
Effect of Long-Term Low-Dose Arsenic Exposure on DNA Methylation and Gene Expression in Human Liver Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
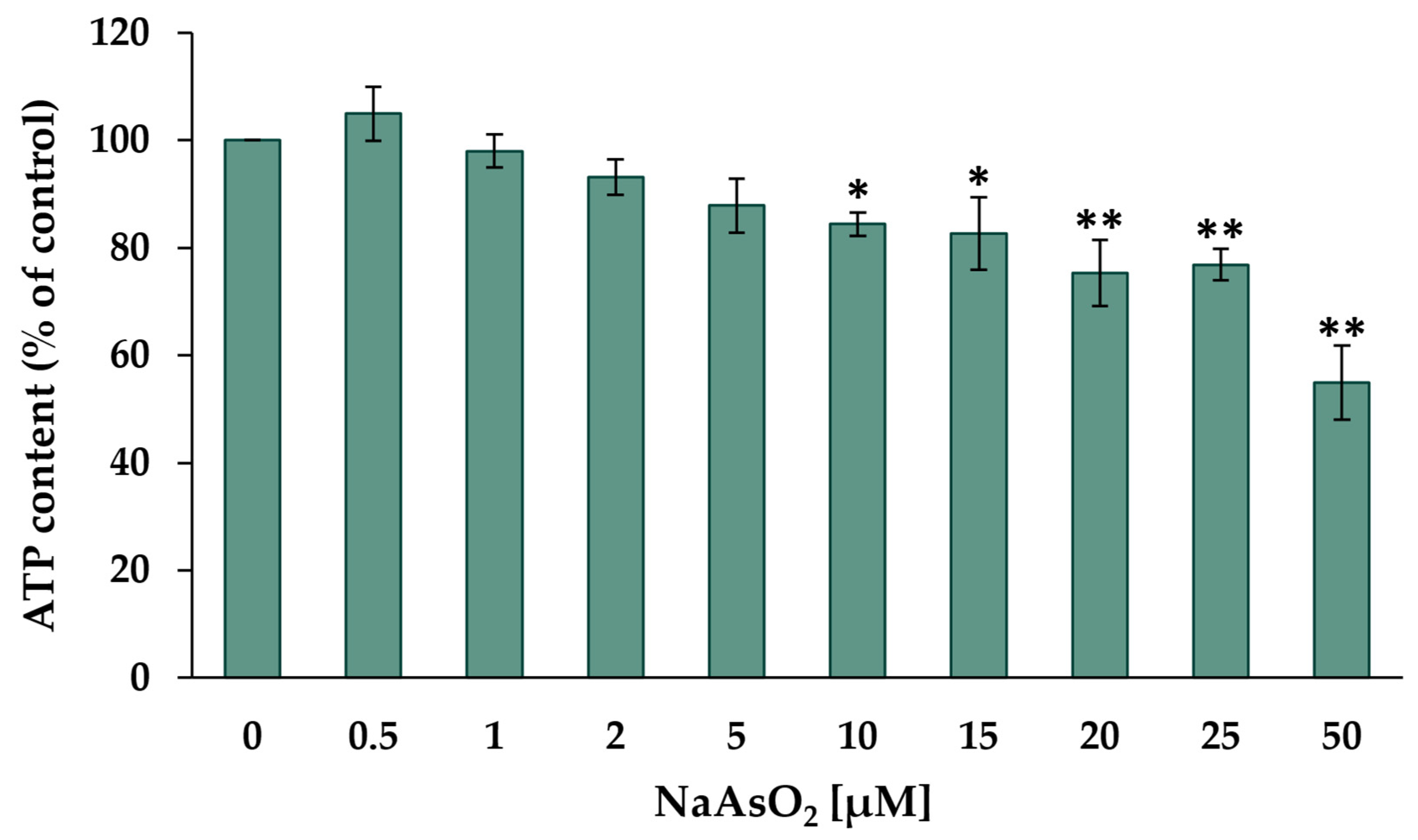
2.1. Cytotoxicity
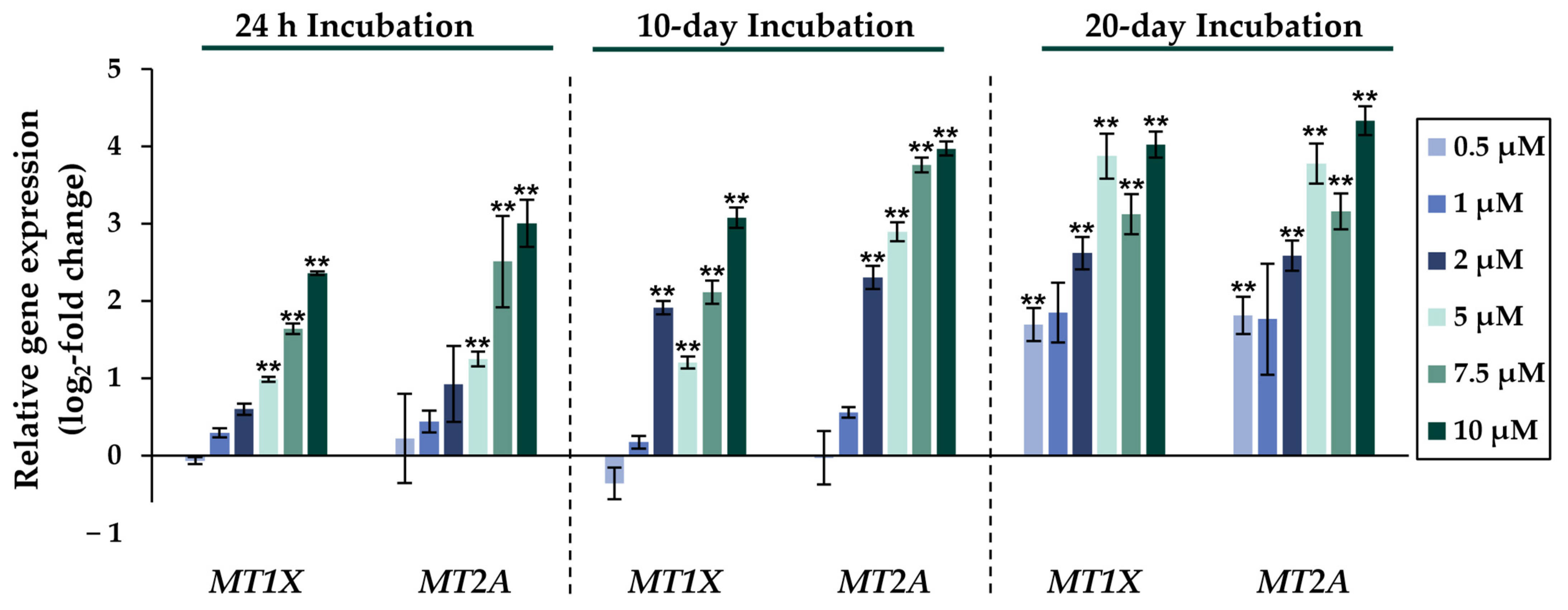
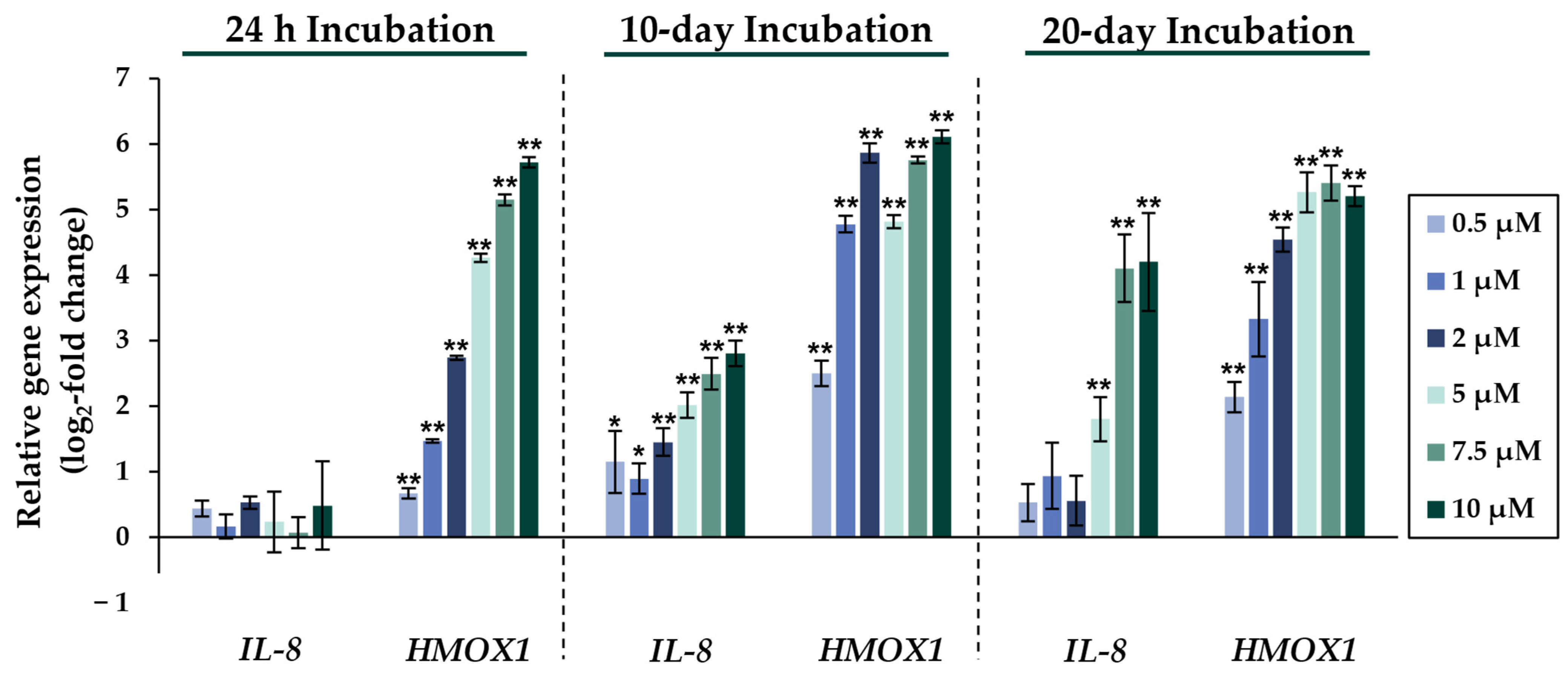
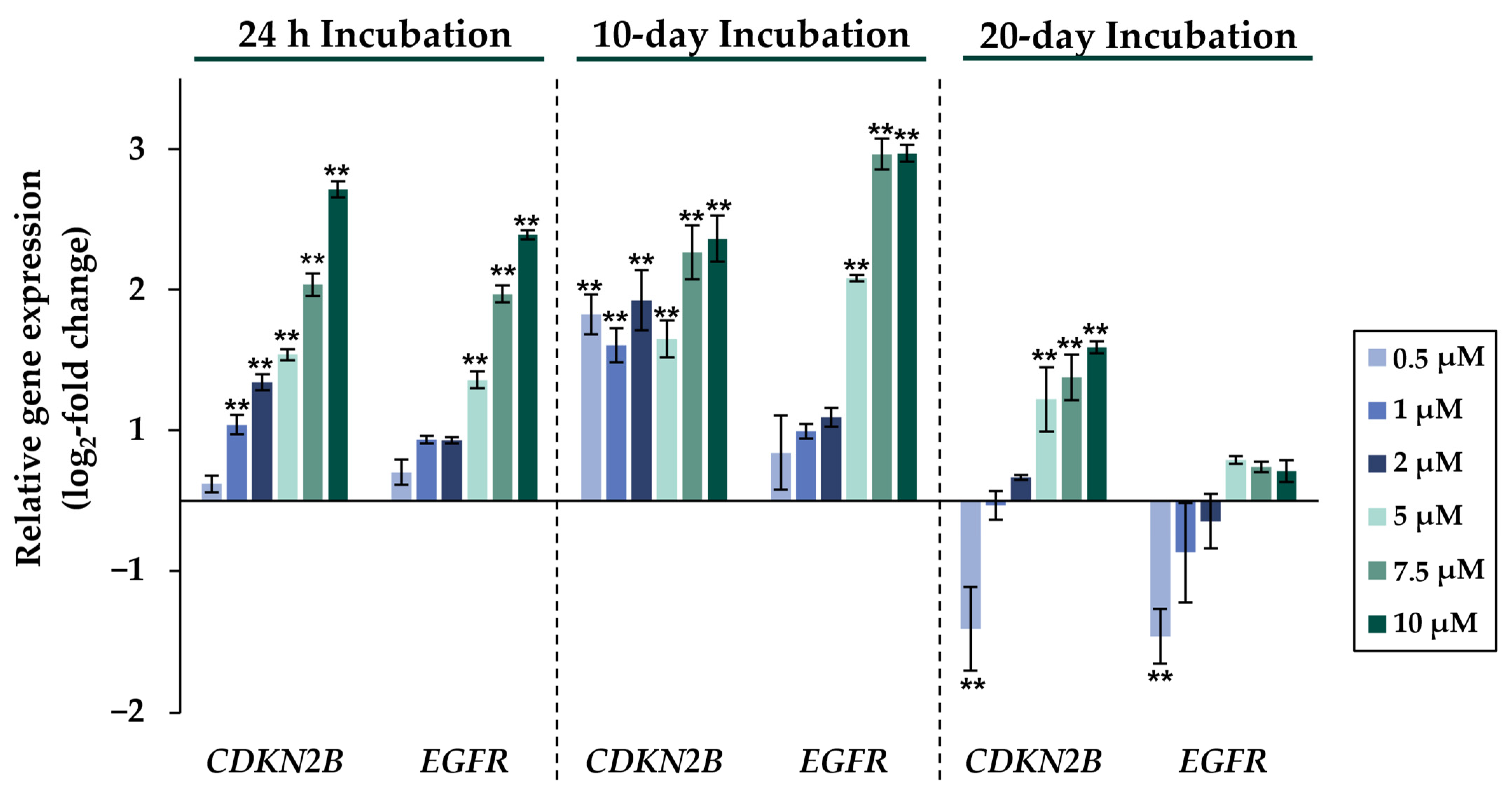
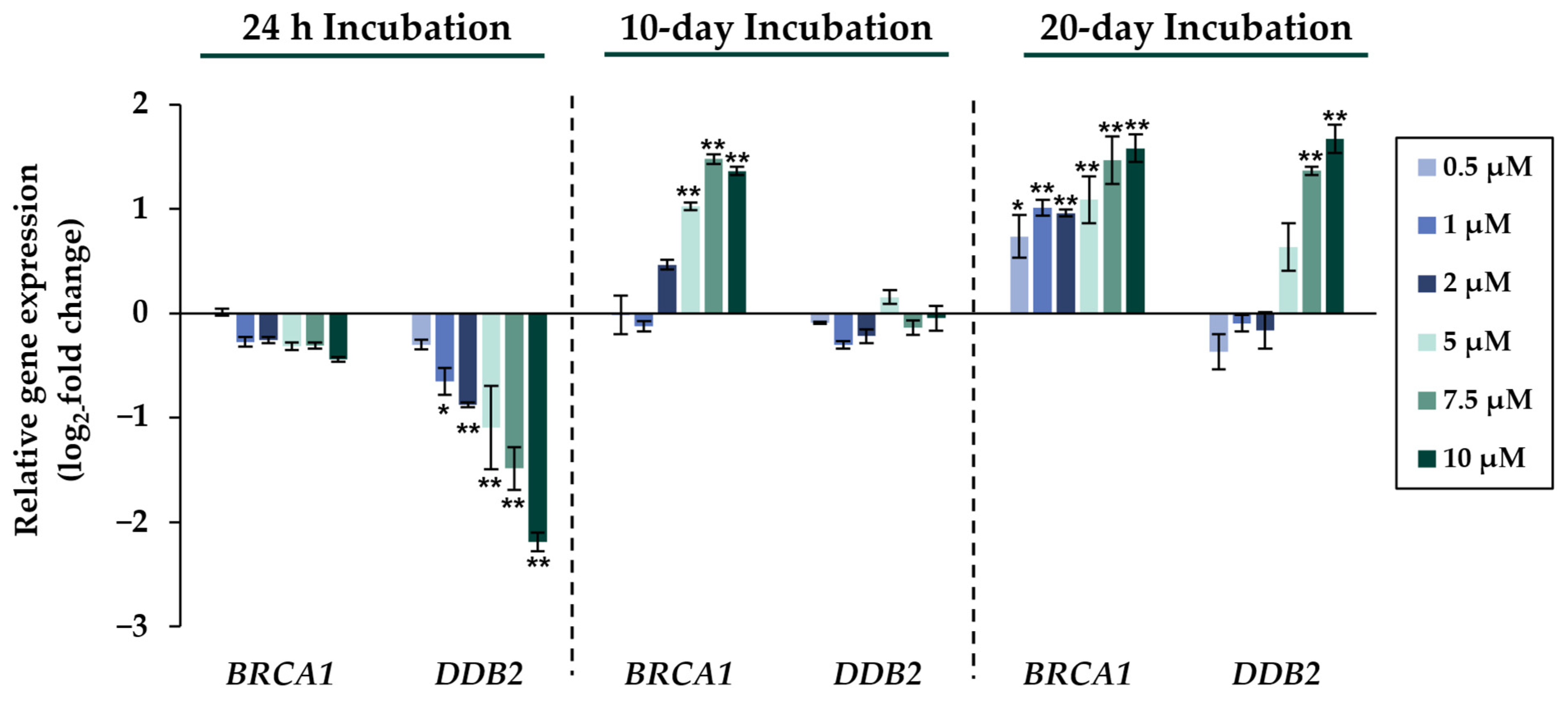
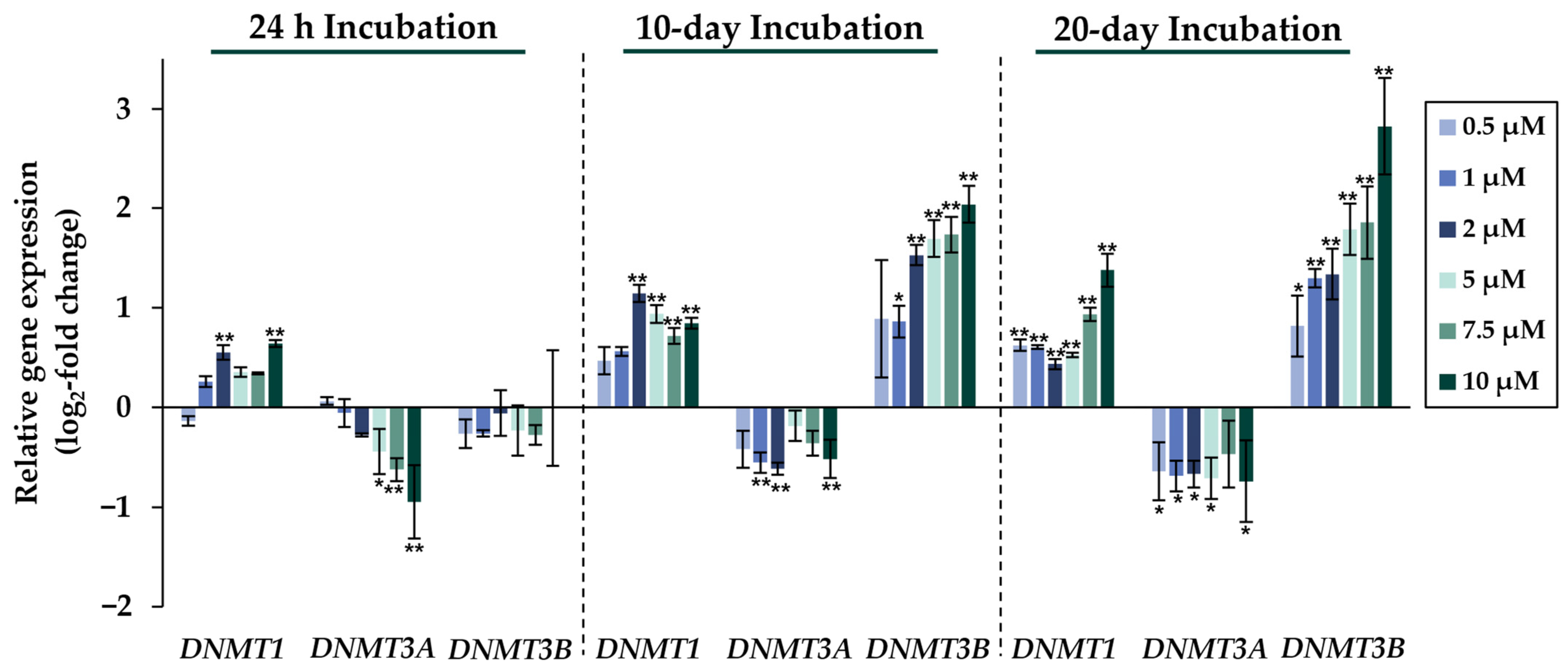
2.2. Gene Expression Analysis
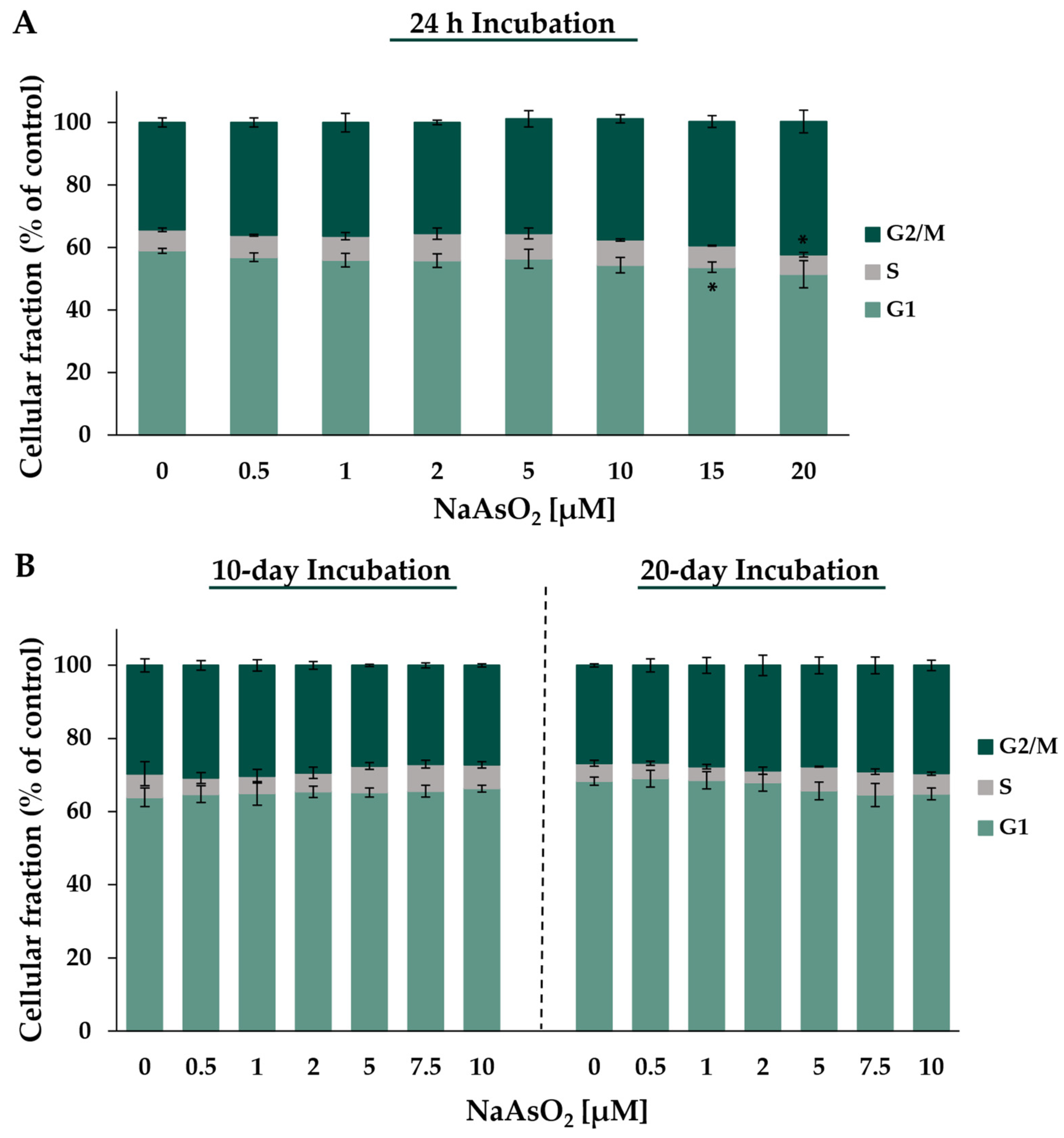
2.3. Cell Cycle Regulation
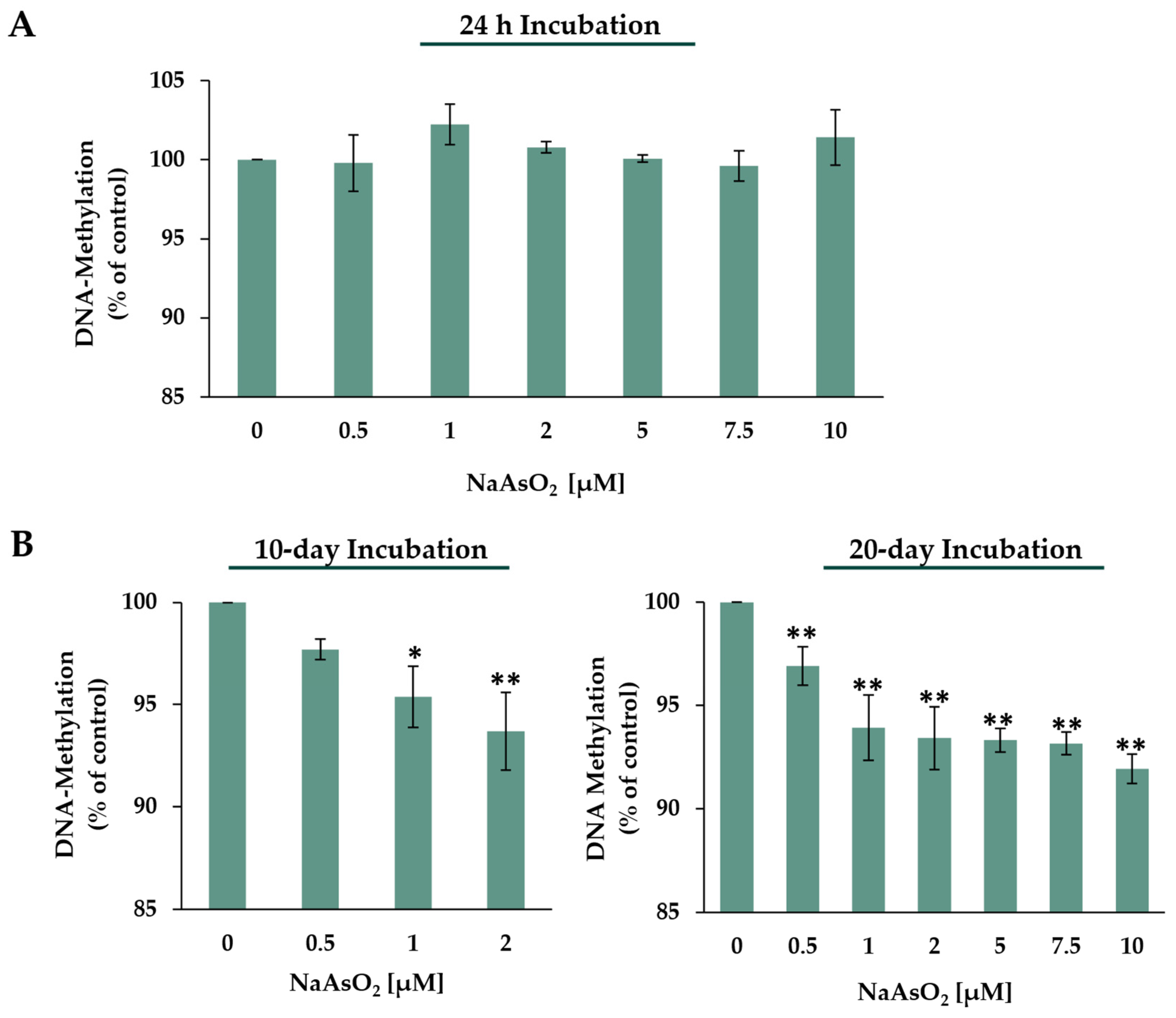
2.4. DNA Methylation
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture and Drug Treatment
4.3. Cytotoxicity Assays
4.4. High-Throughput RT-qPCR
4.5. Analysis of Cell Cycle Distribution via Flow Cytometry
4.6. Quantification of Methyl-dCMP Content
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| As3MT | arsenite methyltransferase |
| dCMP | desoxycytidin-5′-monophosphat |
| DMA III | dimethylarsinic acid |
| DNMT1 | DNA methyltransferase 1 |
| DNMT3A | DNA methyltransferase 3A |
| DNMT3B | DNA methyltransferase 3B |
| GSH | glutathione |
| HPLC | high-performance liquid chromatography |
| IARC | International Agency for Research on Cancer |
| MMA III | monomethylarsonic acid |
| MT | metallothionein |
| SAM | S-adenosylmethionine |
| TXN | thioredoxin |
References
- Zhou, Q.; Xi, S. A review on arsenic carcinogenesis: Epidemiology, metabolism, genotoxicity and epigenetic changes. Regul. Toxicol. Pharmacol. 2018, 99, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.W.; Lee, C.H.; Yu, H.S. Arsenic-Induced Carcinogenesis and Immune Dysregulation. Int. J. Environ. Res. Public Health 2019, 16, 2746. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.Q.; Hua, H.Y.; Naranmandura, H.; Zhu, H.H. Balance between the toxicity and anticancer activity of arsenic trioxide in treatment of acute promyelocytic leukemia. Toxicol. Appl. Pharmacol. 2020, 409, 115299. [Google Scholar] [CrossRef]
