
Analysis of Codon Usage Bias in Xyloglucan Endotransglycosylase (XET) Genes

 , , ,
, , ,
Abstract
1. Introduction
2. Results
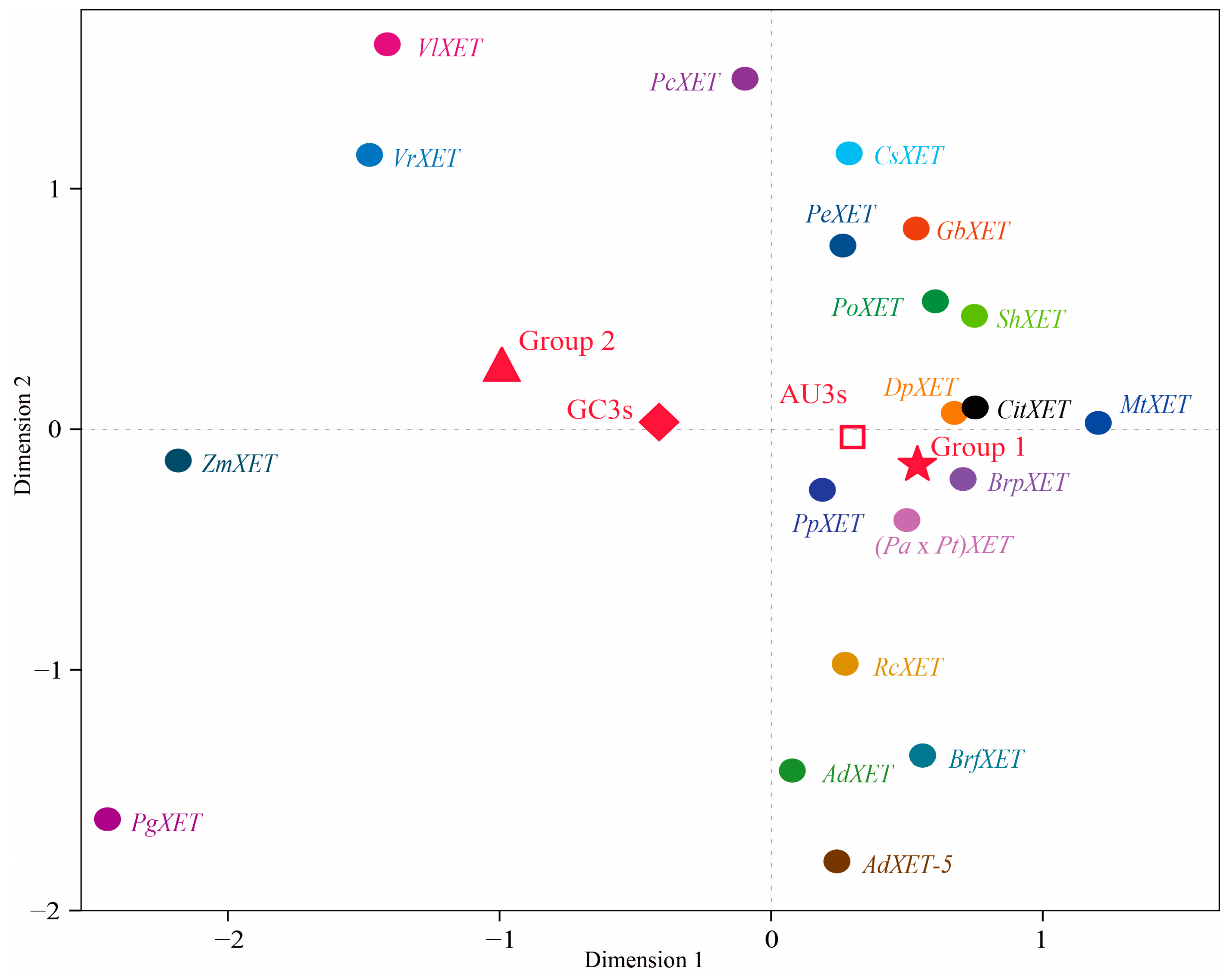
2.1. Clustering Analysis
2.2. Analysis of Codon Usage Bias in 20 XET Genes
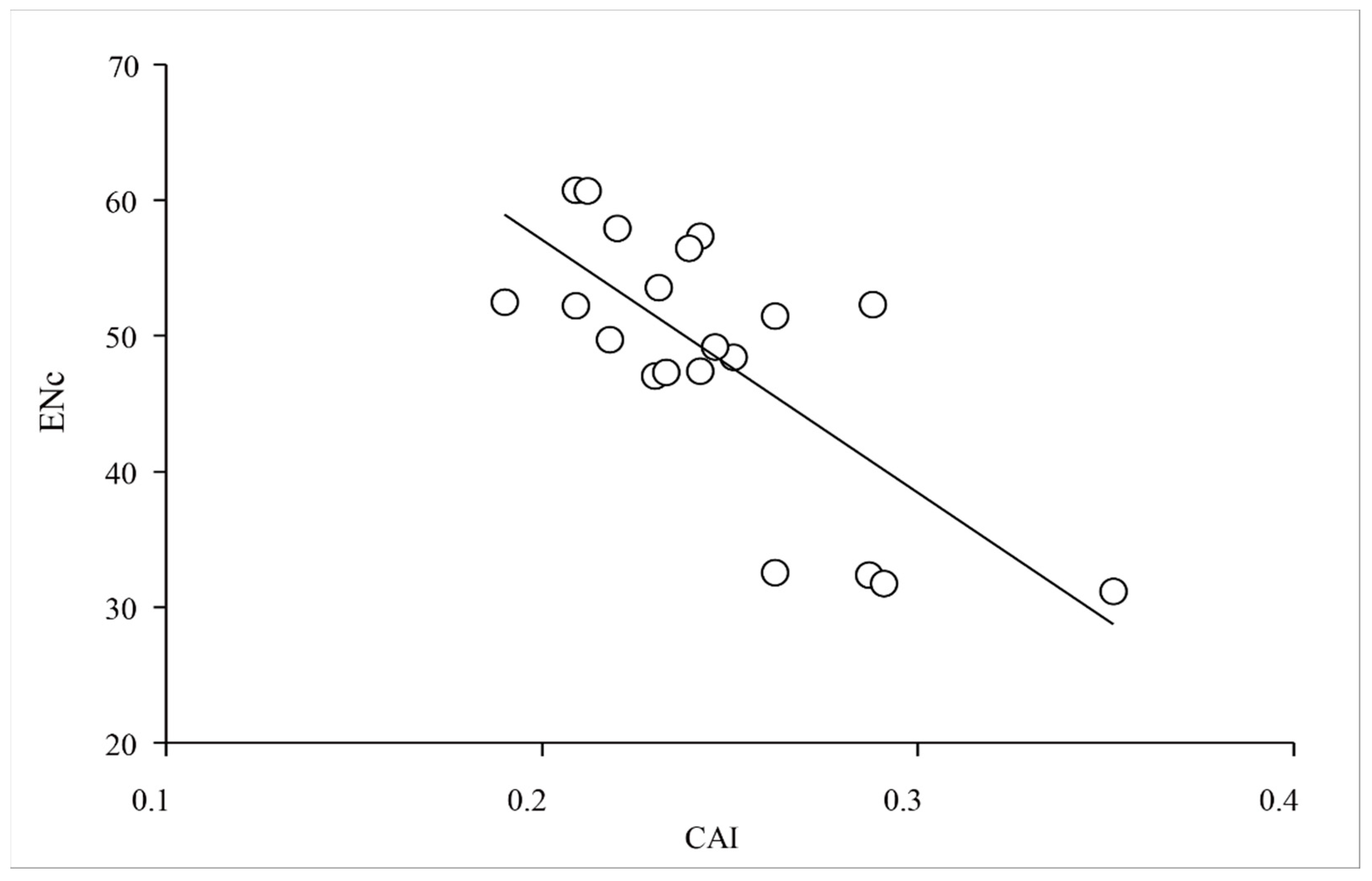
2.3. Base Composition Affects the Formation of Codon Usage Bias
3. Discussion
4. Materials and Methods
4.1. Sequence Data
4.2. Analysis of Codon Usage in XET Coding Sequences
4.3. Phylogenetic and Hierarchical Cluster Analyses
4.4. Comparison of the XET Codon Usage Patterns
4.5. Correspondence and Multidimensional Preference Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ingvarsson, P.K. Molecular evolution of synonymous codon usage in Populus. BMC Evol. Biol. 2008, 8, 307. [Google Scholar] [CrossRef]
- Gao, Y.; Lu, Y.; Song, Y.; Jing, L. Analysis of codon usage bias of WRKY transcription factors in Helianthus annuus. BMC Genom. Data 2022, 23, 46. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zheng, H.; Xu, A.; Yan, D.; Jiang, Z.; Qi, Q.; Sun, J. Analysis of codon usage bias of envelope glycoprotein genes in nuclear polyhedrosis virus (NPV) and its relation to evolution. BMC Genom. 2016, 17, 677. [Google Scholar] [CrossRef]
- Tsai, C.J.; Sauna, Z.E.; Kimchi-Sarfaty, C.; Ambudkar, S.V.; Gottesman, M.M.; Nussinov, R. Synonymous mutations and ribosome stalling can lead to altered folding pathways and distinct minima. J. Mol. Biol. 2008, 383, 281–291. [Google Scholar] [CrossRef] [PubMed]
- Marin, M. Folding at the rhythm of the rare codon beat. Biotechnol. J. 2008, 3, 1047–1057. [Google Scholar] [CrossRef] [PubMed]
- Hao, J.; Liang, Y.; Ping, J.; Li, J.; Shi, W.; Su, Y.; Wang, T. Chloroplast gene expression level is negatively correlated with evolutionary rates and selective pressure while positively with codon usage bias in Ophioglossum vulgatum L. BMC Plant Biol. 2022, 22, 580. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liu, S.; Zhang, B.; Wei, W. Analysis of Synonymous Codon Usage Bias of Zika Virus and Its Adaption to the Hosts. PLoS ONE 2016, 11, e0166260. [Google Scholar] [CrossRef]
- Babbitt, G.A.; Alawad, M.A.; Schulze, K.V.; Hudson, A.O. Synonymous codon bias and functional constraint on GC3-related DNA backbone dynamics in the prokaryotic nucleoid. Nucleic Acids Res. 2014, 42, 10915–10926. [Google Scholar] [CrossRef]
