López, M.C. Immunological exhaustion and functional profile of CD8(+) T lymphocytes as cellular biomarkers of therapeutic efficacy in chronic Chagas disease patients. Acta Trop. 2020, 202, 105242. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, L.R.P.; Ferreira, F.M.; Nakaya, H.I.; Deng, X.; Cândido, D.D.S.; De Oliveira, L.C.; Billaud, J.-N.; Lanteri, M.C.; Rigaud, V.O.-C.; Seielstad, M.; et al. Blood Gene Signatures of Chagas Cardiomyopathy with or without Ventricular Dysfunction. J. Infect. Dis. 2017, 215, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Torrico, F.; Heremans, H.; Rivera, M.T.; Van Marck, E.; Billiau, A.; Carlier, Y. Endogenous IFN-gamma is required for resistance to acute Trypanosoma cruzi infection in mice. J. Immunol. 1991, 146, 3626–3632. [Google Scholar] [CrossRef] [PubMed]
- Hölscher, C.; Köhler, G.; Müller, U.; Mossmann, H.; Schaub, G.N.A.; Brombacher, F. Defective nitric oxide effector functions lead to extreme susceptibility of Trypanosoma cruzi-infected mice deficient in gamma interferon receptor or inducible nitric oxide synthase. Infect. Immun. 1998, 66, 1208–1215. [Google Scholar] [CrossRef] [PubMed]
- Graefe, S.E.; Jacobs, T.; Gaworski, I.; Klauenberg, U.; Steeg, C.; Fleischer, B. Interleukin-12 but not interleukin-18 is required for immunity to Trypanosoma cruzi in mice. Microbes Infect. 2003, 5, 833–839. [Google Scholar] [CrossRef]
- Salvador, F.; Sánchez-Montalvá, A.; Martínez-Gallo, M.; Sulleiro, E.; Franco-Jarava, C.; Avilés, A.S.; Bosch-Nicolau, P.; Moure, Z.; Silgado, A.; Molina, I. Serum IL-10 Levels and Its Relationship with Parasitemia in Chronic Chagas Disease Patients. Am. J. Trop. Med. Hyg. 2020, 102, 159–163. [Google Scholar] [CrossRef]
- Noor, S.; Habashy, A.S.; Nance, J.P.; Clark, R.T.; Nemati, K.; Carson, M.J.; Wilson, E.H. CCR7-dependent immunity during acute Toxoplasma gondii infection. Infect. Immun. 2010, 78, 2257–2263. [Google Scholar] [CrossRef]
- Ploix, C.C.; Noor, S.; Crane, J.; Masek, K.; Carter, W.; Lo, D.D.; Wilson, E.H.; Carson, M.J. CNS-derived CCL21 is both sufficient to drive homeostatic CD4+ T cell proliferation and necessary for efficient CD4+ T cell migration into the CNS parenchyma following Toxoplasma gondii infection. Brain Behav. Immun. 2011, 25, 883–896. [Google Scholar] [CrossRef]
