
An Insight into the Arising Role of MicroRNAs in Hepatocellular Carcinoma: Future Diagnostic and Therapeutic Approaches

 ,
,  , ,
, ,  ,
,  ,
, 

Abstract
1. Introduction
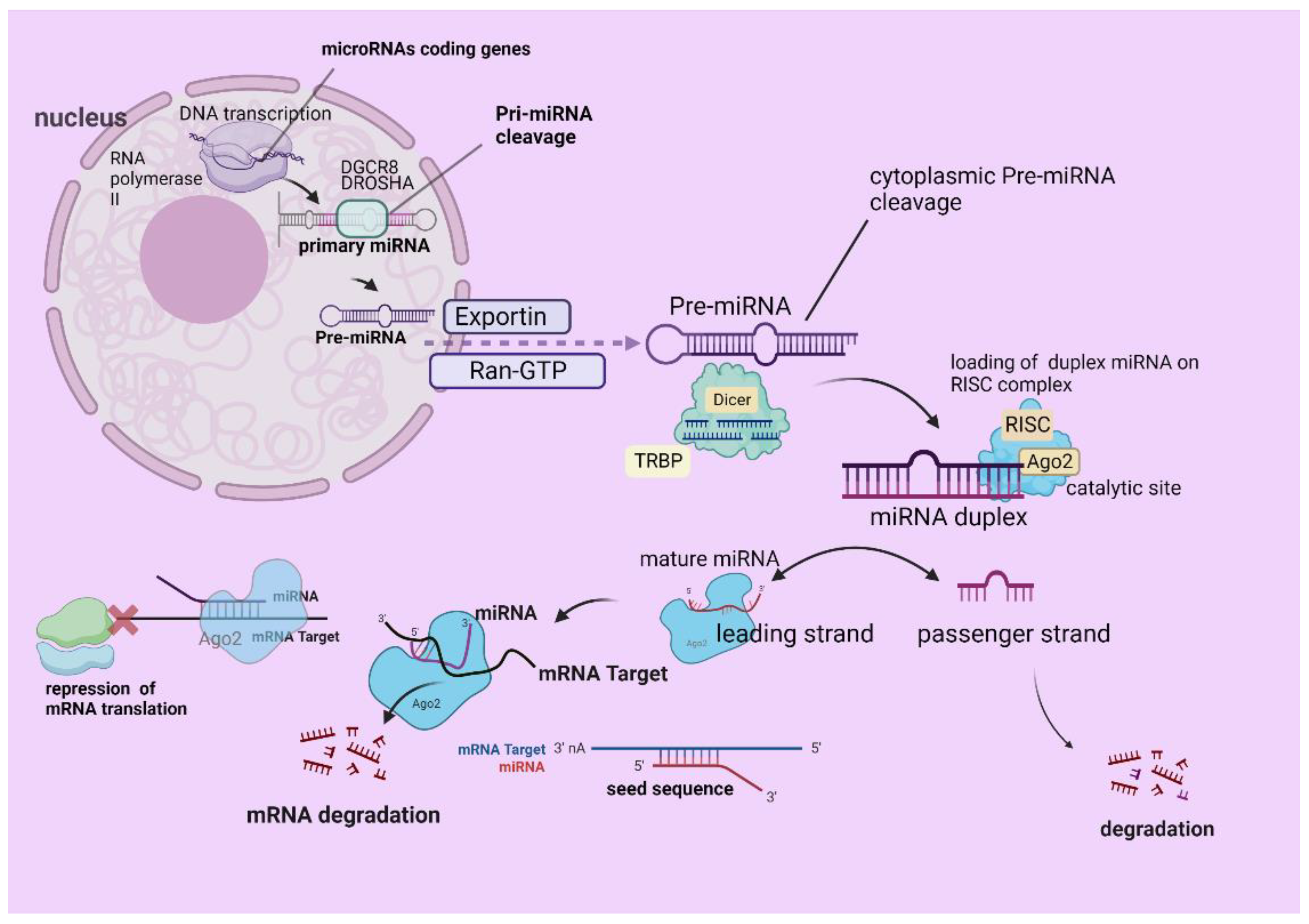
2. A Glance through the Canonical MiRNA Biogenesis
3. The Emerging Role of MiRNAs in HCC
3.1. The Oncogenic MiRNAs in HCC
3.2. MiRNAs as Tumor Suppressors for HCC
3.3. The Interplay between MiRNAs and the Major HCC Predisposing Diseases
3.4. MiRNAs Signature in Premalignant State, HCC Staging, and Prognosis
3.5. MiRNAs as Predictive and Prognostic Biomarkers for HCC
3.6. MiRNAs as Diagnostic or Monitoring Tools
4. The Interplay between EVs and MiRNAs in HCC
5. The Interplay between MiRNAs and Autophagy in HCC
6. The Interplay between MiRNAs and Microbiome in HCC
7. Future Therapeutic Opportunities
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rumgay, H.; Arnold, M.; Ferlay, J.; Lesi, O.; Cabasag, C.J.; Vignat, J.; Laversanne, M.; McGlynn, K.A.; Soerjomataram, I. Global burden of primary liver cancer in 2020 and predictions to 2040. J. Hepatol. 2022, 77, 1598–1606. [Google Scholar] [CrossRef] [PubMed]
- Vogel, A.; Meyer, T.; Sapisochin, G.; Salem, R.; Saborowski, A. Hepatocellular carcinoma. Lancet 2022, 400, 1345–1362. [Google Scholar] [CrossRef]
- Kotsiliti, E. Understanding HCC sex disparities. Nat. Rev. Gastroenterol. Hepatol. 2022, 19, 147. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, L. Changes in the Epidemiology of Hepatocellular Carcinoma in Asia. Cancers 2022, 14, 4473. [Google Scholar] [CrossRef]
- Vitale, A.; Scarpa, M.; Caputo, D.; Spoletini, G.; Giovinazzo, F. (Eds.) Novel Insights into the Treatment of HCC and Liver Tumors; Frontiers Media SA: Lausanne, Switzerland, 2022. [Google Scholar]
- Da Costa, A.C.; Sodergren, M.; Jayant, K.; Cruz, F.S.; Spalding, D.; Pai, M.; Habib, N. Radiofrequency combined with immunomodulation for hepatocellular carcinoma: State of the art and innovations. World J. Gastroenterol. 2020, 26, 2040–2048. [Google Scholar] [CrossRef]
- Koustas, E.; Trifylli, E.-M.; Sarantis, P.; Papadopoulos, N.; Papanikolopoulos, K.; Aloizos, G.; Damaskos, C.; Garmpis, N.; Garmpi, A.; Karamouzis, M.V. The Emerging Role of MicroRNAs and Autophagy Mechanism in Pancreatic Cancer Progression: Future Therapeutic Approaches. Genes 2022, 13, 1868. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Wang, X.; Lv, L.; Liu, J.; Xing, H.; Song, Y.; Xie, M.; Lei, T.; Zhang, N.; Yang, M. The emerging role of microRNAs and long noncoding RNAs in drug resistance of hepatocellular carcinoma. Mol. Cancer 2019, 18, 147. [Google Scholar] [CrossRef]
- Peng, Y.; Croce, C.M. The role of MicroRNAs in human cancer. Signal Transduct. Target. Ther. 2016, 1, 15004. [Google Scholar] [CrossRef]
- Miroshnichenko, S.; Patutina, O. Enhanced Inhibition of Tumorigenesis Using Combinations of miRNA-Targeted Therapeutics. Front. Pharmacol. 2019, 10, 488. [Google Scholar] [CrossRef]
- Rani, V.; Sengar, R.S. Biogenesis and mechanisms of microRNA-mediated gene regulation. Biotechnol. Bioeng. 2022, 119, 685–692. [Google Scholar] [CrossRef]
- Lee, Y.; Kim, M.; Han, J.; Yeom, K.-H.; Lee, S.; Baek, S.H.; Kim, V.N. MicroRNA genes are transcribed by RNA polymerase II. EMBO J. 2004, 23, 4051–4060. [Google Scholar] [CrossRef]
- Lee, D.; Shin, C. Emerging roles of DROSHA beyond primary microRNA processing. RNA Biol. 2018, 15, 186–193. [Google Scholar] [CrossRef]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef]
- Paturi, S.; Deshmukh, M.V. A Glimpse of “Dicer Biology” through the Structural and Functional Perspective. Front. Mol. Biosci. 2021, 8, 643657. [Google Scholar] [CrossRef] [PubMed]
- Doench, J.G.; Sharp, P.A. Specificity of microRNA target selection in translational repression. Genes Dev. 2004, 18, 504–511. [Google Scholar] [CrossRef]
- Medley, J.C.; Panzade, G.; Zinovyeva, A.Y. microRNA strand selection: Unwinding the rules. Wiley Interdiscip. Rev. RNA 2021, 12, e1627. [Google Scholar] [CrossRef]
- AGO2 Argonaute RISC Catalytic Component 2 [Homo Sapiens (Human)]-Gene-NCBI. Nih.gov. Available online: https://www.ncbi.nlm.nih.gov/gene/27161 (accessed on 5 March 2023).
