

Enhancement of Sphingomyelinase-Induced Endothelial Nitric Oxide Synthase-Mediated Vasorelaxation in a Murine Model of Type 2 Diabetes

 , , , and
, , , and 
Abstract
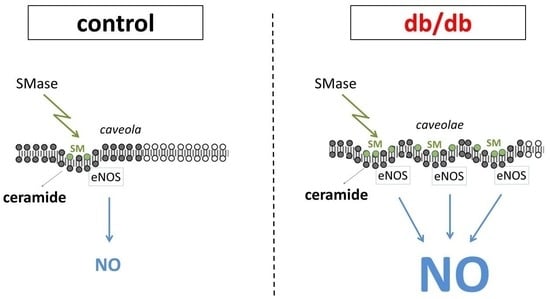
1. Introduction
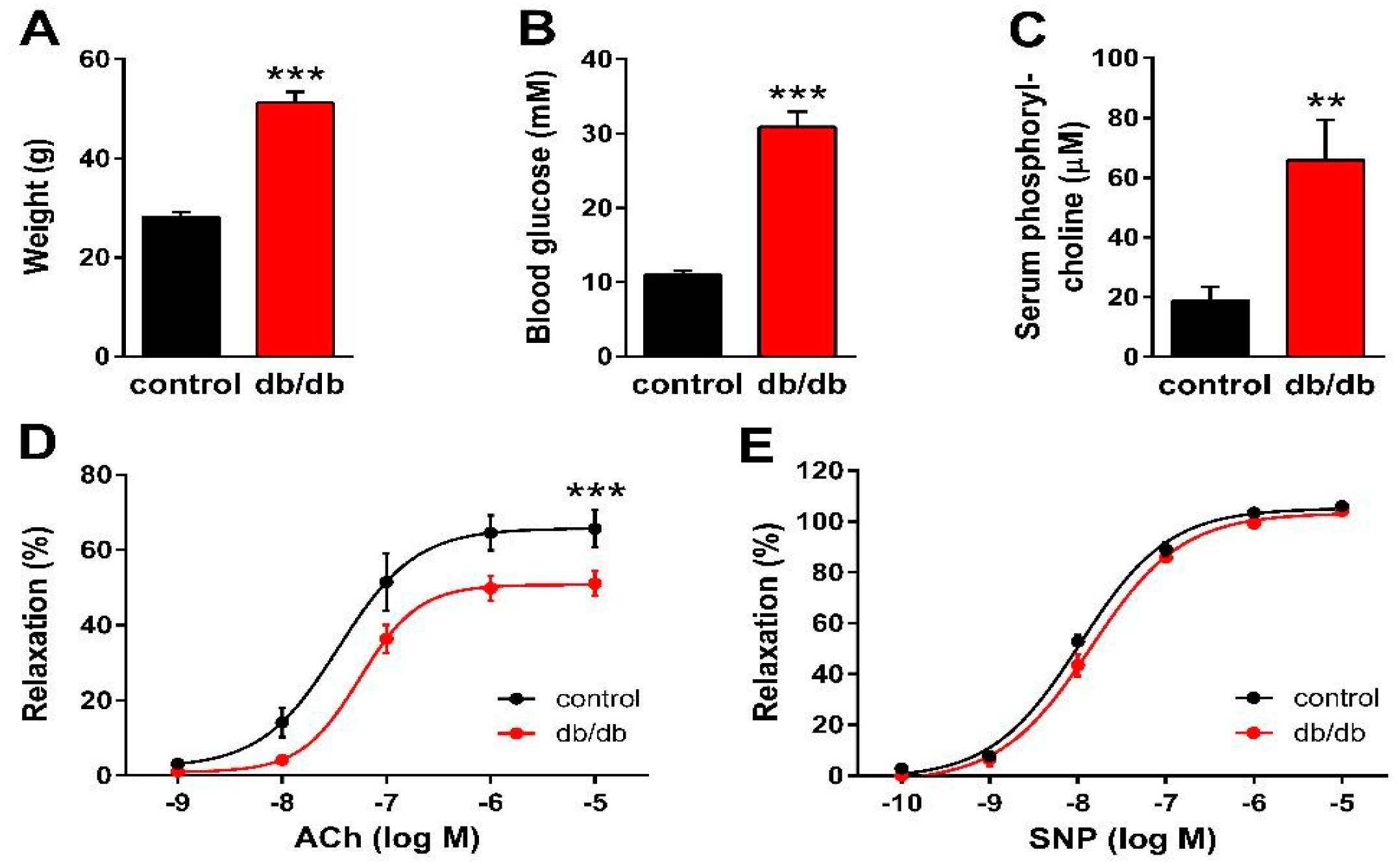
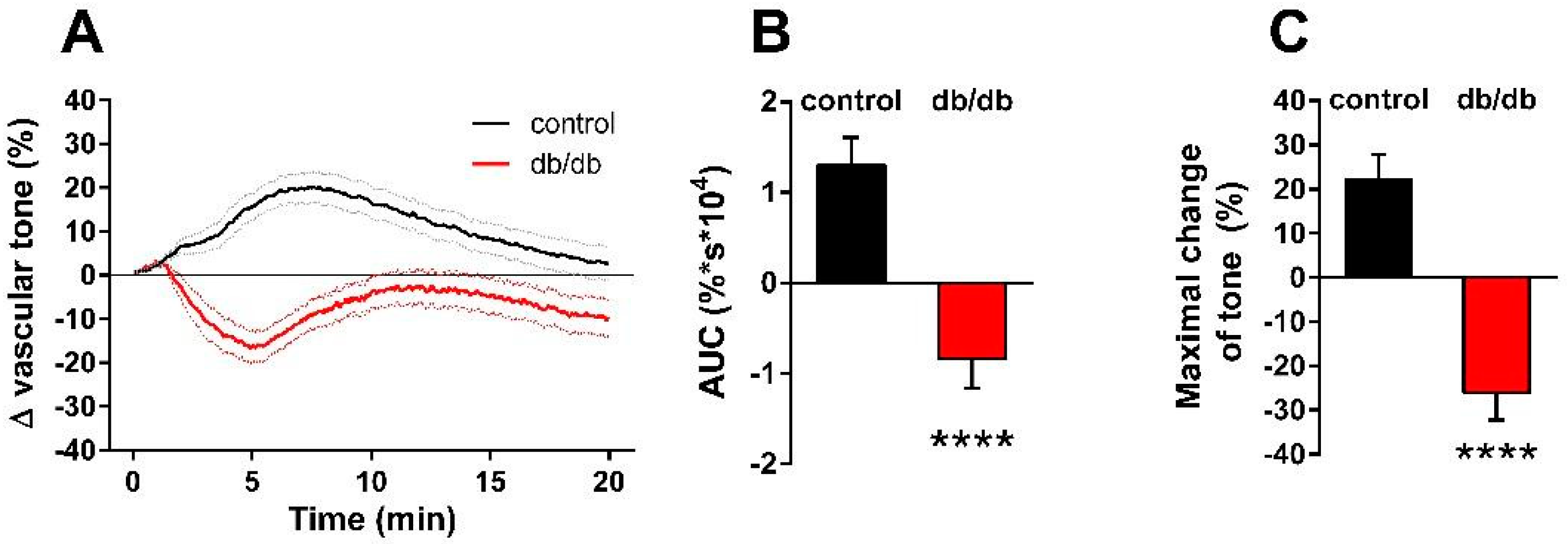
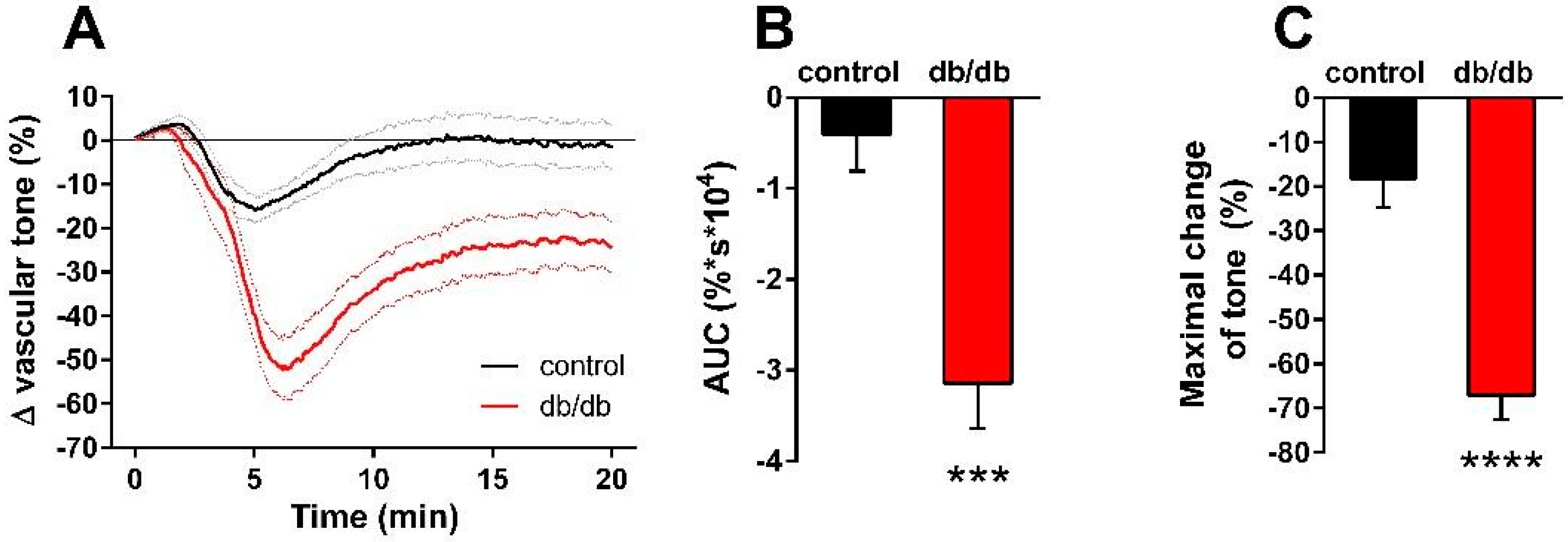
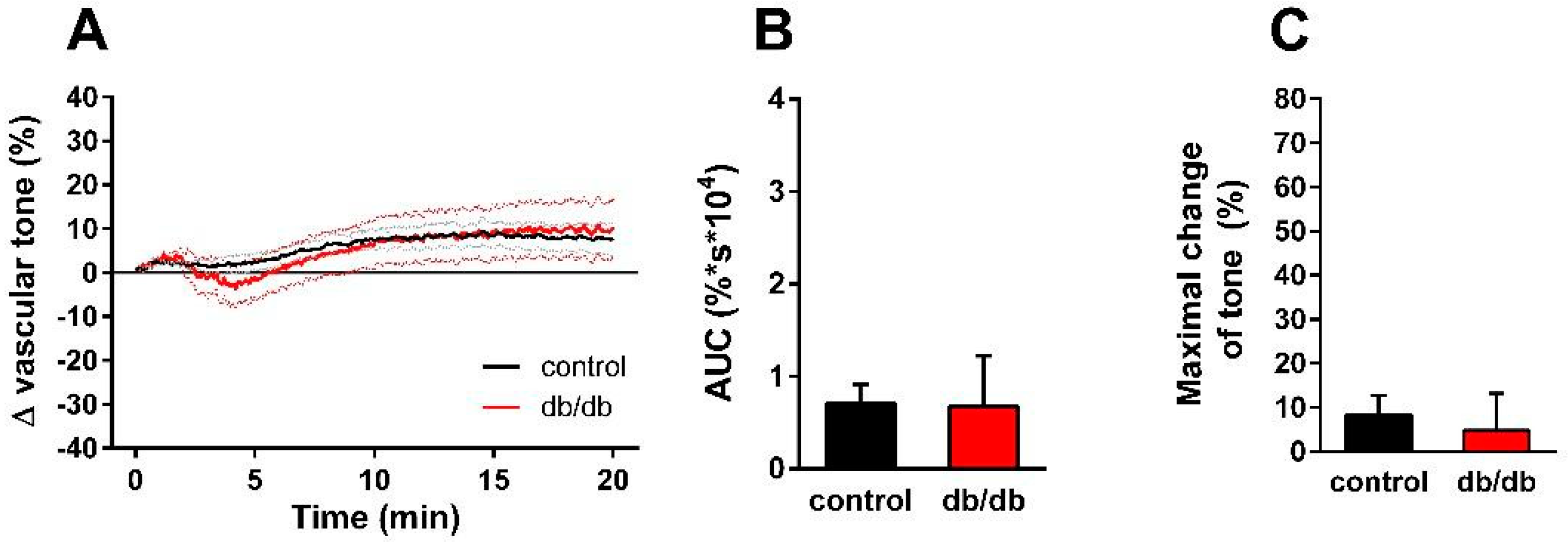
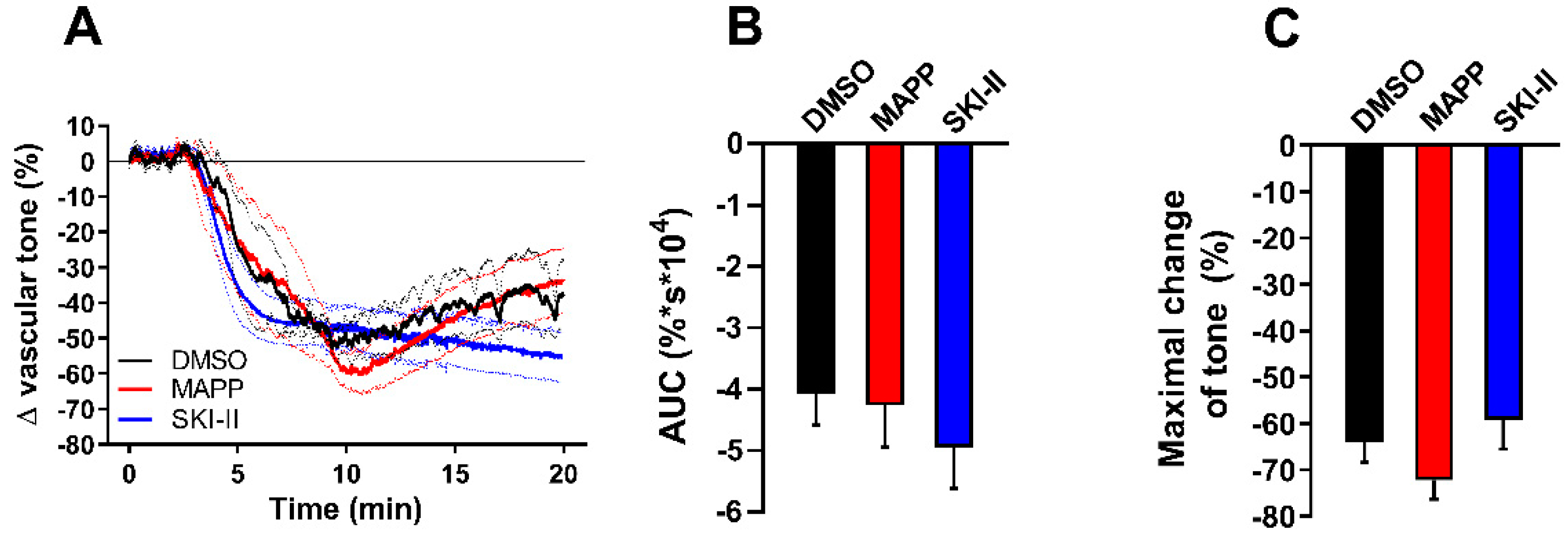
2. Results
3. Discussion
4. Materials and Methods
4.1. Animals and General Procedures
4.2. Myography
4.3. Data Analysis
4.4. Reagents
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Peters, S.L.; Alewijnse, A.E. Sphingosine-1-phosphate signaling in the cardiovascular system. Curr. Opin. Pharmacol. 2007, 7, 186–192. [Google Scholar] [CrossRef] [PubMed]
- Igarashi, J.; Michel, T. Sphingosine-1-phosphate and modulation of vascular tone. Cardiovasc. Res. 2009, 82, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Kerage, D.; Brindley, D.N.; Hemmings, D.G. Review: Novel insights into the regulation of vascular tone by sphingosine 1-phosphate. Placenta 2014, 35, S86–S92. [Google Scholar] [CrossRef] [PubMed]
- Proia, R.L.; Hla, T. Emerging biology of sphingosine-1-phosphate: Its role in pathogenesis and therapy. J. Clin. Investig. 2015, 125, 1379–1387. [Google Scholar] [CrossRef]
- Hemmings, D.G. Signal transduction underlying the vascular effects of sphingosine 1-phosphate and sphingosylphosphorylcholine. Naunyn. Schmiedebergs Arch. Pharmacol. 2006, 373, 18–29. [Google Scholar] [CrossRef] [PubMed]
- De Palma, C.; Meacci, E.; Perrotta, C.; Bruni, P.; Clementi, E. Endothelial nitric oxide synthase activation by tumor necrosis factor alpha through neutral sphingomyelinase 2, sphingosine kinase 1, and sphingosine 1 phosphate receptors: A novel pathway relevant to the pathophysiology of endothelium. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Fyrst, H.; Saba, J.D. An update on sphingosine-1-phosphate and other sphingolipid mediators. Nat. Chem. Biol. 2010, 6, 489–497. [Google Scholar] [CrossRef] [PubMed]
- Meyer zu Heringdorf, D.; Jakobs, K.H. Lysophospholipid receptors: Signalling, pharmacology and regulation by lysophospholipid metabolism. Biochim. Biophys. Acta 2007, 1768, 923–940. [Google Scholar] [CrossRef]
- Strub, G.M.; Maceyka, M.; Hait, N.C.; Milstien, S.; Spiegel, S. Extracellular and intracellular actions of sphingosine-1-phosphate. Adv. Exp. Med. Biol. 2010, 688, 141–155. [Google Scholar]
