
Prolactin and Growth Hormone Signaling and Interlink Focused on the Mammosomatotroph Paradigm: A Comprehensive Review of the Literature

 ,
, 

Abstract
1. Introduction
2. Methods
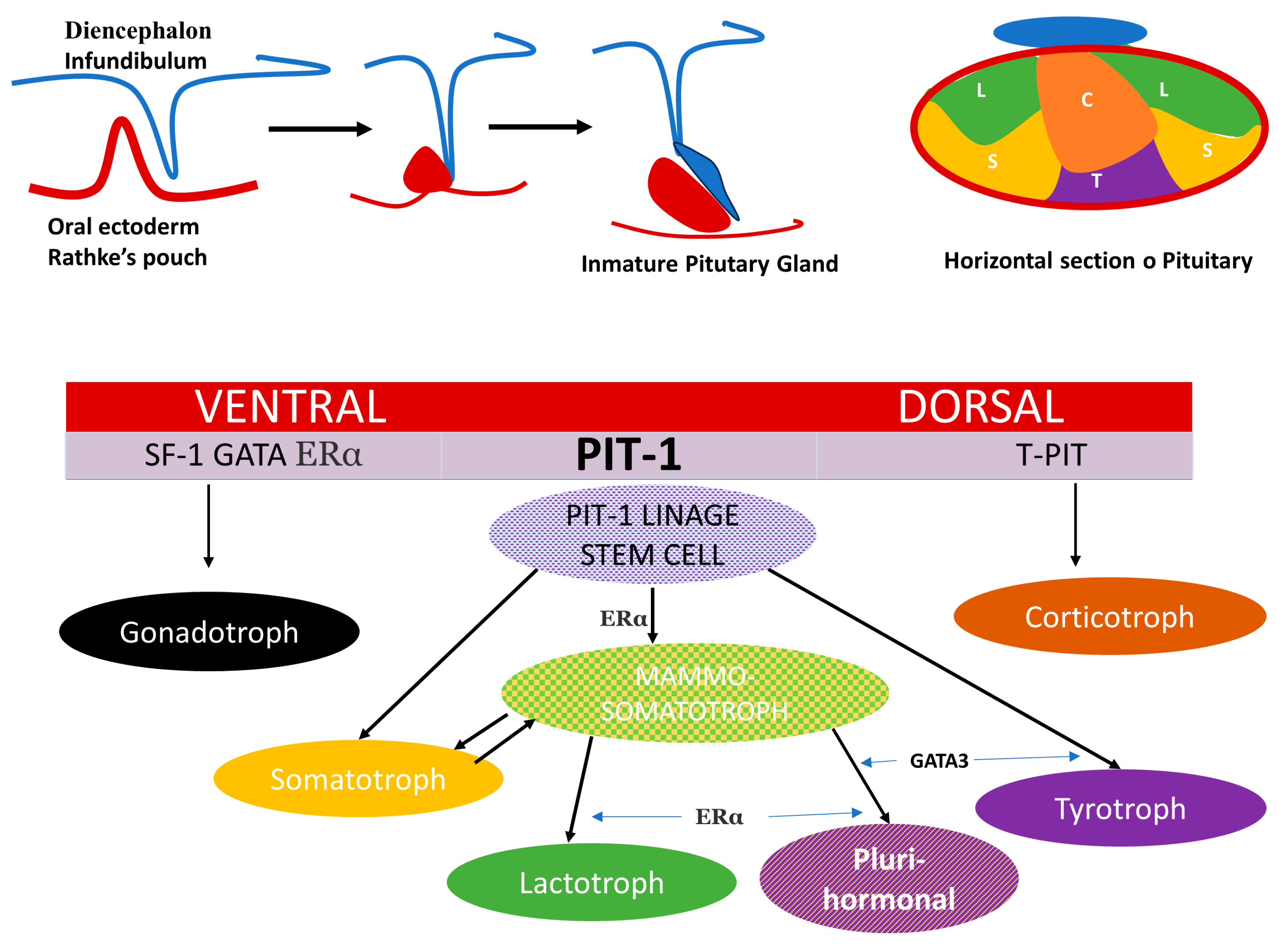
3. Ontogeny and Cell Linage
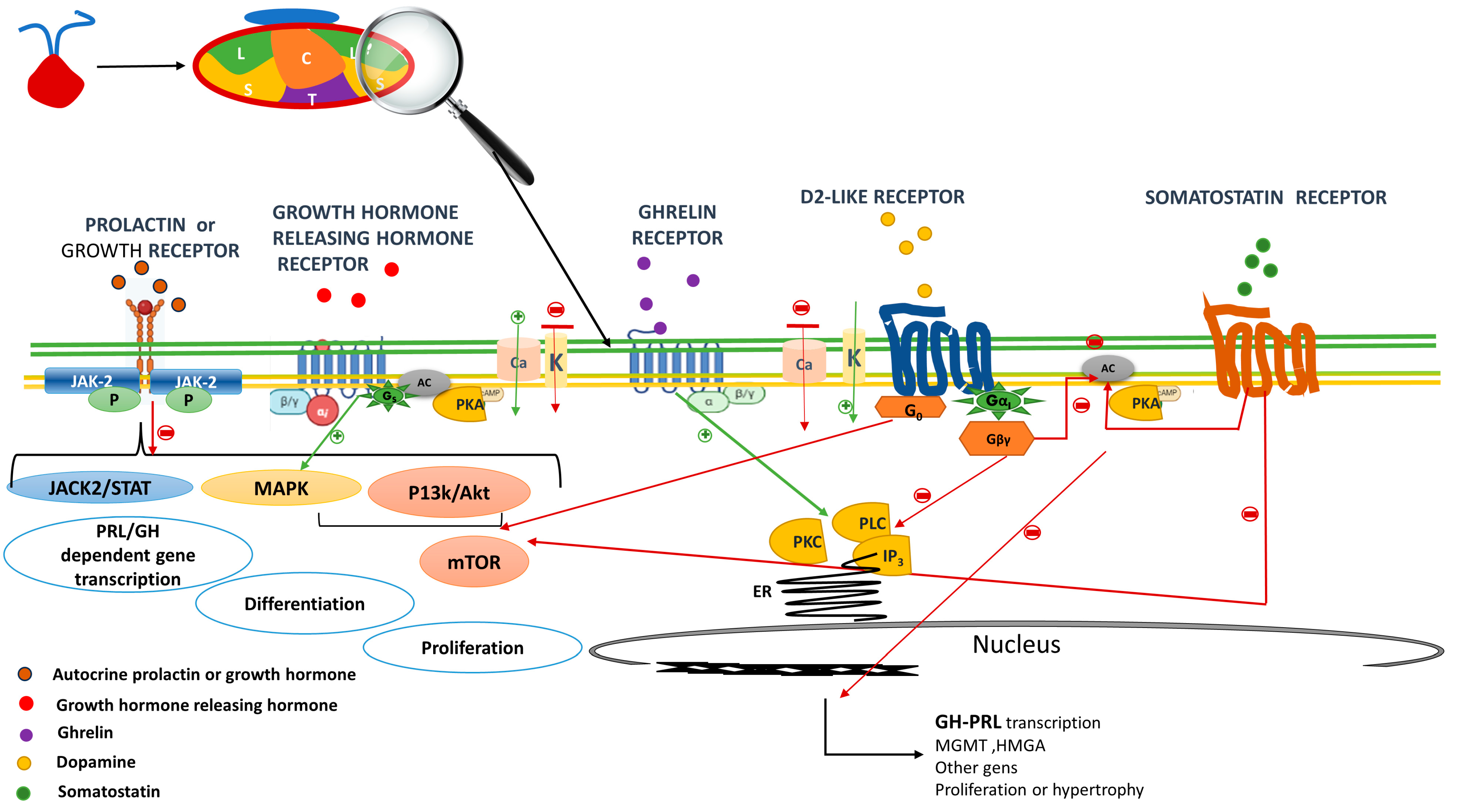
4. Prolactin and GH Receptor in the Mammosomatotroph Cell
5. Dopamine Receptor
6. GHRH Receptor and Ghrelin Receptor
7. Somatostatin Receptors
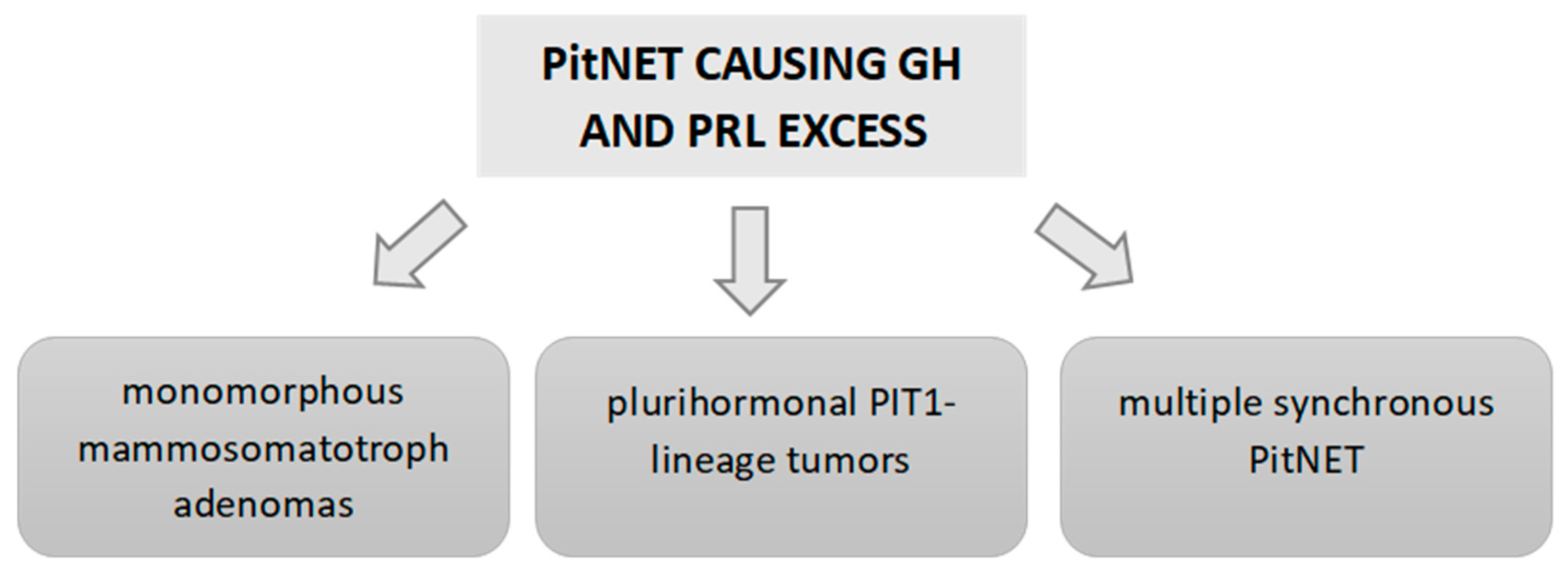
8. Classification of GH–PRL-Secreting PitNETs
9. Clinical Aspects and Outcomes of GH–PRL PitNETs
9.1. Prevalence and Epidemiological Aspects

{kind=link}
{kind=link}
{kind=link}
| Number of Cases | Definitions | Gender and Age | Tumors Size and Invasiveness | Hormonal Data | Surgical and Medical Outcomes | |
|---|---|---|---|---|---|---|
| PATHOLOGICAL CLASSIFICATION (based on PRL and GH staining) | ||||||
| Varlamov, 2020 [109] | Bihormonal group (9 MSA,10 MSLA), 30 DGA, and 28 SGA | Based on 2017 WHO classification staining for GH, PRL and cytokeratin | Patients in the bihormonal group were older than SGA. No differences in sex | Bihormonal adenomas did not differ in tumor size from DGA or SGA and were less frequently invasive than SGA | Bihormonal adenomas had higher baseline IGF1 index compared to SGA | No difference in the surgical remission rates across the groups |
| Liang Lv, 2019 [4] | 53 PSA, 28 MSA and 13 MSLA | Based on staining for GH and PRL * | No differences in sex and age across subtypes | MSLAs were larger and more invasive than MSA and PSA | No differences in baseline GH and IGF-1 level nor in hypopituitarism prevalence across subtypes | MSLAs had worse long-term biological remission rate than MSAs and PSAs |
| Rick, 2019 [95] | 69 GH-positive staining tumors and 22 with GH–PRL-positive staining | Based on staining for GH and PRL | No differences in sex and age between groups | No differences in tumor size nor in cavernous sinus invasion | GH–PRL tumors had higher serum PRL and IGF1 levels than GH tumors | Patients with GH–PRL tumors were less likely to achieve remission with surgery than GH tumors |
