Abstract
Low back pain is an extremely common condition with severe consequences. Among its potential specific causes, degenerative disc disease (DDD) is one of the most frequently observed. Mechanobiology is an emerging science studying the interplay between mechanical stimuli and the biological behavior of cells and tissues. The aim of the presented study is to review, with a systematic approach, the existing literature regarding the mechanobiology of the human intervertebral disc (IVD), define the main pathways involved in DDD and identify novel potential therapeutic targets. The review was carried out in accordance with the Preferential Reporting Items for Systematic Reviews and Meta-analyses (PRISMA) guidelines. Studies were included if they described biological responses of human IVD cells under mechanical stimulation or alterations of mechanical properties of the IVD determined by different gene expression. Fifteen studies were included and showed promising results confirming the mechanobiology of the human IVD as a key element in DDD. The technical advances of the last decade have allowed us to increase our understanding of this topic, enabling us to identify possible therapeutic targets to treat and to prevent DDD. Further research and technological innovations will shed light on the interactions between the mechanics and biology of the human IVD.
1. Introduction
Low back pain (LBP) is an extremely common condition worldwide, with an expected lifetime-prevalence rate of 80% [1] and severe socioeconomic and health care consequences [2]. Among its potential specific causes, degenerative disc disease (DDD) is one of the most frequently observed [3]. On a morphological level, DDD results in dehydration, fissuring and tearing of the nucleus pulposus (NP), annulus fibrosus (AF) and endplate [4].
However, as opposed to being a passive process of wear and tear, disc degeneration is an abnormal, cell-mediated response to structural failure brought on by aging and other environmental variables including abnormal mechanical stress. In fact, substantial degenerative changes also happen at a biological level. A decrease in the diffusion of oxygen, nutrients and waste products through the disc space is seen [5], with a consequent shift towards an anaerobic metabolism, with increased lactate production and a decrease in pH level. The intervertebral disc (IVD) pH ranges from 7.1 in healthy tissue to 6.8 in degenerated tissue, with values reaching pH 5.5 in severely degenerated discs [6,7]. Concerning physiological loading forces acting within human lumbar IVDs, the values range.
Between 0.5 and 1.7 MPa during relaxed standing and lifting a 20 Kg weight with flexed knees, respectively. Concerning the strain stress of human annulus fibrosus in vivo, the values range between 4% and 6% when discs are in compression or torsion and flexion or extension, respectively [8,9,10].
Alongside this, while the normal IVD is avascular and aneural, micronervous [11] and microvascular [12,13] ingrowth is seen in degenerated discs. Moreover, increased cellular apoptosis [14], cellular senescence [15], cell density [16] and the substitution of notochordal cells of the NP with chondrocyte-like cells [17] occur.
Furthermore, substantial changes are seen in the extracellular matrix (ECM). A quantitative decline in the proteoglycan (PG) content occurs, as well as a deterioration of qualitative ECM composition, with decreased aggrecan content and a change in the relative proportions of the glycosaminoglycans in favor of keratan sulfate over chondroitin sulfate [18]. As a result, less water molecules can be bonded by the matrix. Collagen production patterns and compositions within the degenerated disc change: an increase in collagen I appears in the NP and in the inner AF (in which collagen II is normally predominant), while an increase in collagen II appears in the outer AF (in which collagen I is normally predominant).
Finally, degenerated IVDs show a pro-inflammatory microenvironment, as the expression of IL-1β, TNF-α [19,20], Nitric Oxide (NO), prostaglandin E2, IL-6 and matrix metalloproteinases (MMPs) [21,22] are significantly increased compared to controls. Recent studies further demonstrated that axial loading and mechanical stress correlated with CGRP and mPGES-1 [23]. On the first instance, this pro-inflammatory microenvironment plays a crucial role as a “discogenic pain” generator [24]. Secondly, inflammation contributes to the increased degradation of the ECM and to PG loss from the NP, with a consequent decrease in weight-bearing capacity and, eventually, leading to the aforementioned structural degenerative changes, thus establishing a vicious circle.
Mechanobiology is an emerging science studying the interplay between mechanical stimuli and the biological behavior of cells and tissues. The human spine is a complex biomechanical model constantly subjected to mechanical loading, and the IVD is the keystone of spine physiology. Given the important effect that mechanical loadings have on the IVD, it is crucial to understand the molecular pathways that, from loading, lead to DDD, in order to prevent and treat disc degeneration. There is a growing body of literature focusing on IVD mechanobiology; however, most of the studies just focus on one single molecular pathway or on a single form of mechanical stress. Some literature reviews [25,26,27,28,29] were conducted to harmonize the results achieved by the most important mechanobiological studies on IV, but none are systematic, with a consequent increased risk of selection bias. Moreover, many of the existing reviews included both human and animal models, providing a possible additional source of bias, since there are substantial differences between the physiology and the forces acting on animal and human IVDs, given their erect posture. Hence, the aim of the presented study is to review, with a systematic approach, the existing literature regarding the mechanobiology of the human IVD, defining the main pathways involved in DDD to identify novel potential therapeutic targets.
2. Material and Methods
2.1. Review Design
A systematic review of the literature regarding the mechanobiology of the human intervertebral disc was carried out following the Preferential Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) guidelines [30].
Only peer-reviewed publications were considered. Studies were included if they described biological responses of human IVD cells under mechanical stimulation or alterations of mechanical properties of the intervertebral disc determined by different gene expressions. Only articles in English were included. Articles that analyzed samples retrieved from animals or both animals and humans were excluded.