- Hla, T.; Dannenberg, A.J. Sphingolipid signaling in metabolic disorders. Cell Metab. 2012, 16, 420–434. [Google Scholar] [CrossRef]
- Ernst, A.M.; Brugger, B. Sphingolipids as modulators of membrane proteins. Biochim. Biophys. Acta 2014, 1841, 665–670. [Google Scholar] [CrossRef] [PubMed]
- Adada, M.; Luberto, C.; Canals, D. Inhibitors of the sphingomyelin cycle: Sphingomyelin synthases and sphingomyelinases. Chem. Phys. Lipids 2016, 197, 45–59. [Google Scholar] [CrossRef]
- Pavoine, C.; Pecker, F. Sphingomyelinases: Their regulation and roles in cardiovascular pathophysiology. Cardiovasc. Res. 2009, 82, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Murohara, T.; Kugiyama, K.; Ohgushi, M.; Sugiyama, S.; Ohta, Y.; Yasue, H. Effects of sphingomyelinase and sphingosine on arterial vasomotor regulation. J. Lipid Res. 1996, 37, 1601–1608. [Google Scholar] [CrossRef] [PubMed]
- Johns, D.G.; Jin, J.S.; Wilde, D.W.; Webb, R.C. Ceramide-induced vasorelaxation: An inhibitory action on protein kinase C. Gen. Pharmacol. 1999, 33, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Johns, D.G.; Osborn, H.; Webb, R.C. Ceramide: A novel cell signaling mechanism for vasodilation. Biochem. Biophys. Res. Commun. 1997, 237, 95–97. [Google Scholar] [CrossRef]
- Zheng, T.; Li, W.; Wang, J.; Altura, B.T.; Altura, B.M. Effects of neutral sphingomyelinase on phenylephrine-induced vasoconstriction and Ca(2+) mobilization in rat aortic smooth muscle. Eur. J. Pharmacol. 2000, 391, 127–135. [Google Scholar] [CrossRef]
- Zheng, T.; Li, W.; Wang, J.; Altura, B.T.; Altura, B.M. Sphingomyelinase and ceramide analogs induce contraction and rises in [Ca(2+)](i) in canine cerebral vascular muscle. Am. J. Physiol. Heart Circ. Physiol. 2000, 278, H1421–H1428. [Google Scholar] [CrossRef] [PubMed]
- Altura, B.M.; Gebrewold, A.; Zheng, T.; Altura, B.T. Sphingomyelinase and ceramide analogs induce vasoconstriction and leukocyte-endothelial interactions in cerebral venules in the intact rat brain: Insight into mechanisms and possible relation to brain injury and stroke. Brain Res. Bull. 2002, 58, 271–278. [Google Scholar] [CrossRef]
- Jang, G.J.; Ahn, D.S.; Cho, Y.E.; Morgan, K.G.; Lee, Y.H. C2-ceramide induces vasodilation in phenylephrine-induced pre-contracted rat thoracic aorta: Role of RhoA/Rho-kinase and intracellular Ca2+ concentration. Naunyn. Schmiedebergs Arch. Pharmacol. 2005, 372, 242–250. [Google Scholar] [CrossRef] [PubMed]
- Mogami, K.; Kishi, H.; Kobayashi, S. Sphingomyelinase causes endothelium-dependent vasorelaxation through endothelial nitric oxide production without cytosolic Ca2+ elevation. FEBS Lett. 2005, 579, 393–397. [Google Scholar] [CrossRef] [PubMed]