- IARC. Arsenic, metals, fibres, and dusts. IARC Monogr. Eval. Carcinog. Risks Hum. 2012, 100, 11–465. [Google Scholar]
- Greim, H. Arsenic and arsenic compounds (with the exception of arsine). MAK Value Documentation. In The MAK-Collection for Occupational Health and Safety. Part I: MAK Value Documentations; Wiley-VCH: Weinheim, Germany, 2002; Volume 21, pp. 49–106. [Google Scholar]
- Rossman, T.G. Mechanism of arsenic carcinogenesis: An integrated approach. Mutat. Res. 2003, 533, 37–65. [Google Scholar] [CrossRef]
- Chen, H.; Liu, J.; Merrick, B.A.; Waalkes, M.P. Genetic events associated with arsenic-induced malignant transformation: Applications of cDNA microarray technology. Mol. Carcinog. 2001, 30, 79–87. [Google Scholar] [CrossRef]
- Speer, R.M.; Nandi, S.P.; Cooper, K.L.; Zhou, X.; Yu, H.; Guo, Y.; Hudson, L.G.; Alexandrov, L.B.; Liu, K.J. Arsenic is a potent co-mutagen of ultraviolet light. bioRxiv 2023. preprint. [Google Scholar] [CrossRef]
- Riedmann, C.; Ma, Y.; Melikishvili, M.; Godfrey, S.G.; Zhang, Z.; Chen, K.C.; Rouchka, E.C.; Fondufe-Mittendorf, Y.N. Inorganic Arsenic-induced cellular transformation is coupled with genome wide changes in chromatin structure, transcriptome and splicing patterns. BMC Genom. 2015, 16, 212. [Google Scholar] [CrossRef]
- Liu, J.; Yu, L.; Tokar, E.J.; Bortner, C.; Sifre, M.I.; Sun, Y.; Waalkes, M.P. Arsenic-induced aberrant gene expression in fetal mouse primary liver-cell cultures. Ann. N. Y. Acad. Sci. 2008, 1140, 368–375. [Google Scholar] [CrossRef]
- Reichard, J.F.; Puga, A. Effects of arsenic exposure on DNA methylation and epigenetic gene regulation. Epigenomics 2010, 2, 87–104. [Google Scholar] [CrossRef]
- Zhang, W.; Xu, J. DNA methyltransferases and their roles in tumorigenesis. Biomark. Res. 2017, 5, 1. [Google Scholar] [CrossRef] [PubMed]