- Nogueira, L.G.; Santos, R.H.B.; Ianni, B.M.; Fiorelli, A.I.; Mairena, E.C.; Benvenuti, L.A.; Frade, A.; Donadi, E.; Dias, F.; Saba, B.; et al. Myocardial chemokine expression and intensity of myocarditis in Chagas cardiomyopathy are controlled by polymorphisms in CXCL9 and CXCL10. PLoS Negl. Trop. Dis. 2012, 6, e1867. [Google Scholar] [CrossRef] [PubMed]
- Barinov, A.; Luo, L.; Gasse, P.; Meas-Yedid, V.; Donnadieu, E.; Arenzana-Seisdedos, F.; Vieira, P. Essential role of immobilized chemokine CXCL12 in the regulation of the humoral immune response. Proc. Natl. Acad. Sci. USA 2017, 114, 2319–2324. [Google Scholar] [CrossRef] [PubMed]
- Garnica, M.R.; De Moraes, L.V.; Rizzo, L.V.; De Andrade, H.F., Jr. Supplementation of CXCL12 (CXCL12) induces homing of CD11c+ dendritic cells to the spleen and enhances control of Plasmodium berghei malaria in BALB/c mice. Immunology 2005, 115, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Berbert, L.R.; González, F.B.; Villar, S.R.; Vigliano, C.; Lioi, S.; Beloscar, J.; Bottasso, O.A.; Silva-Barbosa, S.D.; Savino, W.; Pérez, A.R.; et al. Enhanced Migratory Capacity of T Lymphocytes in Severe Chagasic Patients Is Correlated With VLA-4 and TNF-α Expression. Front. Cell Infect. Microbiol. 2021, 11, 713150. [Google Scholar] [CrossRef] [PubMed]
- Santamaría, M.H.; Ríos, L.D.; Corral, R.S. Chronic Trypanosoma cruzi infection activates the TWEAK/Fn14 axis in cardiac myocytes and fibroblasts driving structural and functional changes that affect the heart. Exp. Parasitol. 2023, 248, 108491. [Google Scholar] [CrossRef] [PubMed]
- Castro-Ochoa, K.F.; Guerrero-Fonseca, I.M.; Schnoor, M. Hematopoietic cell-specific lyn substrate (HCLS1 or HS1): A versatile actin-binding protein in leukocytes. J. Leukoc. Biol. 2019, 105, 881–890. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Cao, J.; Fan, Y.; Li, C.; Hu, X. Comprehensive analysis of miRNA-mRNA regulatory network and potential drugs in chronic chagasic cardiomyopathy across human and mouse. BMC Med. Genomics 2021, 14, 283. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Clinton, J.; Yao, C.; Chang, S.H. Interleukin-17D Promotes Pathogenicity during Infection by Suppressing CD8 T Cell Activity. Front. Immunol. 2019, 10, 1172. [Google Scholar] [CrossRef]