- Cogolludo, A.; Moreno, L.; Frazziano, G.; Moral-Sanz, J.; Menendez, C.; Castaneda, J.; Gonzalez, C.; Villamor, E.; Perez-Vizcaino, F. Activation of neutral sphingomyelinase is involved in acute hypoxic pulmonary vasoconstriction. Cardiovasc. Res. 2009, 82, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Spijkers, L.J.; van den Akker, R.F.; Janssen, B.J.; Debets, J.J.; De Mey, J.G.; Stroes, E.S.; van den Born, B.J.; Wijesinghe, D.S.; Chalfant, C.E.; MacAleese, L.; et al. Hypertension is associated with marked alterations in sphingolipid biology: A potential role for ceramide. PLoS ONE 2011, 6, e21817. [Google Scholar] [CrossRef] [PubMed]
- Spijkers, L.J.; Janssen, B.J.; Nelissen, J.; Meens, M.J.; Wijesinghe, D.; Chalfant, C.E.; De Mey, J.G.; Alewijnse, A.E.; Peters, S.L. Antihypertensive treatment differentially affects vascular sphingolipid biology in spontaneously hypertensive rats. PLoS ONE 2011, 6, e29222. [Google Scholar] [CrossRef] [PubMed]
- van den Elsen, L.W.; Spijkers, L.J.; van den Akker, R.F.; van Winssen, A.M.; Balvers, M.; Wijesinghe, D.S.; Chalfant, C.E.; Garssen, J.; Willemsen, L.E.; Alewijnse, A.E.; et al. Dietary fish oil improves endothelial function and lowers blood pressure via suppression of sphingolipid-mediated contractions in spontaneously hypertensive rats. J. Hypertens. 2014, 32, 1050–1058; discussion 1058. [Google Scholar] [CrossRef] [PubMed]
- Shamseddine, A.A.; Airola, M.V.; Hannun, Y.A. Roles and regulation of neutral sphingomyelinase-2 in cellular and pathological processes. Adv. Biol. Regul. 2015, 57, 24–41. [Google Scholar] [CrossRef]
- Russo, S.B.; Ross, J.S.; Cowart, L.A. Sphingolipids in obesity, type 2 diabetes, and metabolic disease. Handb. Exp. Pharmacol. 2013, 216, 373–401. [Google Scholar] [CrossRef]
- Cowart, L.A. Sphingolipids: Players in the pathology of metabolic disease. Trends Endocrinol. Metab. 2009, 20, 34–42. [Google Scholar] [CrossRef]
- Hannun, Y.A.; Obeid, L.M. The ceramide-centric universe of lipid-mediated cell regulation: Stress encounters of the lipid kind. J. Biol. Chem. 2002, 277, 25847–25850. [Google Scholar] [CrossRef] [PubMed]
- Sawai, H.; Hannun, Y.A. Ceramide and sphingomyelinases in the regulation of stress responses. Chem. Phys. Lipids 1999, 102, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Hannun, Y.A.; Luberto, C. Ceramide in the eukaryotic stress response. Trends Cell Biol. 2000, 10, 73–80. [Google Scholar] [CrossRef] [PubMed]