- Del Castillo Falconi, V.M.; Torres-Arciga, K.; Matus-Ortega, G.; Diaz-Chavez, J.; Herrera, L.A. DNA Methyltransferases: From Evolution to Clinical Applications. Int. J. Mol. Sci. 2022, 23, 8994. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Tan, J.; Wang, M.; Wen, W.; He, Y. Inorganic arsenic-mediated upregulation of AS3MT promotes proliferation of nonsmall cell lung cancer cells by regulating cell cycle genes. Environ. Toxicol. 2021, 36, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Medda, N.; De, S.K.; Maiti, S. Different mechanisms of arsenic related signaling in cellular proliferation, apoptosis and neo-plastic transformation. Ecotoxicol. Environ. Saf. 2021, 208, 111752. [Google Scholar] [CrossRef]
- Fischer, B.M.; Neumann, D.; Piberger, A.L.; Risnes, S.F.; Koeberle, B.; Hartwig, A. Use of high-throughput RT-qPCR to assess modulations of gene expression profiles related to genomic stability and interactions by cadmium. Arch. Toxicol. 2016, 90, 2745–2761. [Google Scholar] [CrossRef]
- Drexler, H. Arsenic and inorganic arsenic compounds (with the exception of arsenic hydride and its salts). BAT Value Documentation. In The MAK-Collection for Occupational Health and Safety. Part II: BAT Value Documentations; Drexler, H., Greim, H., Eds.; Wiley-VCH: Weinheim, Germany, 2003; Volume 4, pp. 171–178. [Google Scholar]
- Hata, A.; Yamanaka, K.; Habib, M.A.; Endo, Y.; Fujitani, N.; Endo, G. Arsenic speciation analysis of urine samples from individuals living in an arsenic-contaminated area in Bangladesh. Environ. Health Prev. Med. 2012, 17, 235–245. [Google Scholar] [CrossRef]
- Zhao, C.Q.; Young, M.R.; Diwan, B.A.; Coogan, T.P.; Waalkes, M.P. Association of arsenic-induced malignant transformation with DNA hypomethylation and aberrant gene expression. Proc. Natl. Acad. Sci. USA 1997, 94, 10907–10912. [Google Scholar] [CrossRef]
- Caudill, M.A.; Wang, J.C.; Melnyk, S.; Pogribny, I.P.; Jernigan, S.; Collins, M.D.; Santos-Guzman, J.; Swendseid, M.E.; Cogger, E.A.; James, S.J. Intracellular S-adenosylhomocysteine concentrations predict global DNA hypomethylation in tissues of methyl-deficient cystathionine beta-synthase heterozygous mice. J. Nutr. 2001, 131, 2811–2818. [Google Scholar] [CrossRef]
