

Different Dietary Sources of Selenium Alleviate Hepatic Lipid Metabolism Disorder of Heat-Stressed Broilers by Relieving Endoplasmic Reticulum Stress
 , ,
, , 
Abstract
:1. Introduction
2. Results
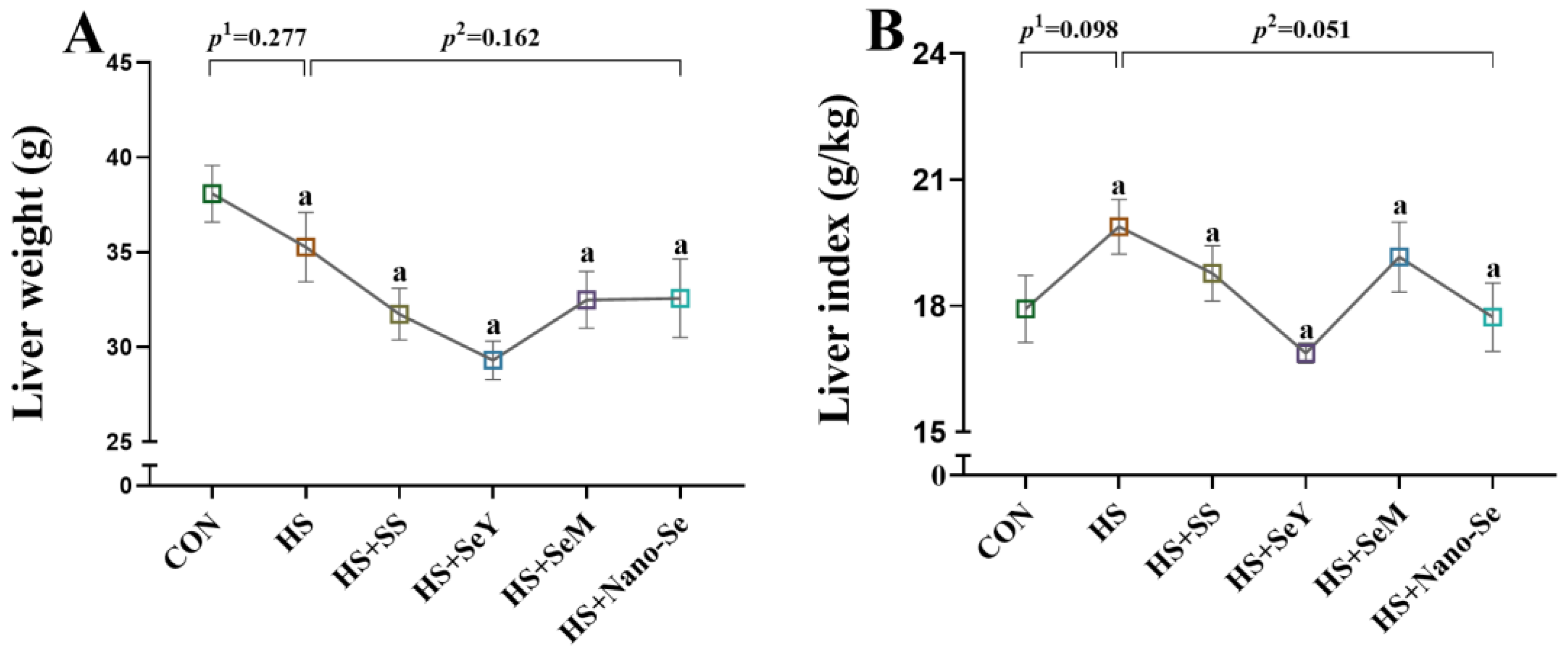
2.1. Liver Weight and Liver Index
2.2. Serum Biochemical Indicators
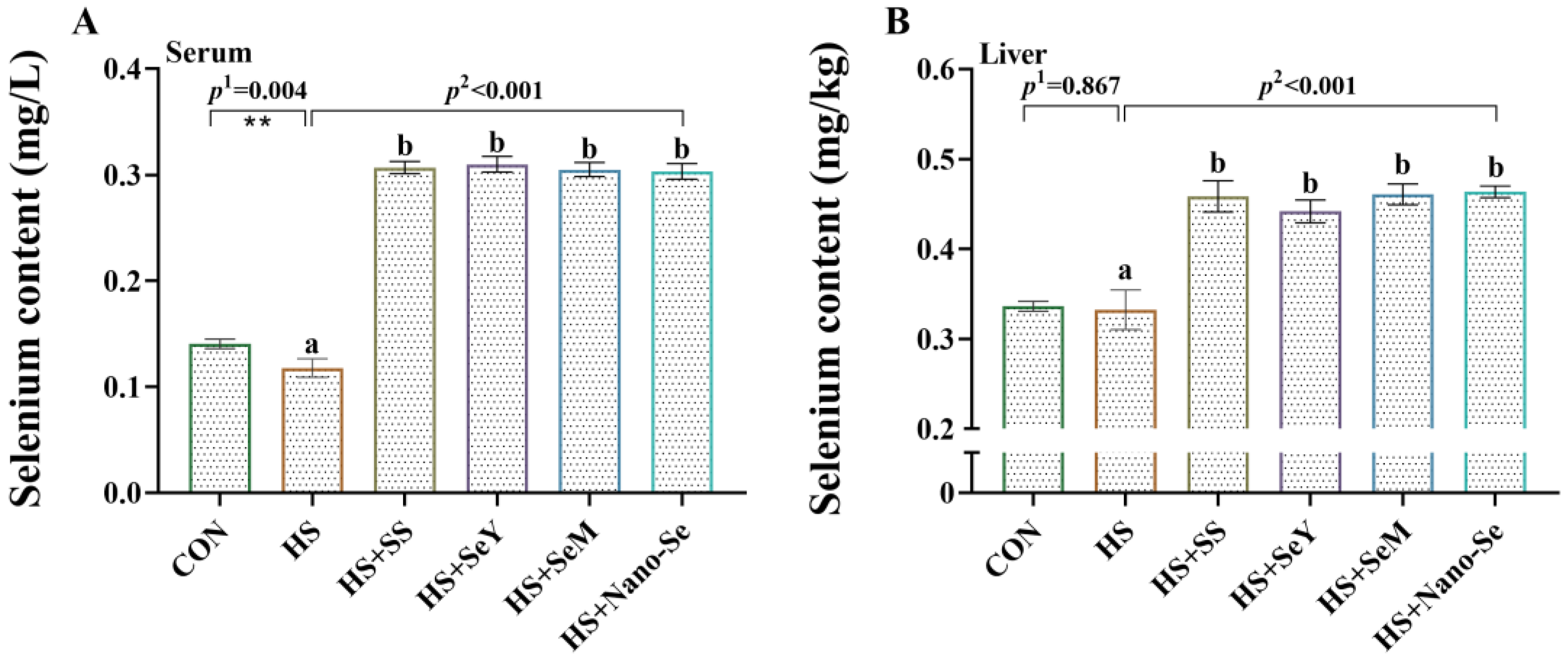
2.3. Se Concentration in Serum and the Liver
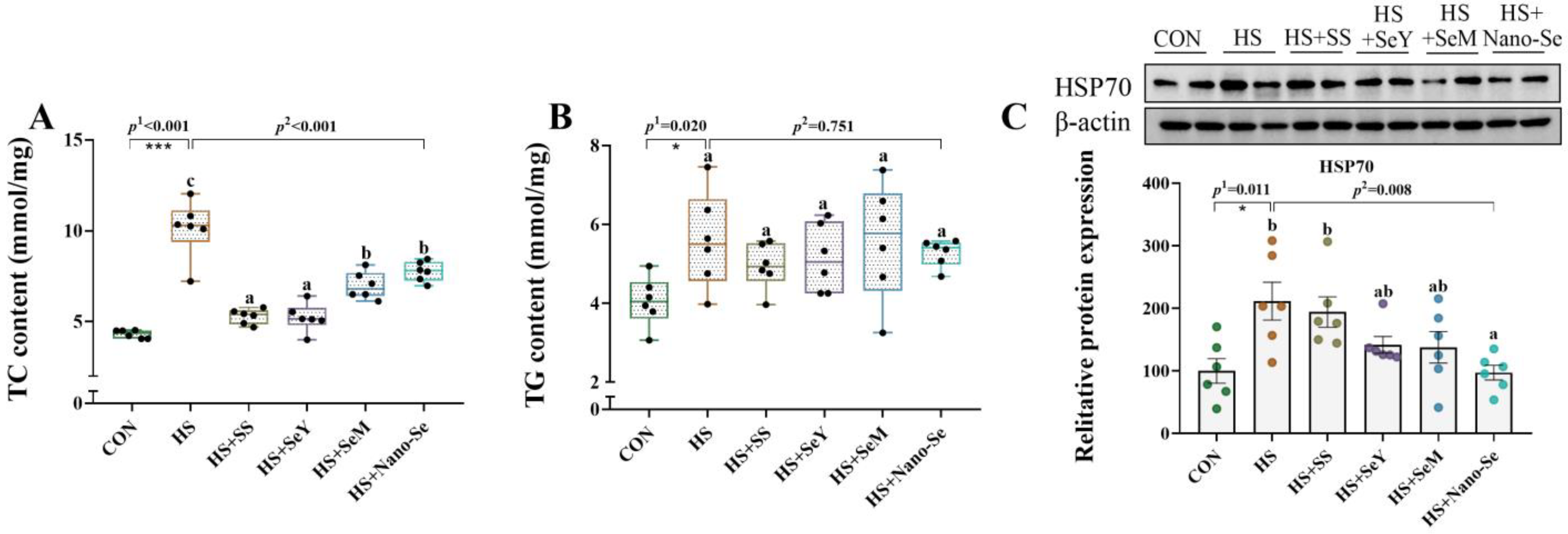
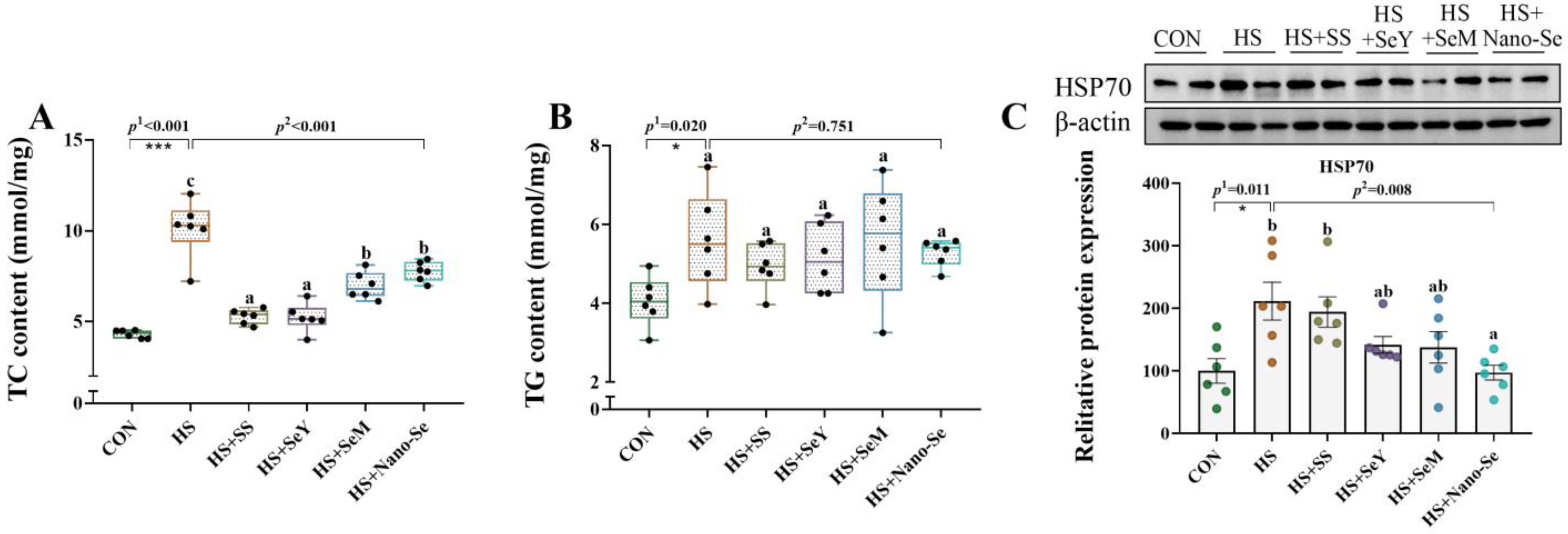
2.4. TC Content, TG Content, and HSP70 Protein Abundance in the Liver
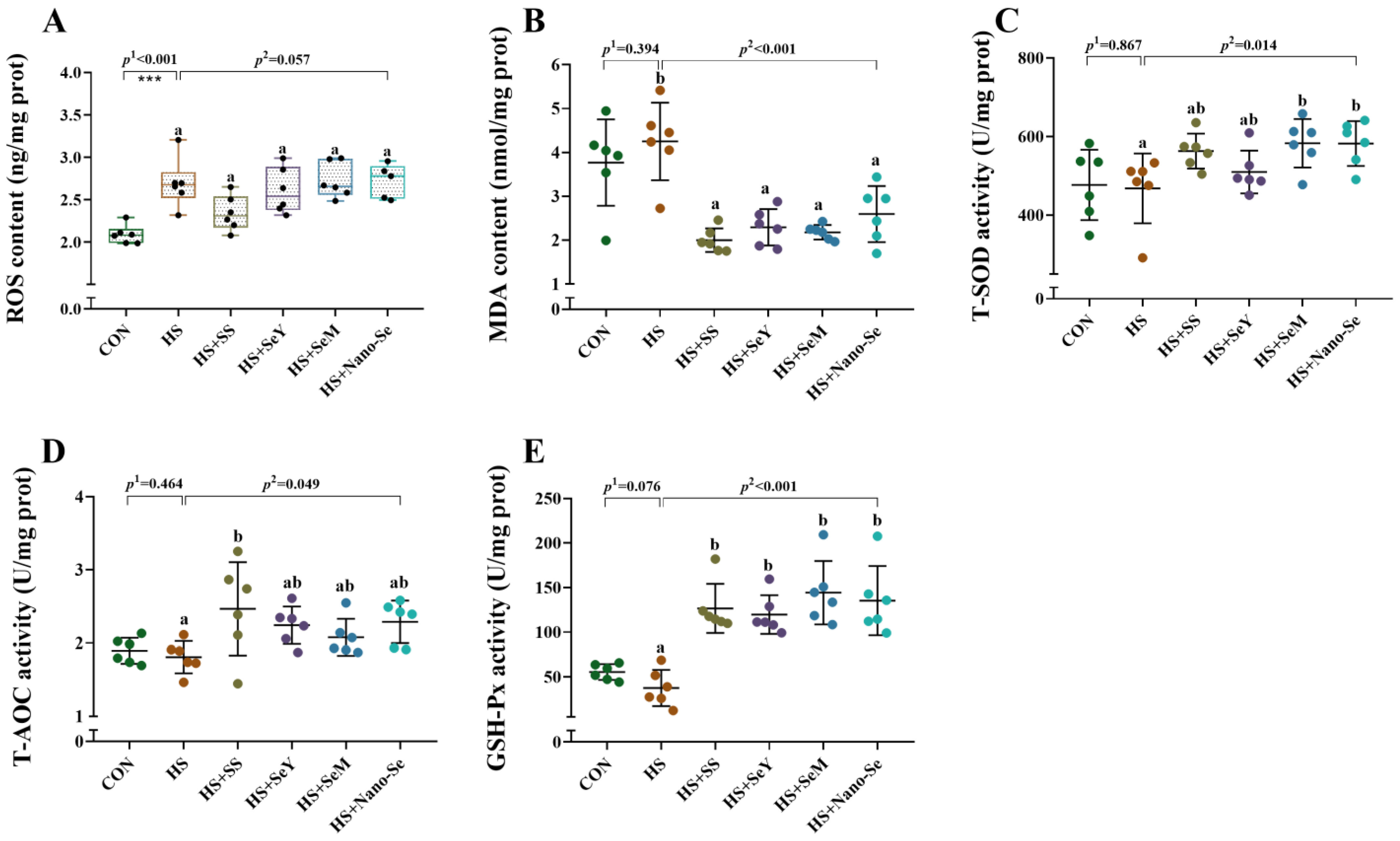
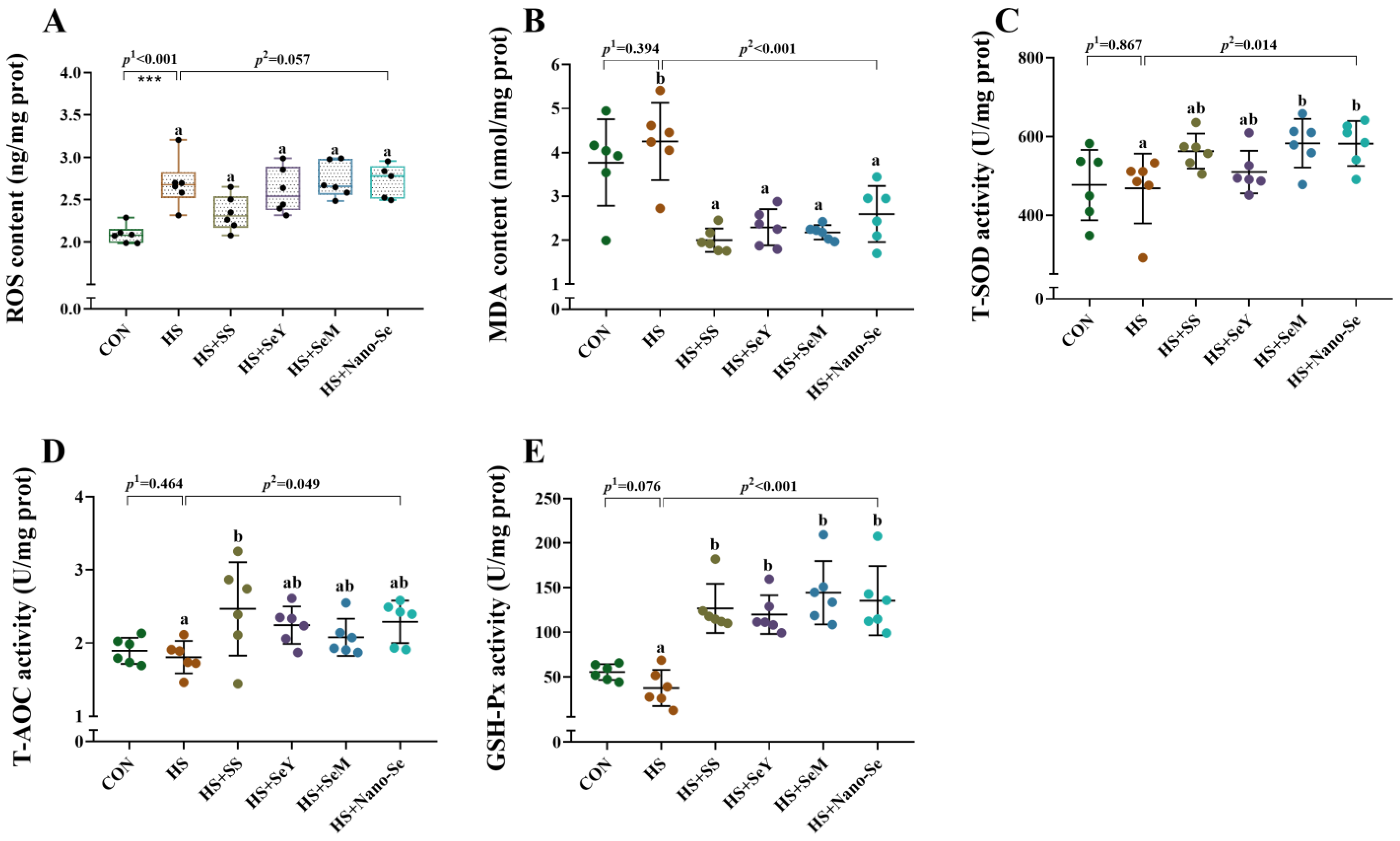
2.5. Hepatic Antioxidant Variables
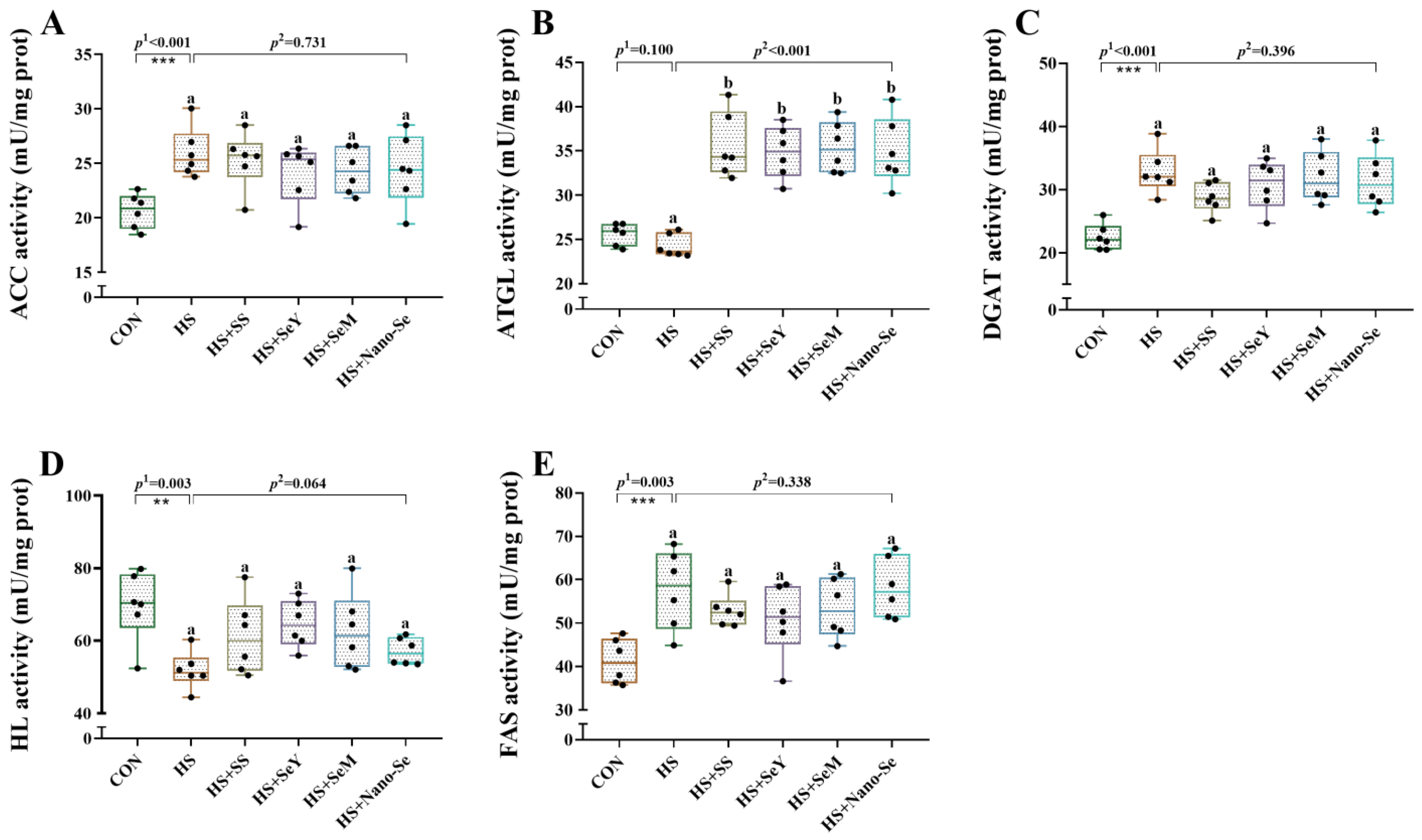
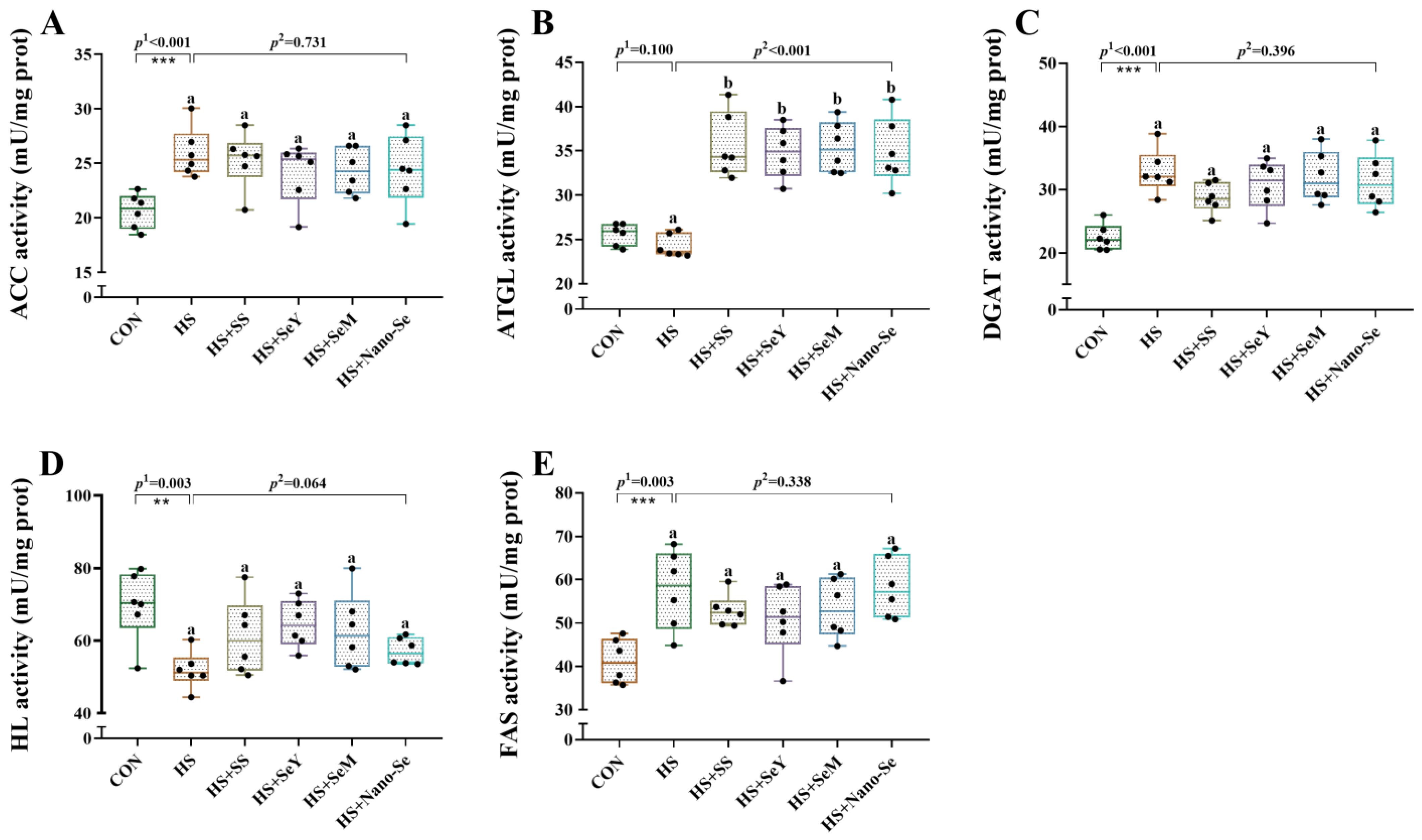
2.6. The Activity of Lipid Metabolic-Related Enzymes in the Liver
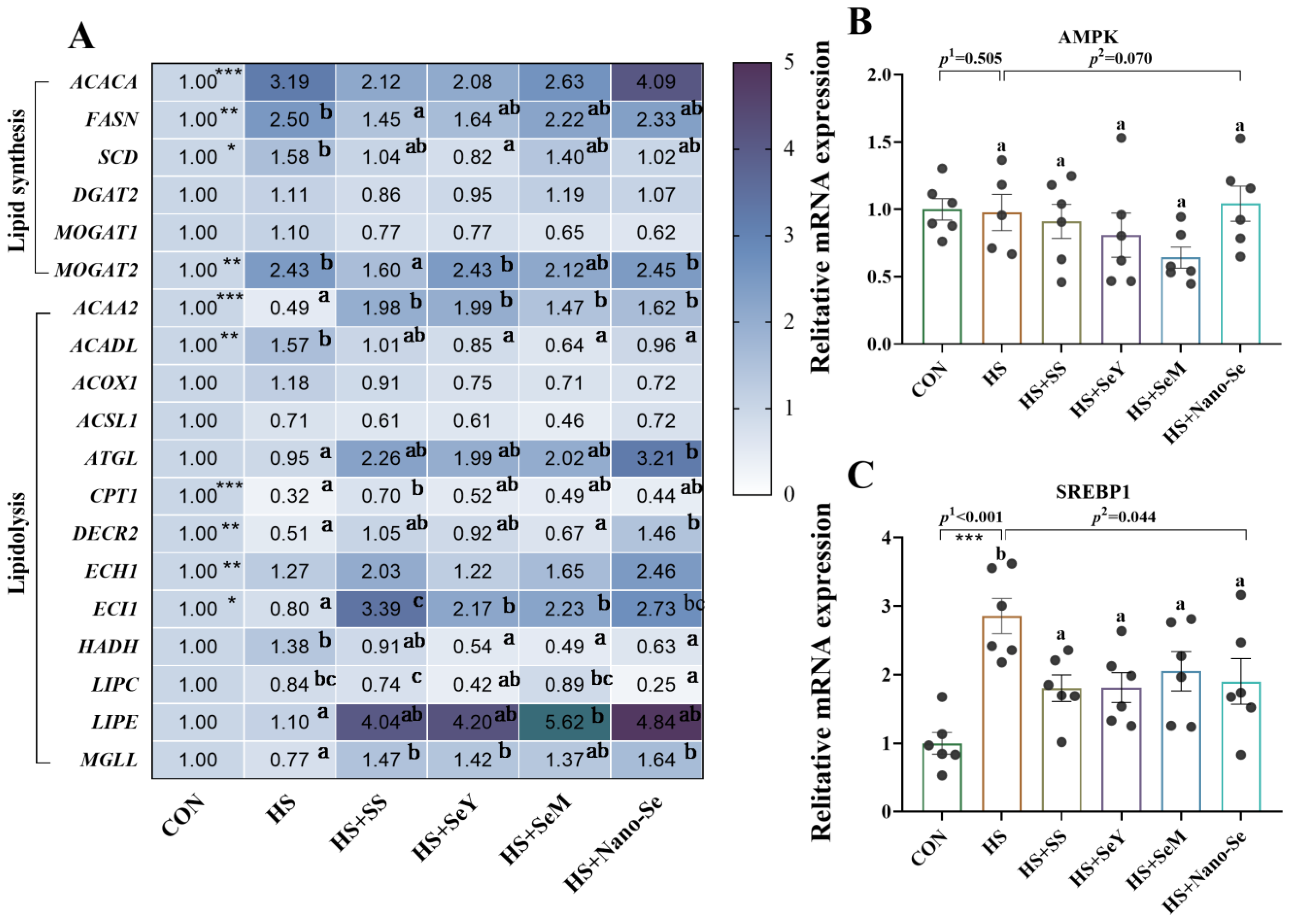
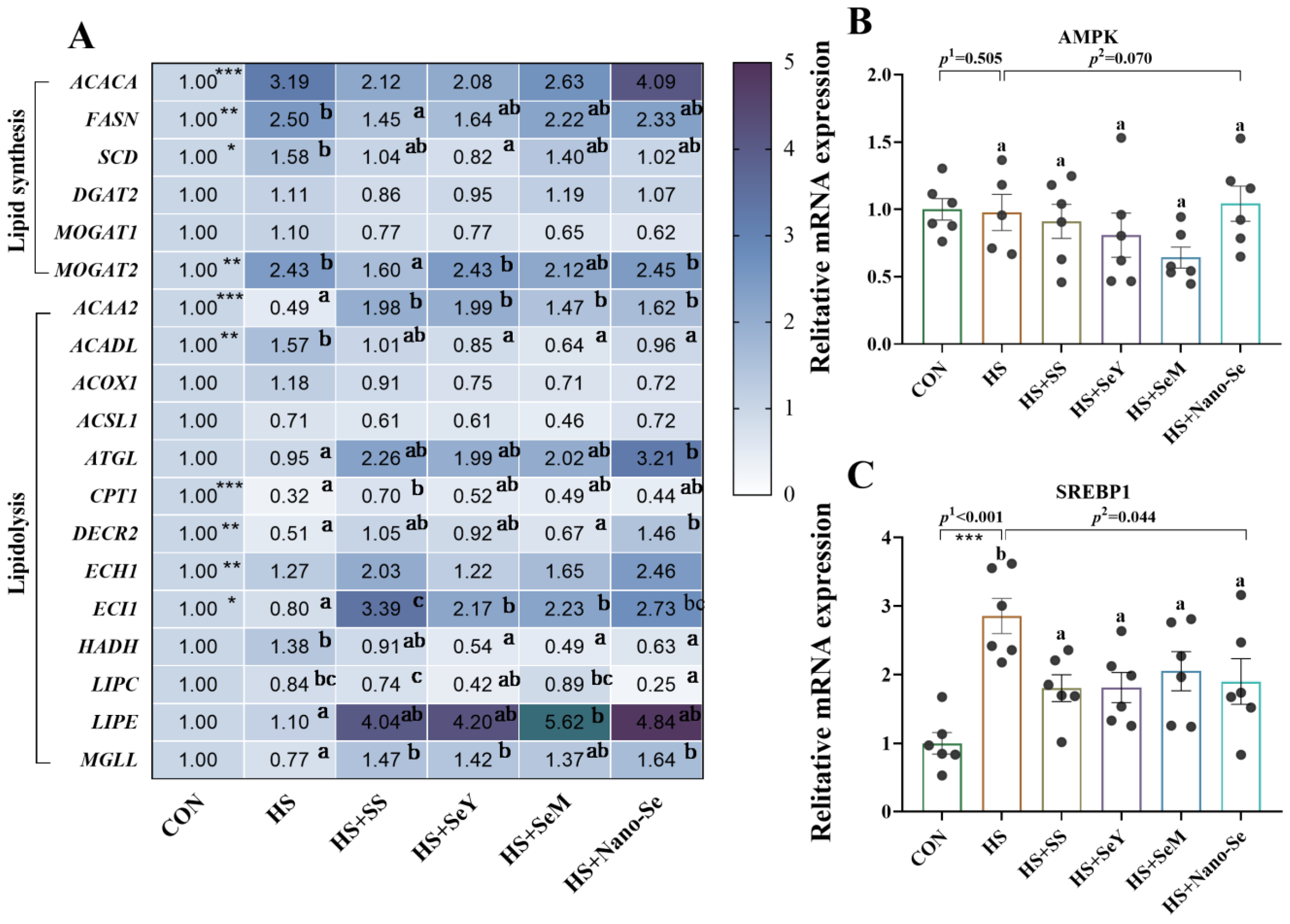
2.7. The Expression of Lipid Metabolic-Related Genes in the Liver
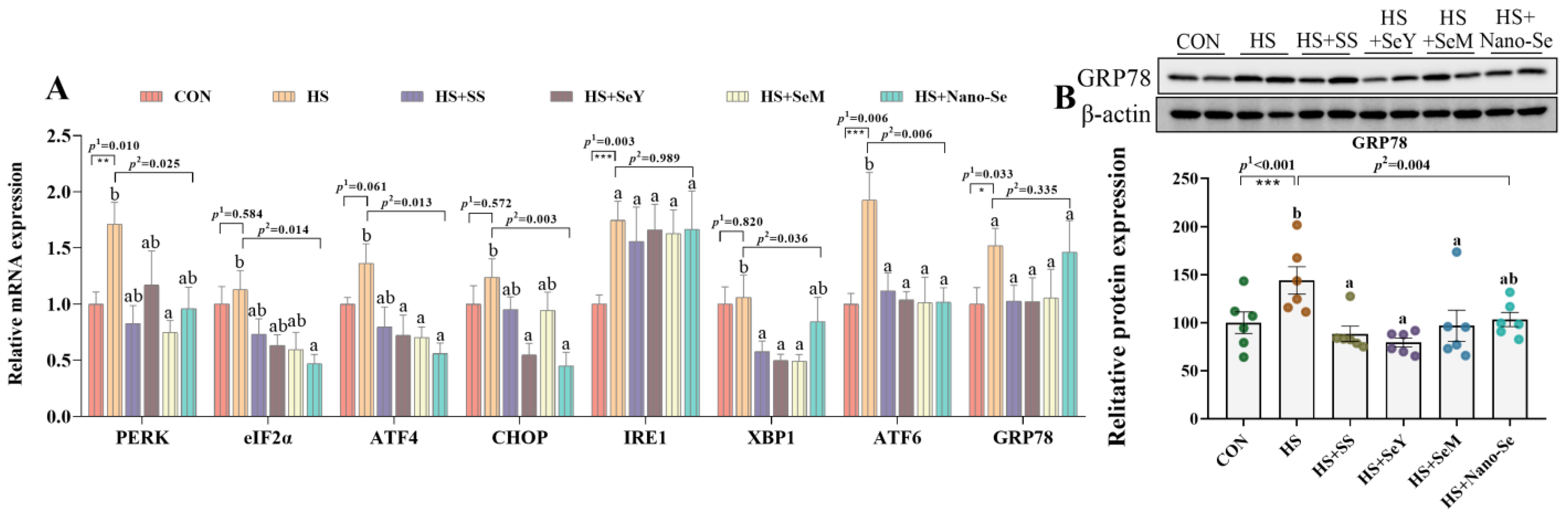
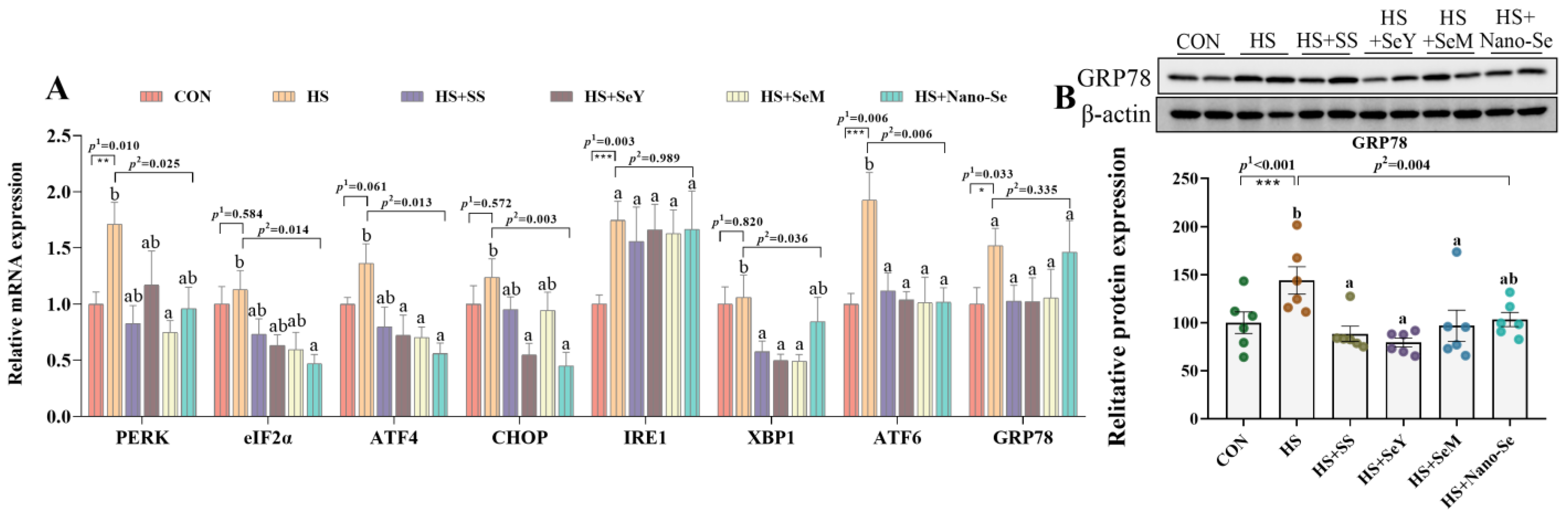
2.8. ER Stress Biomarkers in the Liver
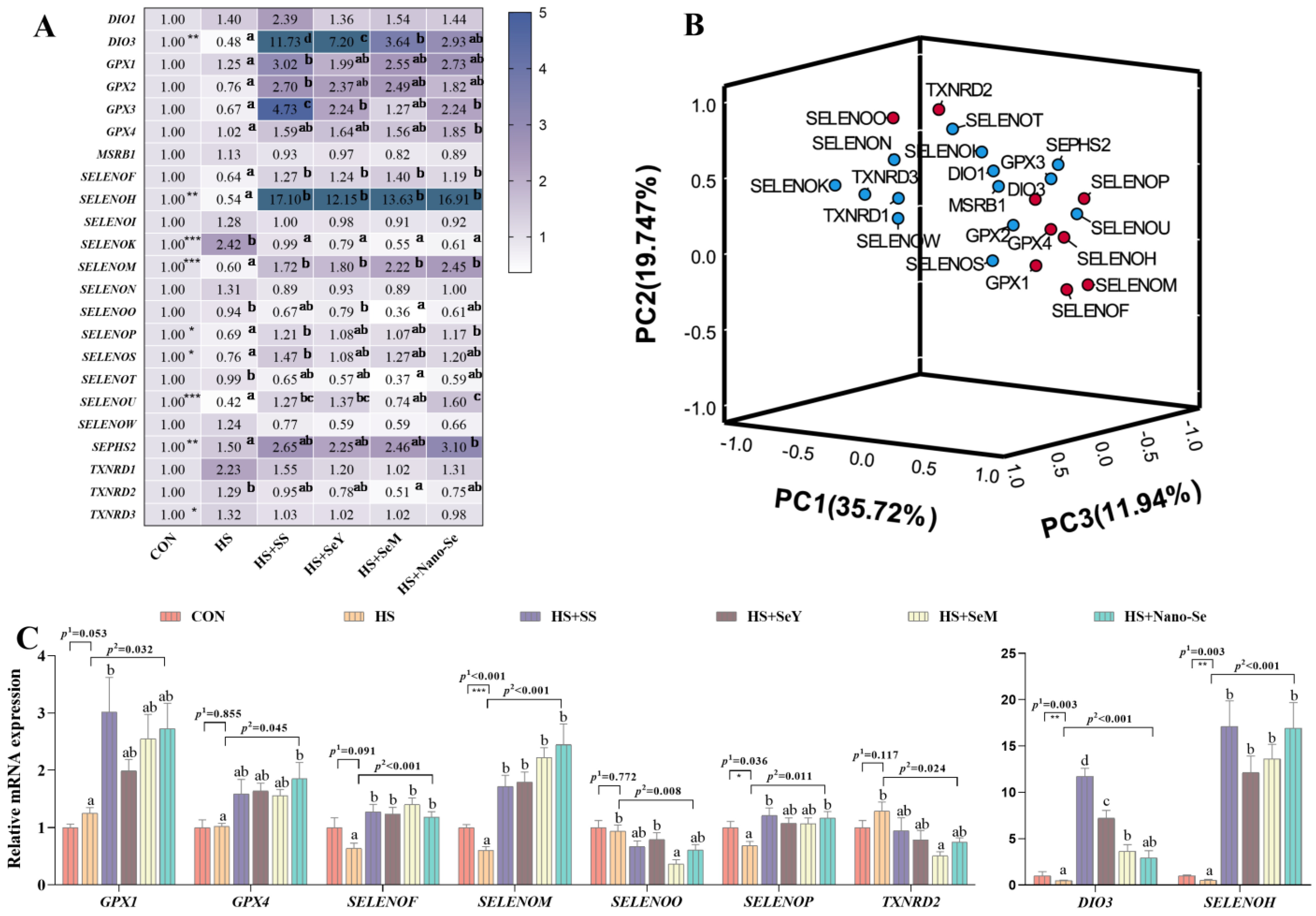
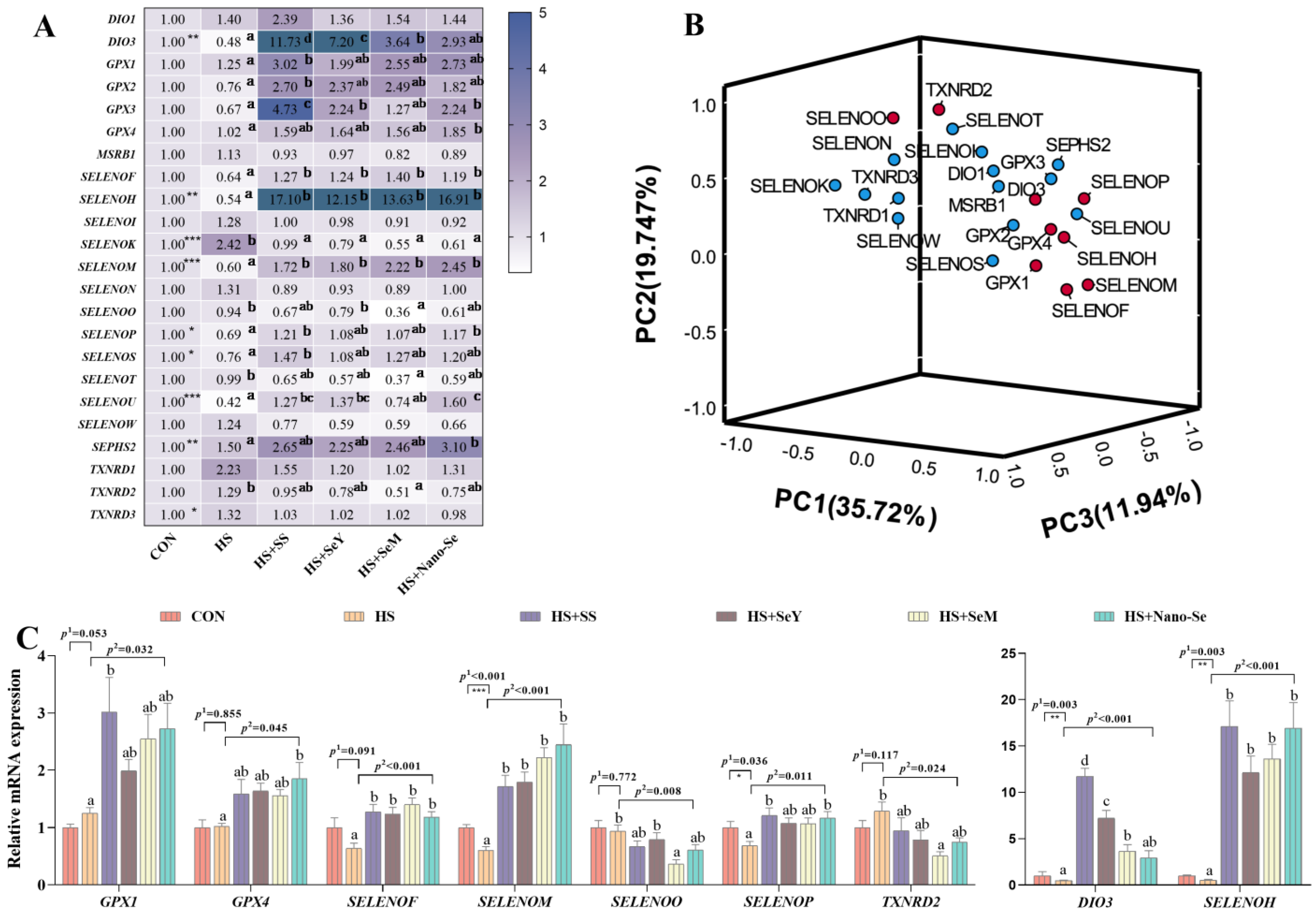
2.9. The mRNA Expression of Selenotranscriptome in the Liver
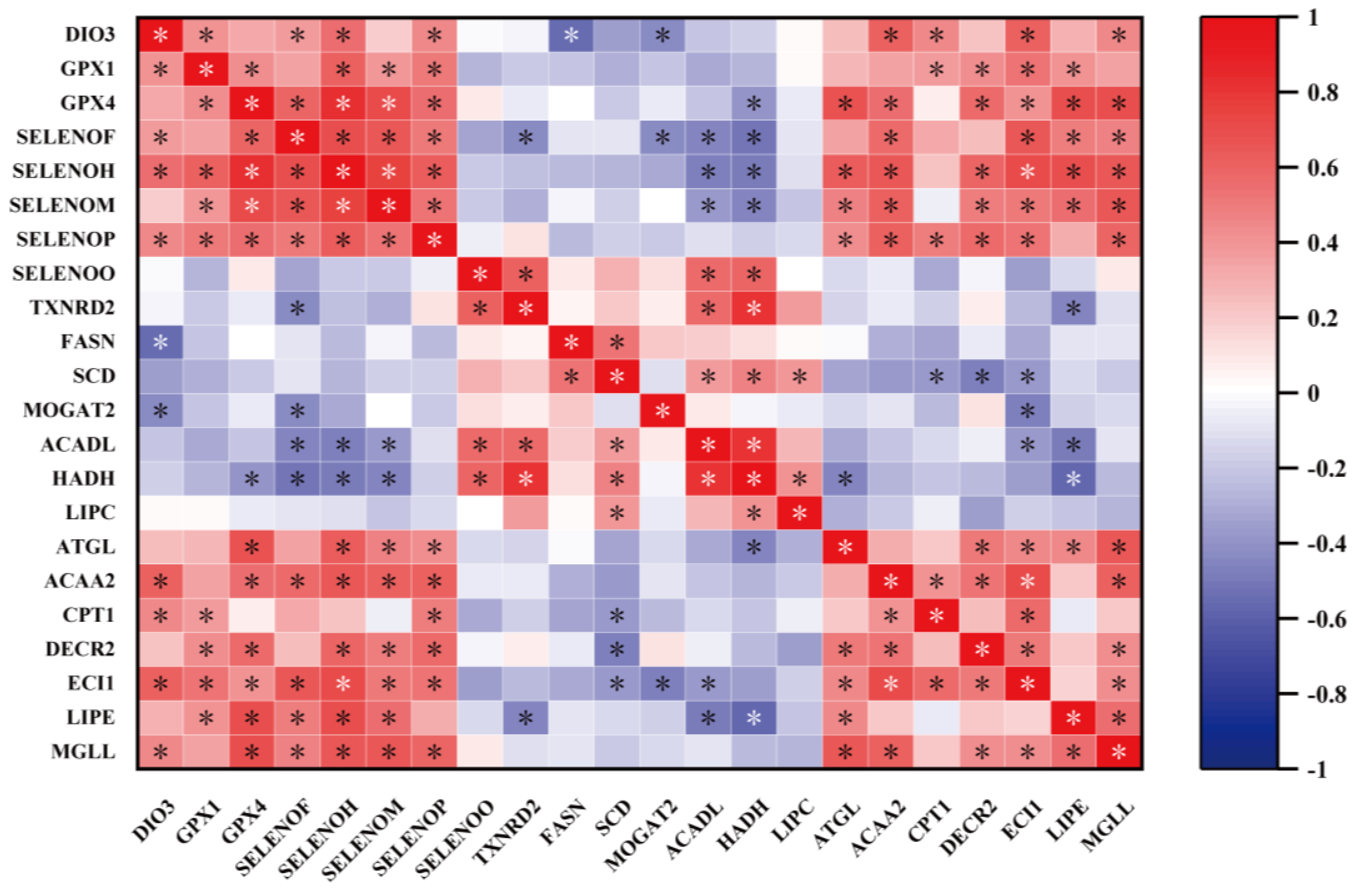
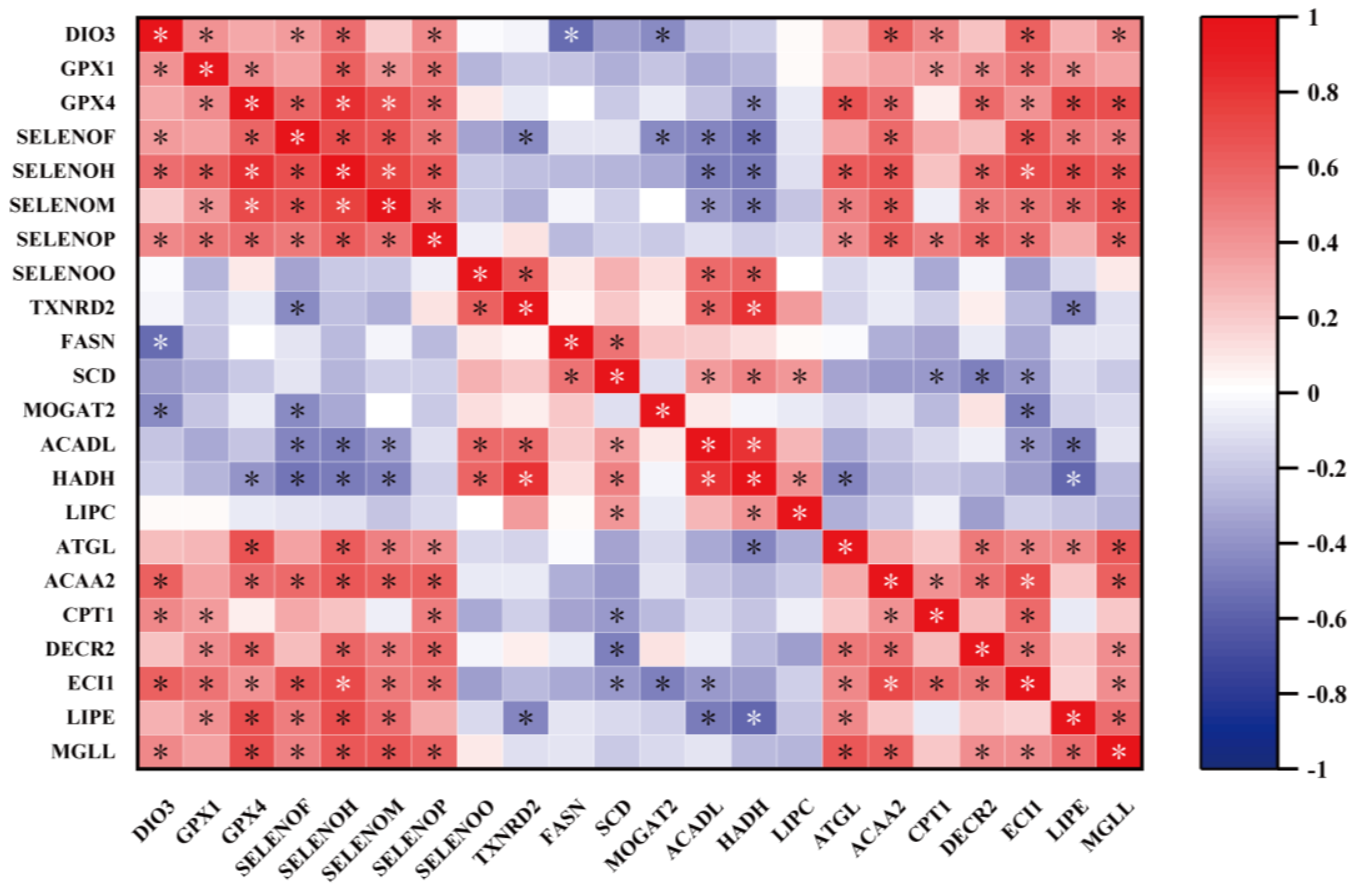
2.10. Correlation Analysis
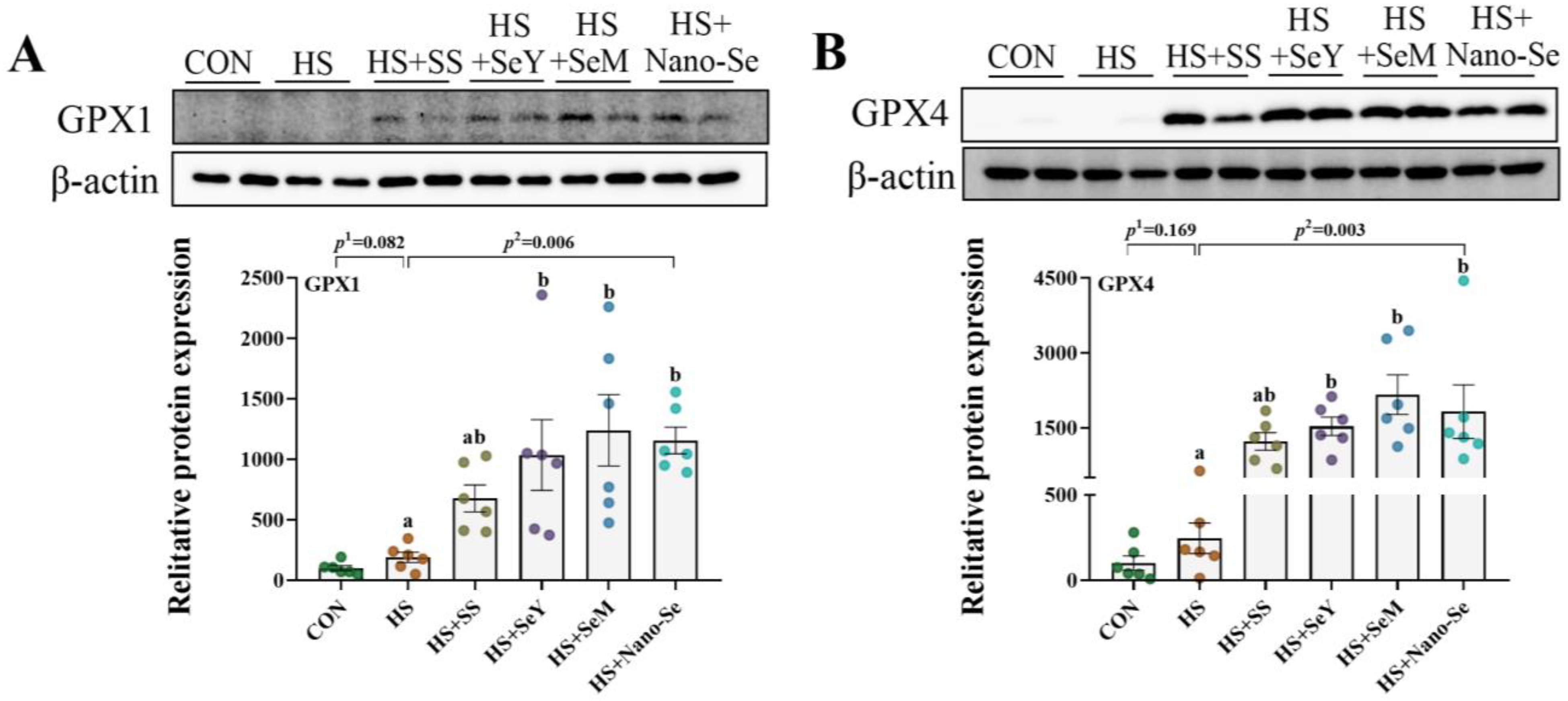
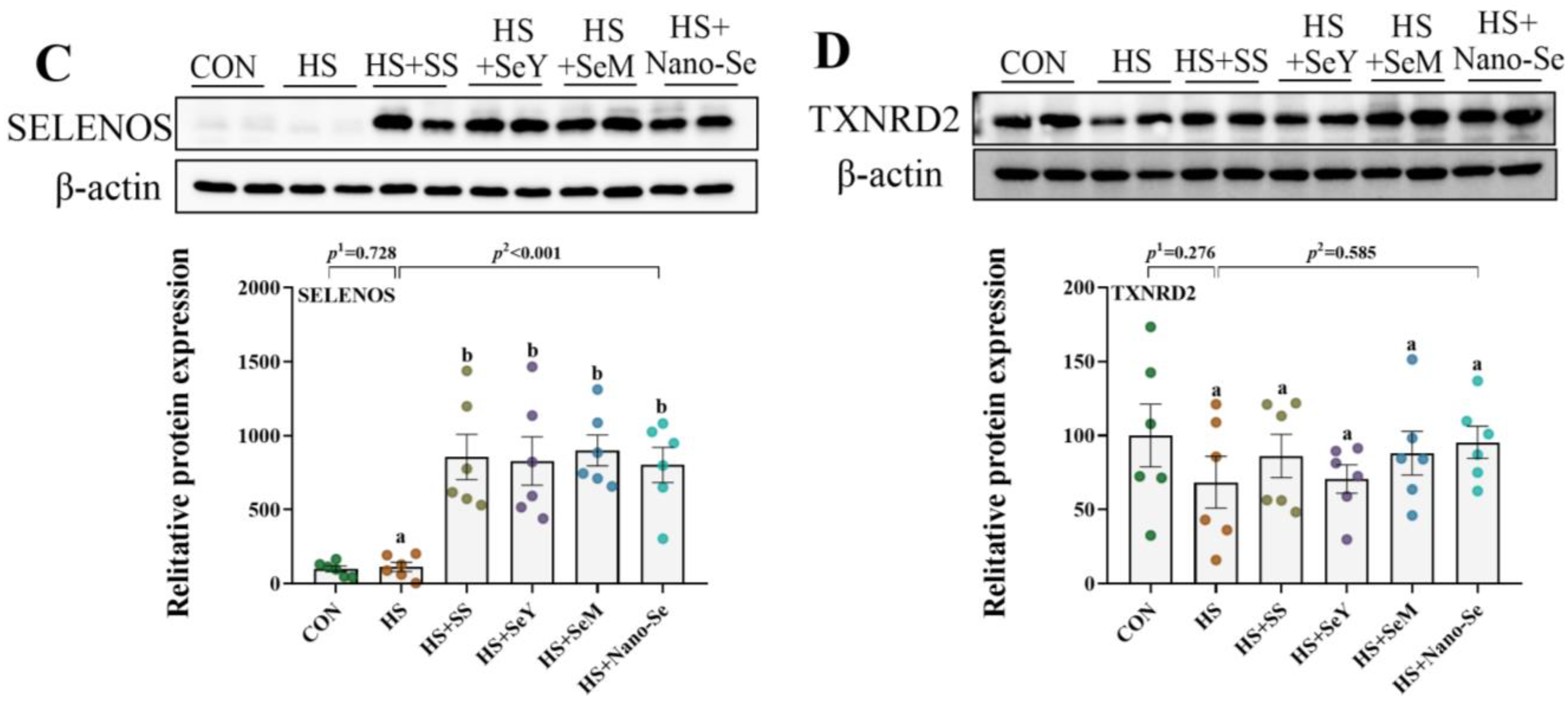
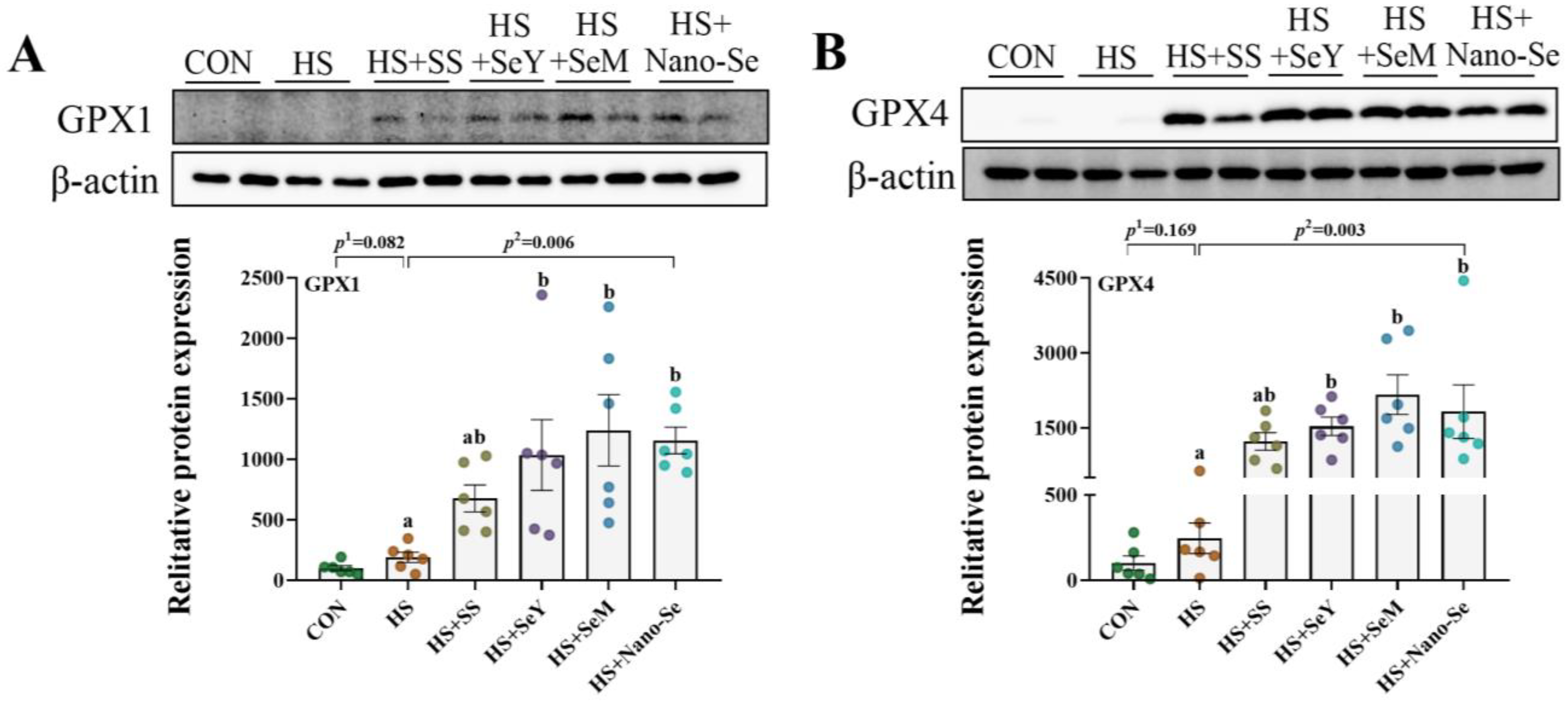
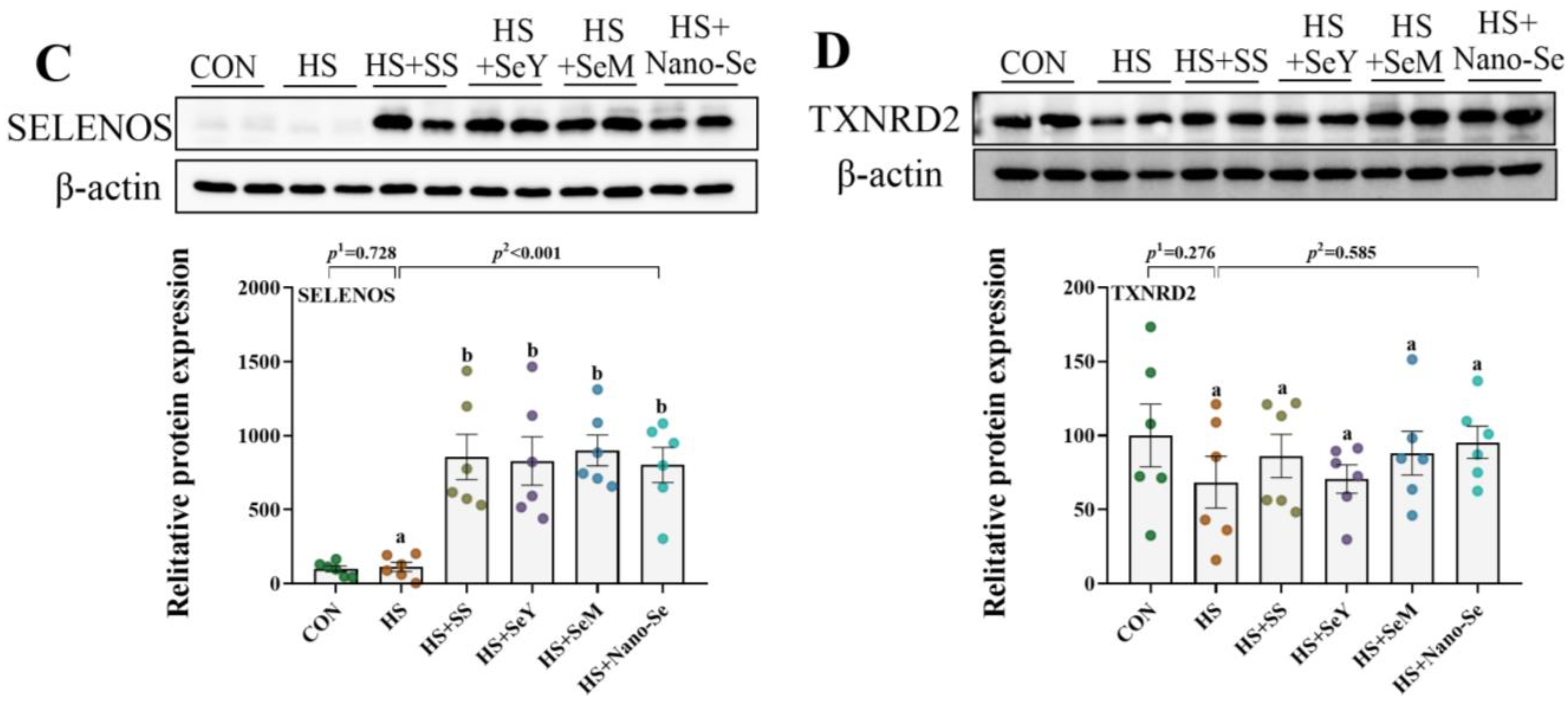
2.11. The Protein Abundance of Selenoproteins
3. Discussion
4. Materials and Methods
4.1. Animal, Diet, and Experimental Design
4.2. Liver Weight and Sample Collection
4.3. Serum Biochemical Analyses
4.4. Selenium Concentration in Diets, Serum and Liver
4.5. Liver Biochemical Analyses
4.6. Q-PCR Analyses of mRNA Abundance
4.7. Western Blot Analyses
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ali, M.Z.; Carlile, G.; Giasuddin, M. Impact of global climate change on livestock health: Bangladesh perspective. Open Vet. J. 2020, 10, 178–188. [Google Scholar] [CrossRef] [PubMed]
- St-Pierre, N.R.; Cobanov, B.; Schnitkey, G. Economic losses from heat stress by US livestock industries. J. Dairy Sci. 2003, 86, E52–E77. [Google Scholar] [CrossRef]
- Lara, L.J.; Rostagno, M.H. Impact of heat stress on poultry production. Animals 2013, 3, 356–369. [Google Scholar] [CrossRef] [PubMed]
- Leveille, G.A.; Romsos, D.R.; Yeh, Y.; O’Hea, E.K. Lipid biosynthesis in the chick. A consideration of site of synthesis, influence of diet and possible regulatory mechanisms. Poult. Sci. 1975, 54, 1075–1093. [Google Scholar] [CrossRef]