- van Meer, G.; Holthuis, J.C. Sphingolipid transport in eukaryotic cells. Biochim. Biophys. Acta 2000, 1486, 145–170. [Google Scholar] [CrossRef] [PubMed]
- Holland, W.L.; Summers, S.A. Sphingolipids, insulin resistance, and metabolic disease: New insights from in vivo manipulation of sphingolipid metabolism. Endocr. Rev. 2008, 29, 381–402. [Google Scholar] [CrossRef] [PubMed]
- Unger, R.H.; Orci, L. Lipoapoptosis: Its mechanism and its diseases. Biochim. Biophys. Acta 2002, 1585, 202–212. [Google Scholar] [CrossRef]
- Boden, G. Pathogenesis of type 2 diabetes. Insulin resistance. Endocrinol. Metab. Clin. N. Am. 2001, 30, 801–815. [Google Scholar] [CrossRef] [PubMed]
- Samad, F. Contribution of sphingolipids to the pathogenesis of obesity. Future Lipidol. 2007, 2, 625–639. [Google Scholar] [CrossRef]
- Symons, J.D.; Abel, E.D. Lipotoxicity contributes to endothelial dysfunction: A focus on the contribution from ceramide. Rev. Endocr. Metab. Disord. 2013, 14, 59–68. [Google Scholar] [CrossRef]
- Aasum, E.; Hafstad, A.D.; Severson, D.L.; Larsen, T.S. Age-dependent changes in metabolism, contractile function, and ischemic sensitivity in hearts from db/db mice. Diabetes 2003, 52, 434–441. [Google Scholar] [CrossRef] [PubMed]
- Coleman, D.L. Obese and diabetes: Two mutant genes causing diabetes-obesity syndromes in mice. Diabetologia 1978, 14, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Do, O.H.; Low, J.T.; Gaisano, H.Y.; Thorn, P. The secretory deficit in islets from db/db mice is mainly due to a loss of responding beta cells. Diabetologia 2014, 57, 1400–1409. [Google Scholar] [CrossRef] [PubMed]
- Sallam, N.A.; Laher, I. Redox Signaling and Regional Heterogeneity of Endothelial Dysfunction in db/db Mice. Int. J. Mol. Sci. 2020, 21, 6147. [Google Scholar] [CrossRef]
- Ramadan, F.M.; Upchurch, G.R., Jr.; Keagy, B.A.; Johnson, G., Jr. Endothelial cell thromboxane production and its inhibition by a calcium-channel blocker. Ann. Thorac. Surg. 1990, 49, 916–919. [Google Scholar] [CrossRef]
- Epand, R.M.; So, V.; Jennings, W.; Khadka, B.; Gupta, R.S.; Lemaire, M. Diacylglycerol Kinase-epsilon: Properties and Biological Roles. Front. Cell. Dev. Biol. 2016, 4, 112. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, P.; Vora, M.; Gentile, L.B.; Stahelin, R.V.; Chalfant, C.E. Anionic lipids activate group IVA cytosolic phospholipase A2 via distinct and separate mechanisms. J. Lipid Res. 2007, 48, 2701–2708. [Google Scholar] [CrossRef] [PubMed]