| HORMONAL CLASSIFICATION (based on serum PRL levels) | ||||||
| Wang, 2012 [104] | 182 patients with normal PRL levels and 97 with high levels of PRL | PRL threshold: any PRL elevation above the ULN | The GH group had a higher age than GH–PRL group. No differences by gender | The GH group had a smaller mean maximal diameter | Higher GH levels in the GH group than in the GH–PRL group | No differences in surgical control rates were observed between both groups |
| Laethem, 2020 [105] | 25 with normal PRL levels and 19 with high levels | PRL threshold: any PRL elevation above the ULN | No differences in age at diagnosis and gender between both groups | No differences in tumor size and invasiveness between groups | Similar GH and IGF1 levels between groups | The rate of surgical remission of acromegaly was not reported |
| Guo, 2022 [96] | 322 with normal PRL levels and 207 with high PRL levels | Hyperprolactinemia and hypopituitarism in acromegaly and effect of pituitary surgery: long-term follow-up on 529 patients | Patients with hyperprolactinemia were younger and more likely to be females | Patients with hyperprolactinemia had larger and more invasive tumors | Higher baseline and GH nadir levels in patients with hyperprolactinemia | Patients with preoperative hyperprolactinemia had higher recurrence rates after surgery than those without |
| IN VITRO PRESENCE OF DETECTABLE PRL SECRETION | ||||||
| Gatto, 2022 [110] | 15 with normal PRL and 8 with high PRL | Based on in vitro secretion of GH and/or PRL ** | No differences in age and gender between both groups | The prevalence of macroadenomas was comparable | No differences in GH and IGF1 levels between groups | No differences in the response to octreotide and cabergoline between groups |
9.2. Clinical and Hormonal Behavior
9.3. Radiological Features
9.4. Surgical and Medical Outcomes