- Ramos, L.F.; Silva, C.M.; Pansa, C.C.; Moraes, K.C.M. Non-alcoholic fatty liver disease: Molecular and cellular interplays of the lipid metabolism in a steatotic liver. Expert Rev. Gastroenterol. Hepatol. 2021, 51, 25–40. [Google Scholar] [CrossRef]
- Slimen, I.B.; Najar, T.; Ghram, A.; Dabbebi, H.; Ben Mrad, M.; Abdrabbah, M. Reactive oxygen species, heat stress and oxidative-induced mitochondrial damage. A review. Int. J. Hyperth. 2014, 30, 513–523. [Google Scholar] [CrossRef]
- Marí, M.; Colell, A.; Morales, A.; von Montfort, C.; Garcia-Ruiz, C.; Fernández-Checa, J.C. Redox control of liver function in health and disease. Antioxid. Redox Signal. 2010, 12, 1295–1331. [Google Scholar] [CrossRef]
- Schwarz, D.S.; Blower, M.D. The endoplasmic reticulum: Structure, function and response to cellular signaling. Cell. Mol. Life Sci. 2016, 73, 79–94. [Google Scholar] [CrossRef]
- Ma, B.; Xing, T.; Li, J.; Zhang, L.; Jiang, Y.; Gao, F. Chronic heat stress causes liver damage via endoplasmic reticulum stress-induced apoptosis in broilers. Poult. Sci. 2022, 101, 102063. [Google Scholar] [CrossRef]
- Lemmer, I.L.; Willemsen, N.; Hilal, N.; Bartelt, A. A guide to understanding endoplasmic reticulum stress in metabolic disorders. Mol. Metab. 2021, 47, 101169. [Google Scholar] [CrossRef]
- Sicari, D.; Delaunay-Moisan, A.; Combettes, L.; Chevet, E.; Igbaria, A. A guide to assessing endoplasmic reticulum homeostasis and stress in mammalian systems. FEBS J. 2020, 287, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Rawson, R.B.; Komuro, R.; Chen, X.; Davé, U.P.; Prywes, R.; Brown, M.S.; Goldstein, J.L. ER stress induces cleavage of membrane-bound ATF6 by the same proteases that process SREBPs. Mol. Cell 2000, 6, 1355–1364. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, H.; Matsui, T.; Yamamoto, A.; Okada, T.; Mori, K. XBP1 mRNA is induced by ATF6 and spliced by IRE1 in response to ER stress to produce a highly active transcription factor. Cell 2001, 107, 881–891. [Google Scholar] [CrossRef] [PubMed]
- Harding, H.P.; Zhang, Y.; Ron, D. Protein translation and folding are coupled by an endoplasmic-reticulum-resident kinase. Nature 1999, 397, 271–274. [Google Scholar] [CrossRef] [PubMed]
- Wek, R.C.; Jiang, H.Y.; Anthony, T.G. Coping with stress: eIF2 kinases and translational control. Biochem. Soc. Trans. 2006, 34, 7–11. [Google Scholar] [CrossRef]
- Tang, J.; Cao, L.; Jia, G.; Liu, G.; Chen, X.; Tian, G.; Cai, J.; Shang, H.; Zhao, H. The protective effect of selenium from heat stress-induced porcine small intestinal epithelial cell line (IPEC-J2) injury is associated with regulation expression of selenoproteins. Br. J. Nutr. 2019, 122, 1081–1090. [Google Scholar] [CrossRef]