- Pettus, B.J.; Bielawska, A.; Spiegel, S.; Roddy, P.; Hannun, Y.A.; Chalfant, C.E. Ceramide kinase mediates cytokine- and calcium ionophore-induced arachidonic acid release. J. Biol. Chem. 2003, 278, 38206–38213. [Google Scholar] [CrossRef] [PubMed]
- Mitsutake, S.; Date, T.; Yokota, H.; Sugiura, M.; Kohama, T.; Igarashi, Y. Ceramide kinase deficiency improves diet-induced obesity and insulin resistance. FEBS Lett. 2012, 586, 1300–1305. [Google Scholar] [CrossRef] [PubMed]
- Machida, T.; Matamura, R.; Iizuka, K.; Hirafuji, M. Cellular function and signaling pathways of vascular smooth muscle cells modulated by sphingosine 1-phosphate. J. Pharmacol. Sci. 2016, 132, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Goto, K.; Kitazono, T. Endothelium-dependent hyperpolarization (EDH) in diet-induced obesity. Endocr. Metab. Sci. 2020, 1, 100062. [Google Scholar] [CrossRef]
- Pilarczyk, M.; Mateuszuk, L.; Rygula, A.; Kepczynski, M.; Chlopicki, S.; Baranska, M.; Kaczor, A. Endothelium in spots—high-content imaging of lipid rafts clusters in db/db mice. PLoS ONE 2014, 9, e106065. [Google Scholar] [CrossRef]
- Mitsutake, S.; Zama, K.; Yokota, H.; Yoshida, T.; Tanaka, M.; Mitsui, M.; Ikawa, M.; Okabe, M.; Tanaka, Y.; Yamashita, T.; et al. Dynamic modification of sphingomyelin in lipid microdomains controls development of obesity, fatty liver, and type 2 diabetes. J. Biol. Chem. 2011, 286, 28544–28555. [Google Scholar] [CrossRef] [PubMed]
- Romiti, E.; Meacci, E.; Tanzi, G.; Becciolini, L.; Mitsutake, S.; Farnararo, M.; Ito, M.; Bruni, P. Localization of neutral ceramidase in caveolin-enriched light membranes of murine endothelial cells. FEBS Lett. 2001, 506, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Jasmin, J.F.; Frank, P.G.; Lisanti, M.P. Caveolins and caveolae: Roles in signaling and disease mechanism. In Advances in Experimental Medicine and Biology; Springer Science + Business Media, LCC.: New York, NY, USA, 2012; pp. 3–13. [Google Scholar]
- Garcia-Cardena, G.; Martasek, P.; Masters, B.S.; Skidd, P.M.; Couet, J.; Li, S.; Lisanti, M.P.; Sessa, W.C. Dissecting the interaction between nitric oxide synthase (NOS) and caveolin. Functional significance of the nos caveolin binding domain in vivo. J. Biol. Chem. 1997, 272, 25437–25440. [Google Scholar] [CrossRef] [PubMed]
- Frank, P.G.; Woodman, S.E.; Park, D.S.; Lisanti, M.P. Caveolin, caveolae, and endothelial cell function. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 1161–1168. [Google Scholar] [CrossRef] [PubMed]
