

Effect of Probiotic Lactobacillus plantarum on Streptococcus mutans and Candida albicans Clinical Isolates from Children with Early Childhood Caries

 , , ,
, , ,
Abstract
1. Introduction
2. Results
2.1. Characteristics of S-ECC Children whose S. mutans and C. albicans Were Isolated
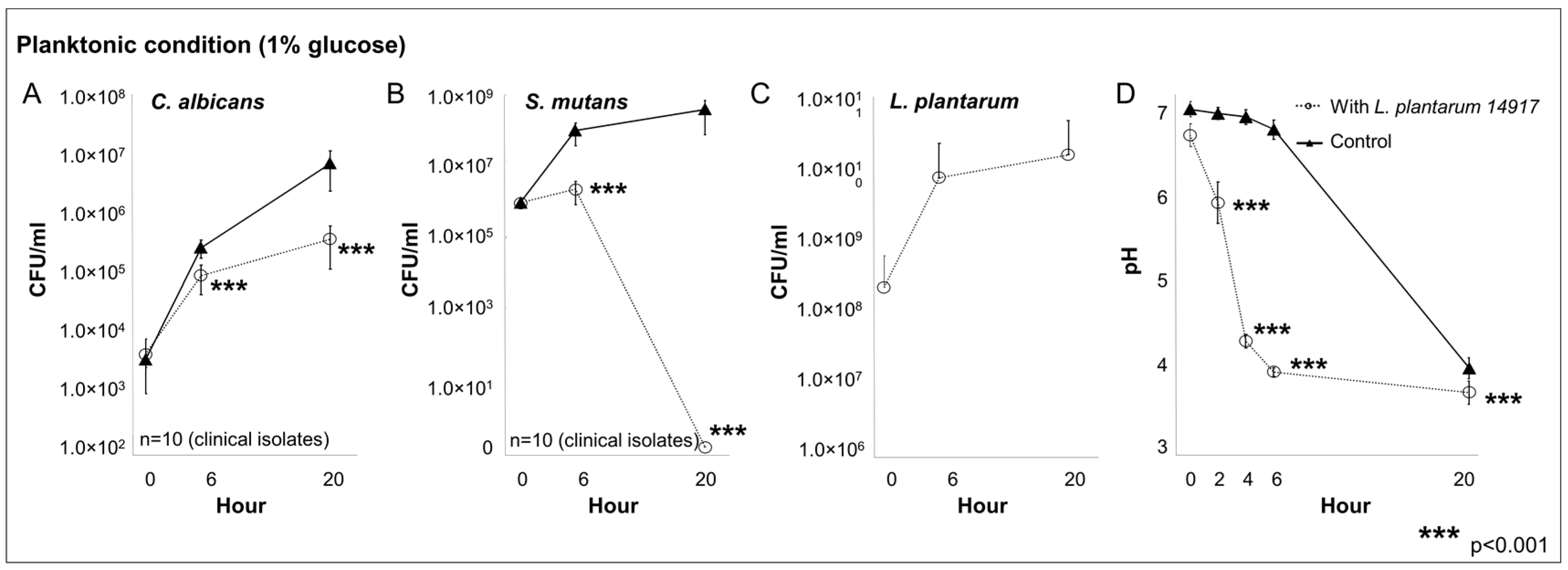
2.2. L. plantarum 14917 Inhibited the Growth of S. mutans and C. albicans Clinical Isolates in Planktonic Condition
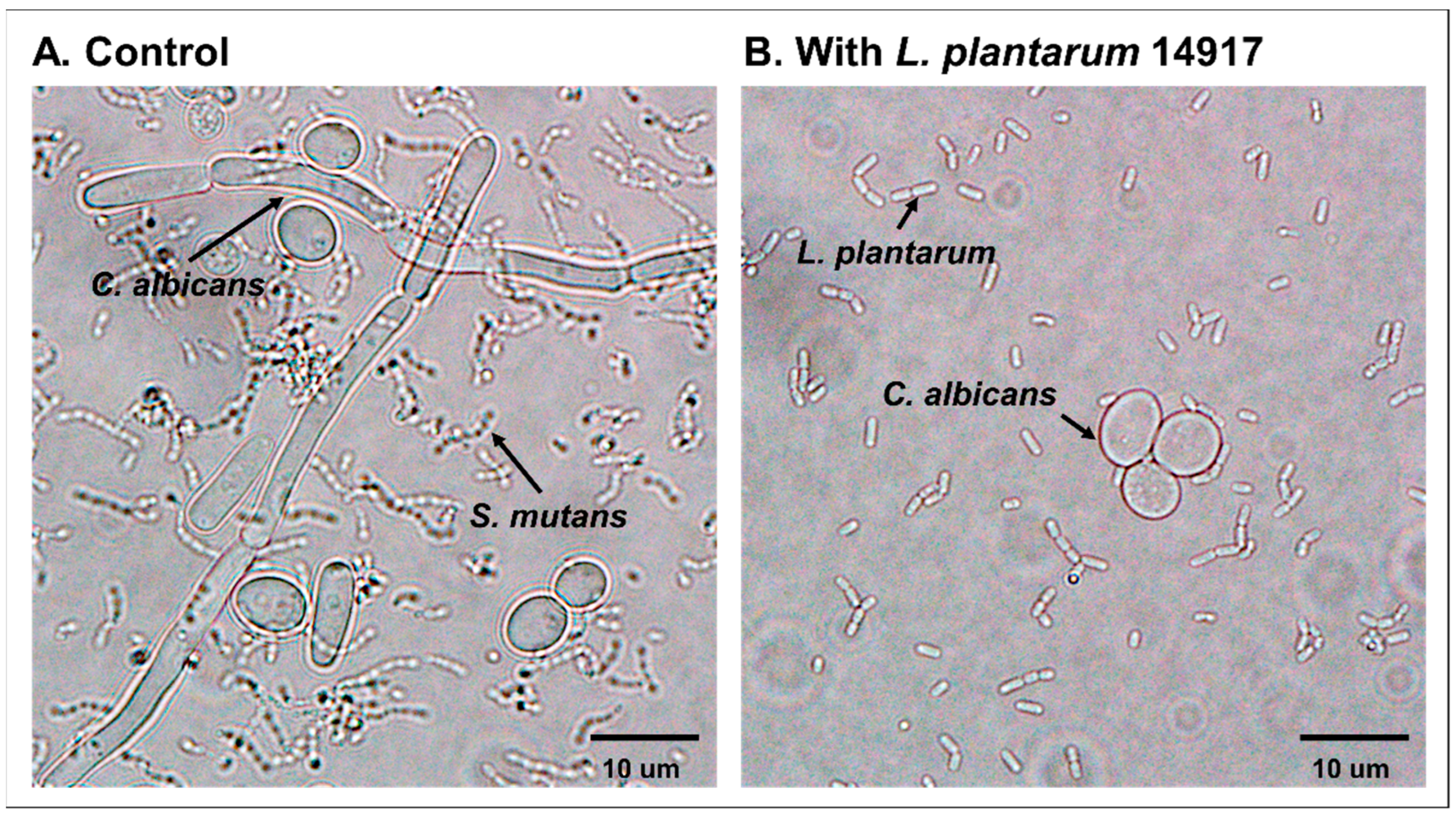
2.3. Inhibition of C. albicans Hypha Formation by L. plantarum
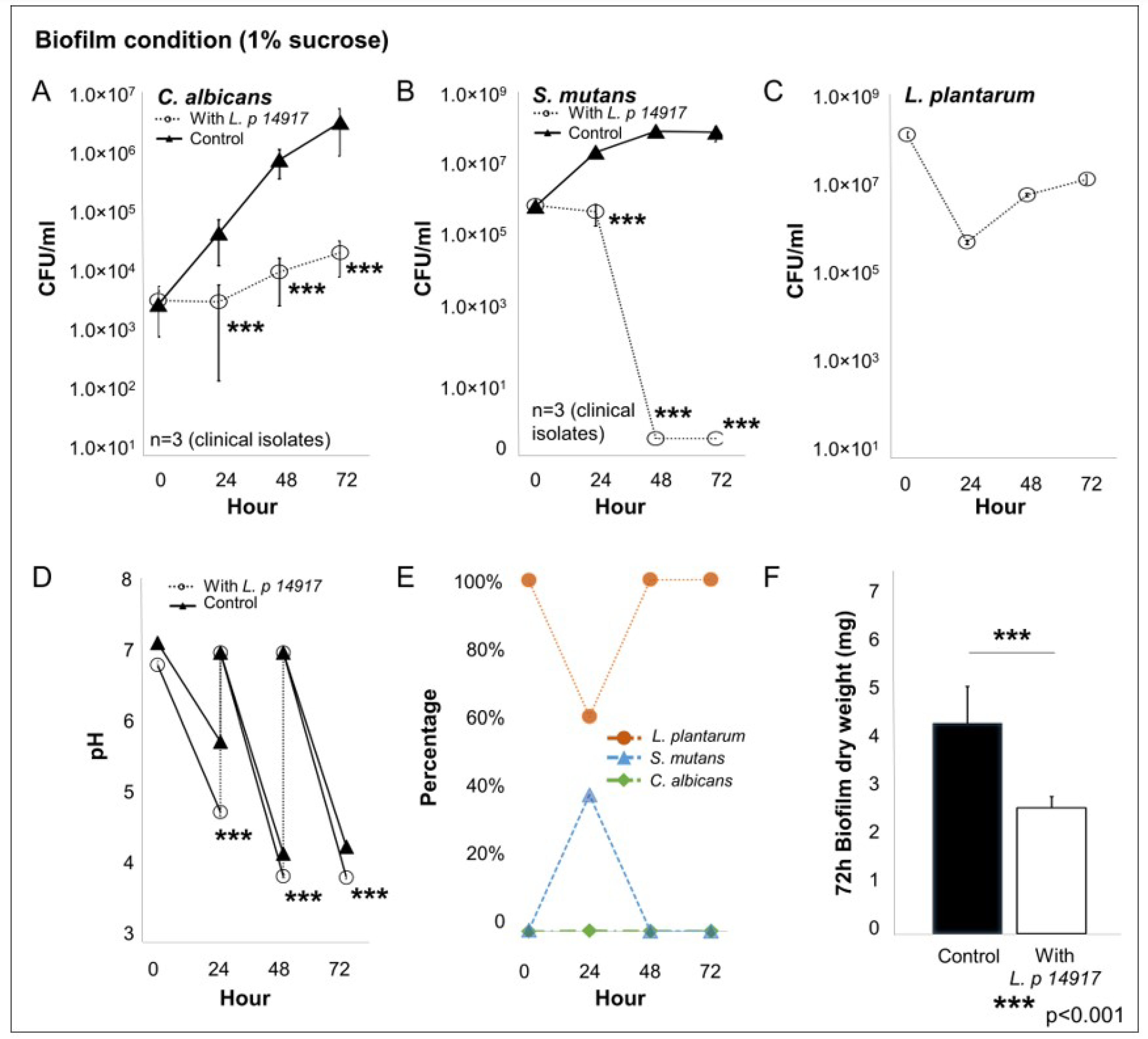
2.4. L. plantarum 14917 Inhibited Biofilm Formation by S. mutans and C. albicans Clinical Isolates
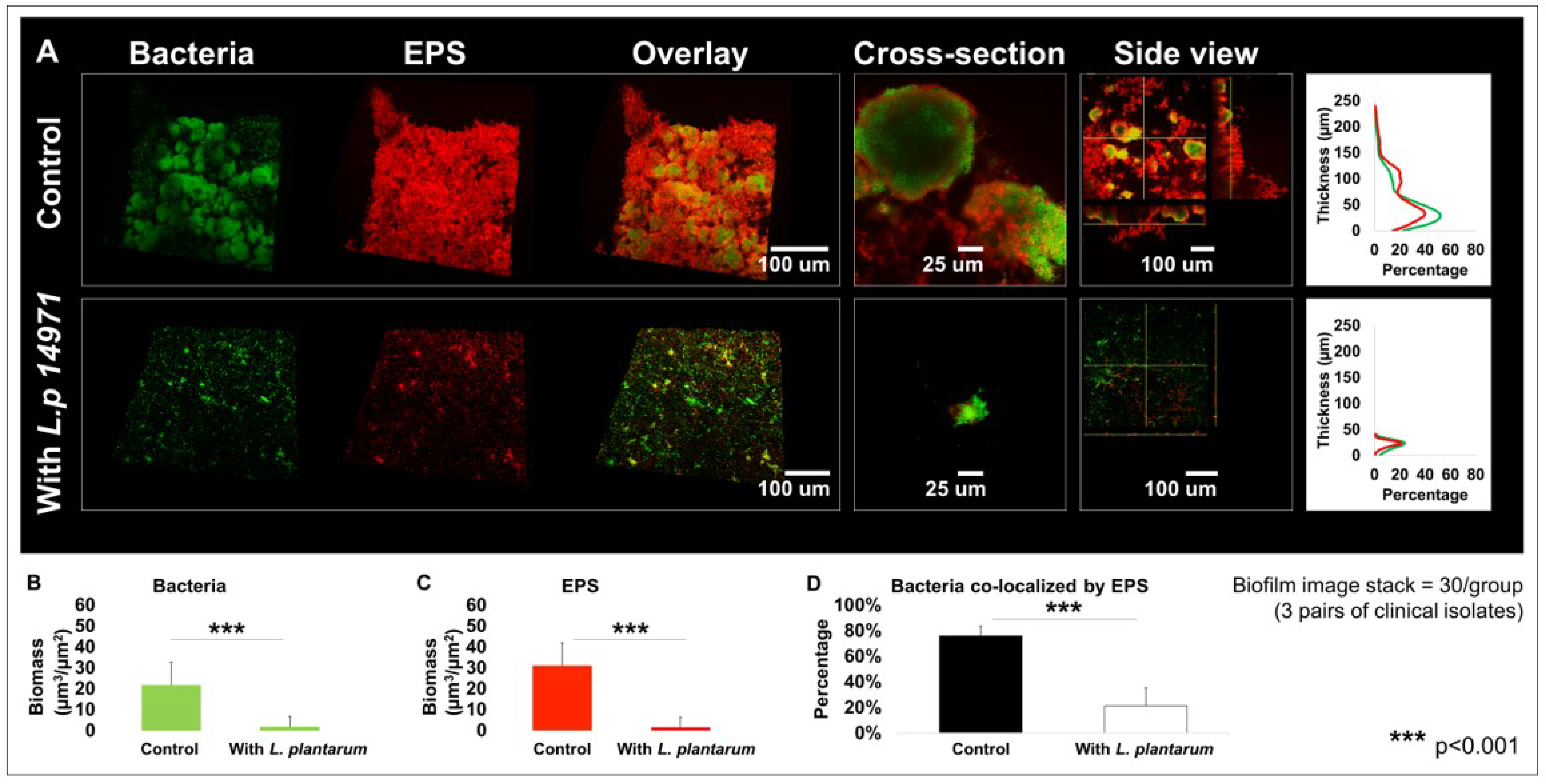
2.5. L. plantarum 14917 Altered 3D Structure of Biofilms Formed by S. mutans and C. albicans Clinical Isolates
2.6. Plantaricin Inhibited the Growth of S. mutans and C. albicans Clinical Isolates
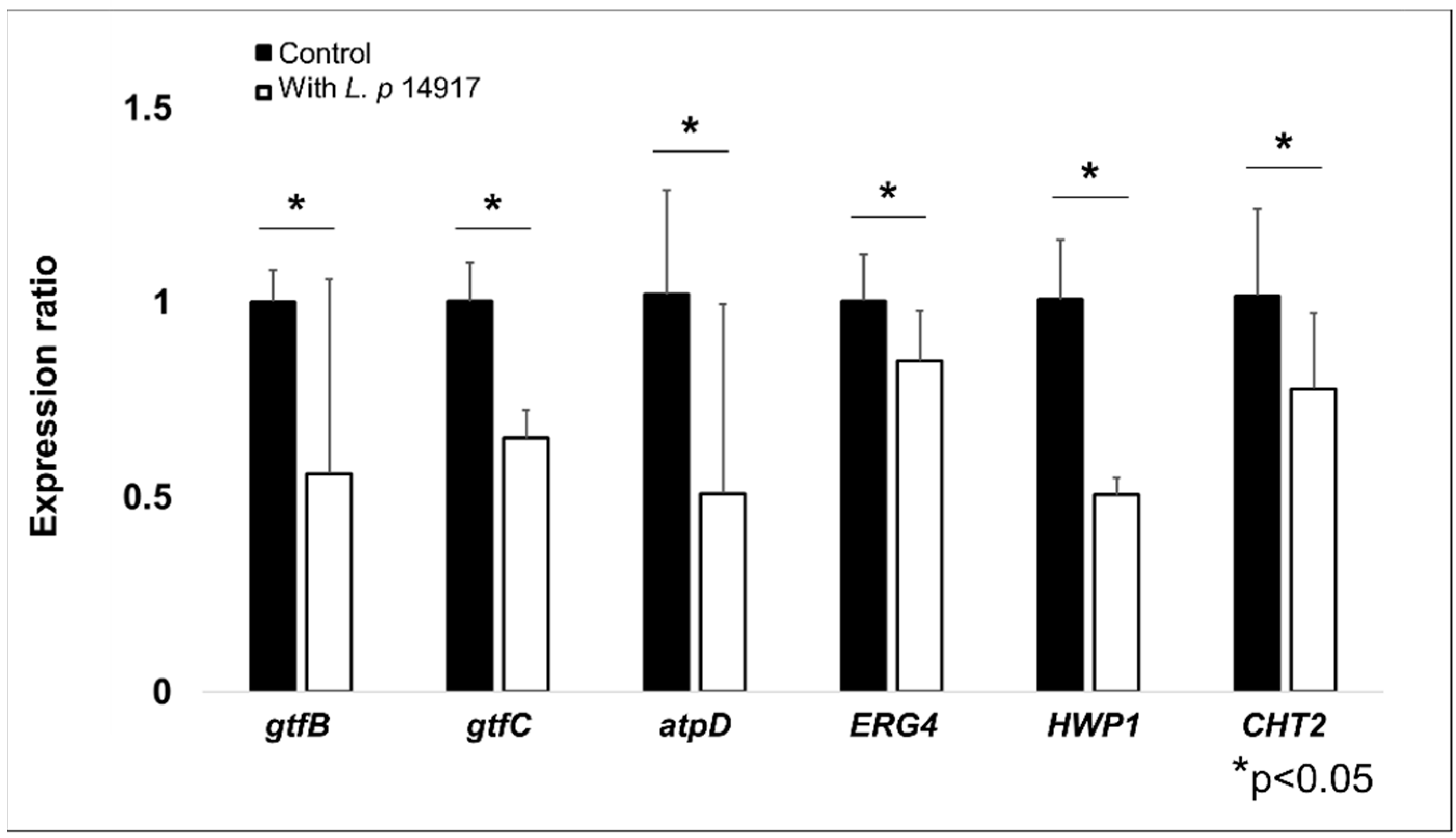
2.7. L. plantarum 14917 Downregulated C. albicans and S. mutans Virulence Genes in Biofilms
3. Discussion
3.1. Susceptibility of Clinically Isolated S. mutans and C. albicans to Common Antimicrobial Agents
3.2. Equal Effectiveness of L. plantarum 14917 on Clinical Isoalted C. albicans and S. mutans Compared to Wild-Type Strains
4. Materials and Methods
4.1. Study Design
4.2. Bacterial Strains and Starter Preparation
4.3. Planktonic Model
4.4. Mixed-Species Biofilm Model
4.5. Microbiological Analysis of the Mixed-Species Bacterial Population
4.6. Laser Scanning Confocal Fluorescence Microscopy (LCSFM) Imaging of Biofilm Matrix
4.7. Computational Analyses of the Confocal Biofilm Images
4.8. Assessment of Total Protein in Multispecies Biofilms
4.9. Inhibition of S. mutans and C. albicans by Plantaricin
4.10. Real-Time Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
4.11. Assessment of Morphology of C. albicans
4.12. Statistical Analysis
4.13. Ethical Approval
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Heimer, M.; Teschler, M.; Schmitz, B.; Mooren, F.C. Health Benefits of Probiotics in Sport and Exercise—Non-existent or a Matter of Heterogeneity? A Systematic Review. Front. Nutr. 2022, 9, 804046. [Google Scholar] [CrossRef] [PubMed]