- Wu, L.; Li, H.; Ye, F.; Wei, Y.; Li, W.; Xu, Y.; Xia, H.; Zhang, J.; Guo, L.; Zhang, G.; et al. As3MT-mediated SAM consumption, which inhibits the methylation of histones and LINE1, is involved in arsenic-induced male reproductive damage. Environ. Pollut. 2022, 313, 120090. [Google Scholar] [CrossRef]
- Drobna, Z.; Waters, S.B.; Walton, F.S.; LeCluyse, E.L.; Thomas, D.J.; Styblo, M. Interindividual variation in the metabolism of arsenic in cultured primary human hepatocytes. Toxicol. Appl. Pharmacol. 2004, 201, 166–177. [Google Scholar] [CrossRef]
- Drobna, Z.; Xing, W.; Thomas, D.J.; Styblo, M. shRNA silencing of AS3MT expression minimizes arsenic methylation capacity of HepG2 cells. Chem. Res. Toxicol. 2006, 19, 894–898. [Google Scholar] [CrossRef][Green Version]
- Watanabe, T.; Ohta, Y.; Mizumura, A.; Kobayashi, Y.; Hirano, S. Analysis of arsenic metabolites in HepG2 and AS3MT-transfected cells. Arch. Toxicol. 2011, 85, 577–588. [Google Scholar] [CrossRef]
- Hartwig, A.; Pelzer, A.; Asmuss, M.; Buerkle, A. Very low concentrations of arsenite suppress poly(ADP-ribosyl)ation in mammalian cells. Int. J. Cancer 2003, 104, 1–6. [Google Scholar] [CrossRef]
- Yeh, J.Y.; Cheng, L.C.; Liang, Y.C.; Ou, B.R. Modulation of the arsenic effects on cytotoxicity, viability, and cell cycle in porcine endothelial cells by selenium. Endothelium 2003, 10, 127–139. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, M.J.; Shaki, F.; Ghazi-Khansari, M.; Pourahmad, J. Toxicity of Arsenic (III) on Isolated Liver Mitochondria: A New Mechanistic Approach. Iran. J. Pharm. Res. 2013, 12, 121–138. [Google Scholar] [PubMed]
- Ronchetti, S.A.; Bianchi, M.S.; Duvilanski, B.H.; Cabilla, J.P. In Vivo and In Vitro Arsenic Exposition Induces Oxidative Stress in Anterior Pituitary Gland. Int. J. Toxicol. 2016, 35, 463–475. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Wang, Y.F.; Liu, J.; Jiang, B.H.; Liu, L.Z. Human endothelial cells promote arsenic-transformed lung epithelial cells to induce tumor growth and angiogenesis through interleukin-8 induction. Aging 2022, 14, 2113–2130. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zhao, Y.; Xu, W.; Luo, F.; Wang, B.; Li, Y.; Pang, Y.; Liu, Q. Involvement of HIF-2alpha-mediated inflammation in arsenite-induced transformation of human bronchial epithelial cells. Toxicol. Appl. Pharmacol. 2013, 272, 542–550. [Google Scholar] [CrossRef]
- Guenther, V.; Lindert, U.; Schaffner, W. The taste of heavy metals: Gene regulation by MTF-1. Biochim. Biophys. Acta 2012, 1823, 1416–1425. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, X.W.; Shu, L.S.; Liu, C.D. The correlation and role analysis of SLC30A1 and SLC30A10 in cervical carcinoma. J. Cancer 2022, 13, 1031–1047. [Google Scholar] [CrossRef]
- Kim, C.; States, J.C.; Ceresa, B.P. Chronic and acute arsenic exposure enhance EGFR expression via distinct molecular mechanisms. Toxicol. In Vitro 2020, 67, 104925. [Google Scholar] [CrossRef] [PubMed]
- Yih, L.H.; Hsueh, S.W.; Luu, W.S.; Chiu, T.H.; Lee, T.C. Arsenite induces prominent mitotic arrest via inhibition of G2 checkpoint activation in CGL-2 cells. Carcinogenesis 2005, 26, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Jiao, G.; Ren, T.; Guo, W.; Ren, C.; Yang, K. Arsenic trioxide inhibits growth of human chondrosarcoma cells through G2/M arrest and apoptosis as well as autophagy. Tumour Biol. 2015, 36, 3969–3977. [Google Scholar] [CrossRef] [PubMed]
- Ganapathy, S.; Liu, J.; Xiong, R.; Yu, T.; Makriyannis, A.; Chen, C. Chronic low dose arsenic exposure preferentially perturbs mitotic phase of the cell cycle. Genes. Cancer 2019, 10, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Andrew, A.S.; Karagas, M.R.; Hamilton, J.W. Decreased DNA repair gene expression among individuals exposed to arsenic in United States drinking water. Int. J. Cancer 2003, 104, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.B.; Vahter, M.; Concha, G.; Broberg, K. Environmental arsenic exposure and DNA methylation of the tumor suppressor gene p16 and the DNA repair gene MLH1: Effect of arsenic metabolism and genotype. Metallomics 2012, 4, 1167–1175. [Google Scholar] [CrossRef]