- Shah, P.; Gilchrist, M.A. Explaining complex codon usage patterns with selection for translational efficiency, mutation bias, and genetic drift. Proc. Natl. Acad. Sci. USA 2011, 108, 10231–10236. [Google Scholar] [CrossRef]
- Supek, F.; Skunca, N.; Repar, J.; Vlahovicek, K.; Smuc, T. Translational selection is ubiquitous in prokaryotes. PLoS Genet. 2010, 6, e1001004. [Google Scholar] [CrossRef]
- Plotkin, J.B.; Dushoff, J.; Desai, M.M.; Fraser, H.B. Codon usage and selection on proteins. J. Mol. Evol. 2006, 63, 635–653. [Google Scholar] [CrossRef]
- Bulmer, M. The selection-mutation-drift theory of synonymous codon usage. Genetics 1991, 129, 897–907. [Google Scholar] [CrossRef] [PubMed]
- Dos Reis, M.; Savva, R.; Wernisch, L. Solving the riddle of codon usage preferences: A test for translational selection. Nucleic Acids Res. 2004, 32, 5036–5044. [Google Scholar] [CrossRef] [PubMed]
- Stoletzki, N. The surprising negative correlation of gene length and optimal codon use—Disentangling translational selection from GC-biased gene conversion in yeast. BMC Evol. Biol. 2011, 11, 93. [Google Scholar] [CrossRef] [PubMed]
- Qin, H.; Wu, W.B.; Comeron, J.M.; Kreitman, M.; Li, W.H. Intragenic spatial patterns of codon usage bias in prokaryotic and eukaryotic genomes. Genetics 2004, 168, 2245–2260. [Google Scholar] [CrossRef] [PubMed]
- Kanaya, S.; Kinouchi, M.; Abe, T.; Kudo, Y.; Yamada, Y.; Nishi, T.; Mori, H.; Ikemura, T. Analysis of codon usage diversity of bacterial genes with a self-organizing map (SOM): Characterization of horizontally transferred genes with emphasis on the E. coli O157 genome. Gene 2001, 276, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Van Sandt, V.S.; Suslov, D.; Verbelen, J.P.; Vissenberg, K. Xyloglucan endotransglucosylase activity loosens a plant cell wall. Ann. Bot. 2007, 100, 1467–1473. [Google Scholar] [CrossRef]
- Dogra, V.; Sharma, R.; Yelam, S. Xyloglucan endo-transglycosylase/hydrolase (XET/H) gene is expressed during the seed germination in Podophyllum hexandrum: A high altitude Himalayan plant. Planta 2016, 244, 505–515. [Google Scholar] [CrossRef]
- Baumann, M.J.; Eklof, J.M.; Michel, G.; Kallas, A.M.; Teeri, T.T.; Czjzek, M.; Brumer, H., 3rd. Structural evidence for the evolution of xyloglucanase activity from xyloglucan endo-transglycosylases: Biological implications for cell wall metabolism. Plant Cell 2007, 19, 1947–1963. [Google Scholar] [CrossRef]