- Nanatsue, K.; Ninomiya, T.; Tsuchiya, M.; Tahara-Hanaoka, S.; Shibuya, A.; Masuko, H.; Sakamoto, T.; Hizawa, N.; Arinami, T.; Noguchi, E. Influence of MILR1 promoter polymorphism on expression levels and the phenotype of atopy. J. Human. Genet. 2014, 59, 480–483. [Google Scholar] [CrossRef]
- Nagasawa, T. CXC chemokine ligand 12 (CXCL12) and its receptor CXCR4. J. Mol. Med. 2014, 92, 433–439. [Google Scholar] [CrossRef]
- Sabino, E.C.; Ribeiro, A.L.; Salemi, V.M.; Oliveira, C.D.L.; Antunes, A.P.; Menezes, M.M.; Ianni, B.M.; Nastari, L.; Fernandes, F.; Patavino, G.M.; et al. Ten-year incidence of Chagas cardiomyopathy among asymptomatic Trypanosoma cruzi-seropositive former blood donors. Circulation 2013, 127, 1105–1115. [Google Scholar] [CrossRef] [PubMed]
- Johansen, F.E.; Braathen, R.; Brandtzaeg, P. Role of J chain in secretory immunoglobulin formation. Scand. J. Immunol. 2000, 52, 240–248. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Sabino, E.C.; Cunha-Neto, E.; Ribeiro, A.L.; Ianni, B.; Mady, C.; Busch, M.P.; Seielstad, M.; International Component; The REDSII Chagas Study Group from the NHLBI Retrovirus Epidemiology Donor Study-II (REDS-II). Genome Wide Association Study (GWAS) of Chagas Cardiomyopathy in Trypanosoma cruzi Seropositive Subjects. PLoS ONE 2013, 8, e79629. [Google Scholar] [CrossRef] [PubMed]
- Salles, N.; Sabino, E.; Cliquet, M.; Eluf-Neto, J.; Mayer, A.; Almeida-Neto, C.; Mendonça, M.C.; Dorliach-Llacer, P.; Chamone, D.F.; MSc, A.S.-A.; et al. Risk of exposure to Chagas’ disease among seroreactive Brazilian blood donors. Transfusion 1996, 36, 969–973. [Google Scholar] [CrossRef]
- Saéz-Alquézar, A.; Otani, M.M.; Sabino, E.C.; Ribeiro-dos-Santos, G.; Salles, N.; Chamone, D.F. Evaluation of the performance of Brazilian blood banks in testing for Chagas’ disease. Vox Sang. 1998, 74, 228–231. [Google Scholar] [CrossRef]
- Sanders-van Wijk, S.; Tromp, J.; Beussink-Nelson, L.; Hage, C.; Svedlund, S.; Saraste, A.; Swat, S.A.; Sanchez, C.; Njoroge, J.; Tan, R.-S.; et al. Proteomic Evaluation of the Comorbidity-Inflammation Paradigm in Heart Failure With Preserved Ejection Fraction: Results From the PROMIS-HFpEF Study. Circulation 2020, 142, 2029–2044. [Google Scholar] [CrossRef]