- Kato, Y.; Kaneda, M.; Hata, K.; Kumaki, K.; Hisano, M.; Kohara, Y.; Okano, M.; Li, E.; Nozaki, M.; Sasaki, H. Role of the Dnmt3 family in de novo methylation of imprinted and repetitive sequences during male germ cell development in the mouse. Hum. Mol. Genet. 2007, 16, 2272–2280. [Google Scholar] [CrossRef]
- Okano, M.; Bell, D.W.; Haber, D.A.; Li, E. DNA methyltransferases Dnmt3a and Dnmt3b are essential for de novo methylation and mammalian development. Cell 1999, 99, 247–257. [Google Scholar] [CrossRef]
- Goll, M.G.; Bestor, T.H. Eukaryotic cytosine methyltransferases. Annu. Rev. Biochem. 2005, 74, 481–514. [Google Scholar] [CrossRef]
- Chen, T.; Li, E. Structure and function of eukaryotic DNA methyltransferases. Curr. Top. Dev. Biol. 2004, 60, 55–89. [Google Scholar] [CrossRef]
- Dodge, J.E.; Okano, M.; Dick, F.; Tsujimoto, N.; Chen, T.; Wang, S.; Ueda, Y.; Dyson, N.; Li, E. Inactivation of Dnmt3b in mouse embryonic fibroblasts results in DNA hypomethylation, chromosomal instability, and spontaneous immortalization. J. Biol. Chem. 2005, 280, 17986–17991. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Zhou, Y.; Campbell, S.L.; Le, T.; Li, E.; Sweatt, J.D.; Silva, A.J.; Fan, G. Dnmt1 and Dnmt3a maintain DNA methylation and regulate synaptic function in adult forebrain neurons. Nat. Neurosci. 2010, 13, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Jair, K.W.; Bachman, K.E.; Suzuki, H.; Ting, A.H.; Rhee, I.; Yen, R.W.; Baylin, S.B.; Schuebel, K.E. De novo CpG island methylation in human cancer cells. Cancer Res. 2006, 66, 682–692. [Google Scholar] [CrossRef] [PubMed]
- Feltus, F.A.; Lee, E.K.; Costello, J.F.; Plass, C.; Vertino, P.M. Predicting aberrant CpG island methylation. Proc. Natl. Acad. Sci. USA 2003, 100, 12253–12258. [Google Scholar] [CrossRef]