The included articles met the PICO criteria for systematic reviews (Population, Intervention, Comparison, and Outcomes).
2.2. Search Strategy
Eligible studies were searched in Pubmed-MEDLINE, Embase Biomedical Database, Google Scholar, and The Cochrane Central Registry of Controlled Trials between 1990 and 2022. Three reviewers (G.V., M.T. and M.M.) performed the literature search in October 2022.
The search string used included the following items: mechanobiology OR mechanotransduction OR (mechanics OR overload OR load) AND (genes OR gene OR genome OR epigenetic OR genetic OR pathway) AND (“intervertebral disc” OR “nucleus pulpous” OR “annulus fibrosus” OR “intervertebral discs” OR “intervertebral disc degeneration” OR “degenerative disc disease” OR “IVD” OR “DDD”).
2.3. Study Selection
Titles and abstracts were screened, and then, relevant articles obtained via an electronic search were retrieved in full-text, followed by a hand check of their bibliography in order to recover additional related articles. This search was extended to reviews and meta-analyses for the identification of potentially missed eligible papers. Duplicates were subsequently removed. In Figure 1, the selection process carried out in accordance with the PRISMA flowchart is represented. This systematic review was registered in the PROSPERO database for systematic reviews [31] (ID: CRD42022371280).

Figure 1.
Prisma 2009 flow diagram of the included studies.
2.4. Data Extraction
Two reviewers (G.V. and M.T.) extracted the data through a standardized data collection form. Three reviewers (G.V., M.T. and S.N.) checked the data for accuracy, and inconsistent results were analyzed for discussion. The extracted data concerning the study design, number of samples, demographics of patients, type of IVD cells considered for analysis, model of mechanical stimulation, cellular response to mechanical stimulation and results are summarized in Table 1.

Table 1.
Details of the included studies. ↑ Increase of expression, ↓ decrease of expression, ↑↑ high increase of expression, ↓↓ high decrease of expression, ⊗ inhibition.
3. Results
3.1. Included Studies
A total of 213 studies were found through the electronic search, according to the research performed. A total of 15 papers [32,33,34,35,37,38,39,40,41,42,43,44,45,46] met the inclusion criteria and were considered for review. All of the included studies were in vitro or ex vivo studies.
The following studies were not included in the final analysis: research using animal models, finite element analyses, studies that did not evaluate mechanical stimuli on samples and review articles.
The studies analyzed mainly small-sized groups of samples (n = 3 to 30), describing the relationship between mechanical stimuli and the biology of human IVD cells and the mechanical properties of human IVD cells under different biological conditions.The included studies were largely heterogeneous in terms of patients’ characteristics, the degeneration degree of different samples, gender, the type of applied mechanical stimuli, the analysis technique and the analyzed molecular pathway (Table 1).
3.2. Cohort Characteristics
The included studies reported data on a total of 212 IVD samples derived from 126 patients who underwent spinal surgery for DDD, 38 patients who underwent spinal surgery for idiopathic scoliosis, 14 patients who underwent spinal surgery for spinal cord injury and 30 cadavers. One study did not report accurate information about the sample origin [37].
The studies included 62 males (58%) and 45 females (42%). Seven studies did not report the patients’ sex [33,37,40,41,42,43,44]. The mean age of the patients ranged from 56.2 to 15.2 years. No study reported accurate information about patients’ ethnicity.
3.3. Evaluated Cell Types and Molecular Pathways
Six studies evaluated the biological response of NP cells isolated from IVD components and cultured before mechanical stimulation [32,35,36,37,44,46]. Five studies evaluated the biological response of AF cells isolated from IVD components and cultured before mechanical stimulation [34,37,38,39,41]. Three studies evaluated the biological response of both isolated NP and AF cells to mechanical stimulation [34,38,45]. Sun et al. [33] evaluated the anti-angiogenic role of exosomes obtained via the in vitro mechanical stimulation of isolated and cultured notochordal cells. Aladin et al. [45] analyzed the relationship between the mechanical properties of frozen NP tissue explants and type IX collagen variant typing.
As for molecular pathways, most of the studies analyzed the expression of catabolic and anabolic genes in IVD cells under different types and conditions of mechanical stimulation [34,35,36,41,42,43,44,45,46]. Two studies focused on the role of ion channels in mechanotransduction [38,39]. Song et al. [32] evaluated the role of the RhoA/ROCK2–MRTF-A signaling pathway in the development of the IDD. Sun et al. [33] evaluated the role of the Wnt/b-catenin axis in the inhibition of miR-140-5p. Likhitpanichktul et al. [37] investigated the interaction between TNF-α and tensile strain in IVD degeneration. Some of the key findings are summarized in Figure 2.
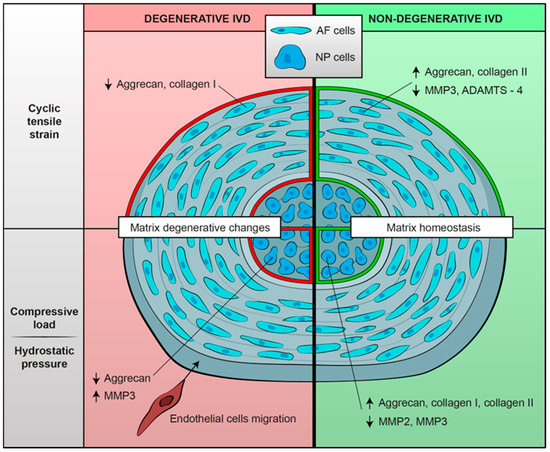
Figure 2.