- Korada, S.K.; Yarla, N.S.; Mishra, V.; Daim, M.A.; Sharma, B.; Gm, A.; Reggi, R.; Palmery, M.; Peluso, I.; Kamal, M.A. Single Probiotic versus Multiple Probiotics—A Debate On Current Scenario for Alleviating Health Benefits. Curr. Pharm. Des. 2018, 24, 4150–4153. [Google Scholar] [CrossRef] [PubMed]
- Kechagia, M.; Basoulis, D.; Konstantopoulou, S.; Dimitriadi, D.; Gyftopoulou, K.; Skarmoutsou, N.; Fakiri, E.M. Health benefits of probiotics: A review. ISRN Nutr. 2013, 2013, 481651. [Google Scholar] [CrossRef] [PubMed]
- Schwendicke, F.; Dorfer, C.; Kneist, S.; Meyer-Lueckel, H.; Paris, S. Cariogenic effects of probiotic Lactobacillus rhamnosus GG in a dental biofilm model. Caries Res. 2014, 48, 186–192. [Google Scholar] [CrossRef]
- Haukioja, A. Probiotics and oral health. Eur. J. Dent. 2010, 4, 348–355. [Google Scholar] [CrossRef]
- Rastogi, P.; Saini, H.; Dixit, J.; Singhal, R. Probiotics and oral health. Natl. J. Maxillofac. Surg. 2011, 2, 6–9. [Google Scholar] [CrossRef]
- Twetman, L.; Larsen, U.; Fiehn, N.-E.; Stecksén-Blicks, C.; Twetman, S. Coaggregation between probiotic bacteria and caries-associated strains: An in vitro study. Acta Odontol. Scand. 2009, 67, 284–288. [Google Scholar] [CrossRef]
- Nishikawara, F.; Nomura, Y.; Imai, S.; Senda, A.; Hanada, N. Evaluation of cariogenic bacteria. Eur. J. Dent. 2007, 1, 31–39. [Google Scholar] [CrossRef]
- Zeng, Y.; Nikitkova, A.; Abdelsalam, H.; Li, J.; Xiao, J. Activity of quercetin and kaemferol against Streptococcus mutans biofilm. Arch. Oral Biol. 2019, 98, 9–16. [Google Scholar] [CrossRef]
- de Carvalho, F.G.; Silva, D.S.; Hebling, J.; Spolidorio, L.C.; Spolidorio, D.M. Presence of mutans streptococci and Candida spp. in dental plaque/dentine of carious teeth and early childhood caries. Arch. Oral Biol. 2006, 51, 1024–1028. [Google Scholar] [CrossRef]
- Signoretto, C.; Burlacchini, G.; Faccioni, F.; Zanderigo, M.; Bozzola, N.; Canepari, P. Support for the role of Candida spp. in extensive caries lesions of children. New Microbiol. 2009, 32, 101–107. [Google Scholar] [PubMed]