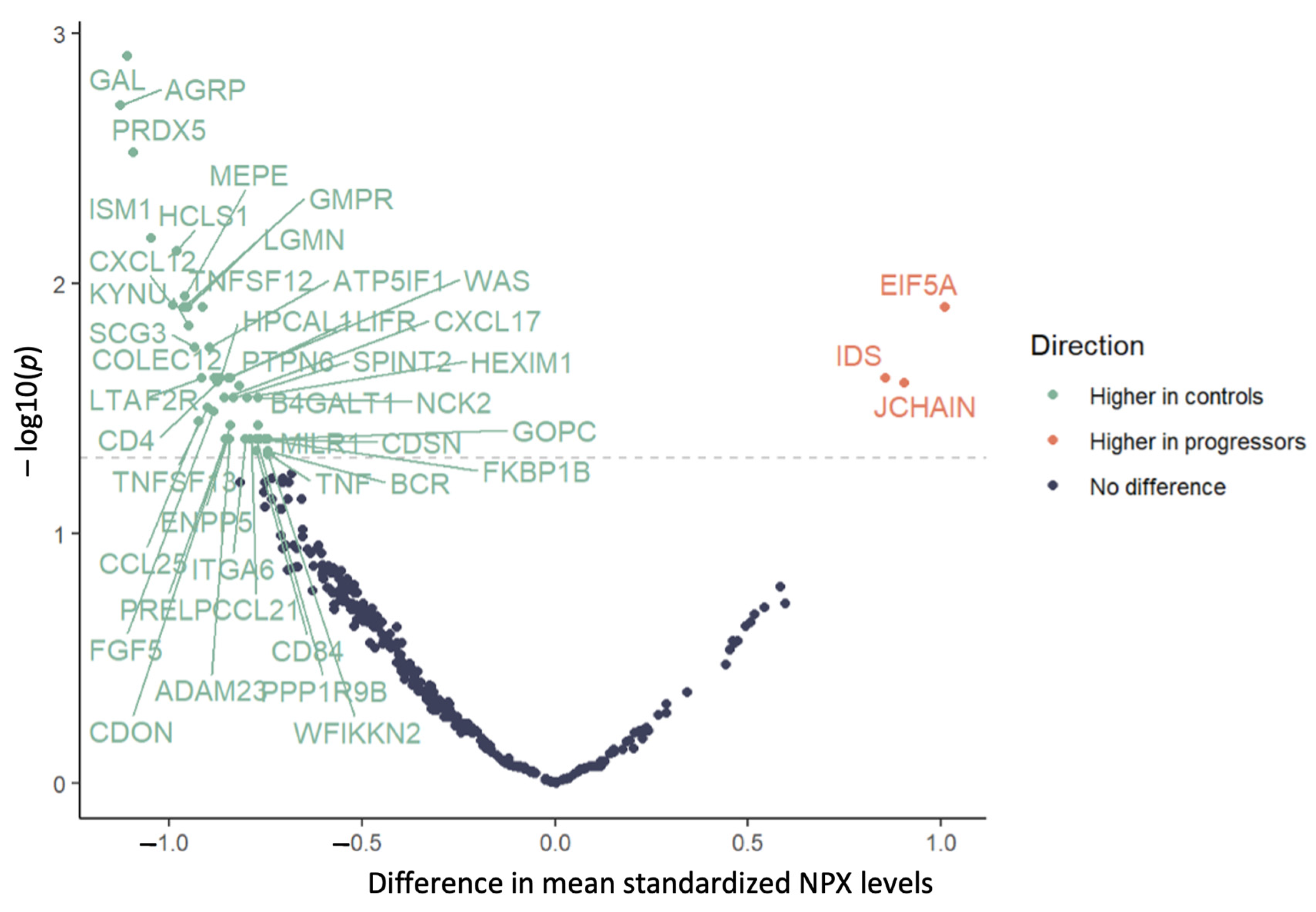

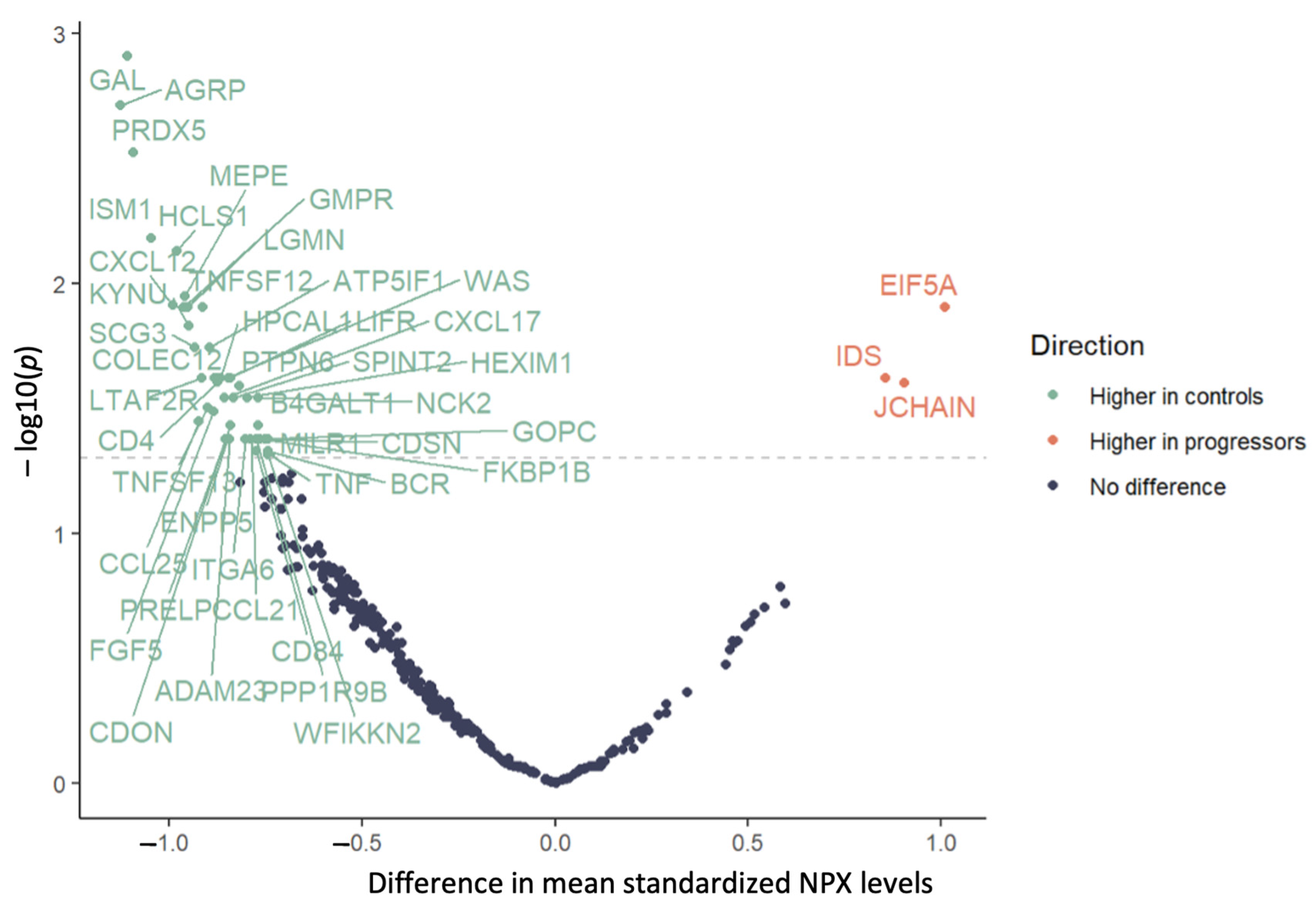{kind=link}
| Characteristics | CONTROLS (n = 31) | PROGRESSORS (n = 21) | p Value |
|---|---|---|---|
| Sex, n (%) | 1 | ||
| Male | 18 (58.1%) | 13 (61.9%) | |
| Female | 13 (41.9%) | 8 (38.1%) | |
| Age at T0 1, Mean (SD) | 42.8 (7.65) | 48.2 (9.75) | 0.04 |
| Age at T1 2, Mean (SD) | 51.6 (7.60) | 56.3 (9.75) | 0.07 |
| Hypertension at Baseline | <0.01 | ||
| No | 31 (100%) | 11 (52.4%) | |
| Yes | 0 (0%) | 10 (47.6%) | |
| Hypertension at Follow-up | 0.03 | ||
| No | 30 (96.8%) | 16 (76.2%) | |
| Yes | 1 (3.2%) | 5 (23.8%) | |
| Diabetes at Baseline | 0.68 | ||
| No | 28 (90.3%) | 18 (85.7%) | |
| Yes | 3 (9.7%) | 3 (14.3%) | |
| Diabetes at Follow-up | 1 | ||
| No | 27 (87.1%) | 18 (85.7%) | |
| Yes | 4 (6.5%) | 3 (14.3%) | |
| EF 3 at Baseline, Mean (SD) | 62.5 (2.59) | 53.3 (8.43) | <0.01 |
| EF 3 at Follow-up, Mean (SD) | 67.6 (4.90) | 39.2 (12.8) | <0.01 |
| T. cruzi DNA 4 at T0, Mean (SD) | 0.605 (1.63) | 18.3 (43.6) | 0.02 |
| Primary Analysis n = 52 | Restricted Analysis 1 n = 43 | |||||
|---|---|---|---|---|---|---|
| Protein | Change | Mean Difference | p Value 2 | Change | Mean Difference | p Value 2 |
| ADAM23 | Decrease | −0.842 | 0.042 | – | −0.758 | 0.084 |
| AGRP | Decrease | −1.125 | 0.002 | Decrease | −1.073 | 0.012 |
| AMN | – | −0.71 | 0.102 | Decrease | −1.054 | 0.040 |
| ATP5IF1 | Decrease | −0.894 | 0.018 | Decrease | −1.073 | 0.001 |
| B4GALT1 | Decrease | −0.768 | 0.037 | Decrease | −0.923 | 0.012 |
| BCR | Decrease | −0.743 | 0.047 | Decrease | −0.973 | 0.001 |
| CCL21 | Decrease | −0.788 | 0.042 | Decrease | −1.017 | 0.010 |
| CCL25 | Decrease | −0.924 | 0.036 | – | −0.915 | 0.097 |
| CD4 | Decrease | −0.874 | 0.025 | – | −0.815 | 0.073 |
| CD79B | – | −0.705 | 0.060 | Decrease | −0.702 | 0.028 |
| CD84 | Decrease | −0.773 | 0.047 | – | −0.863 | 0.052 |
| CDON | Decrease | −0.84 | 0.037 | – | −0.845 | 0.067 |
| CDSN | Decrease | −0.743 | 0.042 | – | −0.659 | 0.111 |
| COLEC12 | Decrease | −0.874 | 0.024 | Decrease | −0.908 | 0.026 |
| CXCL12 | Decrease | −0.99 | 0.012 | Decrease | −1.094 | 0.026 |