10. Future Directions and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Freeman, M.E.; Kanyicska, B.; Lerant, A.; Nagy, G. Prolactin: Structure, Function, and Regulation of Secretion. Physiol. Rev. 2000, 80, 1523–1631. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Flanagan, J.U.; Langley, R.J.; Hay, M.P.; Perry, J.K. Targeting Growth Hormone Function: Strategies and Therapeutic Applications. Signal Transduct. Target. Ther. 2019, 4, 3. [Google Scholar] [CrossRef] [PubMed]
- Asa, S.L.; Mete, O.; Perry, A.; Osamura, R.Y. Overview of the 2022 WHO Classification of Pituitary Tumors. Endocr. Pathol. 2022, 33, 6–26. [Google Scholar] [CrossRef] [PubMed]
- Lv, L.; Jiang, Y.; Yin, S.; Hu, Y.; Chen, C.; Ma, W.; Jiang, S.; Zhou, P. Mammosomatotroph and mixed somatotroph-lactotroph adenoma in acromegaly: A retrospective study with long-term follow-up. Endocrine 2019, 66, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Melmed, S.; Casanueva, F.F.; Hoffman, A.R.; Kleinberg, D.L.; Montori, V.M.; Schlechte, J.A.; Wass, J.A.H. Diagnosis and Treatment of Hyperprolactinemia: An Endocrine Society Clinical Practice Guideline. J. Clin. Endocrinol. Metab. 2011, 96, 273–288. [Google Scholar] [CrossRef] [PubMed]
- Vroonen, L.; Daly, A.F.; Beckers, A. Epidemiology and Management Challenges in Prolactinomas. Neuroendocrinology 2019, 109, 20–27. [Google Scholar] [CrossRef]
- Di Sarno, A.; Landi, M.L.; Cappabianca, P.; Di Salle, F.; Rossi, F.W.; Pivonello, R.; Di Somma, C.; Faggiano, A.; Lombardi, G.; Colao, A. Resistance to Cabergoline as Compared with Bromocriptine in Hyperprolactinemia: Prevalence, Clinical Definition, and Therapeutic Strategy. J. Clin. Endocrinol. Metab. 2001, 86, 5256–5261. [Google Scholar] [CrossRef]
- Molitch, M.E. Pharmacologic Resistance in Prolactinoma Patients. Pituitary 2005, 8, 43–52. [Google Scholar] [CrossRef]
- Colao, A.; Di Sarno, A.; Landi, M.L.; Scavuzzo, F.; Cappabianca, P.; Pivonello, R.; Volpe, R.; Di Salle, F.; Cirillo, S.; Annunziato, L.; et al. Macroprolactinoma Shrinkage during Cabergoline Treatment Is Greater in Naive Patients Than in Patients Pretreated with Other Dopamine Agonists: A Prospective Study in 110 Patients. J. Clin. Endocrinol. Metab. 2000, 85, 2247–2252. [Google Scholar] [CrossRef]