- Raja, M.; Hannan, A.; Ali, K. Association of oral candidal carriage with dental caries in children. Caries Res. 2010, 44, 272–276. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.Q.; Zhang, Q.; Lu, L.Y.; Yang, R.; Liu, Y.; Zou, J. Genotypic distribution of Candida albicans in dental biofilm of Chinese children associated with severe early childhood caries. Arch. Oral Biol. 2012, 57, 1048–1053. [Google Scholar] [CrossRef] [PubMed]
- Hasslof, P.; Hedberg, M.; Twetman, S.; Stecksen-Blicks, C. Growth inhibition of oral mutans streptococci and candida by commercial probiotic lactobacilli—An in vitro study. BMC Oral Health 2010, 10, 18. [Google Scholar] [CrossRef]
- Meurman, J.H.; Stamatova, I. Probiotics: Contributions to oral health. Oral Dis. 2007, 13, 443–451. [Google Scholar] [CrossRef]
- Comelli, E.M.; Guggenheim, B.; Stingele, F.; Neeser, J.R. Selection of dairy bacterial strains as probiotics for oral health. Eur. J. Oral Sci. 2002, 110, 218–224. [Google Scholar] [CrossRef]
- Ahola, A.J.; Yli-Knuuttila, H.; Suomalainen, T.; Poussa, T.; Ahlstrom, A.; Meurman, J.H.; Korpela, R. Short-term consumption of probiotic-containing cheese and its effect on dental caries risk factors. Arch. Oral Biol. 2002, 47, 799–804. [Google Scholar] [CrossRef]
- Hatakka, K.; Ahola, A.J.; Yli-Knuuttila, H.; Richardson, M.; Poussa, T.; Meurman, J.H.; Korpela, R. Probiotics reduce the prevalence of oral candida in the elderly—A randomized controlled trial. J. Dent. Res. 2007, 86, 125–130. [Google Scholar] [CrossRef]
- Krzysciak, W.; Koscielniak, D.; Papiez, M.; Vyhouskaya, P.; Zagorska-Swiezy, K.; Kolodziej, I.; Bystrowska, B.; Jurczak, A. Effect of a Lactobacillus salivarius Probiotic on a Double-Species Streptococcus mutans and Candida albicans Caries Biofilm. Nutrients 2017, 9, 1242. [Google Scholar] [CrossRef]
- Jiang, Q.R.; Stamatova, I.; Kainulainen, V.; Korpela, R.; Meurman, J.H. Interactions between Lactobacillus rhamnosus GG and oral micro-organisms in an in vitro biofilm model. BMC Microbiol. 2016, 16, 149. [Google Scholar] [CrossRef]