- Kryukov, G.V.; Castellano, S.; Novoselov, S.V.; Lobanov, A.V.; Zehtab, O.; Guigó, R.; Gladyshev, V.N. Characterization of mammalian selenoproteomes. Science 2003, 300, 1439–1443. [Google Scholar] [CrossRef]
- Chen, F.; Zhu, L.; Qiu, H.; Qin, S. Selenium-enriched Saccharomyces cerevisiae improves growth, antioxidant status and selenoprotein gene expression in Arbor Acres broilers. J. Anim. Physiol. Anim. Nutr. 2017, 101, 259–266. [Google Scholar] [CrossRef]
- Li, S.; Gao, F.; Huang, J.; Wu, Y.; Wu, S.; Lei, X.G. Regulation and function of avian selenogenome. Biochim. Biophys. Acta Gen. Subj. 2018, 1862, 2473–2479. [Google Scholar] [CrossRef]
- Zhao, L.; Chu, X.H.; Liu, S.; Li, R.; Zhu, Y.F.; Li, F.N.; Jiang, J.; Zhou, J.C.; Lei, X.G.; Sun, L.H. Selenium-enriched cardamine violifolia increases selenium and decreases cholesterol concentrations in liver and pectoral muscle of broilers. J. Nutr. 2022, 152, 2072–2079. [Google Scholar] [CrossRef]
- Jing, J.; Yin, S.; Liu, Y.; Liu, Y.; Wang, L.; Tang, J.; Jia, G.; Liu, G.; Tian, G.; Chen, X.; et al. Hydroxy selenomethionine alleviates hepatic lipid metabolism disorder of pigs induced by dietary oxidative stress via relieving the endoplasmic reticulum stress. Antioxidants 2022, 11, 552. [Google Scholar] [CrossRef]
- Liu, Y.; Tang, J.; He, Y.; Jia, G.; Liu, G.; Tian, G.; Chen, X.; Cai, J.; Kang, B.; Zhao, H. Selenogenome and AMPK signal insight into the protective effect of dietary selenium on chronic heat stress-induced hepatic metabolic disorder in growing pigs. J. Anim. Sci. Biotechnol. 2021, 12, 68. [Google Scholar] [CrossRef]
- Addinsall, A.B.; Wright, C.R.; Andrikopoulos, S.; van der Poel, C.; Stupka, N. Emerging roles of endoplasmic reticulum-resident selenoproteins in the regulation of cellular stress responses and the implications for metabolic disease. Biochem. J. 2018, 475, 1037–1057. [Google Scholar] [CrossRef]
- Hadrup, N.; Ravn-Haren, G. Absorption, distribution, metabolism and excretion (ADME) of oral selenium from organic and inorganic sources: A review. J. Trace Elem. Med. Biol. 2021, 67, 126801. [Google Scholar] [CrossRef]
- Ringuet, M.T.; Hunne, B.; Lenz, M.; Bravo, D.M.; Furness, J.B. Analysis of bioavailability and Induction of Glutathione Peroxidase by Dietary Nanoelemental. Organic and Inorganic Selenium. Nutrients 2021, 13, 1073. [Google Scholar] [CrossRef]
- Surai, P.F.; Kochish, I.I.; Fisinin, V.I.; Velichko, O.A. Selenium in poultry nutrition: From sodium selenite to organic selenium sources. J. Poult. Sci. 2018, 55, 79–93. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.S.; Gao, X.Y.; Zhang, L.D.; Bao, Y.P. Biological effects of a nano red elemental selenium. BioFactors 2001, 15, 27–38. [Google Scholar] [CrossRef]