| CXCL17 | Decrease | −0.856 | 0.029 | – | −0.667 | 0.149 |
| EIF5A | Increase | 1.01 | 0.012 | – | 0.798 | 0.145 |
| ENPP5 | Decrease | −0.848 | 0.042 | – | −0.847 | 0.077 |
| F2R | Decrease | −0.883 | 0.024 | Decrease | −1.022 | 0.006 |
| FGF5 | Decrease | −0.884 | 0.032 | – | −0.916 | 0.077 |
| FKBP1B | Decrease | −0.752 | 0.042 | Decrease | −0.897 | 0.013 |
| FOXO1 | – | −0.603 | 0.134 | Decrease | −0.818 | 0.020 |
| GAL | Decrease | −1.108 | 0.001 | Decrease | −1.17 | 0.001 |
| GMPR | Decrease | −0.962 | 0.012 | Decrease | −0.993 | 0.012 |
| GOPC | Decrease | −0.765 | 0.042 | Decrease | −0.905 | 0.006 |
| HCLS1 | Decrease | −0.979 | 0.007 | Decrease | −1.017 | 0.006 |
| HEXIM1 | Decrease | −0.798 | 0.029 | Decrease | −0.9 | 0.012 |
| HPCAL1 | Decrease | −0.867 | 0.024 | Decrease | −0.907 | 0.012 |
| IDS | Increase | 0.856 | 0.024 | – | 0.649 | 0.149 |
| ISM1 | Decrease | −1.045 | 0.007 | Decrease | −1.08 | 0.006 |
| ITGA6 | Decrease | −0.802 | 0.042 | – | −0.816 | 0.069 |
| JCHAIN | Increase | 0.904 | 0.025 | Increase | 1.124 | 0.014 |
| KYNU | Decrease | −0.949 | 0.015 | Decrease | −1.032 | 0.012 |
| LGMN | Decrease | −0.952 | 0.012 | Decrease | −0.977 | 0.026 |
| LIFR | Decrease | −0.841 | 0.024 | Decrease | −0.938 | 0.024 |
| LTA | Decrease | −0.915 | 0.024 | – | −0.83 | 0.054 |
| MEPE | Decrease | −0.959 | 0.011 | Decrease | −0.964 | 0.010 |
| MGMT | – | −0.633 | 0.121 | Decrease | −0.797 | 0.043 |
| MILR1 | Decrease | −0.744 | 0.042 | Decrease | −0.807 | 0.026 |
| NBN | – | −0.491 | 0.206 | Decrease | −0.672 | 0.042 |
| NCK2 | Decrease | −0.77 | 0.029 | Decrease | −0.912 | 0.006 |
| PPP1R9B | Decrease | −0.773 | 0.042 | Decrease | −0.868 | 0.015 |
| PRDX5 | Decrease | −1.093 | 0.003 | Decrease | −1.055 | 0.012 |
| PRELP | Decrease | −0.854 | 0.042 | – | −0.708 | 0.077 |
| PTPN6 | Decrease | −0.818 | 0.026 | Decrease | −0.971 | 0.011 |
| SCG3 | Decrease | −0.933 | 0.018 | Decrease | −0.974 | 0.015 |
| SCRN1 | – | −0.734 | 0.060 | Decrease | −0.841 | 0.049 |
| SMOC2 | – | −0.411 | 0.282 | Decrease | −0.762 | 0.040 |
| SPINT2 | Decrease | −0.832 | 0.029 | Decrease | −0.93 | 0.012 |
| TNF | Decrease | −0.738 | 0.049 | – | −0.635 | 0.102 |
| TNFRSF4 | – | −0.349 | 0.395 | Decrease | −0.736 | 0.040 |