- Jungers, C.F.; Djuranovic, S. Modulation of miRISC-Mediated Gene Silencing in Eukaryotes. Front. Mol. Biosci. 2022, 9, 832916. [Google Scholar] [CrossRef] [PubMed]
- Ergin, K.; Çetinkaya, R. Regulation of MicroRNAs. In Methods in Molecular Biology; Springer: New York, NY, USA, 2022; pp. 1–32. [Google Scholar]
- Naeli, P.; Winter, T.; Hackett, A.P.; Alboushi, L.; Jafarnejad, S.M. The intricate balance between microRNA-induced mRNA decay and translational repression. FEBS J. 2022. [Google Scholar] [CrossRef]
- Stavast, C.; Erkeland, S. The Non-Canonical Aspects of MicroRNAs: Many Roads to Gene Regulation. Cells 2019, 8, 1465. [Google Scholar] [CrossRef]
- Smolarz, B.; Durczyński, A.; Romanowicz, H.; Szyłło, K.; Hogendorf, P. miRNAs in Cancer (Review of Literature). Int. J. Mol. Sci. 2022, 23, 2805. [Google Scholar] [CrossRef] [PubMed]
- Hussen, B.M.; Hidayat, H.J.; Salihi, A.; Sabir, D.K.; Taheri, M.; Ghafouri-Fard, S. MicroRNA: A signature for cancer progression. Biomed. Pharmacother. 2021, 138, 111528. [Google Scholar] [CrossRef]
- Sarkar, F.H.; Li, Y.; Wang, Z.; Kong, D.; Ali, S. Implication of microRNAs in drug resistance for designing novel cancer therapy. Drug Resist. Updates 2010, 13, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Chen, E.; Xu, X.; Liu, R.; Liu, T. Small but Heavy Role: MicroRNAs in Hepatocellular Carcinoma Progression. BioMed Res. Int. 2018, 2018, 6784607. [Google Scholar] [CrossRef] [PubMed]
- Otmani, K.; Rouas, R.; Lewalle, P. OncomiRs as noncoding RNAs having functions in cancer: Their role in immune suppression and clinical implications. Front. Immunol. 2022, 13, 913951. [Google Scholar] [CrossRef]
- Oura, K.; Morishita, A.; Masaki, T. Molecular and Functional Roles of MicroRNAs in the Progression of Hepatocellular Carcinoma—A Review. Int. J. Mol. Sci. 2020, 21, 8362. [Google Scholar] [CrossRef]
- Wang, L.; Wu, J.; Xie, C. miR-92a promotes hepatocellular carcinoma cells proliferation and invasion by FOXA2 targeting. Iran. J. Basic Med. Sci. 2017, 20, 783–790. [Google Scholar] [CrossRef] [PubMed]
- Huo, W.; Du, M.; Pan, X.; Zhu, X.; Gao, Y.; Li, Z. miR-203a-3p.1 targets IL-24 to modulate hepatocellular carcinoma cell growth and metastasis. FEBS Open Bio 2017, 7, 1085–1091. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Zhang, W.; Chang, Z.; Pan, Y.; Zong, H.; Fan, Q.; Wang, L. miR-4417 Targets Tripartite Motif-Containing 35 (TRIM35) and Regulates Pyruvate Kinase Muscle 2 (PKM2) Phosphorylation to Promote Proliferation and Suppress Apoptosis in Hepatocellular Carcinoma Cells. Med. Sci. Monit. 2017, 23, 1741–1750. [Google Scholar] [CrossRef]
- Lai, C.-Y.; Yeh, K.-Y.; Lin, C.-Y.; Hsieh, Y.-W.; Lai, H.-H.; Chen, J.-R.; Hsu, C.-C.; Her, G.M. MicroRNA-21 Plays Multiple Oncometabolic Roles in the Process of NAFLD-Related Hepatocellular Carcinoma via PI3K/AKT, TGF-β, and STAT3 Signaling. Cancers 2021, 13, 940. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Tao, R.; Wang, S.; Wang, C.; Zhao, X.; Zhao, H.; Li, L.; Zhu, S.; He, Y.; Jiang, X.; et al. MicroRNA-21 promotes cell proliferation in human hepatocellular carcinoma partly by targeting HEPN1. Tumor Biol. 2015, 36, 5467–5472. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Lu, J.; Cao, J.; Ma, B.; Gao, C.; Qi, F. MicroRNA-18a promotes hepatocellular carcinoma proliferation, migration, and invasion by targeting Bcl2L10. Onco Targets Ther. 2018, 11, 7919–7934. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Wen, H.; Jing, L.; Yang, Y.; Wang, W.; Liang, X.; Nan, K.; Yao, Y.; Tian, T. MicroRNA-155-5p promotes hepatocellular carcinoma progression by suppressing PTEN through the PI3K/Akt pathway. Cancer Sci. 2017, 108, 620–631. [Google Scholar] [CrossRef]
- Jin, W.; Zhong, N.; Wang, L.; Yu, J.; Yin, F.; Zhang, K. MiR-331-3p Inhibition of the Hepatocellular Carcinoma (HCC) Bel-7402 Cell Line by Down-Regulation of E2F1. J. Nanosci. Nanotechnol. 2019, 19, 5476–5482. [Google Scholar] [CrossRef] [PubMed]
- Xie, B.-H.; He, X.; Hua, R.-X.; Zhang, B.; Tan, G.-S.; Xiong, S.-Q.; Liu, L.-S.; Chen, W.; Yang, J.-Y.; Wang, X.-N.; et al. Mir-765 promotes cell proliferation by downregulating INPP4B expression in human hepatocellular carcinoma. Cancer Biomark. 2016, 16, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-L.; Xu, Q.-P.; Guo, F.; Guan, W.-H. MicroRNA-302d downregulates TGFBR2 expression and promotes hepatocellular carcinoma growth and invasion. Exp. Ther. Med. 2017, 13, 681–687. [Google Scholar] [CrossRef]
- Chang, R.-M.; Xiao, S.; Lei, X.; Yang, H.; Fang, F.; Yang, L.-Y. miRNA-487a Promotes Proliferation and Metastasis in Hepatocellular Carcinoma. Clin. Cancer Res. 2017, 23, 2593–2604. [Google Scholar] [CrossRef] [PubMed]
- Lei, Z.; Tang, X.; Si, A.; Yang, P.; Wang, L.; Luo, T.; Guo, G.; Zhang, Q.; Cheng, Z. microRNA-454 promotes liver tumor-initiating cell expansion by regulating SOCS6. Exp. Cell Res. 2020, 390, 111955. [Google Scholar] [CrossRef]
- Guo, Z.; Li, J.; Sun, J.; Sun, L.; Zhou, Y.; Yu, Z. miR-346 Promotes HCC Progression by Suppressing Breast Cancer Metastasis Suppressor 1 Expression. Oncol. Res. Featur. Preclin. Clin. Cancer Ther. 2018, 26, 1073–1081. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Wang, H.; Han, S. MiR-3910 Promotes the Growth and Migration of Cancer Cells in the Progression of Hepatocellular Carcinoma. Dig. Dis. Sci. 2017, 62, 2812–2820. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, C.; Zhao, Q.; Wei, W.; Dong, Z.; Shao, L.; Li, J.; Wu, W.; Zhang, H.; Huang, H.; et al. The MiR-873/NDFIP1 Axis Promotes Hepatocellular Carcinoma Growth and Metastasis through the AKT/MTOR-Mediated Warburg Effect. Am. J. Cancer Res. 2019, 9, 927–944. [Google Scholar] [PubMed]
- Li, B.; He, L.; Zuo, D.; He, W.; Wang, Y.; Zhang, Y.; Liu, W.; Yuan, Y. Mutual Regulation of MiR-199a-5p and HIF-1α Modulates the Warburg Effect in Hepatocellular Carcinoma. J. Cancer 2017, 8, 940–949. [Google Scholar] [CrossRef]
- Feng, J.; Li, J.; Wu, L.; Yu, Q.; Ji, J.; Wu, J.; Dai, W.; Guo, C. Emerging roles and the regulation of aerobic glycolysis in hepatocellular carcinoma. J. Exp. Clin. Cancer Res. 2020, 39, 126. [Google Scholar] [CrossRef]
- Zeng, B.; Shi, W.; Tan, G. MiR-199a/b-3p inhibits gastric cancer cell proliferation via down-regulating PAK4/MEK/ERK signaling pathway. BMC Cancer 2018, 18, 34. [Google Scholar] [CrossRef]
- Huang, Z.; Wen, J.; Yu, J.; Liao, J.; Liu, S.; Cai, N.; Liang, H.; Chen, X.; Ding, Z.; Zhang, B. MicroRNA-148a-3p inhibits progression of hepatocelluar carcimoma by repressing SMAD2 expression in an Ago2 dependent manner. J. Exp. Clin. Cancer Res. 2020, 39, 150. [Google Scholar] [CrossRef]
- Xu, T.; Zhu, Y.; Xiong, Y.; Ge, Y.-Y.; Yun, J.-P.; Zhuang, S.-M. MicroRNA-195 suppresses tumorigenicity and regulates G1/S transition of human hepatocellular carcinoma cells. Hepatology 2009, 50, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.-M.; Shi, X.-L.; Xing, K.-L.; Zhou, H.-X.; Lu, L.-L.; Wu, W.-Z. miR-296-5p suppresses stem cell potency of hepatocellular carcinoma cells via regulating Brg1/Sall4 axis. Cell. Signal. 2020, 72, 109650. [Google Scholar] [CrossRef]
- Shi, D.-M.; Li, L.-X.; Bian, X.-Y.; Shi, X.-J.; Lu, L.-L.; Zhou, H.-X.; Pan, T.-J.; Zhou, J.; Fan, J.; Wu, W.-Z. miR-296-5p suppresses EMT of hepatocellular carcinoma via attenuating NRG1/ERBB2/ERBB3 signaling. J. Exp. Clin. Cancer Res. 2018, 37, 294. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tai, Q.; Zhang, J.; Kang, J.; Gao, F.; Zhong, F.; Cai, L.; Fang, F.; Gao, Y. MiRNA-206 inhibits hepatocellular carcinoma cell proliferation and migration but promotes apoptosis by modulating cMET expression. Acta Biochim. Biophys. Sin. 2019, 51, 243–253. [Google Scholar] [CrossRef]