- Nguyen, P.; Leray, V.; Diez, M.; Serisier, S.; Le Bloc’h, J.; Siliart, B.; Dumon, H. Liver lipid metabolism. J. Anim. Physiol. Anim. Nutr. 2008, 92, 272–283. [Google Scholar] [CrossRef] [PubMed]
- Baumgard, L.H.; Wheelock, J.B.; Sanders, S.R.; Moore, C.E.; Green, H.B.; Waldron, M.R.; Rhoads, R.P. Postabsorptive carbohydrate adaptations to heat stress and monensin supplementation in lactating Holstein cows. J. Dairy Sci. 2011, 94, 5620–5633. [Google Scholar] [CrossRef]
- Zhang, Q.; Ke, Y.; Hong, H. HDL and Lipid Metabolism. Adv. Exp. Med. Biol. 2022, 1377, 49–61. [Google Scholar] [CrossRef]
- Mandell, B.F. Alanine aminotransferase: A nonspecific marker of liver disease. Arch. Intern. Med. 1992, 152, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Otto-Ślusarczyk, D.; Graboń, W.; Mielczarek-Puta, M. Aspartate aminotransferase–key enzyme in the human systemic metabolism. Postepy. Hig. Med. Dosw. 2016, 70, 219–230. [Google Scholar] [CrossRef]
- Jing, J.; Zeng, H.; Shao, Q.; Tang, J.; Wang, L.; Jia, G.; Liu, G.; Chen, X.; Tian, G.; Cai, J.; et al. Selenomethionine alleviates environmental heat stress induced hepatic lipid accumulation and glycogen infiltration of broilers via maintaining mitochondrial and endoplasmic reticulum homeostasis. Redox Biol. 2023, 67, 102912. [Google Scholar] [CrossRef]
- Jing, J.; He, Y.; Liu, Y.; Tang, J.; Wang, L.; Jia, G.; Liu, G.; Chen, X.; Tian, G.; Cai, J.; et al. Selenoproteins synergistically protect porcine skeletal muscle from oxidative damage via relieving mitochondrial dysfunction and endoplasmic reticulum stress. J. Anim. Sci. Biotechnol. 2023, 14, 79. [Google Scholar] [CrossRef]
- Lu, Z.; He, X.F.; Ma, B.B.; Zhang, L.; Li, J.L.; Jiang, Y.; Zhou, G.H.; Gao, F. Increased fat synthesis and limited apolipoprotein B cause lipid accumulation in the liver of broiler chickens exposed to chronic heat stress. Poult. Sci. 2019, 98, 3695–3704. [Google Scholar] [CrossRef]
- Richter, K.; Haslbeck, M.; Buchner, J. The heat shock response: Life on the verge of death. Mol. Cell 2010, 40, 253–266. [Google Scholar] [CrossRef]
- Hartl, F.U.; Bracher, A.; Hayer-Hartl, M. Molecular chaperones in protein folding and proteostasis. Nature 2011, 475, 324–332. [Google Scholar] [CrossRef] [PubMed]
- Mujahid, A.; Yoshiki, Y.; Akiba, Y.; Toyomizu, M. Superoxide radical production in chicken skeletal muscle induced by acute heat stress. Poult. Sci. 2005, 84, 307–314. [Google Scholar] [CrossRef]
- Zheng, Y.; Xie, T.; Li, S.; Wang, W.; Wang, Y.; Cao, Z.; Yang, H. Effects of selenium as a dietary source on performance, inflammation, cell damage, and reproduction of livestock induced by heat stress: A review. Front. Immunol. 2021, 12, 820853. [Google Scholar] [CrossRef] [PubMed]