- Biagetti, B.; Sarria-Estrada, S.; Ng-Wong, Y.K.; Martinez-Saez, E.; Casteràs, A.; Asanza, E.C.; Hernandez, I.; Giralt-Arnaiz, M.; Simò, R. Shrinkage by the third month predicts long-term response of macroprolactinoma after cabergoline. Eur. J. Endocrinol. 2021, 185, 587–595. [Google Scholar] [CrossRef]
- Lopes, M.B.S. The 2017 World Health Organization classification of tumors of the pituitary gland: A summary. Acta Neuropathol. 2017, 134, 521–535. [Google Scholar] [CrossRef] [PubMed]
- Besser, G.M.; Burman, P.; Daly, A.F. Predictors and rates of treatment-resistant tumor growth in acromegaly. Eur. J. Endocrinol. 2005, 153, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Gil, J.; Marques-Pamies, M.; Sampedro, M.; Webb, S.M.; Serra, G.; Salinas, I.; Blanco, A.; Valassi, E.; Carrato, C.; Picó, A.; et al. Data mining analyses for precision medicine in acromegaly: A proof of concept. Sci. Rep. 2022, 12, 8979. [Google Scholar] [CrossRef] [PubMed]
- Gadelha, M.R.; Kasuki, L. Refractory somatotroph adenomas. Pituitary 2023, 26, 266–268. [Google Scholar] [CrossRef] [PubMed]
- Baethge, C.; Goldbeck-Wood, S.; Mertens, S. SANRA—A scale for the quality assessment of narrative review articles. Res. Integr. Peer Rev. 2019, 4, 5. [Google Scholar] [CrossRef] [PubMed]
- Ganapathy, M.K.; Tadi, P. Anatomy, Head and Neck, Pituitary Gland. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- de Moraes, D.C.; Vaisman, M.; Conceição, F.L.; Ortiga-Carvalho, T.M. Pituitary development: A complex, temporal regulated process dependent on specific transcriptional factors. J. Endocrinol. 2012, 215, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, Y.G. Effects of brain and mesenchyme upon the cytogenesis of rat adenohypophysis in vitro. I. Differentiation of Adrenocorticotropes. Cell Tissue Res. 1982, 227, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Ingraham, H.A.; Treacy, M.N.; Albert, V.R.; Wilson, L.; Rosenfeld, M.G. Autoregulation of pit-1 gene expression mediated by two cis-active promoter elements. Nature 1990, 346, 583–586. [Google Scholar] [CrossRef]
- Akirov, A.; Asa, S.L.; Amer, L.; Shimon, I.; Ezzat, S. The Clinicopathological Spectrum of Acromegaly. J. Clin. Med. 2019, 8, E1962. [Google Scholar] [CrossRef]