- Yang, Y.-L.; Tsai, M.-C.; Chang, Y.-H.; Wang, C.-C.; Chu, P.-Y.; Lin, H.-Y.; Huang, Y.-H. MIR29A Impedes Metastatic Behaviors in Hepatocellular Carcinoma via Targeting LOX, LOXL2, and VEGFA. Int. J. Mol. Sci. 2021, 22, 6001. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.-L.; Chang, Y.-H.; Li, C.-J.; Huang, Y.-H.; Tsai, M.-C.; Chu, P.-Y.; Lin, H.-Y. New Insights into the Role of miR-29a in Hepatocellular Carcinoma: Implications in Mechanisms and Theragnostics. J. Pers. Med. 2021, 11, 219. [Google Scholar] [CrossRef]
- You, Y.; Que, K.; Zhou, Y.; Zhang, Z.; Zhao, X.; Gong, J.; Liu, Z. MicroRNA-766-3p Inhibits Tumour Progression by Targeting Wnt3a in Hepatocellular Carcinoma. Mol. Cells 2018, 41, 830–841. [Google Scholar] [CrossRef]
- Nie, X.; Liu, Y.; Chen, W.-D.; Wang, Y.-D. Interplay of miRNAs and Canonical Wnt Signaling Pathway in Hepatocellular Carcinoma. Front. Pharmacol. 2018, 9, 657. [Google Scholar] [CrossRef]
- Roy, S.; Hooiveld, G.J.; Seehawer, M.; Caruso, S.; Heinzmann, F.; Schneider, A.T.; Frank, A.K.; Cardenas, D.V.; Sonntag, R.; Luedde, M.; et al. microRNA 193a-5p Regulates Levels of Nucleolar- and Spindle-Associated Protein 1 to Suppress Hepatocarcinogenesis. Gastroenterology 2018, 155, 1951–1966.e26. [Google Scholar] [CrossRef]
- Wang, Y.; Ju, L.; Xiao, F.; Liu, H.; Luo, X.; Chen, L.; Lu, Z.; Bian, Z. Downregulation of nucleolar and spindle-associated protein 1 expression suppresses liver cancer cell function. Exp. Ther. Med. 2019, 17, 2969–2978. [Google Scholar] [CrossRef] [PubMed]
- Gramantieri, L.; Pollutri, D.; Gagliardi, M.; Giovannini, C.; Quarta, S.; Ferracin, M.; Casadei-Gardini, A.; Callegari, E.; De Carolis, S.; Marinelli, S.; et al. MiR-30e-3p Influences Tumor Phenotype through MDM2/TP53 Axis and Predicts Sorafenib Resistance in Hepatocellular Carcinoma. Cancer Res. 2020, 80, 1720–1734. [Google Scholar] [CrossRef]
- Gramantieri, L.; Giovannini, C.; Piscaglia, F.; Fornari, F. MicroRNAs as Modulators of Tumor Metabolism, Microenvironment, and Immune Response in Hepatocellular Carcinoma. J. Hepatocell. Carcinoma 2021, 8, 369–385. [Google Scholar] [CrossRef]
- Ye, Y.; Wei, Y.; Xu, Y.; Li, Y.; Wang, R.; Chen, J.; Zhou, Y.; Fu, Z.; Chen, Y.; Wang, X.; et al. Induced MiR-1249 expression by aberrant activation of Hedegehog signaling pathway in hepatocellular carcinoma. Exp. Cell Res. 2017, 355, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Gatfield, D.; Le Martelot, G.; Vejnar, C.E.; Gerlach, D.; Schaad, O.; Fleury-Olela, F.; Ruskeepää, A.-L.; Oresic, M.; Esau, C.C.; Zdobnov, E.M.; et al. Integration of microRNA miR-122 in hepatic circadian gene expression. Genes Dev. 2009, 23, 1313–1326. [Google Scholar] [CrossRef]
- Castro-Muñoz, L.J.; Ulloa, E.V.; Sahlgren, C.; Lizano, M.; De La Cruz-Hernández, E.; Contreras-Paredes, A. Modulating epigenetic modifications for cancer therapy (Review). Oncol. Rep. 2023, 49, 59. [Google Scholar] [CrossRef] [PubMed]
- Fornari, F.; Gramantieri, L.; Giovannini, C.; Veronese, A.; Ferracin, M.; Sabbioni, S.; Calin, G.A.; Grazi, G.L.; Croce, C.M.; Tavolari, S.; et al. MiR-122/Cyclin G1 Interaction Modulates p53 Activity and Affects Doxorubicin Sensitivity of Human Hepatocarcinoma Cells. Cancer Res. 2009, 69, 5761–5767. [Google Scholar] [CrossRef] [PubMed]
- Stella, L.; Santopaolo, F.; Gasbarrini, A.; Pompili, M.; Ponziani, F.R. Viral hepatitis and hepatocellular carcinoma: From molecular pathways to the role of clinical surveillance and antiviral treatment. World J. Gastroenterol. 2022, 28, 2251–2281. [Google Scholar] [CrossRef]
- Loureiro, D.; Tout, I.; Narguet, S.; Benazzouz, S.M.; Mansouri, A.; Asselah, T. miRNAs as Potential Biomarkers for Viral Hepatitis B and C. Viruses 2020, 12, 1440. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.-X.; Tang, H. Complex interactions between microRNAs and hepatitis B/C viruses. World J. Gastroenterol. 2014, 20, 13477–13492. [Google Scholar] [CrossRef] [PubMed]
- Kunden, R.D.; Khan, J.Q.; Ghezelbash, S.; Wilson, J.A. The Role of the Liver-Specific microRNA, miRNA-122 in the HCV Replication Cycle. Int. J. Mol. Sci. 2020, 21, 5677. [Google Scholar] [CrossRef] [PubMed]
- Ottosen, S.; Parsley, T.B.; Yang, L.; Zeh, K.; van Doorn, L.-J.; van der Veer, E.; Raney, A.K.; Hodges, M.R.; Patick, A.K. In Vitro Antiviral Activity and Preclinical and Clinical Resistance Profile of Miravirsen, a Novel Anti-Hepatitis C Virus Therapeutic Targeting the Human Factor miR-122. Antimicrob. Agents Chemother. 2015, 59, 599–608. [Google Scholar] [CrossRef]
- Li, H.; Jiang, J.-D.; Peng, Z.-G. MicroRNA-mediated interactions between host and hepatitis C virus. World J. Gastroenterol. 2016, 22, 1487–1496. [Google Scholar] [CrossRef]
- Ahmad, I.; Valverde, A.; Siddiqui, H.; Schaller, S.; Naqvi, A.R. Viral MicroRNAs: Interfering the Interferon Signaling. Curr. Pharm. Des. 2020, 26, 446–454. [Google Scholar] [CrossRef]
- Sartorius, K.; Makarova, J.; Sartorius, B.; An, P.; Winkler, C.; Chuturgoon, A.; Kramvis, A. The Regulatory Role of MicroRNA in Hepatitis-B Virus-Associated Hepatocellular Carcinoma (HBV-HCC) Pathogenesis. Cells 2019, 8, 1504. [Google Scholar] [CrossRef]
- Mahmoudian-Sani, M.R.; Asgharzade, S.; Alghasi, A.; Saeedi-Boroujeni, A.; Sadati, S.J.A.; Moradi, M.T. MicroRNA-122 in patients with hepatitis B and hepatitis B virus-associated hepatocellular carcinoma. J. Gastrointest. Oncol. 2019, 10, 789–796. [Google Scholar] [CrossRef]
- Sartorius, K.; Swadling, L.; An, P.; Makarova, J.; Winkler, C.; Chuturgoon, A.; Kramvis, A. The Multiple Roles of Hepatitis B Virus X Protein (HBx) Dysregulated MicroRNA in Hepatitis B Virus-Associated Hepatocellular Carcinoma (HBV-HCC) and Immune Pathways. Viruses 2020, 12, 746. [Google Scholar] [CrossRef]
- Xie, K.-L.; Zhang, Y.-G.; Liu, J.; Zeng, Y.; Wu, H. MicroRNAs Associated with HBV Infection and HBV-related HCC. Theranostics 2014, 4, 1176–1192. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Wan, P.; Zhang, Y.; Tan, Q.; Qudus, M.S.; Yue, Z.; Luo, W.; Zhang, W.; Ouyang, J.; Li, Y.; et al. Innate Immunity, Inflammation, and Intervention in HBV Infection. Viruses 2022, 14, 2275. [Google Scholar] [CrossRef] [PubMed]
- Dong, M.-L.; Wen, X.; He, X.; Ren, J.-H.; Yu, H.-B.; Qin, Y.-P.; Yang, Z.; Yang, M.-L.; Zhou, C.-Y.; Zhang, H.; et al. HBx Mediated Increase of DDX17 Contributes to HBV-Related Hepatocellular Carcinoma Tumorigenesis. Front. Immunol. 2022, 13, 871558. [Google Scholar] [CrossRef]
- Fang, Z.; Dou, G.; Wang, L. MicroRNAs in the Pathogenesis of Nonalcoholic Fatty Liver Disease. Int. J. Biol. Sci. 2021, 17, 1851–1863. [Google Scholar] [CrossRef]
- Torres, J.-L.; Novo-Veleiro, I.; Manzanedo, L.; Alvela-Suárez, L.; Macías, R.; Laso, F.-J.; Marcos, M. Role of microRNAs in alcohol-induced liver disorders and non-alcoholic fatty liver disease. World J. Gastroenterol. 2018, 24, 4104–4118. [Google Scholar] [CrossRef]
- Chai, C.; Rivkin, M.; Berkovits, L.; Simerzin, A.; Zorde-Khvalevsky, E.; Rosenberg, N.; Klein, S.; Yaish, D.; Durst, R.; Shpitzen, S.; et al. Metabolic Circuit Involving Free Fatty Acids, microRNA 122, and Triglyceride Synthesis in Liver and Muscle Tissues. Gastroenterology 2017, 153, 1404–1415. [Google Scholar] [CrossRef]
- Li, M.; Tang, Y.; Wu, L.; Mo, F.; Wang, X.; Li, H.; Qi, R.; Zhang, H.; Srivastava, A.; Ling, C. The hepatocyte-specific HNF4α/miR-122 pathway contributes to iron overload–mediated hepatic inflammation. Blood 2017, 130, 1041–1051. [Google Scholar] [CrossRef]