Summary of biological mechanoresponses of human intervertebral cells, either degenerative or non-degenerative, to different mechanical stimuli in terms of extracellular matrix changes. Mechanical load seems to determine a shift towards an anabolic phenotype in non-degenerated IVD cells; on the contrary, degenerated IVD cells exposed to mechanical stimulation show matrix catabolic changes.
3.4. Applied Mechanical Stimuli and Cell Environmental Conditions
The included studies used different types of mechanical stimulation in order to produce a biological response in different types of cells. It is noteworthy that in a physiological situation (in vivo), IVD cells were subjected to different loads depending on their anatomical position, with NP and inner AF cells being mainly subjected to compressive loading, while outer AF cells were subjected to strain forces [47]. Most of the studies applied cyclic tensile strain [32,34,39,40,42,43,45]. Studies focusing on NP cells mainly applied compressive load [33,36,37,44,46]. Few studies exposed cells to hydrostatic pressure [34,35,45]. Franco-Obregon et al. simulated microgravity using a random positioning machine and analyzed isolated and cultured IVD cells’ biological responses [35].
3.4.1. Cyclic Tensile Strain (CTS)
Authors applied different strain protocols to study cellular responses (Table 2). Most of the experiments used a strain frequency of 1 Hz, which is similar to physiological human motion [34,39,41,42,43,45,47]. The strain magnitude varied, ranging from 1% [46] to 20% [39,43].
Table 2.
Key findings of the included studies.
Neidlinger-Wilke at al. [46] found that CTS has an anabolic effect on AF cells, with the increased gene expression of aggrecan and collagen II and decreased MMP-3 gene expression. No difference was found by applying different strain magnitudes (1, 2, 4 and 8% strain). Mechanically stimulated cells tended to decrease the mRNA expression of MMP-2 and MMP-3. It is important to notice that cells obtained by different donors showed high interindividual variations in cellular responses to mechanical stimuli, suggesting that different degrees of disc degeneration could affect mechanotransduction pathways in a similar manner.
Gilbert et al. [41] investigated the effect of CTS on AF cells derived either from degenerated or healthy IVDs. Interestingly, the response appeared to be frequency-dependent and strongly dependent on the degree of disc degeneration. At 1 Hz CTS, AF cells derived from non-degenerative IVDs showed a shift to a less catabolic phenotype, decreasing ADAMTS-4 and MMP-3 expression. Conversely, AF cells exposed to 0.33 Hz CTS showed a catabolic response. AF cells derived from degenerative tissues appeared to have lost their ability to reduce metalloproteinase gene expression and also decreased matrix protein gene expression (aggrecan and collagen I). Subsequently, Gilbert et al. [39] observed that the reduction in the catabolic response in non-degenerative AF cells exposed to 1 Hz CTS may be IL-1- and IL-4-dependent. In AF cells derived from degenerative IVDs, neither IL-1 nor IL-4 showed a significant role in the matrix remodeling mechanoresponse, confirming their loss of response to stimuli.
AF cells derived from degenerative and non-degenerative tissue display strong differences in the regulation of matrix protein and matrix-degrading enzyme gene expression. Those differences could be attributed to mechanotransduction pathways altered in degenerated tissues [38,39,41]. Integrins have been shown to play a pivotal role in the mechanoresponse of AF cells to CTS [38]. In non-degenerative AF cells exposed to 1 Hz 10% CTS, the addition of an integrin-recognizing RAD peptide determined the upregulation of ADAMTS-4 gene expression and increased FAK phosphorylation. The addition of integrin-function-blocking peptide to non-degenerative AF cells exposed to 1 Hz 10% CTS was able to inhibit CTS-induced FAK phosphorylation and to upregulate ADAMTS-4 gene expression. AF cells derived from degenerative IVDs did not show integrin-dependent differences in mechanoresponses. These findings suggest that the involvement of RGD integrins in mechanoresponse is lost with degeneration; thus, integrins may not be involved in degenerative AF cells’ altered mechanoresponse, which seems to happen via an alternative mechanotransduction pathway.
Likhitpanichkul et al. [37] further evaluated the role of inflammation in matrix homeostasis and the mechanoresponse of AF cells subjected to CTS, showing that the concomitant exposure of AF cells to TNF-α and pathological strains determined increased IL-1β and IL-8 production and a modified AF cytoskeleton. These results suggested that TNF-α induced cytoskeletal changes in AF cells, suggestive of enhanced mechanosensitivity, having a detrimental effect on IVD matrix homeostasis.
Cambria et al. [34] investigated TRPV4 ion channel signaling in stretch-induced AF cells’ inflammation. In addition, they established a novel model of acute pro-inflammatory response to hyperphysiological stretching typical for early-stage AF injury (increased gene expression of IL-6, IL-8, COX2 and MMP-1 and reduced expression of collagen II and aggrecan). These findings showed that TRPV4 pharmacological inhibition and TRPV4 gene knock-out via CRISPR-Cas9 gene editing effectively mitigated the inflammation caused by hyperphysiological cyclic stretching, thus demonstrating that TRPV4 may play a role in stretch-induced inflammation, possibly via the activation of the p38 MAPK pathway.