- Syro, L.V.; Rotondo, F.; Serna, C.A.; Ortiz, L.D.; Kovacs, K. Pathology of GH-producing pituitary adenomas and GH cell hyperplasia of the pituitary. Pituitary 2017, 20, 84–92. [Google Scholar] [CrossRef]
- Niall, H.D.; Hogan, M.L.; Sauer, R.; Rosenblum, I.Y.; Greenwood, F.C. Sequences of Pituitary and Placental Lactogenic and Growth Hormones: Evolution from a Primordial Peptide by Gene Reduplication. Proc. Natl. Acad. Sci. USA 1971, 68, 866–869. [Google Scholar] [CrossRef]
- Wallis, M. The expanding growth hormone/prolactin family. J. Mol. Endocrinol. 1992, 9, 185–188. [Google Scholar] [CrossRef] [PubMed]
- Bernard, V.; Lamothe, S.; Beau, I.; Guillou, A.; Martin, A.; Le Tissier, P.; Grattan, D.; Young, J.; Binart, N. Autocrine actions of prolactin contribute to the regulation of lactotroph function in vivo. FASEB J. 2018, 32, 4791–4797. [Google Scholar] [CrossRef] [PubMed]
- Leung, K.-C.; Ranji, P.; Akbarzadeh, A.; Rahmati-Yamchi, M.; Glad, C.A.M.; Barbosa, E.J.L.; Nystrom, H.F.; Carlsson, L.M.S.; Nilsson, S.; Nilsson, A.G.; et al. Regulation of Cytokine Receptor Signaling by Nuclear Hormone Receptors: A New Paradigm for Receptor Interaction. DNA Cell Biol. 2004, 23, 463–474. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, L.A.; Liongue, C.; Lewis, R.S.; Stephenson, S.E.; Ward, A.C. Cytokine receptor signaling through the Jak–Stat–Socs pathway in disease. Mol. Immunol. 2007, 44, 2497–2506. [Google Scholar] [CrossRef] [PubMed]
- Gorvin, C.M. The prolactin receptor: Diverse and emerging roles in pathophysiology. J. Clin. Transl. Endocrinol. 2015, 2, 85–91. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Martín-Pérez, J.; García-Martínez, J.M.; Sánchez-Bailón, M.P.; Mayoral-Varo, V.; Calcabrini, A. Role of Src Family Kinases in Prolactin Signaling. Adv. Exp. Med. Biol. 2015, 846, 163–188. [Google Scholar] [CrossRef] [PubMed]
- Ferraris, J.; Zárate, S.; Jaita, G.; Boutillon, F.; Bernadet, M.; Auffret, J.; Seilicovich, A.; Binart, N.; Goffin, V.; Pisera, D. Prolactin Induces Apoptosis of Lactotropes in Female Rodents. PLoS ONE 2014, 9, e97383. [Google Scholar] [CrossRef]