- Chai, C.; Cox, B.; Yaish, D.; Gross, D.; Rosenberg, N.; Amblard, F.; Shemuelian, Z.; Gefen, M.; Korach, A.; Tirosh, O.; et al. Agonist of RORA Attenuates Nonalcoholic Fatty Liver Progression in Mice via Up-regulation of MicroRNA 122. Gastroenterology 2020, 159, 999–1014.e9. [Google Scholar] [CrossRef]
- De Sousa, M.C.; Calo, N.; Sobolewski, C.; Gjorgjieva, M.; Clément, S.; Maeder, C.; Dolicka, D.; Fournier, M.; Vinet, L.; Montet, X.; et al. Mir-21 Suppression Promotes Mouse Hepatocarcinogenesis. Cancers 2021, 13, 4983. [Google Scholar] [CrossRef] [PubMed]
- Fodor, A.; Lazar, A.; Buchman, C.; Tiperciuc, B.; Orasan, O.; Cozma, A. MicroRNAs: The Link between the Metabolic Syndrome and Oncogenesis. Int. J. Mol. Sci. 2021, 22, 6337. [Google Scholar] [CrossRef]
- McDaniel, K.; Huang, L.; Sato, K.; Wu, N.; Annable, T.; Zhou, T.; Ramos-Lorenzo, S.; Wan, Y.; Huang, Q.; Francis, H.; et al. The let-7/Lin28 axis regulates activation of hepatic stellate cells in alcoholic liver injury. J. Biol. Chem. 2017, 292, 11336–11347. [Google Scholar] [CrossRef] [PubMed]
- Mohr, R.; Özdirik, B.; Lambrecht, J.; Demir, M.; Eschrich, J.; Geisler, L.; Hellberg, T.; Loosen, S.H.; Luedde, T.; Tacke, F.; et al. From Liver Cirrhosis to Cancer: The Role of Micro-RNAs in Hepatocarcinogenesis. Int. J. Mol. Sci. 2021, 22, 1492. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Ma, H.; Wu, R.; Wang, H.; Xu, H.; Li, S.; Wang, G.; Lv, G.; Niu, J. Identification of Liver Fibrosis-Related MicroRNAs in Human Primary Hepatic Stellate Cells Using High-Throughput Sequencing. Genes 2022, 13, 2201. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Cai, X.; Wang, Z.; Huang, L.; Wang, C.; Jiang, S.; Hua, Y.; Liu, Q. miR-200c Accelerates Hepatic Stellate Cell-Induced Liver Fibrosis via Targeting the FOG2/PI3K Pathway. BioMed Res. Int. 2017, 2017, 2670658. [Google Scholar] [CrossRef]
- Trifylli, E.-M.; Kriebardis, A.G.; Koustas, E.; Papadopoulos, N.; Deutsch, M.; Aloizos, G.; Fortis, S.P.; Papageorgiou, E.G.; Tsagarakis, A.; Manolakopoulos, S. The Emerging Role of Extracellular Vesicles and Autophagy Machinery in NASH—Future Horizons in NASH Management. Int. J. Mol. Sci. 2022, 23, 12185. [Google Scholar] [CrossRef]
- Kogure, A.; Kosaka, N.; Ochiya, T.; Kogure, A.; Kosaka, N.; Ochiya, T. Cross-talk between cancer cells and their neighbors via miRNA in extracellular vesicles: An emerging player in cancer metastasis. J. Biomed. Sci. 2019, 26, 7. [Google Scholar] [CrossRef]
- Liu, L.; Lu, L.; Zheng, A.; Xie, J.; Xue, Q.; Wang, F.; Wang, X.; Zhou, H.; Tong, X.; Li, Y.; et al. MiR-199a-5p and let-7c cooperatively inhibit migration and invasion by targeting MAP4K3 in hepatocellular carcinoma. Oncotarget 2017, 8, 13666–13677. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Y.; Zheng, N.; Teng, F.; Bao, L.; Liu, F.; Zhang, M.; Guo, M.; Guo, W.; Ding, G.; Wang, Q. MiR-199a/b-5p inhibits hepatocellular carcinoma progression by post-transcriptionally suppressing ROCK1. Oncotarget 2017, 8, 67169–67180. [Google Scholar] [CrossRef]
- Song, T.; Li, L.; Wu, S.; Liu, Y.; Guo, C.; Wang, W.; Dai, L.; Zhang, T.; Wu, H.; Su, B. Peripheral Blood Genetic Biomarkers for the Early Diagnosis of Hepatocellular Carcinoma. Front. Oncol. 2021, 11, 583714. [Google Scholar] [CrossRef]
- Nagy, Á.; Lánczky, A.; Menyhárt, O.; Győrffy, B. Validation of miRNA prognostic power in hepatocellular carcinoma using expression data of independent datasets. Sci. Rep. 2018, 8, 9227. [Google Scholar] [CrossRef]
- Morishita, A.; Oura, K.; Tadokoro, T.; Fujita, K.; Tani, J.; Masaki, T. MicroRNAs in the Pathogenesis of Hepatocellular Carcinoma: A Review. Cancers 2021, 13, 514. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Liu, M.; Qu, S.; Ma, J.; Zhang, Y.; Shi, T.; Wen, H.; Yang, Y.; Wang, S.; Wang, J.; et al. Exosomal microRNA-32-5p induces multidrug resistance in hepatocellular carcinoma via the PI3K/Akt pathway. J. Exp. Clin. Cancer Res. 2018, 37, 52. [Google Scholar] [CrossRef]
- Sasaki, R.; Kanda, T.; Yokosuka, O.; Kato, N.; Matsuoka, S.; Moriyama, M. Exosomes and Hepatocellular Carcinoma: From Bench to Bedside. Int. J. Mol. Sci. 2019, 20, 1406. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Liu, M.; Li, F.; Qian, L.; Zhang, P.; Lv, F.; Cheng, W.; Hou, R. MiR-221 Promotes Hepatocellular Carcinoma Cells Migration via Targeting PHF2. BioMed Res. Int. 2019, 2019, 4371405. [Google Scholar] [CrossRef]
- Yuan, B.; Liang, Y.; Wang, D.; Luo, F. MiR-940 inhibits hepatocellular carcinoma growth and correlates with prognosis of hepatocellular carcinoma patients. Cancer Sci. 2015, 106, 819–824. [Google Scholar] [CrossRef]
- Wei, Q.; Zhao, L.; Jiang, L.; Bi, J.; Yu, Z.; Zhao, L.; Song, X.; Sun, M.; Chen, Y.; Wei, M. Prognostic relevance of miR-137 and its liver microenvironment regulatory target gene AFM in hepatocellular carcinoma. J. Cell. Physiol. 2019, 234, 11888–11899. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Bo, X.; Zheng, Q.; Xiao, X.; Wu, L.; Li, B. miR-296 inhibits proliferation and induces apoptosis by targeting FGFR1 in human hepatocellular carcinoma. FEBS Lett. 2016, 590, 4252–4262. [Google Scholar] [CrossRef] [PubMed]
- Jun, L.; Yang, G.; Zhisu, L. The utility of serum exosomal microRNAs in hepatocellular carcinoma. Biomed. Pharmacother. 2019, 111, 1221–1227. [Google Scholar] [CrossRef] [PubMed]
- Yokota, Y.; Noda, T.; Okumura, Y.; Kobayashi, S.; Iwagami, Y.; Yamada, D.; Tomimaru, Y.; Akita, H.; Gotoh, K.; Takeda, Y.; et al. Serum exosomal miR-638 is a prognostic marker of HCC via downregulation of VE-cadherin and ZO-1 of endothelial cells. Cancer Sci. 2021, 112, 1275–1288. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, D.; Jiang, J.; Dong, L. Loss of miR-638 promotes invasion and epithelial-mesenchymal transition by targeting SOX2 in hepatocellular carcinoma. Oncol. Rep. 2017, 37, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Li, B.; Zhao, S.; Du, H.; Du, Y. Exosomal miR-638 Inhibits Hepatocellular Carcinoma Progression by Targeting SP1. OncoTargets Ther. 2020, 13, 6709–6720. [Google Scholar] [CrossRef]
- Chen, S.; Fu, Z.; Wen, S.; Yang, X.; Yu, C.; Zhou, W.; Lin, Y.; Lv, Y. Expression and Diagnostic Value of miR-497 and miR-1246 in Hepatocellular Carcinoma. Front. Genet. 2021, 12, 666306. [Google Scholar] [CrossRef]
- Peng, Y.; Huang, D.; Qing, X.; Tang, L.; Shao, Z. Investigation of MiR-92a as a Prognostic Indicator in Cancer Patients: A Meta-Analysis. J. Cancer 2019, 10, 4430–4441. [Google Scholar] [CrossRef]
- Cao, X.; Yang, Q.; Yu, Q. Increased Expression of miR-487b Is Associated With Poor Prognosis and Tumor Progression of HBV-Related Hepatocellular Carcinoma. Open Forum Infect. Dis. 2020, 7, ofaa498. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Li, J.; Qian, Y.; Liu, W.; Liang, J.; Huang, Z.; Wang, S.; Zhao, C. Identification of plasma miR-148a as a noninvasive biomarker for hepatocellular carcinoma. Clin. Res. Hepatol. Gastroenterol. 2019, 43, 585–593. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.; Huang, S.; He, R.; Rong, M.; Dang, Y.; Chen, G. Decreased expression and clinical significance of miR-148a in hepatocellular carcinoma tissues. Eur. J. Med. Res. 2014, 19, 68. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Deng, X.; Zeng, X.; Peng, X. The Role of Mir-148a in Cancer. J. Cancer 2016, 7, 1233–1241. [Google Scholar] [CrossRef]