Unlike other studies, Song et al. [32] evaluated NP cells exposed to cyclic strain, focusing specifically on the RhoA/MRTF-A signaling pathway and its role in DDD development. They applied CTS with or without an MRTF-A inhibitor and observed that the RhoA/MRTF-A signaling pathway is of key importance in NP cells’ strain-induced matrix degeneration. MRTF-A inhibition may alleviate the fibrosis caused by mechanical overload, with a potential therapeutic effect.
3.4.2. Compressive Load
The differences in mechanoresponses between degenerative and non-degenerative IVD cells have been further investigated by Le Maitre et al. [43]. The compressive load of degenerative and non-degenerative NP cells seeded in alginate constructs appeared to have subtle differences in terms of mechanoresponse, determining decreased aggrecan expression. When exposed to an RGD integrin ligand site inhibitor, degenerative NP cells continued to show decreased aggrecan expression, while non-degenerative NP cells under the same loading conditions did not show load-induced aggrecan downregulation, revealing an integrin-mediated anti-catabolic effect. Thus, the differences in cellular response to mechanical load between degenerative and non-degenerative cells may be due to the altered mechanotransduction pathway. In non-degenerative NP cells, mechanotransduction seems to occur via integrin receptors, while degenerative NP cells might use different receptors.
Huang et al. [40] showed that a dynamic compressive load applied to NP tissue determined matrix enzymes’ (metalloproteinases and ADAMTSs) upregulation, leading to a decrease in aggrecan content in the NP cell matrix, therefore being an etiological factor in the development of DDD. Interestingly, ADAMTS-1, 4, 9, 15, and TIMP-3 are also physiologically expressed in healthy disc matrixes, suggesting that these proteins may play a role in regular IVD matrix turnover [40]. TIMP-3 demonstrated the potential capability to prevent matrix catabolism and to determine an increase in aggrecan expression [40].
In order to better understand the importance of the cell microenvironment, Hodson et al. [36] tested NP cells embedded in agarose constructs by applying cycling compressive loads under different pH conditions, representative of degenerative (acidic pH) and non-degenerative (neutral pH) IVD microenvironments. The compression response of NP cells at a pH representative of degenerate tissue (pH 6.5) was represented by a metabolism shift to matrix catabolism. The progression of IVD degeneration may therefore recognize acidity-induced aberrant mechanotransduction as a potential pathologic mechanism. In addition, Hodson et al. investigated whether mechanotransduction might be integrin-dependent under various pH settings, finding that aggrecan expression was modulated by integrins at a pH representative of non-degenerative IVD, while the mechanoresponsiveness of aggrecan was aborted at an acidic pH [36].
The potential role of notochordal cells’ products obtained by notochordal cells’ compression in preventing IVD angiogenesis was evaluated by Sun et al. [33]. Exosomes derived from compressed notochordal cells showed a strong inhibitive angiogenesis effect, demonstrating that exosomes are effective carriers in transferring miRNA from cell to cell. The anti-angiogenetic effect seems to occur through the inhibition of the Wnt/b-catenin pathway.
3.4.3. Hydrostatic Pressure
Consistent with aforementioned results, Neidlinger-Wilke et al. [46] also showed that hydrostatic pressure, if applied in protocols similar to physiological loads, determines a shift to a less catabolic metabolism of both NP and AF cells embedded in collagen gel.
These findings might suggest that moderate mechanical loading can prevent disc matrix degradation by inhibiting MMPs’ expression.
Most importantly, for the first time, it has been assumed that the strong interindividual variations in gene expression among donors could be due to the degree of disc degeneration.
Subsequent studies [44] showed that NP cells derived from healthy IVD tissue exposed to dynamic hydrostatic pressure responded by upregulating the gene expression of anabolic genes, while NP cells derived from degenerative IVDs exposed to the same stress did not show any effect on gene expression.
3.4.4. Microgravity
To elucidate the effect of reduced load on IVD cells, Franco-Obregon et al. [35] simulated microgravity using a random positioning machine and tested the mechanoresponse of both AF and NP cells in the presence or absence of TRPC6 channel pharmacological inhibition. Surprisingly, cells subjected to simulated microgravity or to TRPC channel inhibition showed reduced proliferation and increased senescence.
4. Discussion
In the present work, we systematically reviewed the available literature from 1990 to the present date regarding the mechanobiology of the human IVD. Fifteen papers met the inclusion criteria, exploring multiple aspects of the mechanobiology of the human IVD and showing promising results in terms of the definition of molecular pathways potentially involved in the development of DDD. Many of the included studies evaluated the mechanical response of IVD cells to different mechanical stimuli in terms of matrix homeostasis. In detail, Neidlinger-Wilke [46] demonstrated that moderate mechanical stimulation, both by HP and CTS, determines an anabolic shift of the matrix homeostasis, with some differences between the NP cells (more responsive to HP) and the AF cells (more responsive to CTS). As both hyperphysiological stresses and mechanical unloading are reported to determine a catabolic shift in the IVD [40,43,48], moderate mechanical loading may be important in preventing disc matrix degradation The clinical reflection of this is that moderate exercise may be have a protective role against disc degeneration [49]. However, this does not seem to be true for both degenerated and non-degenerated IVD. In fact, physiological HP only seems to increase the expression of collagen II and aggrecan in healthy NP cells and not in degenerated ones [44]; on the other hand, the application of 20% CTS at 1 Hz on degenerated AF cells not only fails to downregulate matrix proteases’ gene expression, but it also results in reduced aggrecan and collagen I gene expression [41]. In this light, in a clinical view, moderate physical exercise on extracellular matrix metabolism in a degenerated disc may have detrimental consequences, although this requires further research.