- Becnel, J.; Natarajan, M.; Kipp, A.; Braam, J. Developmental expression patterns of Arabidopsis XTH genes reported by transgenes and Genevestigator. Plant Mol. Biol. 2006, 61, 451–467. [Google Scholar] [CrossRef]
- Ma, Y.S.; Jie, H.D.; Zhao, L.; Lv, X.Y.; Liu, X.C.; Tang, Y.Y.; Zhang, Y.; He, P.L.; Xing, H.C.; Jie, Y.C. Identification of the Xyloglucan Endotransglycosylase/Hydrolase (XTH) Gene Family Members Expressed in Boehmeria nivea in Response to Cadmium Stress. Int. J. Mol. Sci. 2022, 23, 16104. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.E.; Fry, S.C. Restructuring of wall-bound xyloglucan by transglycosylation in living plant cells. Plant J. 2001, 26, 23–34. [Google Scholar] [CrossRef]
- Fry, S.C. Primary cell wall metabolism: Tracking the careers of wall polymers in living plant cells. New Phytol. 2004, 161, 641–675. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Li, H.; Yin, C.; Wang, X.; Jiang, Q.; Zhang, R.; Ge, F.; Chen, Y.; Yang, L. Genome-Wide Identification and Characterization of Xyloglucan Endotransglycosylase/Hydrolase in Ananas comosus during Development. Genes 2019, 10, 537. [Google Scholar] [CrossRef]
- Frankova, L.; Fry, S.C. Biochemistry and physiological roles of enzymes that ‘cut and paste’ plant cell-wall polysaccharides. J. Exp. Bot. 2013, 64, 3519–3550. [Google Scholar] [CrossRef] [PubMed]
- Iurlaro, A.; De Caroli, M.; Sabella, E.; De Pascali, M.; Rampino, P.; De Bellis, L.; Perrotta, C.; Dalessandro, G.; Piro, G.; Fry, S.C.; et al. Drought and Heat Differentially Affect XTH Expression and XET Activity and Action in 3-Day-Old Seedlings of Durum Wheat Cultivars with Different Stress Susceptibility. Front. Plant Sci. 2016, 7, 1686. [Google Scholar] [CrossRef] [PubMed]
- Holland, C.; Simmons, T.J.; Meulewaeter, F.; Hudson, A.; Fry, S.C. Three highly acidic Equisetum XTHs differ from hetero-trans-beta-glucanase in donor substrate specificity and are predominantly xyloglucan homo-transglucosylases. J. Plant Physiol. 2020, 251, 153210. [Google Scholar] [CrossRef]
- Van Sandt, V.S.; Stieperaere, H.; Guisez, Y.; Verbelen, J.P.; Vissenberg, K. XET activity is found near sites of growth and cell elongation in bryophytes and some green algae: New insights into the evolution of primary cell wall elongation. Ann. Bot. 2007, 99, 39–51. [Google Scholar] [CrossRef]
- Vissenberg, K.; Oyama, M.; Osato, Y.; Yokoyama, R.; Verbelen, J.P.; Nishitani, K. Differential expression of AtXTH17, AtXTH18, AtXTH19 and AtXTH20 genes in Arabidopsis roots. Physiological roles in specification in cell wall construction. Plant Cell Physiol. 2005, 46, 192–200. [Google Scholar] [CrossRef]