- Fu, X.; Calin, G.A. miR-122 and hepatocellular carcinoma: From molecular biology to therapeutics. Ebiomedicine 2018, 37, 17–18. [Google Scholar] [CrossRef] [PubMed]
- Mariam, A.; Miller-Atkins, G.; Moro, A.; Rodarte, A.I.; Siddiqi, S.; Acevedo-Moreno, L.-A.; Brown, J.M.; Allende, D.S.; Aucejo, F.; Rotroff, D.M. Salivary miRNAs as non-invasive biomarkers of hepatocellular carcinoma: A pilot study. PeerJ 2022, 10, e12715. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Wong, Y.S.; Goh, B.K.P.; Chan, C.Y.; Cheow, P.C.; Chow, P.K.H.; Lim, T.K.H.; Goh, G.B.B.; Krishnamoorthy, T.L.; Kumar, R.; et al. Circulating microRNAs as Potential Diagnostic and Prognostic Biomarkers in Hepatocellular Carcinoma. Sci. Rep. 2019, 9, 10464. [Google Scholar] [CrossRef]
- Dat, V.H.X.; Nhung, B.T.H.; Chau, N.N.B.; Cuong, P.H.; Hieu, V.D.; Linh, N.T.M.; Quoc, N.B. Identification of potential microRNA groups for the diagnosis of hepatocellular carcinoma (HCC) using microarray datasets and bioinformatics tools. Heliyon 2022, 8, e08987. [Google Scholar] [CrossRef]
- Zhang, L.; Lou, J.; Lu, M.; Gao, C.; Zhao, S.; Li, B.; Liang, S.; Li, Y.; Li, D.; Liu, M. Suppression of miR-199a maturation by HuR is crucial for hypoxia-induced glycolytic switch in hepatocellular carcinoma. EMBO J. 2015, 34, 2671–2685. [Google Scholar] [CrossRef] [PubMed]
- Dabbish, A.M.; Abdelzaher, H.M.; Abohawya, M.; Shamma, S.; Mahmoud, Y.H.; Maged, A.; Manaa, M.; Hassany, M.; Kobeissy, F.; Bazgir, O.; et al. Prognostic MicroRNA Panel for HCV-Associated HCC: Integrating Computational Biology and Clinical Validation. Cancers 2022, 14, 3036. [Google Scholar] [CrossRef]
- Di Bella, M.A. Overview and Update on Extracellular Vesicles: Considerations on Exosomes and Their Application in Modern Medicine. Biology 2022, 11, 804. [Google Scholar] [CrossRef] [PubMed]
- Bartel, S.; Deshane, J.; Wilkinson, T.; Gabrielsson, S. Extracellular Vesicles as Mediators of Cellular Cross Talk in the Lung Microenvironment. Front. Med. 2020, 7, 326. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, A.; Natsui, K.; Ishii, Y.; Koseki, Y.; Takeda, N.; Tomiyoshi, K.; Yamazaki, F.; Yoshida, Y.; Terai, S. Small extracellular vesicles and liver diseases: From diagnosis to therapy. World J. Hepatol. 2022, 14, 1307–1318. [Google Scholar] [CrossRef]
- Liu, C.; Wu, H.; Mao, Y.; Chen, W.; Chen, S. Exosomal microRNAs in hepatocellular carcinoma. Cancer Cell Int. 2021, 21, 254. [Google Scholar] [CrossRef]
- Li, S.; Yao, J.; Xie, M.; Liu, Y.; Zheng, M. Exosomal miRNAs in hepatocellular carcinoma development and clinical responses. J. Hematol. Oncol. 2018, 11, 54. [Google Scholar] [CrossRef]
- Sorop, A.; Constantinescu, D.; Cojocaru, F.; Dinischiotu, A.; Cucu, D.; Dima, S. Exosomal microRNAs as Biomarkers and Therapeutic Targets for Hepatocellular Carcinoma. Int. J. Mol. Sci. 2021, 22, 4997. [Google Scholar] [CrossRef]
- Wang, H.; Yu, L.; Huang, P.; Zhou, Y.; Zheng, W.; Meng, N.; He, R.; Xu, Y.; Keong, T.S.; Cui, Y. Tumor-associated Exosomes Are Involved in Hepatocellular Carcinoma Tumorigenesis, Diagnosis, and Treatment. J. Clin. Transl. Hepatol. 2022, 10, 496–508. [Google Scholar] [CrossRef]
- Ghosh, S.; Bhowmik, S.; Majumdar, S.; Goswami, A.; Chakraborty, J.; Gupta, S.; Aggarwal, S.; Ray, S.; Chatterjee, R.; Bhattacharyya, S.; et al. The exosome encapsulated microRNAs as circulating diagnostic marker for hepatocellular carcinoma with low alpha-fetoprotein. Int. J. Cancer 2020, 147, 2934–2947. [Google Scholar] [CrossRef]
- Wang, H.; Hou, L.; Li, A.; Duan, Y.; Gao, H.; Song, X. Expression of Serum Exosomal MicroRNA-21 in Human Hepatocellular Carcinoma. BioMed Res. Int. 2014, 2014, 864894. [Google Scholar] [CrossRef] [PubMed]
- Sorop, A.; Iacob, R.; Iacob, S.; Constantinescu, D.; Chitoiu, L.; Fertig, T.E.; Dinischiotu, A.; Chivu-Economescu, M.; Bacalbasa, N.; Savu, L.; et al. Plasma Small Extracellular Vesicles Derived miR-21-5p and miR-92a-3p as Potential Biomarkers for Hepatocellular Carcinoma Screening. Front. Genet. 2020, 11, 712. [Google Scholar] [CrossRef]
- Nakano, T.; Chen, I.-H.; Wang, C.-C.; Chen, P.-J.; Tseng, H.-P.; Huang, K.-T.; Hu, T.-H.; Li, L.-C.; Goto, S.; Cheng, Y.-F.; et al. Circulating exosomal miR-92b: Its role for cancer immunoediting and clinical value for prediction of posttransplant hepatocellular carcinoma recurrence. Am. J. Transplant. 2019, 19, 3250–3262. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Xu, M.; Zhang, G.; Dong, L.; Wu, J.; Wei, C.; Xu, K.; Zhang, L. Identification of Circulating Exosomal miR-101 and miR-125b Panel Act as a Potential Biomarker for Hepatocellular Carcinoma. Int. J. Genom. 2021, 2021, 1326463. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Hu, J.; Zhou, K.; Chen, F.; Wang, Z.; Liao, B.; Dai, Z.; Cao, Y.; Fan, J.; Zhou, J. Serum exosomal miR-125b is a novel prognostic marker for hepatocellular carcinoma. OncoTargets Ther. 2017, 10, 3843–3851. [Google Scholar] [CrossRef]
- Abdelaziz, H.E.; El-Guendy, N.; Radwan, E.M.; Sharawi, S.K.; E El-Houseini, M.; Abdel-Wahab, A.-H.A. Possible Use of miR-223-3p as a Prognostic Marker in Transarterial Chemoembolization Treatment of Hepatocellular Carcinoma Patients. Asian Pac. J. Cancer Prev. 2022, 23, 4125–4135. [Google Scholar] [CrossRef]
- Sohn, W.; Kim, J.; Kang, S.H.; Yang, S.R.; Cho, J.-Y.; Cho, H.C.; Shim, S.G.; Paik, Y.-H. Serum exosomal microRNAs as novel biomarkers for hepatocellular carcinoma. Exp. Mol. Med. 2015, 47, e184. [Google Scholar] [CrossRef] [PubMed]
- Fründt, T.; Krause, L.; Hussey, E.; Steinbach, B.; Köhler, D.; von Felden, J.; Schulze, K.; Lohse, A.; Wege, H.; Schwarzenbach, H. Diagnostic and Prognostic Value of miR-16, miR-146a, miR-192 and miR-221 in Exosomes of Hepatocellular Carcinoma and Liver Cirrhosis Patients. Cancers 2021, 13, 2484. [Google Scholar] [CrossRef] [PubMed]
- Deng, P.; Li, M.; Wu, Y. The Predictive Efficacy of Serum Exosomal microRNA-122 and microRNA-148a for Hepatocellular Carcinoma Based on Smart Healthcare. J. Healthc. Eng. 2022, 2022, 5914541. [Google Scholar] [CrossRef]
- Suehiro, T.; Miyaaki, H.; Kanda, Y.; Shibata, H.; Honda, T.; Ozawa, E.; Miuma, S.; Taura, N.; Nakao, K. Serum exosomal microRNA-122 and microRNA-21 as predictive biomarkers in transarterial chemoembolization-treated hepatocellular carcinoma patients. Oncol. Lett. 2018, 16, 3267–3273. [Google Scholar] [CrossRef]
- Ma, Y.-S.; Liu, J.-B.; Lin, L.; Zhang, H.; Wu, J.-J.; Shi, Y.; Jia, C.-Y.; Zhang, D.-D.; Yu, F.; Wang, H.-M.; et al. Exosomal microRNA-15a from mesenchymal stem cells impedes hepatocellular carcinoma progression via downregulation of SALL4. Cell Death Discov. 2021, 7, 224. [Google Scholar] [CrossRef]
- Zhou, Y.; Ren, H.; Dai, B.; Li, J.; Shang, L.; Huang, J.; Shi, X. Hepatocellular carcinoma-derived exosomal miRNA-21 contributes to tumor progression by converting hepatocyte stellate cells to cancer-associated fibroblasts. J. Exp. Clin. Cancer Res. 2018, 37, 324. [Google Scholar] [CrossRef]
- Liu, J.; Fan, L.; Yu, H.; Zhang, J.; He, Y.; Feng, D.; Wang, F.; Li, X.; Liu, Q.; Li, Y.; et al. Endoplasmic Reticulum Stress Causes Liver Cancer Cells to Release Exosomal miR-23a-3p and Up-regulate Programmed Death Ligand 1 Expression in Macrophages. Hepatology 2019, 70, 241–258. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Zou, B.; Nan, T.; Zheng, X.; Zheng, L.; Lan, J.; Chen, W.; Yu, J. MiR-25 enhances autophagy and promotes sorafenib resistance of hepatocellular carcinoma via targeting FBXW7. Int. J. Med. Sci. 2022, 19, 257–266. [Google Scholar] [CrossRef]