- Benbrahim-Tallaa, L.; Waterland, R.A.; Styblo, M.; Achanzar, W.E.; Webber, M.M.; Waalkes, M.P. Molecular events associated with arsenic-induced malignant transformation of human prostatic epithelial cells: Aberrant genomic DNA methylation and K-ras oncogene activation. Toxicol. Appl. Pharmacol. 2005, 206, 288–298. [Google Scholar] [CrossRef]
- Cui, X.; Wakai, T.; Shirai, Y.; Yokoyama, N.; Hatakeyama, K.; Hirano, S. Arsenic trioxide inhibits DNA methyltransferase and restores methylation-silenced genes in human liver cancer cells. Hum. Pathol. 2006, 37, 298–311. [Google Scholar] [CrossRef] [PubMed]
- Rea, M.; Eckstein, M.; Eleazer, R.; Smith, C.; Fondufe-Mittendorf, Y.N. Genome-wide DNA methylation reprogramming in response to inorganic arsenic links inhibition of CTCF binding, DNMT expression and cellular transformation. Sci. Rep. 2017, 7, 41474. [Google Scholar] [CrossRef]
- Hartwig, A. Metal interaction with redox regulation: An integrating concept in metal carcinogenesis? Free Radic. Biol. Med. 2013, 55, 63–72. [Google Scholar] [CrossRef]
- Walter, I.; Schwerdtle, T.; Thuy, C.; Parsons, J.L.; Dianov, G.L.; Hartwig, A. Impact of arsenite and its methylated metabolites on PARP-1 activity, PARP-1 gene expression and poly(ADP-ribosyl)ation in cultured human cells. DNA Repair 2007, 6, 61–70. [Google Scholar] [CrossRef]
- Zhou, X.; Sun, X.; Cooper, K.L.; Wang, F.; Liu, K.J.; Hudson, L.G. Arsenite interacts selectively with zinc finger proteins containing C3H1 or C4 motifs. J. Biol. Chem. 2011, 286, 22855–22863. [Google Scholar] [CrossRef]
- Schwerdtle, T.; Walter, I.; Hartwig, A. Arsenite and its biomethylated metabolites interfere with the formation and repair of stable BPDE-induced DNA adducts in human cells and impair XPAzf and Fpg. DNA Repair 2003, 2, 1449–1463. [Google Scholar] [CrossRef] [PubMed]
- Piatek, K.; Schwerdtle, T.; Hartwig, A.; Bal, W. Monomethylarsonous acid destroys a tetrathiolate zinc finger much more efficiently than inorganic arsenite: Mechanistic considerations and consequences for DNA repair inhibition. Chem. Res. Toxicol. 2008, 21, 600–606. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Speer, R.M.; Volk, L.; Hudson, L.G.; Liu, K.J. Arsenic co-carcinogenesis: Inhibition of DNA repair and interaction with zinc finger proteins. Semin. Cancer Biol. 2021, 76, 86–98. [Google Scholar] [CrossRef] [PubMed]