- Zeng, Y.; Fadaak, A.; Alomeir, N.; Wu, T.T.; Rustchenko, E.; Qing, S.; Bao, J.; Gilbert, C.; Xiao, J. Lactobacillus plantarum Disrupts S. mutans–C. albicans Cross-Kingdom Biofilms. Front. Cell Infect. Microbiol. 2022, 12, 872012. [Google Scholar] [CrossRef] [PubMed]
- Arvola, T.; Laiho, K.; Torkkeli, S.; Mykkanen, H.; Salminen, S.; Maunula, L.; Isolauri, E. Prophylactic Lactobacillus GG reduces antibiotic-associated diarrhea in children with respiratory infections: A randomized study. Pediatrics 1999, 104, e64. [Google Scholar] [CrossRef] [PubMed]
- Kelesidis, T.; Pothoulakis, C. Efficacy and safety of the probiotic Saccharomyces boulardii for the prevention and therapy of gastrointestinal disorders. Ther. Adv. Gastroenterol. 2012, 5, 111–125. [Google Scholar] [CrossRef]
- D’Haens, G.R.; Jobin, C. Fecal Microbial Transplantation for Diseases Beyond Recurrent Clostridium Difficile Infection. Gastroenterology 2019, 157, 624–636. [Google Scholar] [CrossRef] [PubMed]
- Iannitti, T.; Palmieri, B. Therapeutical use of probiotic formulations in clinical practice. Clin. Nutr. 2010, 29, 701–725. [Google Scholar] [CrossRef]
- Garcia-Gonzalez, N.; Battista, N.; Prete, R.; Corsetti, A. Health-Promoting Role of Lactiplantibacillus plantarum Isolated from Fermented Foods. Microorganisms 2021, 9, 349. [Google Scholar] [CrossRef]
- Sales-Campos, H.; Soares, S.C.; Oliveira, C.J.F. An introduction of the role of probiotics in human infections and autoimmune diseases. Crit. Rev. Microbiol. 2019, 45, 413–432. [Google Scholar] [CrossRef]
- Caglar, E.; Kargul, B.; Tanboga, I. Bacteriotherapy and probiotics’ role on oral health. Oral Dis. 2005, 11, 131–137. [Google Scholar] [CrossRef]
- Meurman, J.H. Probiotics: Do they have a role in oral medicine and dentistry? Eur. J. Oral Sci. 2005, 113, 188–196. [Google Scholar] [CrossRef]
- Cagetti, M.G.; Mastroberardino, S.; Milia, E.; Cocco, F.; Lingstrom, P.; Campus, G. The Use of Probiotic Strains in Caries Prevention: A Systematic Review. Nutrients 2013, 5, 2530–2550. [Google Scholar] [CrossRef]
- Lin, T.H.; Lin, C.H.; Pan, T.M. The implication of probiotics in the prevention of dental caries. Appl. Microbiol. Biotechnol. 2018, 102, 577–586. [Google Scholar] [CrossRef] [PubMed]