- De Antonio, J.; Fernandez-Alarcon, M.F.; Lunedo, R.; Squassoni, G.H.; Ferraz, A.L.; Macari, M.; Furlan, R.L.; Furlan, L.R. Chronic heat stress and feed restriction affects carcass composition and the expression of genes involved in the control of fat deposition in broilers. J. Agric. Sci. 2017, 155, 1487–1496. [Google Scholar] [CrossRef]
- Ameer, F.; Scandiuzzi, L.; Hasnain, S.; Kalbacher, H.; Zaidi, N. De novo lipogenesis in health and disease. Metabolism 2014, 63, 895–902. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.H.; Ginsberg, H.N. The role of acyl-CoA: Diacylglycerol acyltransferase (DGAT) in energy metabolism. Ann. Med. 2004, 36, 252–261. [Google Scholar] [CrossRef]
- Olivecrona, T.; Bengtsson, O. Lipoprotein lipase and hepatic lipase. Curr. Opin. Liodiol. 1993, 4, 187–196. [Google Scholar] [CrossRef]
- Li, T.; Guo, W.; Zhou, Z. Adipose triglyceride lipase in hepatic physiology and pathophysiology. Biomolecules 2021, 12, 57. [Google Scholar] [CrossRef]
- Waite, M.; Wakil, S.J. Studies on the mechanism of fatty acid synthesis. XII. Acetyl coenzyme A carboxylase. J. Biol. Chem. 1962, 237, 2750–2757. [Google Scholar] [CrossRef]
- Eberlé, D.; Hegarty, B.; Bossard, P.; Ferré, P.; Foufelle, F. SREBP transcription factors: Master regulators of lipid homeostasis. Biochimie 2004, 86, 839–848. [Google Scholar] [CrossRef] [PubMed]
- Günenc, A.N.; Graf, B.; Stark, H.; Chari, A. Fatty acid synthase: Structure, function, and regulation. In Macromolecular Protein Complexes IV; Subcellular Biochemistry Book Series; Springer: Cham, Switzerland, 2022; Volume 99, pp. 1–33. [Google Scholar] [CrossRef]
- Cohen, P.; Friedman, J.M. Leptin and the control of metabolism: Role for stearoyl-CoA desaturase-1 (SCD-1). J. Nutr. 2004, 134, 2455s–2463s. [Google Scholar] [CrossRef]
- Nelson, D.W.; Gao, Y.; Yen, M.I.; Yen, C.L. Intestine-specific deletion of acyl-CoA: Monoacylglycerol acyltransferase (MGAT) 2 protects mice from diet-induced obesity and glucose intolerance. J. Biol. Chem. 2014, 289, 17338–17349. [Google Scholar] [CrossRef]
- Woeltje, K.F.; Esser, V.; Weis, B.C.; Cox, W.F.; Schroeder, J.G.; Liao, S.T.; Foster, D.W.; McGarry, J.D. Inter-tissue and inter-species characteristics of the mitochondrial carnitine palmitoyltransferase enzyme system. J. Biol. Chem. 1990, 265, 10714–10719. [Google Scholar] [CrossRef]
- Adeva-Andany, M.M.; Carneiro-Freire, N.; Seco-Filgueira, M.; Fernández-Fernández, C.; Mouriño-Bayolo, D. Mitochondrial β-oxidation of saturated fatty acids in humans. Mitochondrion 2019, 46, 73–90. [Google Scholar] [CrossRef]