- Koustas, E.; Sarantis, P.; Papavassiliou, A.G.; Karamouzis, M.V. Upgraded role of autophagy in colorectal carcinomas. World J. Gastrointest. Oncol. 2018, 10, 367–369. [Google Scholar] [CrossRef]
- Fang, J.H.; Zhang, Z.J.; Shang, L.R.; Luo, Y.W.; Lin, Y.F.; Yuan, Y.; Zhuang, S.M. Hepatoma cell-secreted exosomal microRNA-103 increases vascular permeability and promotes metastasis by targeting junction proteins. Hepatology 2018, 68, 1459–1475. [Google Scholar] [CrossRef] [PubMed]
- Lou, G.; Song, X.; Yang, F.; Wu, S.; Wang, J.; Chen, Z.; Liu, Y. Exosomes derived from miR-122-modified adipose tissue-derived MSCs increase chemosensitivity of hepatocellular carcinoma. J. Hematol. Oncol. 2015, 8, 122. [Google Scholar] [CrossRef]
- Li, S.; Chen, L. Exosomes in Pathogenesis, Diagnosis, and Treatment of Hepatocellular Carcinoma. Front. Oncol. 2022, 12, 793432. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Jiang, J.; Badawi, M.; Schmittgen, T.D. miR-221 regulates CD44 in hepatocellular carcinoma through the PI3K-AKT-mTOR pathway. Biochem. Biophys. Res. Commun. 2017, 487, 709–715. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, X.; Sun, W.; Yue, S.; Yang, J.; Li, J.; Ma, B.; Wang, J.; Yang, X.; Pu, M.; et al. Loss of exosomal miR-320a from cancer-associated fibroblasts contributes to HCC proliferation and metastasis. Cancer Lett. 2017, 397, 33–42. [Google Scholar] [CrossRef]
- Sun, W.; Fu, S.; Wu, S.; Tu, R. Growing Evidence of Exosomal MicroRNA-Related Metastasis of Hepatocellular Carcinoma. BioMed Res. Int. 2020, 2020, 4501454. [Google Scholar] [CrossRef] [PubMed]
- Fang, T.; Lv, H.; Lv, G.; Li, T.; Wang, C.; Han, Q.; Yu, L.; Su, B.; Guo, L.; Huang, S.; et al. Tumor-derived exosomal miR-1247-3p induces cancer-associated fibroblast activation to foster lung metastasis of liver cancer. Nat. Commun. 2018, 9, 191. [Google Scholar] [CrossRef]
- Dong, S.-S.; Dong, D.-D.; Yang, Z.-F.; Zhu, G.-Q.; Gao, D.-M.; Chen, J.; Zhao, Y.; Liu, B.-B. Exosomal miR-3682-3p Suppresses Angiogenesis by Targeting ANGPT1 via the RAS-MEK1/2-ERK1/2 Pathway in Hepatocellular Carcinoma. Front. Cell Dev. Biol. 2021, 9, 633358. [Google Scholar] [CrossRef]
- Majeed, S.T.; Majeed, R.; Andrabi, K.I. Expanding the view of the molecular mechanisms of autophagy pathway. J. Cell. Physiol. 2022, 237, 3257–3277. [Google Scholar] [CrossRef]
- Koustas, E.; Sarantis, P.; Karamouzis, M.; Vielh, P.; Theocharis, S. The Controversial Role of Autophagy in Ewing Sarcoma Pathogenesis—Current Treatment Options. Biomolecules 2021, 11, 355. [Google Scholar] [CrossRef] [PubMed]
- Koustas, E.; Trifylli, E.-M.; Sarantis, P.; Papadopoulos, N.; Papanikolopoulos, K.; Aloizos, G.; Damaskos, C.; Garmpis, N.; Garmpi, A.; Matthaios, D.; et al. Exploiting Autophagy-Dependent Neoantigen Presentation in Tumor Microenvironment. Genes 2023, 14, 474. [Google Scholar] [CrossRef]
- Shan, C.; Chen, X.; Cai, H.; Hao, X.; Li, J.; Zhang, Y.; Gao, J.; Zhou, Z.; Li, X.; Liu, C.; et al. The Emerging Roles of Autophagy-Related MicroRNAs in Cancer. Int. J. Biol. Sci. 2021, 17, 134–150. [Google Scholar] [CrossRef] [PubMed]
- Akkoc, Y.; Gozuacik, D. MicroRNAs as major regulators of the autophagy pathway. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2020, 1867, 118662. [Google Scholar] [CrossRef]
- Wang, P.; Liu, X.-M.; Ding, L.; Zhang, X.-J.; Ma, Z.-L. mTOR signaling-related MicroRNAs and Cancer involvement. J. Cancer 2018, 9, 667–673. [Google Scholar] [CrossRef]
- Lu, G.; Wang, Y.; Shi, Y.; Zhang, Z.; Huang, C.; He, W.; Wang, C.; Shen, H. Autophagy in health and disease: From molecular mechanisms to therapeutic target. Medcomm 2022, 3, e150. [Google Scholar] [CrossRef]
- Yang, Y.; Liang, C.; Isidoro, C. MicroRNAs: An Emerging Player In Autophagy. Sci. Res. 2015, 10, 14293. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, Z.; Lu, Y.; Song, K.; Liu, X.; Xia, F.; Sun, W. Downregulation of ULK 1 by micro RNA -372 inhibits the survival of human pancreatic adenocarcinoma cells. Cancer Sci. 2017, 108, 1811–1819. [Google Scholar] [CrossRef]
- Roy, S.G. Regulation of autophagy by miRNAs in human diseases. Nucleus 2021, 64, 317–329. [Google Scholar] [CrossRef]
- Zhu, H.; Wu, H.; Liu, X.; Li, B.; Chen, Y.; Ren, X.; Liu, C.-G.; Yang, J.-M. Regulation of autophagy by a beclin 1-targeted microRNA, miR-30a, in cancer cells. Autophagy 2009, 5, 816–823. [Google Scholar] [CrossRef] [PubMed]
- Gozuacik, D.; Akkoc, Y.; Ozturk, D.G.; Kocak, M. Autophagy-Regulating microRNAs and Cancer. Front. Oncol. 2017, 7, 65. [Google Scholar] [CrossRef] [PubMed]
- Hyttinen, J.M.; Blasiak, J.; Felszeghy, S.; Kaarniranta, K. MicroRNAs in the regulation of autophagy and their possible use in age-related macular degeneration therapy. Ageing Res. Rev. 2021, 67, 101260. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, Z.; Zhang, W.; Zhang, L. MicroRNAs play an essential role in autophagy regulation in various disease phenotypes. Biofactors 2019, 45, 844–856. [Google Scholar] [CrossRef]
- Xu, X.; Tao, Y.; Shan, L.; Chen, R.; Jiang, H.; Qian, Z.; Cai, F.; Ma, L.; Yu, Y. The Role of MicroRNAs in Hepatocellular Carcinoma. J. Cancer 2018, 9, 3557–3569. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Q.; Song, J. Inhibition of autophagy potentiates the proliferation inhibition activity of microRNA-7 in human hepatocellular carcinoma cells. Oncol. Lett. 2017, 14, 3566–3572. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Li, C.; Hao, R. miR-559 Inhibits Proliferation, Autophagy, and Angiogenesis of Hepatocellular Carcinoma Cells by Targeting PARD3. Mediat. Inflamm. 2022, 2022, 3121492. [Google Scholar] [CrossRef]
- Xu, Y.; An, Y.; Wang, Y.; Zhang, C.; Zhang, H.; Huang, C.; Jiang, H.; Wang, X.; Li, X. miR-101 inhibits autophagy and enhances cisplatin-induced apoptosis in hepatocellular carcinoma cells. Oncol. Rep. 2013, 29, 2019–2024. [Google Scholar] [CrossRef]
- Zhang, K.; Chen, J.; Zhou, H.; Chen, Y.; Zhi, Y.; Zhang, B.; Chen, L.; Chu, X.; Wang, R.; Zhang, C. PU.1/microRNA-142-3p targets ATG5/ATG16L1 to inactivate autophagy and sensitize hepatocellular carcinoma cells to sorafenib. Cell Death Dis. 2018, 9, 312. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; He, Y.; Zhai, N.; Ding, S.; Li, J.; Peng, Z.Z. MicroRNA-181a inhibits autophagy by targeting Atg5 in hepatocellular carcinoma. Front. Biosci. 2018, 23, 388–396. [Google Scholar]
- Zhang, Y.-J.; Pan, Q.; Yu, Y.; Zhong, X.-P. microRNA-519d Induces Autophagy and Apoptosis of Human Hepatocellular Carcinoma Cells through Activation of the AMPK Signaling Pathway via Rab10. Cancer Manag. Res. 2020, 12, 2589–2602. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Yang, L.; Li, X.; Li, B.; Li, Y.; Zhang, X.; Ma, Y.; Peng, X.; Jin, H.; Li, H. New Insights into Autophagy in Hepatocellular Carcinoma: Mechanisms and Therapeutic Strategies. Am. J. Cancer Res. 2019, 9, 1329–1353. [Google Scholar]
- Yang, S.; Wang, M.; Yang, L.; Li, Y.; Ma, Y.; Peng, X.; Li, X.; Li, B.; Jin, H.; Li, H. MicroRNA-375 Targets ATG14 to Inhibit Autophagy and Sensitize Hepatocellular Carcinoma Cells to Sorafenib. OncoTargets Ther. 2020, 13, 3557–3570. [Google Scholar] [CrossRef] [PubMed]
- Kaur, R.; Kanthaje, S.; Taneja, S.; Dhiman, R.K.; Chakraborti, A. miR-23b-3p Modulating Cytoprotective Autophagy and Glutamine Addiction in Sorafenib Resistant HepG2, a Hepatocellular Carcinoma Cell Line. Genes 2022, 13, 1375. [Google Scholar] [CrossRef]
- Xie, C.-M.; Sun, Y. The MTORC1-mediated autophagy is regulated by the FBXW7-SHOC2-RPTOR axis. Autophagy 2019, 15, 1470–1472. [Google Scholar] [CrossRef]