- Shaul, P.W. Regulation of endothelial nitric oxide synthase: Location, location, location. Annu. Rev. Physiol. 2002, 64, 749–774. [Google Scholar] [CrossRef]
- Lam, T.Y.; Seto, S.W.; Lau, Y.M.; Au, L.S.; Kwan, Y.W.; Ngai, S.M.; Tsui, K.W. Impairment of the vascular relaxation and differential expression of caveolin-1 of the aorta of diabetic +db/+db mice. Eur. J. Pharmacol. 2006, 546, 134–141. [Google Scholar] [CrossRef]
- Miller, M.A.; Morgan, R.J.; Thompson, C.S.; Mikhailidis, D.P.; Jeremy, J.Y. Adenylate and guanylate cyclase activity in the penis and aorta of the diabetic rat: An in vitro study. Br. J. Urol. 1994, 74, 106–111. [Google Scholar] [CrossRef]
- Samad, F.; Hester, K.D.; Yang, G.; Hannun, Y.A.; Bielawski, J. Altered adipose and plasma sphingolipid metabolism in obesity: A potential mechanism for cardiovascular and metabolic risk. Diabetes 2006, 55, 2579–2587. [Google Scholar] [CrossRef]
- Arora, T.; Velagapudi, V.; Pournaras, D.J.; Welbourn, R.; le Roux, C.W.; Oresic, M.; Backhed, F. Roux-en-Y gastric bypass surgery induces early plasma metabolomic and lipidomic alterations in humans associated with diabetes remission. PLoS ONE 2015, 10, e0126401. [Google Scholar] [CrossRef]
- Fox, T.E.; Bewley, M.C.; Unrath, K.A.; Pedersen, M.M.; Anderson, R.E.; Jung, D.Y.; Jefferson, L.S.; Kim, J.K.; Bronson, S.K.; Flanagan, J.M.; et al. Circulating sphingolipid biomarkers in models of type 1 diabetes. J. Lipid Res. 2011, 52, 509–517. [Google Scholar] [CrossRef]
- Gorska, M.; Baranczuk, E.; Dobrzyn, A. Secretory Zn2+-dependent sphingomyelinase activity in the serum of patients with type 2 diabetes is elevated. Horm. Metab. Res. 2003, 35, 506–507. [Google Scholar] [CrossRef]
- Gorska, M.; Dobrzyn, A.; Baranowski, M. Concentrations of sphingosine and sphinganine in plasma of patients with type 2 diabetes. Med. Sci. Monit. 2005, 11, CR35–CR38. [Google Scholar] [PubMed]
- Donath, M.Y.; Shoelson, S.E. Type 2 diabetes as an inflammatory disease. Nat. Rev. Immunol. 2011, 11, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Dressler, K.A.; Mathias, S.; Kolesnick, R.N. Tumor necrosis factor-alpha activates the sphingomyelin signal transduction pathway in a cell-free system. Science 1992, 255, 1715–1718. [Google Scholar] [CrossRef] [PubMed]
- Wiegmann, K.; Schutze, S.; Machleidt, T.; Witte, D.; Kronke, M. Functional dichotomy of neutral and acidic sphingomyelinases in tumor necrosis factor signaling. Cell 1994, 78, 1005–1015. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Chandrasekera, P.C.; Pippin, J.J. Leptin- and leptin receptor-deficient rodent models: Relevance for human type 2 diabetes. Curr. Diabetes Rev. 2014, 10, 131–145. [Google Scholar] [CrossRef]