- van Weeghel, M.; Brinke, H.T.; van Lenthe, H.; Kulik, W.; Minkler, P.E.; Stoll, M.S.; Sass, J.O.; Janssen, U.; Stoffel, W.; Schwab, K.O.; et al. Functional redundancy of mitochondrial enoyl-CoA isomerases in the oxidation of unsaturated fatty acids. FASEB J. 2012, 26, 4316–4326. [Google Scholar] [CrossRef]
- Kimura, C.; Mizugaki, M.; Yamanaka, H.; Fujino, M.; Morishima, T. 2,4-Dienoyl-CoA reductases: From discovery toward pathophysiological significance. Nihon Rinsho 2004, 62, 1577–1583. [Google Scholar]
- Andresen, B.S.; Olpin, S.; Poorthuis, B.J.; Scholte, H.R.; Vianey-Saban, C.; Wanders, R.; Ijlst, L.; Morris, A.; Pourfarzam, M.; Bartlett, K.; et al. Clear correlation of genotype with disease phenotype in very-long-chain acyl-CoA dehydrogenase deficiency. Am. J. Hum. Genet. 1999, 64, 479–494. [Google Scholar] [CrossRef] [PubMed]
- Nido, S.A.; Shituleni, S.A.; Mengistu, B.M.; Liu, Y.; Khan, A.Z.; Gan, F.; Kumbhar, S.; Huang, K. Effects of selenium-enriched probiotics on lipid metabolism, antioxidative status, histopathological lesions, and related gene expression in mice fed a high-fat diet. Biol. Trace Elem. Res. 2016, 171, 399–409. [Google Scholar] [CrossRef]
- Ibrahim, I.M.; Abdelmalek, D.H.; Elfiky, A.A. GRP78: A cell’s response to stress. Life Sci. 2019, 226, 156–163. [Google Scholar] [CrossRef]
- Liu, Y.; Yin, S.; He, Y.; Tang, J.; Pu, J.; Jia, G.; Liu, G.; Tian, G.; Chen, X.; Cai, J.; et al. Hydroxy-Selenomethionine mitigated chronic heat stress-induced porcine splenic damage via activation of Nrf2/Keap1 signal and suppression of NFκb and STAT signal. Int. J. Mol. Sci. 2023, 24, 6461. [Google Scholar] [CrossRef]
- Cai, J.; Huang, J.; Yang, J.; Chen, X.; Zhang, H.; Zhu, Y.; Liu, Q.; Zhang, Z. The protective effect of selenoprotein M on non-alcoholic fatty liver disease: The role of the AMPKα1-MFN2 pathway and Parkin mitophagy. Cell. Mol. Life Sci. 2022, 79, 354. [Google Scholar] [CrossRef]
- Pitts, M.W.; Hoffmann, P.R. Endoplasmic reticulum-resident selenoproteins as regulators of calcium signaling and homeostasis. Cell Calcium 2018, 70, 76–86. [Google Scholar] [CrossRef]
- Ma, Y.M.; Guo, Y.Z.; Ibeanu, G.; Wang, L.Y.; Dong, J.D.; Wang, J.; Jing, L.; Zhang, J.Z.; Li, P.A. Overexpression of selenoprotein H prevents mitochondrial dynamic imbalance induced by glutamate exposure. Int. J. Biol. Sci. 2017, 13, 1458–1469. [Google Scholar] [CrossRef]
- Jiang, Y.Y.; Huang, J.Q.; Lin, G.C.; Guo, H.Y.; Ren, F.Z.; Zhang, H. Characterization and expression of chicken selenoprotein U. Biol. Trace Elem. Res. 2015, 166, 216–224. [Google Scholar] [CrossRef]