- Jurkowska, R.Z.; Jurkowski, T.P.; Jeltsch, A. Structure and function of mammalian DNA methyltransferases. Chembiochem 2011, 12, 206–222. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, M.; Esteve, P.O.; Chin, H.G.; Samaranayke, M.; Kim, G.D.; Pradhan, S. CXXC domain of human DNMT1 is essential for enzymatic activity. Biochemistry 2008, 47, 10000–10009. [Google Scholar] [CrossRef] [PubMed]
- Yan, N.; Li, Y.; Xing, Y.; Wu, J.; Li, J.; Liang, Y.; Tang, Y.; Wang, Z.; Song, H.; Wang, H.; et al. Developmental arsenic exposure impairs cognition, directly targets DNMT3A, and reduces DNA methylation. EMBO Rep. 2022, 23, e54147. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stößer, S.; Lumpp, T.; Fischer, F.; Gunesch, S.; Schumacher, P.; Hartwig, A. Effect of Long-Term Low-Dose Arsenic Exposure on DNA Methylation and Gene Expression in Human Liver Cells. Int. J. Mol. Sci. 2023, 24, 15238. https://doi.org/10.3390/ijms242015238
Stößer S, Lumpp T, Fischer F, Gunesch S, Schumacher P, Hartwig A. Effect of Long-Term Low-Dose Arsenic Exposure on DNA Methylation and Gene Expression in Human Liver Cells. International Journal of Molecular Sciences. 2023; 24(20):15238. https://doi.org/10.3390/ijms242015238
Chicago/Turabian StyleStößer, Sandra, Tatjana Lumpp, Franziska Fischer, Sarah Gunesch, Paul Schumacher, and Andrea Hartwig. 2023. "Effect of Long-Term Low-Dose Arsenic Exposure on DNA Methylation and Gene Expression in Human Liver Cells" International Journal of Molecular Sciences 24, no. 20: 15238. https://doi.org/10.3390/ijms242015238
APA StyleStößer, S., Lumpp, T., Fischer, F., Gunesch, S., Schumacher, P., & Hartwig, A. (2023). Effect of Long-Term Low-Dose Arsenic Exposure on DNA Methylation and Gene Expression in Human Liver Cells. International Journal of Molecular Sciences, 24(20), 15238. https://doi.org/10.3390/ijms242015238