- Cohen, M.P.; Clements, R.S.; Hud, E.; Cohen, J.A.; Ziyadeh, F.N. Evolution of renal function abnormalities in the db/db mouse that parallels the development of human diabetic nephropathy. Exp. Nephrol. 1996, 4, 166–171. [Google Scholar]
- Clarke, C.J.; Snook, C.F.; Tani, M.; Matmati, N.; Marchesini, N.; Hannun, Y.A. The extended family of neutral sphingomyelinases. Biochemistry 2006, 45, 11247–11256. [Google Scholar] [CrossRef]
- Schulz, M.K.; Gutterman, D.K.; Freed, J.K. Activation of Neutral Sphingomyelinase Contributes to Nitric Oxide-Mediated Flow-Induced Dilation in the Human Microvasculature. FASEB J. 2020, 34, 1. [Google Scholar] [CrossRef]
- Bautista-Perez, R.; del Valle-Mondragon, L.; Cano-Martinez, A.; Perez-Mendez, O.; Escalante, B.; Franco, M. Involvement of neutral sphingomyelinase in the angiotensin II signaling pathway. Am. J. Physiol. Renal. Physiol. 2015, 308, F1178–F1187. [Google Scholar] [CrossRef]
- Horvath, B.; Orsy, P.; Benyo, Z. Endothelial NOS-mediated relaxations of isolated thoracic aorta of the C57BL/6J mouse: A methodological study. J. Cardiovasc. Pharmacol. 2005, 45, 225–231. [Google Scholar] [CrossRef]
- Hojjati, M.R.; Jiang, X.C. Rapid, specific, and sensitive measurements of plasma sphingomyelin and phosphatidylcholine. J. Lipid Res. 2006, 47, 673–676. [Google Scholar] [CrossRef] [PubMed]
- Raines, M.A.; Kolesnick, R.N.; Golde, D.W. Sphingomyelinase and ceramide activate mitogen-activated protein kinase in myeloid HL-60 cells. J. Biol. Chem. 1993, 268, 14572–14575. [Google Scholar] [CrossRef] [PubMed]
- Linardic, C.M.; Hannun, Y.A. Identification of a distinct pool of sphingomyelin involved in the sphingomyelin cycle. J. Biol. Chem. 1994, 269, 23530–23537. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Vessel | Vasoactive Effects | Proposed Mechanism | Refs. |
|---|---|---|---|---|
| Yorkshire pig | Coronary artery | Transient endothelium- dependent contraction followed by endothelium- dependent relaxation | Vasoconstriction: prostanoid(s) Vasorelaxation: NO | [14] |
| Sprague- Dawley rat | Thoracic aorta | Endothelium-independent relaxation | Inhibition of protein kinase C (PKC) | [15,16] |
| Wistar rat | Thoracic aorta | Partly endothelium-independent relaxation | Endothelium-mediated components are independent of NO or prostanoids Non-endothelial components are independent of PKC | [17] |