- Zhu, X.F.; Shi, Y.Z.; Lei, G.J.; Fry, S.C.; Zhang, B.C.; Zhou, Y.H.; Braam, J.; Jiang, T.; Xu, X.Y.; Mao, C.Z.; et al. XTH31, encoding an in vitro XEH/XET-active enzyme, regulates aluminum sensitivity by modulating in vivo XET action, cell wall xyloglucan content, and aluminum binding capacity in Arabidopsis. Plant Cell 2012, 24, 4731–4747. [Google Scholar] [CrossRef]
- Hara, Y.; Yokoyama, R.; Osakabe, K.; Toki, S.; Nishitani, K. Function of xyloglucan endotransglucosylase/hydrolases in rice. Ann. Bot. 2014, 114, 1309–1318. [Google Scholar] [CrossRef]
- Vissenberg, K.; Van Sandt, V.; Fry, S.C.; Verbelen, J.P. Xyloglucan endotransglucosylase action is high in the root elongation zone and in the trichoblasts of all vascular plants from Selaginella to Zea mays. J. Exp. Bot. 2003, 54, 335–344. [Google Scholar] [CrossRef]
- Park, Y.B.; Cosgrove, D.J. Xyloglucan and its interactions with other components of the growing cell wall. Plant Cell Physiol. 2015, 56, 180–194. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, S.; Nag, D.; Mazumder, T.H.; Uddin, A. Codon usage pattern and prediction of gene expression level in Bungarus species. Gene 2017, 604, 48–60. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Xing, H.; Yuan, Y.; Wang, X.; Saeed, M.; Tao, J.; Feng, W.; Zhang, G.; Song, X.; Sun, X. Genome-wide analysis of codon usage bias in four sequenced cotton species. PLoS ONE 2018, 13, e0194372. [Google Scholar] [CrossRef] [PubMed]
- Fages-Lartaud, M.; Hundvin, K.; Hohmann-Marriott, M.F. Mechanisms governing codon usage bias and the implications for protein expression in the chloroplast of Chlamydomonas reinhardtii. Plant J. 2022, 112, 919–945. [Google Scholar] [CrossRef]
- Rao, Y.S.; Wu, G.Z.; Wang, Z.F.; Chai, X.W.; Nie, Q.H.; Zhang, X.Q. Mutation Bias is the Driving Force of Codon Usage in the Gallus gallus genome. DNA Res. 2011, 18, 499–512. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Meng, T.; Wei, W. Analysis of synonymous codon usage bias in helicase gene from Autographa californica multiple nucleopolyhedrovirus. Genes Genom. 2018, 40, 767–780. [Google Scholar] [CrossRef] [PubMed]