- Nase, L.; Hatakka, K.; Savilahti, E.; Saxelin, M.; Ponka, A.; Poussa, T.; Korpela, R.; Meurman, J.H. Effect of long-term consumption of a probiotic bacterium, Lactobacillus rhamnosus GG, in milk on dental caries and caries risk in children. Caries Res. 2001, 35, 412–420. [Google Scholar] [CrossRef] [PubMed]
- Laleman, I.; Detailleur, V.; Slot, D.E.; Slomka, V.; Quirynen, M.; Teughels, W. Probiotics reduce mutans streptococci counts in humans: A systematic review and meta-analysis. Clin. Oral Investig. 2014, 18, 1539–1552. [Google Scholar] [CrossRef] [PubMed]
- Wasfi, R.; Abd El-Rahman, O.A.; Zafer, M.M.; Ashour, H.M. Probiotic Lactobacillus sp. inhibit growth, biofilm formation and gene expression of caries-inducing Streptococcus mutans. J. Cell Mol. Med. 2018, 22, 1972–1983. [Google Scholar] [CrossRef]
- Kraft-Bodi, E.; Jorgensen, M.R.; Keller, M.K.; Kragelund, C.; Twetman, S. Effect of Probiotic Bacteria on Oral Candida in Frail Elderly. J. Dent. Res. 2015, 94, 181S–186S. [Google Scholar] [CrossRef] [PubMed]
- Matsubara, V.H.; Wang, Y.; Bandara, H.; Mayer, M.P.A.; Samaranayake, L.P. Probiotic lactobacilli inhibit early stages of Candida albicans biofilm development by reducing their growth, cell adhesion, and filamentation. Appl. Microbiol. Biotechnol. 2016, 100, 6415–6426. [Google Scholar] [CrossRef] [PubMed]
- Rossoni, R.D.; de Barros, P.P.; de Alvarenga, J.A.; Ribeiro, F.C.; Velloso, M.D.S.; Fuchs, B.B.; Mylonakis, E.; Jorge, A.O.C.; Junqueira, J.C. Antifungal activity of clinical Lactobacillus strains against Candida albicans biofilms: Identification of potential probiotic candidates to prevent oral candidiasis. Biofouling 2018, 34, 212–225. [Google Scholar] [CrossRef]
- Al-Shami, I.Z.; Al-Hamzi, M.A.; Al-Shamahy, H.A.; Majeed, A.L.A.A. Efficacy of some Antibiotics against Streptococcus Mutans Associated with Tooth decay in Children and their Mothers. Online J. Dent. Oral Health 2019, 2. [Google Scholar]
- Pasquantonio, G.; Condò, S.; Cerroni, L.; Bikiqu, L.; Nicoletti, M.; Prenna, M.; Ripa, S. Antibacterial activity of various antibiotics against oral streptococci isolated in the oral cavity. Int. J. Immunopathol. Pharmacol. 2012, 25, 805–809. [Google Scholar] [CrossRef]