The reasons for these different biological responses to mechanical stimulation between degenerated and healthy discs have long been researched. Some of the included studies [36,41,46] have demonstrated that RGD-recognizing integrins lose their role in the mechanoresponse of human NP and AF cells derived from degenerated IVD compared to non-degenerated IVD cells. Since both degenerated and non-degenerated IVD cells express RGD-binding integrins, these data suggest that, with DDD, either the RGD integrin transduction is altered or that mechanosensing could occur through a non-RGD integrin. Local inflammation has also been demonstrated to be crucial in disc mechanobiology. In fact, while non-degenerated AF cells subjected to CTS showed a reduced catabolic response which appeared to be IL-1- and IL-4-dependent, in degenerated AF cells, neither IL-1 nor IL-4 appeared to be necessary for the matrix remodeling mechanoresponse [39]. Moreover, an interesting finding is that local inflammation mediated by TNF-α induces deep cytoskeletal changes, enhancing the mechanosensitivity of cells and thus creating a vicious mechanoinflammatory system at the intracellular level [37]. This inflammatory microenvironment is critical not only in the vicious loop that leads to DDD, but also in the genesis of LBP. In fact, LBP is an extremely heterogeneous symptom, and it is unclear why some degenerative discs are responsible for LBP while others are not. However, nerve in-growth in the NP induced by inflammatory cytokines [50,51] undoubtedly plays a role in the development of discogenic pain.
The relationship between mechanical stimuli, inflammation and the progression of degenerative disc disease is still under investigation. Unfortunately, many studies on this topic involved animal models and were therefore excluded. We hope that further studies will be conducted in the future.
Cambria et al. [34] demonstrated that part of this mechanoinflammation in AF cells is signaled through TPRV4 ion channels, making them a possible future pharmacological or gene-editing therapeutic target. Furthermore, other potential therapeutic targets have been identified by some of the included studies. Song et al. [32] demonstrated that the RhoA/MRTF-A inhibitor CCG-1423 can play a potential role as a therapeutic agent, since it is able to stop a signaling pathway leading to ECM degradation in response to NP cells’ mechanical overload. Sun et al. [33] demonstrated that miR-140-5p, contained in NC-exos induced by compressive load, had a significant inhibitive effect on angiogenesis in endothelial cells. This may have a therapeutic role, since mechanical load seems to have a proangiogenetic effect in the IVD [42]. Finally, Huang et al. [40] suggested that TIMP-3, an ADAMT inhibitor, may have a potential role in the prevention of aggrecan loss during DDD.
It is important to note a significant trend towards increasing complexity in study designs and techniques employed in more recent studies compared to older studies. For example, Neidlinger-Wilke [46] and Le Maitre [44] mainly conducted gene expression studies based on classical semi-quantitative RT-PCR, while Song [32] and Sun [33] reached interesting results thanks to the adoption of modern biotechnologies such as CRISPR-Cas9 genome editing and exosome characterization, respectively.
To the best of our knowledge, this is the first systematic scoping review only focusing on the mechanobiology of human IVD. This is crucial, since the results of animal mechanobiological models of IVD may not be transposed to human IVD cells [52]. In fact, the vast majority of animal models involved quadrupeds, whose spine develops in an extremely different biomechanical environment compared to the human spine, which is almost constantly exposed to the action of gravity after the assumption of an erect posture during childhood. The presented study does not come without limitations. Firstly, high heterogeneity among the included studies was registered, especially in terms of the type of cells (AF and/or NP), type of mechanical stimulation (cyclic tensile strain, hydrostatic pressure, compressive load and simulated microgravity), mechanical stimulation protocol (intensity, frequency and duration of the stimuli that are known as crucial factors in determining biochemical cell response) and type of culture environment (2 or 3D). Secondly, many studies adopted disc samples from AIS patients [33,37,44,45] or cadavers [42,46] as controls, while both the categories may show unrecognized subtle alterations in the IVD (the first, due to the underlying condition; the second, due to advanced age). In addition, only one [32] of the included studies reported data regarding systemic diseases (cardiovascular, pulmonary or neoplastic). Finally, all the included studies were based upon in vitro (almost all) or ex vivo models, which are useful in order to explore cellular mechanisms, but their findings need to be validated at the IVD organ scale and in vivo, since cellular phenotype significantly changes during in vitro culture compared to the in vivo one.
Further in vitro studies, with a more standardized stimulation model, as well as organ-scale and in vivo studies, are needed in the forthcoming years to confirm the role of the promising pathways that have been identified to have a potential role in the mechanobiology of DDD. Alongside that, future preclinical and clinical studies investigating the effects of the targeted pharmacological inhibition of these mechanobiological mechanisms may open up promising therapeutical options for the prevention and treatment of DDD.
5. Conclusions
The mechanobiology of the human intervertebral disc is a key element in degenerative disc disease. The technical advances that occurred in the last decade have allowed us to increase our understanding of this fascinating research topic, also enabling us to identify possible therapeutic targets in order to treat and to prevent DDD. Despite this, multiple pathways are involved in a complex and interplayed system, which is still partly unknown. Further research and technological innovation will further shed light on the interactions between the mechanics and biology of the human IVD.