| Mongrel dog | Basilar artery | Endothelium-independent contraction | Activation of VDCC and PKC | [18] |
| Wistar rat | Pial venule (60–70 μm in diameter) | Constriction and spasm | Activation of VDCC, PKC, and MAP kinase | [19] |
| Wistar rat | Thoracic aorta | Endothelium-independent relaxation | Inhibition of both Ca2+-dependent and Ca2+- independent (RhoA-/Rho kinase-mediated) contractile pathways | [20] |
| Cow | Coronary artery | Endothelium-dependent relaxation | Ca2+-independent eNOS activation, involving phosphorylation on serine 1179 and dissociation of eNOS from plasma membrane caveolae | [21] |
| Wistar rat | Pulmonary artery | Endothelium-independent contraction | Activation of VDCC, PKCζ, and Rho kinase | [22] |
| Wistar-Kyoto (WKY) and Spontaneously hypertensive rat (SHR) | Carotid artery | SHR: strong endothelium- dependent contraction WKY: weak endothelium- dependent contraction | Vasoconstriction is mediated by PLA2- and COX2-mediated TXA2 release and attenuated by NO | [23,24,25] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruisanchez, É.; Janovicz, A.; Panta, R.C.; Kiss, L.; Párkányi, A.; Straky, Z.; Korda, D.; Liliom, K.; Tigyi, G.; Benyó, Z. Enhancement of Sphingomyelinase-Induced Endothelial Nitric Oxide Synthase-Mediated Vasorelaxation in a Murine Model of Type 2 Diabetes. Int. J. Mol. Sci. 2023, 24, 8375. https://doi.org/10.3390/ijms24098375
Ruisanchez É, Janovicz A, Panta RC, Kiss L, Párkányi A, Straky Z, Korda D, Liliom K, Tigyi G, Benyó Z. Enhancement of Sphingomyelinase-Induced Endothelial Nitric Oxide Synthase-Mediated Vasorelaxation in a Murine Model of Type 2 Diabetes. International Journal of Molecular Sciences. 2023; 24(9):8375. https://doi.org/10.3390/ijms24098375
Chicago/Turabian StyleRuisanchez, Éva, Anna Janovicz, Rita Cecília Panta, Levente Kiss, Adrienn Párkányi, Zsuzsa Straky, Dávid Korda, Károly Liliom, Gábor Tigyi, and Zoltán Benyó. 2023. "Enhancement of Sphingomyelinase-Induced Endothelial Nitric Oxide Synthase-Mediated Vasorelaxation in a Murine Model of Type 2 Diabetes" International Journal of Molecular Sciences 24, no. 9: 8375. https://doi.org/10.3390/ijms24098375
APA StyleRuisanchez, É., Janovicz, A., Panta, R. C., Kiss, L., Párkányi, A., Straky, Z., Korda, D., Liliom, K., Tigyi, G., & Benyó, Z. (2023). Enhancement of Sphingomyelinase-Induced Endothelial Nitric Oxide Synthase-Mediated Vasorelaxation in a Murine Model of Type 2 Diabetes. International Journal of Molecular Sciences, 24(9), 8375. https://doi.org/10.3390/ijms24098375