- Xu, X.M.; Carlson, B.A.; Irons, R.; Mix, H.; Zhong, N.; Gladyshev, V.N.; Hatfield, D.L. Selenophosphate synthetase 2 is essential for selenoprotein biosynthesis. Biochem. J. 2007, 404, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Shchedrina, V.A.; Everley, R.A.; Zhang, Y.; Gygi, S.P.; Hatfield, D.L.; Gladyshev, V.N. Selenoprotein K binds multiprotein complexes and is involved in the regulation of endoplasmic reticulum homeostasis. J. Biol. Chem. 2011, 286, 42937–42948. [Google Scholar] [CrossRef] [PubMed]
- Han, S.J.; Lee, B.C.; Yim, S.H.; Gladyshev, V.N.; Lee, S.R. Characterization of mammalian selenoprotein o: A redox-active mitochondrial protein. PLoS ONE 2014, 9, e95518. [Google Scholar] [CrossRef] [PubMed]
- Lillig, C.H.; Holmgren, A. Thioredoxin and related molecules--from biology to health and disease. Antioxid. Redox Signal. 2007, 9, 25–47. [Google Scholar] [CrossRef]
- Imai, H.; Nakagawa, Y. Biological significance of phospholipid hydroperoxide glutathione peroxidase (PHGPx, GPx4) in mammalian cells. Free Radic. Biol. Med. 2003, 34, 145–169. [Google Scholar] [CrossRef]
- Quintana-Cabrera, R.; Fernandez-Fernandez, S.; Bobo-Jimenez, V.; Escobar, J.; Sastre, J.; Almeida, A.; Bolaños, J.P. γ-Glutamylcysteine detoxifies reactive oxygen species by acting as glutathione peroxidase-1 cofactor. Nat. Commun. 2012, 3, 718. [Google Scholar] [CrossRef]
- Gong, T.; Hashimoto, A.C.; Sasuclark, A.R.; Khadka, V.S.; Gurary, A.; Pitts, M.W. Selenoprotein M promotes hypothalamic leptin signaling and thioredoxin antioxidant activity. Antioxid. Redox Signal. 2021, 35, 775–787. [Google Scholar] [CrossRef]
- Ferguson, A.D.; Labunskyy, V.M.; Fomenko, D.E.; Araç, D.; Chelliah, Y.; Amezcua, C.A.; Rizo, J.; Gladyshev, V.N.; Deisenhofer, J. NMR structures of the selenoproteins Sep15 and SelM reveal redox activity of a new thioredoxin-like family. J. Biol. Chem. 2006, 281, 3536–3543. [Google Scholar] [CrossRef]
- Liu, Y.; Yin, S.; Tang, J.; Liu, Y.; Jia, G.; Liu, G.; Tian, G.; Chen, X.; Cai, J.; Kang, B.; et al. Hydroxy selenomethionine improves meat quality through optimal skeletal metabolism and functions of selenoproteins of pigs under chronic heat stress. Antioxidants 2021, 10, 1558. [Google Scholar] [CrossRef]
- Briens, M.; Mercier, Y.; Rouffineau, F.; Mercerand, F.; Geraert, P.A. 2-Hydroxy-4-methylselenobutanoic acid induces additional tissue selenium enrichment in broiler chickens compared with other selenium sources. Poult. Sci. 2014, 93, 85–93. [Google Scholar] [CrossRef]
- China Agricultural Industry Standard NY/T33-2004; Nutrient Requirement of Broilers. China Agriculture Press: Beijing, China, 2012. (In Chinese)
- National Research Council. Nutrient Requirements of Poultry; National Academy Press: Washington, DC, USA, 1994. [Google Scholar]