Author Contributions
Conceptualization, G.V., M.T., S.N. and E.A.; methodology, M.T., G.V. and M.M.; formal analysis, G.V., F.B., A.R. and S.N.; data curation, M.T., M.I. and S.N.; writing—original draft preparation, M.T. and G.V.; writing—review and editing, G.V., M.T., S.N. and A.R.; supervision, S.N., E.A., A.R. and C.F.; project administration, C.F. and F.V.; funding acquisition, A.R. and S.N. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Thanks to “Grafica Biomedica” for its valuable artwork in graphically summarizing biological mechanoresponses of human intervertebral cells.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ehrlich, G.E. Low back pain. Bull. World Health Organ. 2003, 81, 671. [Google Scholar] [PubMed]
- Dagenais, S.; Caro, J.; Haldeman, S. A systematic review of low back pain cost of illness studies in the United States and internationally. Spine J. 2008, 8, 8–20. [Google Scholar] [CrossRef] [PubMed]
- DePalma, M.J.; Ketchum, J.M.; Saullo, T. What Is the Source of Chronic Low Back Pain and Does Age Play a Role? Pain Med. 2011, 12, 224–233. [Google Scholar] [CrossRef]
- Guiot, B.H.; Fessler, R.G.; Benzel, E.C.; Parsa, A.T.; McCormick, P.C.; Sonntag, V.K.H. Molecular Biology of Degenerative Disc Disease. Neurosurgery 2000, 47, 1034–1040. [Google Scholar] [CrossRef]
- Eyre, D.; Buckwalter, J.; Caterson, B.; Heinegard, D.; Oegema, T.; Pearce, R.; Pope, M.; Urban, J. Basic Sciences Perspectives: Part B—Intervertebral Discs; American Academy of Orthopaedic Surgeons: Park Ridge, IL, USA, 1989. [Google Scholar]
- Diamant, B.; Karlsson, J.; Nachemson, A. Correlation between lactate levels and pH in discs of patients with lumbar rhizopathies. Cell Mol. Life Sci. 1968, 24, 1195–1196. [Google Scholar] [CrossRef]
- Nachemson, A. Intradiscal Measurements of pH in Patients with Lumbar Rhizopathies. Acta Orthop. 1969, 40, 23–42. [Google Scholar] [CrossRef]
- Wilke, H.J.; Neef, P.; Caimi, M.; Hoogland, T.; Claes, L.E. New In-vivo measurements of pressures in disc in daily. Spine 1999, 8, 755–762. [Google Scholar] [CrossRef]
- Costi, J.; Stokes, I.; Gardner-Morse, M.; Laible, J.; Scoffone, H.; Iatridis, J. Direct measurement of intervertebral disc maximum shear strain in six degrees of freedom: Motions that place disc tissue at risk of injury. J. Biomech. 2007, 40, 2457–2466. [Google Scholar] [CrossRef] [PubMed]
- Ambard, D.; Cherblanc, F. Mechanical Behavior of Annulus Fibrosus: A Microstructural Model of Fibers Reorientation. Ann. Biomed. Eng. 2009, 37, 2256–2265. [Google Scholar] [CrossRef] [PubMed]
- Freemont, A.; Peacock, T.; Goupille, P.; Hoyland, J.; O’Brien, J.; Jayson, M. Nerve ingrowth into diseased intervertebral disc in chronic back pain. Lancet 1997, 350, 178–181. [Google Scholar] [CrossRef] [PubMed]
- David, G.; Ciurea, A.V.; Iencean, S.M.; Mohan, A. Angiogenesis in the degeneration of the lumbar intervertebral disc. J. Med. Life 2010, 3, 154–161. [Google Scholar] [PubMed]
- La Binch, A.; Cole, A.A.; Breakwell, L.M.; Michael, A.L.; Chiverton, N.; Cross, A.K.; Le Maitre, C.L. Expression and regulation of neurotrophic and angiogenic factors during human intervertebral disc degeneration. Thromb. Haemost. 2014, 16, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.-Q.; Jiang, L.-S.; Dai, L.-Y. Programmed cell death in intervertebral disc degeneration. Apoptosis 2006, 11, 2079–2088. [Google Scholar] [CrossRef] [PubMed]
- Le Maitre, C.; Freemont, A.; Hoyland, J. Accelerated cellular senescence in degenerate intervertebral discs: A possible role in the pathogenesis of intervertebral disc degeneration. Thromb. Haemost. 2007, 9, R45. [Google Scholar] [CrossRef] [PubMed]
- Hastreiter, D.; Ozuna, R.M.; Spector, M. Regional variations in certain cellular characteristics in human lumbar intervertebral discs, including the presence of α-smooth muscle actin. J. Orthop. Res. Publ. Orthop. Res. Soc. 2001, 19, 597–604. [Google Scholar] [CrossRef]
- Hunter, C.; Matyas, J.; Duncan, N. The Notochordal Cell in the Nucleus Pulposus: A Review in the Context of Tissue Engineering. Tissue Eng. 2003, 9, 667–677. [Google Scholar] [CrossRef]
- Le Maitre, C.; Pockert, A.; Buttle, D.; Freemont, T.; Hoyland, J. Matrix synthesis and degradation in human intervertebral disc degeneration. Biochem. Soc. Trans. 2007, 35, 652–655. [Google Scholar] [CrossRef]
- Akyol, S.; Eraslan, B.S.; Etyemez, H.; Tanriverdi, T.; Hanci, M. Catabolic cytokine expressions in patients with degenerative disc disease. Turk. Neurosurg. 2010, 20, 492–499. [Google Scholar] [CrossRef]