- Jahanshiri, Z.; Manifar, S.; Moosa, H.; Asghari-Paskiabi, F.; Mahmoodzadeh, H.; Shams-Ghahfarokhi, M.; Razzaghi-Abyaneh, M. Oropharyngeal candidiasis in head and neck cancer patients in Iran: Species identification, antifungal susceptibility and pathogenic characterization. J. Mycol. Med. 2018, 28, 361–366. [Google Scholar] [CrossRef]
- Aitken-Saavedra, J.; Lund, R.G.; González, J.; Huenchunao, R.; Perez-Vallespir, I.; Morales-Bozo, I.; Urzúa, B.; Tarquinio, S.C.; Maturana-Ramírez, A.; Martos, J. Diversity, frequency and antifungal resistance of Candida species in patients with type 2 diabetes mellitus. Acta Odontol. Scand. 2018, 76, 580–586. [Google Scholar] [CrossRef]
- Chandra, J.; Kuhn, D.M.; Mukherjee, P.K.; Hoyer, L.L.; McCormick, T.; Ghannoum, M.A. Biofilm formation by the fungal pathogen Candida albicans: Development, architecture, and drug resistance. J. Bacteriol. 2001, 183, 5385–5394. [Google Scholar] [CrossRef]
- Kim, D.; Liu, Y.; Benhamou, R.I.; Sanchez, H.; Simon-Soro, A.; Li, Y.; Hwang, G.; Fridman, M.; Andes, D.R.; Koo, H. Bacterial-derived exopolysaccharides enhance antifungal drug tolerance in a cross-kingdom oral biofilm. ISME J. 2018, 12, 1427–1442. [Google Scholar] [CrossRef]
- Xiao, J.; Moon, Y.; Li, L.; Rustchenko, E.; Wakabayashi, H.; Zhao, X.; Feng, C.; Gill, S.R.; McLaren, S.; Malmstrom, H.; et al. Candida albicans Carriage in Children with Severe Early Childhood Caries (S-ECC) and Maternal Relatedness. PLoS ONE 2016, 11, e0164242. [Google Scholar] [CrossRef]
- Marsh, P.D. Are dental diseases examples of ecological catastrophes? Microbiology 2003, 149, 279–294. [Google Scholar] [CrossRef]
- Koo, H.; Xiao, J.; Klein, M.I.; Jeon, J.G. Exopolysaccharides produced by Streptococcus mutans glucosyltransferases modulate the establishment of microcolonies within multispecies biofilms. J. Bacteriol. 2010, 192, 3024–3032. [Google Scholar] [CrossRef]
- Xiao, J.; Hara, A.T.; Kim, D.; Zero, D.T.; Koo, H.; Hwang, G. Biofilm three-dimensional architecture influences in situ pH distribution pattern on the human enamel surface. Int. J. Oral Sci. 2017, 9, 74–79. [Google Scholar] [CrossRef]