- Stiuso, P.; Potenza, N.; Lombardi, A.; Ferrandino, I.; Monaco, A.; Zappavigna, S.; Vanacore, D.; Mosca, N.; Castiello, F.; Porto, S.; et al. MicroRNA-423-5p Promotes Autophagy in Cancer Cells and Is Increased in Serum from Hepatocarcinoma Patients Treated with Sorafenib. Mol. Ther. Nucleic Acids 2015, 4, e233. [Google Scholar] [CrossRef]
- Jin, W.; Liang, Y.; Li, S.; Lin, G.; Liang, H.; Zhang, Z.; Zhang, W.; Nie, R. MiR-513b-5p represses autophagy during the malignant progression of hepatocellular carcinoma by targeting PIK3R3. Aging 2021, 13, 16072–16087. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Yan, W.; He, X.; Zhang, L.; Li, C.; Huang, H.; Nace, G.; Geller, D.A.; Lin, J.; Tsung, A. miR-375 Inhibits Autophagy and Reduces Viability of Hepatocellular Carcinoma Cells under Hypoxic Conditions. Gastroenterology 2012, 143, 177–187.e8. [Google Scholar] [CrossRef] [PubMed]
- Lan, S.-H.; Wu, S.-Y.; Zuchini, R.; Lin, X.-Z.; Su, I.-J.; Tsai, T.-F.; Lin, Y.-J.; Wu, C.-T.; Liu, H.-S. Autophagy-preferential degradation of MIR224 participates in hepatocellular carcinoma tumorigenesis. Autophagy 2014, 10, 1687–1689. [Google Scholar] [CrossRef]
- Tripathi, A.; Debelius, J.; Brenner, D.A.; Karin, M.; Loomba, R.; Schnabl, B.; Knight, R. The gut–liver axis and the intersection with the microbiome. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 397–411. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D. Human gut microbiome: Hopes, threats and promises. Gut 2018, 67, 1716–1725. [Google Scholar] [CrossRef] [PubMed]
- Kiriyama, Y.; Nochi, H. Physiological Role of Bile Acids Modified by the Gut Microbiome. Microorganisms 2021, 10, 68. [Google Scholar] [CrossRef]
- Trifylli, E.-M.; Koustas, E.; Papadopoulos, N.; Sarantis, P.; Aloizos, G.; Damaskos, C.; Garmpis, N.; Garmpi, A.; Karamouzis, M.V. An Insight into the Novel Immunotherapy and Targeted Therapeutic Strategies for Hepatocellular Carcinoma and Cholangiocarcinoma. Life 2022, 12, 665. [Google Scholar] [CrossRef]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What Is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef] [PubMed]
- Rattan, P.; Minacapelli, C.D.; Rustgi, V. The Microbiome and Hepatocellular Carcinoma. Liver Transplant. 2020, 26, 1316–1327. [Google Scholar] [CrossRef] [PubMed]
- Allegra, A.; Musolino, C.; Tonacci, A.; Pioggia, G.; Gangemi, S. Interactions between the MicroRNAs and Microbiota in Cancer Development: Roles and Therapeutic Opportunities. Cancers 2020, 12, 805. [Google Scholar] [CrossRef]
- Tian, Y.; Zhang, M.; Fan, M.; Xu, H.; Wu, S.; Zou, S.; Wang, Y.; Tang, D.; Zhang, C.; Han, W.; et al. A miRNA-mediated attenuation of hepatocarcinogenesis in both hepatocytes and Kupffer cells. Mol. Ther. Nucleic Acids 2022, 30, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhang, W.; Jiang, L.; Chen, Y. Recent advances in systemic therapy for hepatocellular carcinoma. Biomark. Res. 2022, 10, 3. [Google Scholar] [CrossRef]
- Pratama, M.Y.; Pascut, D.; Massi, M.N.; Tiribelli, C. The role of microRNA in the resistance to treatment of hepatocellular carcinoma. Ann. Transl. Med. 2019, 7, 577. [Google Scholar] [CrossRef]
- Guo, W.; Qiu, Z.; Wang, Z.; Wang, Q.; Tan, N.; Chen, T.; Chen, Z.; Huang, S.; Gu, J.; Li, J.; et al. MiR-199a-5p is negatively associated with malignancies and regulates glycolysis and lactate production by targeting hexokinase 2 in liver cancer. Hepatology 2015, 62, 1132–1144. [Google Scholar] [CrossRef]
- Bozzato, A.M.; Martingano, P.; Mucelli, R.A.P.; Cavallaro, M.F.M.; Cesarotto, M.; Marcello, C.; Tiribelli, C.; Pascut, D.; Pizzolato, R.; Mucelli, F.P.; et al. MicroRNAs Related to TACE Treatment Response: A Review of the Literature from a Radiological Point of View. Diagnostics 2022, 12, 374. [Google Scholar] [CrossRef]
- Hassan, M.; Elzallat, M.; Aboushousha, T.; Elhusseny, Y.; El-Ahwany, E. MicroRNA-122 mimic/microRNA-221 inhibitor combination as a novel therapeutic tool against hepatocellular carcinoma. Noncoding RNA Res. 2023, 8, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Hong, D.S.; Kang, Y.K.; Borad, M.; Sachdev, J.; Ejadi, S.; Lim, H.Y.; Brenner, A.J.; Park, K.; Lee, J.L.; Kim, T.Y.; et al. Phase 1 study of MRX34, a liposomal miR-34a mimic, in patients with advanced solid tumours. Br. J. Cancer 2020, 122, 1630–1637. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Liao, Y.; Tang, L. MicroRNA-34 family: A potential tumor suppressor and therapeutic candidate in cancer. J. Exp. Clin. Cancer Res. 2019, 38, 53. [Google Scholar] [CrossRef]
- Sousa, D.P.; Conde, J. Gold Nanoconjugates for MiRNA Modulation in Cancer Therapy: From MiRNA Silencing to MiRNA Mimics. ACS Mater. Au 2022, 2, 626–640. [Google Scholar] [CrossRef] [PubMed]
- Mo, Y.; He, L.; Lai, Z.; Wan, Z.; Chen, Q.; Pan, S.; Li, L.; Li, D.; Huang, J.; Xue, F.; et al. Gold Nano-Particles (AuNPs) Carrying MiR-326 Targets PDK1/AKT/c-Myc Axis in Hepatocellular Carcinoma. Artif. Cells Nanomed. Biotechnol. 2019, 47, 2830–2837. [Google Scholar] [CrossRef]
- Huang, S.-T.; Hsu, W.-F.; Huang, H.-S.; Yen, J.-H.; Lin, M.-C.; Peng, C.-Y.; Yen, H.-R. Improved Survival in Hepatocellular Carcinoma Patients with Cardiac Arrhythmia by Amiodarone Treatment through Autophagy. Int. J. Mol. Sci. 2019, 20, 3978. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| OncomiRs | Expression Levels in HCC | Effects | Target |
|---|---|---|---|
| miR-18a | ↑ | Enhances HCC cell proliferation and tumor migratory behavior | Bcl2l10 [34] |
| miR-21 | ↑ | Tumor proliferation, migration, and EMT | PI3K/AKT, STAT3, TGF-β/SMADs HEPN1 [32,33] |
| Involvement in cell cycle | |||
| miR-92a | ↑ | Tumor proliferation/invasion | FOXA2 [29] |
| miR-155-5p | ↑ | HCC progression | PTEN [35] |
| miR-203a-3p.1 | ↑ | Metastatic dissemination | IL24 [30] |
| miR-302d | ↑ | Tumor proliferation | TGFBR2 [38] |
| miR-331-3p | ↑ | HCC progression | E2F1 [36] |
| miR-346 | ↑ | Tumor growth and progression | BRMS1 [41] |
| miR-454 | ↑ | Recurrent HCC, metastatic dissemination resistance to therapeutic modalities | TICs [40] |
| miR-487a | ↑ | Tumor migration, metastatic dissemination | SPRED2 PIK3R1 [39] |
| miR-765 | ↑ | Tumor proliferation | INPP4B [37] |
| miR-873 | ↑ | HCC growth and metastatic dissemination | AKT/mTOR [43] |
| miR-3910 | ↑ | Tumor progression and migration | MST1 [42] |
| Hippo-YAP pathway | |||
| miR-4417 | ↑ | Inhibits apoptosis and tumor proliferation | PKM2, TRIM35 [31] |
| Tumor-Suppressing miRNA | Expression Levels in HCC | Effect | Target |
|---|---|---|---|
| miR-29a | ↓ | Suppresses invasion | VEGFA, LOXL2 LOX [52,53] |
| and metastasis | |||
| miR-30e-3p | ↓ | Tumor suppression | p53 [58,59] |
| Activates p53 | |||
| miR-122 | ↓ or | Regulate lipid metabolism | mdm2, p53 [61,62,63] |
| knockdown | Tumor suppression | ||
| miR-148b | ↓ | Tumor growth suppression | WNT1 [55] |
| miR-193a-5p | ↓ | Inhibits HCC formation and progression | Nucleolar- and |
| spindle-associated protein [56,57] | |||
| miR-195 | ↓ | Suppresses HCC development via interacting the malignant hepatocyte cell cycle. | CDC25A, CDK6, CDK4, CCNE1 [48] |
| miR-199a | ↓ | Impairment of tumor growth, proliferation | Aerobic |
| Glycolysis [45] | |||
| miR-199a-5p | ↓ | Glycometabolism in hepatocytes | 3′ UTR HK2 mRNA [44] |
| Reprograms the glycolysis of the malignant cells | |||
| miR-199a/b-3p | ↓ | Tumor inhibition | MAPK/ERK PAK4 [46] |
| miR-206 | ↓ | Inhibits HCC development in mice | CDK6, CCND1, cMET [51] |
| miR-296-5p | ↓ | Suppresses the HCC stem cell lines/EMT phenomenon | Neuregulin-ERBB pathway [49,50] |