- Stratilova, B.; Stratilova, E.; Hrmova, M.; Kozmon, S. Definition of the Acceptor Substrate Binding Specificity in Plant Xyloglucan Endotransglycosylases Using Computational Chemistry. Int. J. Mol. Sci. 2022, 23, 11838. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Liu, T.F.; Yang, D.Y.; Nong, X.; Xie, Y.; Fu, Y.; Wu, X.H.; Huang, X.; Gu, X.B.; Wang, S.X.; et al. Analysis of codon usage patterns in Taenia pisiformis through annotated transcriptome data. Biochem. Biophys. Res. Commun. 2013, 430, 1344–1348. [Google Scholar] [CrossRef]
- Kawabe, A.; Miyashita, N.T. Patterns of codon usage bias in three dicot and four monocot plant species. Genes Genet. Syst. 2003, 78, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Xiong, B.; Gu, X.; Qiu, X.; Dong, Z.; Ye, S.; Sun, G.; Huang, S.; Liu, X.; Xi, L.; Wang, Z. Variability in CitXET expression and XET activity in Citrus cultivar Huangguogan seedlings with differed degrees of etiolation. PLoS ONE 2017, 12, e0178973. [Google Scholar] [CrossRef]
- Nasrullah, I.; Butt, A.M.; Tahir, S.; Idrees, M.; Tong, Y. Genomic analysis of codon usage shows influence of mutation pressure, natural selection, and host features on Marburg virus evolution. BMC Evol. Biol. 2015, 15, 174. [Google Scholar] [CrossRef] [PubMed]
- Vicario, S.; Moriyama, E.N.; Powell, J.R. Codon usage in twelve species of Drosophila. BMC Evol. Biol. 2007, 7, 226. [Google Scholar] [CrossRef]
- Tao, P.; Dai, L.; Luo, M.; Tang, F.; Tien, P.; Pan, Z. Analysis of synonymous codon usage in classical swine fever virus. Virus Genes 2009, 38, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Qiu, S.; Bergero, R.; Zeng, K.; Charlesworth, D. Patterns of codon usage bias in Silene latifolia. Mol. Biol. Evol. 2011, 28, 771–780. [Google Scholar] [CrossRef]
- Auewarakul, P. Composition bias and genome polarity of RNA viruses. Virus Res. 2005, 109, 33–37. [Google Scholar] [CrossRef]
- Shackelton, L.A.; Parrish, C.R.; Holmes, E.C. Evolutionary basis of codon usage and nucleotide composition bias in vertebrate DNA viruses. J. Mol. Evol. 2006, 62, 551–563. [Google Scholar] [CrossRef]
- Puigbo, P.; Bravo, I.G.; Garcia-Vallve, S. CAIcal: A combined set of tools to assess codon usage adaptation. Biol. Direct. 2008, 3, 38. [Google Scholar] [CrossRef]
- Lu, H.; Zhao, W.M.; Zheng, Y.; Wang, H.; Qi, M.; Yu, X.P. Analysis of synonymous codon usage bias in Chlamydia. Acta Biochim. Biophys. Sin. (Shanghai) 2005, 37, 1–10. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Sharp, P.-M.; Li, W.-H. Codon usage in regulatory genes in Escherichia coli does not reflect selection for ‘rare’ codons. Nucleic Acids Res. 1986, 14, 7737–7749. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.J.; Zhou, J.; Li, Z.F.; Wang, L.; Gu, X.; Zhong, Y. Comparative Analysis of Codon Usage Patterns Among Mitochondrion, Chloroplast and Nuclear Genes in Triticum aestivum L. J. Integr. Plant Biol. 2007, 49, 246–254. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Accession Number | Genes | U3s/% | C3s/% | A3s/% | G3s /% | GC3s/% | GC/% | CAI | CBI | Fop | ENc | Gravy | Aromo |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | KY576851 | Citrus cultivar (Huangguogan) XET | 43.70 | 27.56 | 36.54 | 22.51 | 37.7 | 41.7 | 0.242 | 0.046 | 0.46 | 47.39 | −0.39 | 0.18 |
| 2 | KU705236.1 | Populus euphratica XET | 36.36 | 41.96 | 21.24 | 26.21 | 53.4 | 46.2 | 0.251 | 0.140 | 0.52 | 48.40 | −0.52 | 0.18 |
| 3 | AF052194.1 | Actinidia deliciosa XET | 26.54 | 43.60 | 22.29 | 38.04 | 61.8 | 51.0 | 0.239 | 0.071 | 0.47 | 56.46 | −0.42 | 0.17 |
| 4 | L46792.1 | Actinidia deliciosa XET-5 | 28.99 | 41.18 | 22.28 | 36.67 | 59.4 | 50.4 | 0.231 | 0.083 | 0.48 | 53.56 | −0.50 | 0.17 |
| 5 | XM_009140285.2 | Brassica rapa (field mustard) XET | 39.34 | 25.82 | 37.88 | 24.04 | 38.5 | 44.9 | 0.208 | −0.058 | 0.39 | 60.70 | −0.37 | 0.15 |
| 6 | EU579461.1 | Brassica rapa subsp. pekinensis XET | 39.34 | 25.82 | 38.38 | 23.50 | 38.1 | 44.9 | 0.209 | −0.058 | 0.39 | 60.71 | −0.38 | 0.15 |
| 7 | KU530158.1 | Camellia sinensis XET | 39.78 | 40.86 | 22.67 | 23.88 | 50.0 | 44.8 | 0.242 | 0.054 | 0.46 | 57.34 | −0.41 | 0.17 |
| 8 | HM053613.1 | Dahlia pinnata XET | 38.27 | 29.63 | 35.53 | 22.16 | 40.9 | 46.8 | 0.220 | −0.019 | 0.41 | 57.91 | −0.35 | 0.14 |
| 9 | DQ912942.1 | Gossypium barbadense XET | 39.68 | 30.95 | 33.00 | 26.67 | 43.2 | 45.5 | 0.288 | 0.092 | 0.48 | 52.30 | −0.57 | 0.16 |
| 10 | DQ855285.2 | Medicago truncatula XET | 48.72 | 20.94 | 44.85 | 15.08 | 27.4 | 38.8 | 0.230 | −0.067 | 0.39 | 47.04 | −0.53 | 0.16 |
| 11 | KT890352.1 | Paeonia ostii var. lishizhenii XET | 32.86 | 37.14 | 39.25 | 22.11 | 45.3 | 43.9 | 0.209 | 0.020 | 0.45 | 52.22 | −0.68 | 0.18 |
| 12 | HQ416697.2 | Pennisetum glaucum XET | 5.53 | 67.66 | 2.43 | 47.50 | 93.4 | 67.5 | 0.352 | 0.357 | 0.64 | 31.14 | −0.44 | 0.14 |
| 13 | JX431932.1 | Populus alba x Populus tremula var XET | 29.00 | 38.10 | 30.00 | 33.52 | 54.4 | 48.0 | 0.218 | 0.038 | 0.46 | 49.73 | −0.45 | 0.17 |
| 14 | AJ811689.1 | Pyrus communis XET | 22.35 | 49.41 | 22.67 | 36.11 | 65.4 | 54.3 | 0.246 | 0.085 | 0.49 | 49.18 | −0.90 | 0.16 |