| TNFSF12 | Decrease | −0.914 | 0.012 | Decrease | −0.914 | 0.026 |
| TNFSF13 | Decrease | −0.901 | 0.031 | – | −0.837 | 0.077 |
| WAS | Decrease | −0.846 | 0.024 | Decrease | −0.897 | 0.010 |
| WFIKKN2 | Decrease | −0.744 | 0.049 | Decrease | −0.983 | 0.011 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sunderraj, A.; Cunha, L.M.; Avila, M.; Alexandria, S.; Ferreira, A.M.; de Oliveira-da Silva, L.C.; Ribeiro, A.L.P.; Nunes, M.d.C.P.; Sabino, E.C.; Landay, A.; et al. Parasite DNA and Markers of Decreased Immune Activation Associate Prospectively with Cardiac Functional Decline over 10 Years among Trypanosoma cruzi Seropositive Individuals in Brazil. Int. J. Mol. Sci. 2024, 25, 44. https://doi.org/10.3390/ijms25010044
Sunderraj A, Cunha LM, Avila M, Alexandria S, Ferreira AM, de Oliveira-da Silva LC, Ribeiro ALP, Nunes MdCP, Sabino EC, Landay A, et al. Parasite DNA and Markers of Decreased Immune Activation Associate Prospectively with Cardiac Functional Decline over 10 Years among Trypanosoma cruzi Seropositive Individuals in Brazil. International Journal of Molecular Sciences. 2024; 25(1):44. https://doi.org/10.3390/ijms25010044
Chicago/Turabian StyleSunderraj, Ashwin, Luisa Marin Cunha, Matheus Avila, Shaina Alexandria, Ariela Mota Ferreira, Léa Campos de Oliveira-da Silva, Antonio L. P. Ribeiro, Maria do Carmo Pereira Nunes, Ester C. Sabino, Alan Landay, and et al. 2024. "Parasite DNA and Markers of Decreased Immune Activation Associate Prospectively with Cardiac Functional Decline over 10 Years among Trypanosoma cruzi Seropositive Individuals in Brazil" International Journal of Molecular Sciences 25, no. 1: 44. https://doi.org/10.3390/ijms25010044
APA StyleSunderraj, A., Cunha, L. M., Avila, M., Alexandria, S., Ferreira, A. M., de Oliveira-da Silva, L. C., Ribeiro, A. L. P., Nunes, M. d. C. P., Sabino, E. C., Landay, A., Kalil, J., Chevillard, C., Cunha-Neto, E., & Feinstein, M. J. (2024). Parasite DNA and Markers of Decreased Immune Activation Associate Prospectively with Cardiac Functional Decline over 10 Years among Trypanosoma cruzi Seropositive Individuals in Brazil. International Journal of Molecular Sciences, 25(1), 44. https://doi.org/10.3390/ijms25010044