- Xiao, J.; Klein, M.I.; Falsetta, M.L.; Lu, B.; Delahunty, C.M.; Yates, J.R., 3rd; Heydorn, A.; Koo, H. The exopolysaccharide matrix modulates the interaction between 3D architecture and virulence of a mixed-species oral biofilm. PLoS Pathog. 2012, 8, e1002623. [Google Scholar] [CrossRef]
- Xiao, J.; Zeng, Y.; Rustchenko, E.; Huang, X.; Wu, T.T.; Falsetta, M.L. Dual transcriptome of Streptococcus mutans and Candida albicans interplay in biofilms. J. Oral Microbiol. 2023, 15, 2144047. [Google Scholar] [CrossRef]
- Alomeir, N.; Zeng, Y.; Fadaak, A.; Wu, T.T.; Malmstrom, H.; Xiao, J. Effect of Nystatin on Candida albicans–Streptococcus mutans duo-species biofilms. Arch. Oral Biol. 2023, 145, 105582. [Google Scholar] [CrossRef]
- Guggenheim, B.; Giertsen, E.; Schupbach, P.; Shapiro, S. Validation of an in vitro biofilm model of supragingival plaque. J. Dent. Res. 2001, 80, 363–370. [Google Scholar] [CrossRef]
- Xiao, J.; Koo, H. Structural organization and dynamics of exopolysaccharide matrix and microcolonies formation by Streptococcus mutans in biofilms. J. Appl. Microbiol. 2010, 108, 2103–2113. [Google Scholar] [CrossRef]
- Klein, M.I.; Xiao, J.; Heydorn, A.; Koo, H. An analytical tool-box for comprehensive biochemical, structural and transcriptome evaluation of oral biofilms mediated by mutans streptococci. J. Vis. Exp. 2011, 2512. [Google Scholar] [CrossRef]
- Sabo, S.D.; Vitolo, M.; Gonzalez, J.M.D.; Oliveira, R.P.D. Overview of Lactobacillus plantarum as a promising bacteriocin producer among lactic acid bacteria. Food Res. Int. 2014, 64, 527–536. [Google Scholar] [CrossRef]
- Zeng, L.; Burne, R.A. Comprehensive mutational analysis of sucrose-metabolizing pathways in Streptococcus mutans reveals novel roles for the sucrose phosphotransferase system permease. J. Bacteriol. 2013, 195, 833–843. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Mean (SD) or % |
|---|---|
| Age (Year) | 3.5 ± 1.0 |
| Gender (Male) | 50% |
| Race: Caucasian | 60% |
| African American | 30% |
| Asian | 10% |
| Ethnicity: Hispanic | 10% |