| 15 | EU432411.1 | Pyrus pyrifolia XET | 28.14 | 42.42 | 23.20 | 37.36 | 60.1 | 49.7 | 0.262 | 0.148 | 0.51 | 51.44 | −0.45 | 0.16 |
| 16 | GU320707.1 | Rosa chinensis XET | 26.81 | 45.11 | 22.61 | 34.22 | 61.2 | 50.3 | 0.233 | 0.126 | 0.50 | 47.28 | −0.53 | 0.16 |
| 17 | KM034907.1 | Sinopodophyllum hexandrum XET | 35.11 | 29.79 | 40.88 | 26.00 | 42.0 | 44.4 | 0.190 | −0.146 | 0.36 | 52.47 | −0.60 | 0.16 |
| 18 | FJ896376.1 | Vigna luteola XET | 2.38 | 68.25 | 0.00 | 53.85 | 97.9 | 67.8 | 0.291 | 0.309 | 0.61 | 31.75 | −0.45 | 0.14 |
| 19 | FJ896377.1 | Vigna radiata cultivar T44 XET | 2.40 | 68.00 | 0.93 | 53.92 | 97.2 | 67.1 | 0.287 | 0.301 | 0.60 | 32.36 | −0.48 | 0.14 |
| 20 | FJ940680.1 | Zea mays XET | 1.68 | 59.66 | 2.88 | 58.91 | 96.3 | 69.5 | 0.262 | 0.313 | 0.61 | 32.54 | −0.41 | 0.12 |
| Amino Acid | Codon | Number | RSCU | Amino Acid | Codon | Number | RSCU |
|---|---|---|---|---|---|---|---|
| Phe | UUU * | 105 | 0.58 | Ala | GCU * | 106 | 1.42 |
| UUC | 221 | 1.42 | GCC | 104 | 1.40 | ||
| Leu | UUA * | 28 | 0.63 | GCA * | 50 | 0.64 | |
| UUG * | 57 | 1.14 | GCG * | 48 | 0.54 | ||
| CUU * | 47 | 1.03 | Tyr | UAU * | 73 | 0.54 | |
| CUC | 68 | 1.42 | UAC | 182 | 1.46 | ||
| CUA | 21 | 0.43 | His | CAU * | 44 | 0.83 | |
| CUG | 67 | 1.36 | CAC | 58 | 1.18 | ||
| Ile | AUU * | 76 | 0.94 | Gln | CAA * | 85 | 0.82 |
| AUC | 89 | 1.55 | CAG | 132 | 1.18 | ||
| AUA * | 42 | 0.51 | Asn | AAU * | 66 | 0.62 | |
| Val | GUU * | 67 | 0.96 | AAC | 132 | 1.38 | |
| GUC | 60 | 0.94 | Lys | AAA * | 99 | 0.71 | |
| GUA * | 38 | 0.60 | AAG | 146 | 1.29 | ||
| GUG | 113 | 1.50 | Asp | GAU * | 153 | 0.86 | |
| Ser | UCU * | 73 | 1.24 | GAC | 183 | 1.14 | |
| UCC | 68 | 1.44 | Glu | GAA * | 59 | 0.62 | |
| UCA | 49 | 0.87 | GAG | 115 | 1.38 | ||
| UCG | 42 | 0.72 | Cys | UGU * | 32 | 0.64 | |
| AGU * | 39 | 0.77 | UGC | 47 | 1.06 | ||
| AGC | 51 | 0.96 | Arg | CGU * | 25 | 0.45 | |
| Pro | CCU * | 56 | 0.85 | CGC | 51 | 0.92 | |
| CCC | 58 | 1.03 | CGA * | 22 | 0.39 | ||
| CCA * | 76 | 1.19 | CGG | 30 | 0.61 | ||
| CCG | 49 | 0.93 | Arg | AGA * | 86 | 1.89 | |
| Thr | ACU * | 84 | 1.24 | AGG * | 84 | 1.74 | |
| ACC | 74 | 1.17 | Gly | GGU * | 77 | 0.97 | |
| ACA * | 69 | 1.02 | GGC | 112 | 1.06 | ||
| ACG | 36 | 0.57 | GGA * | 99 | 1.15 | ||
| GGG | 77 | 0.82 |
| Amino Acid | Codon | Number | RSCU | Amino Acid | Codon | Number | RSCU |
|---|---|---|---|---|---|---|---|
| Phe | UUU * | 13 | 0.90 | Ala | GCU * | 10 | 2.67 |