- Le Maitre, C.L.; Hoyland, J.A.; Freemont, A.J. Catabolic cytokine expression in degenerate and herniated human intervertebral discs: IL-1β and TNFα expression profile. Thromb. Haemost. 2007, 9, R77. [Google Scholar] [CrossRef]
- Kang, J.D.; Georgescu, H.I.; McIntyre-Larkin, L.; Stefanovic-Racic, M.; Evans, C.H. Herniated Cervical Intervertebral Discs Spontaneously Produce Matrix Metalloproteinases, Nitric Oxide, Interleukin-6, and Prostaglandin E2. Spine 1995, 20, 2373–2378. [Google Scholar] [CrossRef]
- Kang, J.D.; Georgescu, H.I.; McIntyre-Larkin, L.; Stefanovic-Racic, M.; Donaldson, W.F., III; Evans, C.H. Herniated Lumbar Intervertebral Discs Spontaneously Produce Matrix Metalloproteinases, Nitric Oxide, Interleukin-6, and Prostaglandin E2. Spine 1996, 21, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Miyagi, M.; Uchida, K.; Inoue, S.; Takano, S.; Nakawaki, M.; Kawakubo, A.; Sekiguchi, H.; Nakazawa, T.; Imura, T.; Saito, W.; et al. A High Body Mass Index and the Vacuum Phenomenon Upregulate Pain-Related Molecules in Human Degenerated Intervertebral Discs. Int. J. Mol. Sci. 2022, 23, 2973. [Google Scholar] [CrossRef] [PubMed]
- Lyu, F.-J.; Cui, H.; Pan, H.; MC Cheung, K.; Cao, X.; Iatridis, J.C.; Zheng, Z. Painful intervertebral disc degeneration and inflammation: From laboratory evidence to clinical interventions. Bone Res. 2021, 9, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Neidlinger-Wilke, C.; Galbusera, F.; Pratsinis, H.; Mavrogonatou, E.; Mietsch, A.; Kletsas, D.; Wilke, H.-J. Mechanical loading of the intervertebral disc: From the macroscopic to the cellular level. Eur. Spine J. 2014, 23, 333–343. [Google Scholar] [CrossRef]
- Setton, L.A.; Chen, J. Cell Mechanics and Mechanobiology in the Intervertebral Disc. Spine 2004, 29, 2710–2723. [Google Scholar] [CrossRef]
- Setton, L.A.; Chen, J. Mechanobiology of the Intervertebral Disc and Relevance to Disc Degeneration. J. Bone Jt. Surg. 2006, 88, 52–57. [Google Scholar] [CrossRef]
- Steele, J.; Bruce-Low, S.; Smith, D.; Osborne, N.; Thorkeldsen, A. Can specific loading through exercise impart healing or regeneration of the intervertebral disc? Spine J. 2015, 15, 2117–2121. [Google Scholar] [CrossRef]
- Hsieh, A.H.; Twomey, J.D. Cellular mechanobiology of the intervertebral disc: New directions and approaches. J. Biomech. 2010, 43, 137–145. [Google Scholar] [CrossRef]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; Altman, D.; Antes, G.; Atkins, D.; Barbour, V.; Barrowman, N.; Berlin, J.A.; et al. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef]
- Sideri, S.; Papageorgiou, S.N.; Eliades, T. Registration in the international prospective register of systematic reviews (PROSPERO) of systematic review protocols was associated with increased review quality. J. Clin. Epidemiology 2018, 100, 103–110. [Google Scholar] [CrossRef]
- Song, M.; Zhang, Y.; Sun, Y.; Kong, M.; Han, S.; Wang, C.; Wang, Y.; Xu, D.; Tu, Q.; Zhu, K.; et al. Inhibition of RhoA/MRTF-A signaling alleviates nucleus pulposus fibrosis induced by mechanical stress overload. Connect. Tissue Res. 2022, 63, 53–68. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Liu, B.; Liu, Z.-H.; Song, W.; Wang, D.; Chen, B.-Y.; Fan, J.; Xu, Z.; Geng, D.; Luo, Z.-J. Notochordal-Cell-Derived Exosomes Induced by Compressive Load Inhibit Angiogenesis via the miR-140-5p/Wnt/β-Catenin Axis. Mol. Ther. Nucl. Acids 2020, 22, 1092–1106. [Google Scholar] [CrossRef] [PubMed]
- Neidlinger-Wilke, C.; Würtz, K.; Liedert, A.; Schmidt, C.; Börm, W.; Ignatius, A.; Wilke, H.-J.; Claes, L. A three-dimensional collagen matrix as a suitable culture system for the comparison of cyclic strain and hydrostatic pressure effects on intervertebral disc cells. J. Neurosurgery: Spine 2005, 2, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Le Maitre, C.L.; Frain, J.; Fotheringham, A.P.; Freemont, A.J.; Hoyland, J.A. Human cells derived from degenerate intervertebral discs respond differently to those derived from non-degenerate intervertebral discs following application of dynamic hydrostatic pressure. Biorheology 2008, 45, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Le Maitre, C.L.; Frain, J.; Millward-Sadler, J.; Fotheringham, A.P.; Freemont, A.J.; Hoyland, J.A. Altered integrin mechanotransduction in human nucleus pulposus cells derived from degenerated discs. Arthritis Rheum. 2009, 60, 460–469. [Google Scholar] [CrossRef]