| miR-766-3p | ↓ | Tumor suppression | Wnt/β-catenin signaling pathway [54] |
| miR-1249 | ↑ | Tumor suppression | Hedgehog pathway |
| PTCH1 [60] |
| Utilization | MiRNAs | Expression Levels | Isolation | References |
|---|---|---|---|---|
| Diagnostic markers | mir-92b | Decreased in saliva/ increased in tumor tissue | Saliva and Tumor tissue | [112] |
| mir-548i-2 | Decreased in saliva/ decreased in tumor tissue | |||
| mir-548l | Decreased in saliva/ decreased in tumor tissue | |||
| Diagnostic markers | miR-1972 | Increased only in HCC | Plasma | [113] |
| miR-193a-5p | Increased only in HCC | |||
| miR-214-3p | Increased only in HCC | |||
| miR-365a-3p | Increased only in HCC | |||
| Poor prognostic markers | miR-137 | Decreased | Tumor tissue | [99] |
| miR-296 | Decreased | Tumor tissue | [100] | |
| miR-638 | Decreased | Serum | [101] | |
| miR-940 | Decreased | Tumor tissue | [98] | |
| miR-32-5p | Increased | Tumor tissue | [95] | |
| miR-92a | Increased | Tumor tissue | [106] | |
| miR-221 | Increased | Tumor tissue | [97] | |
| miR-1246 | Increased | Serum | [105] | |
| Good prognostic markers | miR-296-5p | Notably decreased | Tumor tissue | [49,50] |
| Monitoring markers | miR-199a-5p | [18F]-FDG PET-CT performance | [115] | |
| Follow up for reoccurrence | miR-148 | Serum | [108] | |
| Screening markers | miR-148a in ↓ or absent AFP | Increased | Serum | [108] |
| Staging | Pre-metastatic | Increased | Serum | [102] |
| miR-638 | ||||
| miR-663 | ||||
| miR-3648 | ||||
| miR-4258 | ||||
| Circulatory miRNAs HCV and HCC | miR-142 miR-150 | Decreased Decreased | Blood | [116] |
| miR-183 | Increased | |||
| miR-199b | Decreased | |||
| miR-215 | Decreased | |||
| miR-217 | Increased | |||
| miR-224 | Increased | |||
| miR-424 | Decreased | |||
| miR-3607 | Decreased | |||
| Bioptic specimens | miR-142 | Tumor tissue | [116] | |
| miR-183 | ||||
| miR-217 | ||||
| miR-3607 |
| Exosomal miRNA | Isolation | Expression Levels | Utilization |
|---|---|---|---|
| miR-21 | Serum | ↑ in HCC patients | Diagnostic tool [125] |
| miR-21-5p | Plasma | ↑ in HCC patients | Diagnostic tool and monitoring together with serum AFP [126] |
| miR-92b | Serum | ↑levels—HCC recurrence after liver transplantation | Predictive biomarker [127] |
| miR-92-3p | Plasma | ↑ in HCC patients/AFP low levels | Diagnostic tool and monitoring [126] |
| miR-101 | Serum | ↓ in HCC patients | Diagnostic tool [128] |
| miR-125b | Serum | ↓ in HCC patients ↓levels—↓ recurrence time ↓overall survival | Diagnostic [128] and prognostic tool [129] |
| miR-223-3p | Serum | ↓↓ levels—non-responders to TACE treatment | Predictive biomarker [130] |
| miR-665 | Serum | ↑ levels—HCC patients ↑↑levels—poor prognosis (stage, tumor size, survival) | Diagnostic tool and prognostic biomarker [121] |
| miR-718 | Serum | ↓levels—HCC recurrence after liver transplantation | Predictive biomarker [121] |
| miR-18a, miR-221, miR-222, miR-224 | Serum | ↑↑levels—HCC patients | Diagnostic tool [131] |
| miR-16, miR-146, miR-192 | Plasma | ↑levels—HCC and cirrhosis | Diagnostic tool and prognostic biomarker [132] |
| miR-122, miR-148a | Serum | Predictive biomarkers [133] | |
| miR-122 | Serum | ↓↓ levels post-TACE in Cirrhotic-HCC patients | Predictive biomarkers [134] |
| EV-Contained miRNA Cargo | Functional Role in HCC |
|---|---|
| EVs-miR let-7b [126] | ↓tumor inflammation by targeting |
| interleukin 6 | |
| EVs-miR 15a [135] | Inhibits tumor proliferation/migration |
| EVs-miR-21 [136] | PDK1/AKT pathway activation |
| Conversion of hepatic stellate cells to CAFs | |
| Tumor migration | |
| Neoangiogenesis | |
| EVs-miR-23a-3p [137] | Uptake by macrophages |
| Promoted tumor escape | |
| phenomenon | |
| T-cell function suppression | |
| EVs-miR-25 [121,138] | Modification of BAX, BCL2 expression |
| (apoptotic markers for sorafenib) | |
| Drug resistance in sorafenib | |
| EVs-miR-32-5p [139] | Multi-drug resistance |
| EVs-miR-103 [140] | Uptake by endothelial cells |
| Modified integrity of vessels | |
| Tumor invasion | |
| EVs-miR-122 [141] | Suppressed tumor proliferation |
| ↑chemosensitivity | |
| EVs-miR-125a [142] | Targets HCC stem cells CD90 |
| Suppressed tumor proliferation and | |
| Migration | |
| EVs-miR-221 [143] | CD44 regulation via interaction with |
| PI3K-AKT-mTOR | |
| Tumor proliferation/migration | |
| EVs-miR-320a [123,144] | Inhibition of MAPK pathway |
| Suppressed tumor migration and invasion | |
| EVs-miR-429 [145] | ↑E2F1 transcription activity |
| Tumor proliferation | |
| EVs-miR-1247-3p [146] | Conversion of fibroblasts to CAFs |
| Promoted EMT phenomenon | |
| Metastatic dissemination and proliferation | |
| EVs-miR 3682 3p [147] | Inhibited neoangiogenesis |
| AutophagomiRs | Target | Role in HCC |
|---|---|---|
| miR-7 [163] | ATG5 | Inhibits invasion and metastasis |
| miR-25 [138] | FBXW7 | Autophagy enhancement Sorafenib resistance |
| miR-23b-3p [171] | ATG12 | ↓survival and ↑apoptosis of sorafenib-resistant HepG2 cells |
| miR-101 [165] | ATG4D RAB5A, STMN1 | Inhibits autophagy Enhances cisplatin-induced apoptosis |
| miR-142-3p [166] | ATG5/ATG16L1 | Autophagy inhibition Sensitization of HCC cells to sorafenib |
| miR-181a [167] | ATG5 | autophagy inhibition ↑apoptosis in HepG2 cells ↓tumor growth (HepG2 cell xenograft tumor models) |
| miR-375 [170] | ATG7 | Inhibits autophagy ↓HCC survival under hypoxia |
| miR-423-5p [173] | Autophagy cell cycle | ↑autophagy in HCC cells-treated with sorafenib Partially remitted/stable disease 6 months after the treatment initiation (predictive biomarker) |
| miR-513b-5p [174] | PIK3R3 | Inhibits proliferation, migration/invasion, Induces HCC cell apoptosis |
| miR-519d [168] | Rab10 | Autophagy induction ↑HCC cell Apoptosis AMPK signaling pathway activation via Rab10 |
| miR-559 [164] | PARD3 | Inhibits proliferation, neoangiogenesis, inhibits autophagy |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koustas, E.; Trifylli, E.-M.; Sarantis, P.; Papadopoulos, N.; Papanikolopoulos, K.; Aloizos, G.; Damaskos, C.; Garmpis, N.; Garmpi, A.; Matthaios, D.; et al. An Insight into the Arising Role of MicroRNAs in Hepatocellular Carcinoma: Future Diagnostic and Therapeutic Approaches. Int. J. Mol. Sci. 2023, 24, 7168. https://doi.org/10.3390/ijms24087168
Koustas E, Trifylli E-M, Sarantis P, Papadopoulos N, Papanikolopoulos K, Aloizos G, Damaskos C, Garmpis N, Garmpi A, Matthaios D, et al. An Insight into the Arising Role of MicroRNAs in Hepatocellular Carcinoma: Future Diagnostic and Therapeutic Approaches. International Journal of Molecular Sciences. 2023; 24(8):7168. https://doi.org/10.3390/ijms24087168
Chicago/Turabian StyleKoustas, Evangelos, Eleni-Myrto Trifylli, Panagiotis Sarantis, Nikolaos Papadopoulos, Konstantinos Papanikolopoulos, Georgios Aloizos, Christos Damaskos, Nikolaos Garmpis, Anna Garmpi, Dimitris Matthaios, and et al. 2023. "An Insight into the Arising Role of MicroRNAs in Hepatocellular Carcinoma: Future Diagnostic and Therapeutic Approaches" International Journal of Molecular Sciences 24, no. 8: 7168. https://doi.org/10.3390/ijms24087168
APA StyleKoustas, E., Trifylli, E.-M., Sarantis, P., Papadopoulos, N., Papanikolopoulos, K., Aloizos, G., Damaskos, C., Garmpis, N., Garmpi, A., Matthaios, D., & Karamouzis, M. V. (2023). An Insight into the Arising Role of MicroRNAs in Hepatocellular Carcinoma: Future Diagnostic and Therapeutic Approaches. International Journal of Molecular Sciences, 24(8), 7168. https://doi.org/10.3390/ijms24087168