| UUC | 16 | 1.10 | GCC | 3 | 0.80 | ||
| Leu | UUA * | 0 | 0.00 | GCA * | 1 | 0.27 | |
| UUG * | 5 | 1.50 | GCG * | 1 | 0.27 | ||
| CUU * | 8 | 2.40 | Tyr | UAU * | 3 | 0.35 | |
| CUC | 2 | 0.60 | UAC | 14 | 1.65 | ||
| CUA | 0 | 0.00 | His | CAU * | 4 | 1.33 | |
| CUG | 5 | 1.50 | CAC | 2 | 0.67 | ||
| Ile | AUU * | 9 | 1.59 | Gln | CAA * | 6 | 0.86 |
| AUC | 3 | 0.53 | CAG | 8 | 1.14 | ||
| AUA * | 5 | 0.88 | Asn | AAU * | 8 | 1.33 | |
| Val | GUU * | 7 | 1.65 | AAC | 4 | 0.67 | |
| GUC | 3 | 0.71 | Lys | AAA * | 17 | 1.62 | |
| GUA * | 1 | 0.24 | AAG | 4 | 0.38 | ||
| GUG | 6 | 1.41 | Asp | GAU * | 16 | 1.45 | |
| Ser | UCU * | 7 | 2.10 | GAC | 6 | 0.55 | |
| UCC | 2 | 0.60 | Glu | GAA * | 6 | 1.09 | |
| UCA | 6 | 1.80 | GAG | 5 | 0.91 | ||
| UCG | 1 | 0.30 | Cys | UGU * | 4 | 1.30 | |
| AGU * | 4 | 1.20 | UGC | 2 | 0.70 | ||
| AGC | 0 | 0.00 | Arg | CGU * | 3 | 0.90 | |
| Pro | CCU * | 5 | 1.33 | CGC | 2 | 0.60 | |
| CCC | 1 | 0.27 | CGA * | 1 | 0.30 | ||
| CCA * | 7 | 1.87 | CGG | 0 | 0.00 | ||
| CCG | 2 | 0.53 | Arg | AGA * | 13 | 3.90 | |
| Thr | ACU * | 6 | 1.50 | AGG * | 1 | 0.30 | |
| ACC | 2 | 0.50 | Gly | GGU * | 4 | 0.70 | |
| ACA * | 6 | 1.50 | GGC | 8 | 1.50 | ||
| ACG | 2 | 0.50 | GGA * | 7 | 1.30 | ||
| GGG | 3 | 0.60 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiong, B.; Wang, T.; Huang, S.; Liao, L.; Wang, X.; Deng, H.; Zhang, M.; He, J.; Sun, G.; He, S.; et al. Analysis of Codon Usage Bias in Xyloglucan Endotransglycosylase (XET) Genes. Int. J. Mol. Sci. 2023, 24, 6108. https://doi.org/10.3390/ijms24076108
Xiong B, Wang T, Huang S, Liao L, Wang X, Deng H, Zhang M, He J, Sun G, He S, et al. Analysis of Codon Usage Bias in Xyloglucan Endotransglycosylase (XET) Genes. International Journal of Molecular Sciences. 2023; 24(7):6108. https://doi.org/10.3390/ijms24076108
Chicago/Turabian StyleXiong, Bo, Tie Wang, Shengjia Huang, Ling Liao, Xun Wang, Honghong Deng, Mingfei Zhang, Jiaxian He, Guochao Sun, Siya He, and et al. 2023. "Analysis of Codon Usage Bias in Xyloglucan Endotransglycosylase (XET) Genes" International Journal of Molecular Sciences 24, no. 7: 6108. https://doi.org/10.3390/ijms24076108
APA StyleXiong, B., Wang, T., Huang, S., Liao, L., Wang, X., Deng, H., Zhang, M., He, J., Sun, G., He, S., & Wang, Z. (2023). Analysis of Codon Usage Bias in Xyloglucan Endotransglycosylase (XET) Genes. International Journal of Molecular Sciences, 24(7), 6108. https://doi.org/10.3390/ijms24076108