- Aladin, D.M.K.; Cheung, K.; Chan, D.; Yee, A.F.Y.; Jim, J.J.T.; Luk, K.D.K.; Lu, W.W. Expression of the Trp2 Allele of COL9A2 is Associated with Alterations in the Mechanical Properties of Human Intervertebral Discs. Spine 2007, 32, 2820–2826. [Google Scholar] [CrossRef]
- Franco-Obregón, A.; Cambria, E.; Greutert, H.; Wernas, T.; Hitzl, W.; Egli, M.; Sekiguchi, M.; Boos, N.; Hausmann, O.; Ferguson, S.J.; et al. TRPC6 in simulated microgravity of intervertebral disc cells. Eur Spine J. 2018, 27, 2621–2630. [Google Scholar] [CrossRef]
- Cambria, E.; Arlt, M.; Wandel, S.; Krupkova, O.; Hitzl, W.; Passini, F.; Hausmann, O.; Snedeker, J.; Ferguson, S.; Wuertz-Kozak, K. TRPV4 Inhibition and CRISPR-Cas9 Knockout Reduce Inflammation Induced by Hyperphysiological Stretching in Human Annulus Fibrosus Cells. Cells 2020, 9, 1736. [Google Scholar] [CrossRef]
- Likhitpanichkul, M.; Torre, O.M.; Gruen, J.; Walter, B.A.; Hecht, A.C.; Iatridis, J.C. Do mechanical strain and TNF-α interact to amplify pro-inflammatory cytokine production in human annulus fibrosus cells? J. Biomech. 2016, 49, 1214–1220. [Google Scholar] [CrossRef]
- Gilbert, H.T.J.; Nagra, N.S.; Freemont, A.J.; Millward-Sadler, S.J.; Hoyland, J.A. Integrin—Dependent Mechanotransduction in Mechanically Stimulated Human Annulus Fibrosus Cells: Evidence for an Alternative Mechanotransduction Pathway Operating with Degeneration. PLoS ONE 2013, 8, e72994. [Google Scholar] [CrossRef]
- Gilbert, H.T.; Hoyland, J.A.; Freemont, A.J.; Millward-Sadler, S.J. The involvement of interleukin-1 and interleukin-4 in the response of human annulus fibrosus cells to cyclic tensile strain: An altered mechanotransduction pathway with degeneration. Thromb. Haemost. Res. Ther. 2011, 13, R8-12. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, H.T.J.; Hoyland, J.A.; Millward-Sadler, S.J. The response of human anulus fibrosus cells to cyclic tensile strain is frequency-dependent and altered with disc degeneration. Arthritis Rheum. 2010, 62, 3385–3394. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Wang, H.-Q.; Zhang, Q.; Yan, X.-D.; Hao, M.; Luo, Z.-J. Alterations of ADAMTSs and TIMP-3 in human nucleus pulposus cells subjected to compressive load: Implications in the pathogenesis of human intervertebral disc degeneration. J. Orthop. Res.Off. Publ. Orthop. Res. Soc. 2012, 30, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Neidlinger-Wilke, C.; Liedert, A.; Wuertz, K.; Buser, Z.; Rinkler, C.; Käfer, W.; Ignatius, A.; Claes, L.; Roberts, S.; Johnson, W.E.B. Mechanical Stimulation Alters Pleiotrophin and Aggrecan Expression by Human Intervertebral Disc Cells and Influences Their Capacity to Stimulate Endothelial Cell Migration. Spine 2009, 34, 663–669. [Google Scholar] [CrossRef] [PubMed]
- Hodson, N.W.; Patel, S.; Richardson, S.M.; Hoyland, J.A.; Gilbert, H.T.J. Degenerate intervertebral disc-like pH induces a catabolic mechanoresponse in human nucleus pulposus cells. JOR Spine 2018, 1, e1004. [Google Scholar] [CrossRef] [PubMed]
- Fearing, B.V.; Hernandez, P.A.; Setton, L.A.; Chahine, N.O. Mechanotransduction and cell biomechanics of the intervertebral disc. JOR Spine 2018, 1, e1026. [Google Scholar] [CrossRef]
- Chan, S.C.W.; Ferguson, S.J.; Gantenbein-Ritter, B. The effects of dynamic loading on the intervertebral disc. Eur. Spine J. 2011, 20, 1796–1812. [Google Scholar] [CrossRef]
- Kadow, T.; Sowa, G.; Vo, N.; Kang, J.D. Molecular Basis of Intervertebral Disc Degeneration and Herniations: What Are the Important Translational Questions? Clin. Orthop. Relat. Res. 2015, 473, 1903–1912. [Google Scholar] [CrossRef]
- Richardson, S.M.; Doyle, P.; Minogue, B.M.; Gnanalingham, K.; Hoyland, J.A. Increased expression of matrix metalloproteinase-10, nerve growth factor and substance P in the painful degenerate intervertebral disc. Thromb. Haemost. Res. Ther. 2009, 11, R126-8. [Google Scholar] [CrossRef]
- Purmessur, D.; Walter, B.; Roughley, P.; Laudier, D.; Hecht, A.; Iatridis, J. A role for TNFα in intervertebral disc degeneration: A non-recoverable catabolic shift. Biochem. Biophys. Res. Commun. 2013, 433, 151–156. [Google Scholar] [CrossRef]
- Daly, C.; Ghosh, P.; Jenkin, G.; Oehme, D.; Goldschlager, T. A Review of Animal Models of Intervertebral Disc Degeneration: Pathophysiology, Regeneration, and Translation to the Clinic. BioMed Res. Int. 2016, 2016, 1–14. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).