| Brushing Frequency (Daily) | 90% |
| Attending Daycare (Yes) | 20% |
| Plaque Index | 1.8 ± 0.6 |
| dt | 11.7 ± 5.1 |
| mt | 0.2 ± 0.6 |
| ft | 0.1 ± 0.3 |
| dmft | 12 ± 4.9 |
| ds | 27.2 ± 17.4 |
| ms | 1.0 ± 3.2 |
| fs | 0.1 ± 0.3 |
| dmfs | 28.3 ± 16.5 |
| Microcolony Parameters | Control (n = 30) | With L. plantarum 14917 (n = 30) |
|---|---|---|
| Number of attached microcolonies | 15.3 ± 4.1 | 9.2 ± 6.7 ** |
| Area of attached microcolonies (um2) | 972.2 ± 924.4 | 283.6 ± 84.3 ** |
| Volume of attached microcolonies (um3) × 103 | 3471.5 ± 2334.8 | 25.6 ± 27.5 *** |
| Number of free microcolonies | 287.4 ± 71.1 | 213.3 ± 77.5 *** |
| Diameter of free microcolonies (um) | 38.0 ± 4.3 | 28.4 ± 3.0 *** |
| Volume of free microcolonies (um3) × 103 | 925.0 ± 255.0 | 145.8 ± 111.8 *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, Y.; Fadaak, A.; Alomeir, N.; Wu, Y.; Wu, T.T.; Qing, S.; Xiao, J. Effect of Probiotic Lactobacillus plantarum on Streptococcus mutans and Candida albicans Clinical Isolates from Children with Early Childhood Caries. Int. J. Mol. Sci. 2023, 24, 2991. https://doi.org/10.3390/ijms24032991
Zeng Y, Fadaak A, Alomeir N, Wu Y, Wu TT, Qing S, Xiao J. Effect of Probiotic Lactobacillus plantarum on Streptococcus mutans and Candida albicans Clinical Isolates from Children with Early Childhood Caries. International Journal of Molecular Sciences. 2023; 24(3):2991. https://doi.org/10.3390/ijms24032991
Chicago/Turabian StyleZeng, Yan, Ahmed Fadaak, Nora Alomeir, Yan Wu, Tong Tong Wu, Shuang Qing, and Jin Xiao. 2023. "Effect of Probiotic Lactobacillus plantarum on Streptococcus mutans and Candida albicans Clinical Isolates from Children with Early Childhood Caries" International Journal of Molecular Sciences 24, no. 3: 2991. https://doi.org/10.3390/ijms24032991
APA StyleZeng, Y., Fadaak, A., Alomeir, N., Wu, Y., Wu, T. T., Qing, S., & Xiao, J. (2023). Effect of Probiotic Lactobacillus plantarum on Streptococcus mutans and Candida albicans Clinical Isolates from Children with Early Childhood Caries. International Journal of Molecular Sciences, 24(3), 2991. https://doi.org/10.3390/ijms24032991