- GB 5009.93-2010; National Food Safety Standard Determination of Selenium in Foods. Ministry of Health: Beijing, China, 2010.
- Zhao, H.; Li, K.; Tang, J.Y.; Zhou, J.C.; Wang, K.N.; Xia, X.J.; Lei, X.G. Expression of selenoprotein genes is affected by obesity of pigs fed a high-fat diet. J. Nutr. 2015, 145, 1394–1401. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Groups | SEM | p-Value 1 | p-Value 2 | |||||
|---|---|---|---|---|---|---|---|---|---|
| CON | HS | HS + SS | HS + SeY | HS + SeM | HS + Nano-Se | ||||
| ALT/(U/L) | 11.05 * | 15.46 | 13.80 | 12.71 | 12.30 | 13.68 | 0.77 | 0.019 | 0.775 |
| AST/(U/L) | 336.04 | 365.50 b | 286.45 ab | 283.33 a | 283.97 a | 276.19 a | 10.25 | 0.310 | 0.016 |
| TC/(mmol/L) | 3.14 | 2.68 | 2.66 | 2.78 | 2.82 | 2.80 | 0.10 | 0.095 | 0.983 |
| TG/(mmol/L) | 0.42 | 0.95 | 0.75 | 0.82 | 0.74 | 0.98 | 0.60 | 0.066 | 0.338 |
| HDL-C/(mmol/L) | 2.28 ** | 1.50 | 1.69 | 1.68 | 1.76 | 1.68 | 0.06 | 0.007 | 0.807 |
| LDL-C/(mmol/L) | 0.45 | 0.44 | 0.34 | 0.36 | 0.28 | 0.43 | 0.03 | 0.937 | 0.198 |
| NEFA/(mmol/L) | 0.78 *** | 0.31 a | 0.51 b | 0.50 b | 0.42 ab | 0.47 b | 0.02 | <0.001 | 0.026 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Jing, J.; Gong, Z.; Tang, J.; Wang, L.; Jia, G.; Liu, G.; Chen, X.; Tian, G.; Cai, J.; et al. Different Dietary Sources of Selenium Alleviate Hepatic Lipid Metabolism Disorder of Heat-Stressed Broilers by Relieving Endoplasmic Reticulum Stress. Int. J. Mol. Sci. 2023, 24, 15443. https://doi.org/10.3390/ijms242015443
Wang J, Jing J, Gong Z, Tang J, Wang L, Jia G, Liu G, Chen X, Tian G, Cai J, et al. Different Dietary Sources of Selenium Alleviate Hepatic Lipid Metabolism Disorder of Heat-Stressed Broilers by Relieving Endoplasmic Reticulum Stress. International Journal of Molecular Sciences. 2023; 24(20):15443. https://doi.org/10.3390/ijms242015443
Chicago/Turabian StyleWang, Jiayi, Jinzhong Jing, Zhengyi Gong, Jiayong Tang, Longqiong Wang, Gang Jia, Guangmang Liu, Xiaoling Chen, Gang Tian, Jingyi Cai, and et al. 2023. "Different Dietary Sources of Selenium Alleviate Hepatic Lipid Metabolism Disorder of Heat-Stressed Broilers by Relieving Endoplasmic Reticulum Stress" International Journal of Molecular Sciences 24, no. 20: 15443. https://doi.org/10.3390/ijms242015443
APA StyleWang, J., Jing, J., Gong, Z., Tang, J., Wang, L., Jia, G., Liu, G., Chen, X., Tian, G., Cai, J., Kang, B., Che, L., & Zhao, H. (2023). Different Dietary Sources of Selenium Alleviate Hepatic Lipid Metabolism Disorder of Heat-Stressed Broilers by Relieving Endoplasmic Reticulum Stress. International Journal of Molecular Sciences, 24(20), 15443. https://doi.org/10.3390/ijms242015443