- de Dios, N.; Orrillo, S.J.; Irizarri, M.; Theas, M.S.; Boutillon, F.; Candolfi, M.; Seilicovich, A.; Goffin, V.; Pisera, D.; Ferraris, J. JAK2/STAT5 Pathway Mediates Prolactin-Induced Apoptosis of Lactotropes. Neuroendocrinology 2019, 108, 84–97. [Google Scholar] [CrossRef]
- Coker-Gurkan, A.; Ayhan-Sahin, B.; Keceloglu, G.; Obakan-Yerlikaya, P.; Arisan, E.-D.; Palavan-Unsal, N. Atiprimod induce apoptosis in pituitary adenoma: Endoplasmic reticulum stress and autophagy pathways. J. Cell. Biochem. 2019, 120, 19749–19763. [Google Scholar] [CrossRef]
- Sabatini, D.M. mTOR and cancer: Insights into a complex relationship. Nat. Rev. Cancer 2006, 6, 729–734. [Google Scholar] [CrossRef] [PubMed]
- Dworakowska, D.; Wlodek, E.; Leontiou, C.A.; Igreja, S.; Cakir, M.; Teng, M.; Prodromou, N.; Góth, M.I.; Grozinsky-Glasberg, S.; Gueorguiev, M.; et al. Activation of RAF/MEK/ERK and PI3K/AKT/mTOR pathways in pituitary adenomas and their effects on downstream effectors. Endocr.Relat. Cancer 2009, 16, 1329–1338. [Google Scholar] [CrossRef] [PubMed]
- Monsalves, E.; Juraschka, K.; Tateno, T.; Agnihotri, S.; Asa, S.L.; Ezzat, S.; Zadeh, G. The PI3K/AKT/mTOR pathway in the pathophysiology and treatment of pituitary adenomas. Endocr.Relat. Cancer 2014, 21, R331–R344. [Google Scholar] [CrossRef] [PubMed]
- Gorvin, C.M.; Newey, P.J.; Rogers, A.; Stokes, V.; Neville, M.J.; Lines, K.E.; Ntali, G.; Lees, P.; Morrison, P.J.; Singhellakis, P.N.; et al. Association of prolactin receptor (PRLR) variants with prolactinomas. Hum. Mol. Genet. 2019, 28, 1023–1037. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Way, J.S.; Zhang, X.; Sergey, M.; Bergsneider, M.; Wang, M.B.; Yong, W.H.; Heaney, A.P. Effect of Everolimus in Treatment of Aggressive Prolactin-Secreting Pituitary Adenomas. J. Clin. Endocrinol. Metab. 2019, 104, 1929–1936. [Google Scholar] [CrossRef] [PubMed]
- Ji, C.; Xu, W.; Ding, H.; Chen, Z.; Shi, C.; Han, J.; Yu, L.; Qiao, N.; Zhang, Y.; Cao, X.; et al. The p300 Inhibitor A-485 Exerts Antitumor Activity in Growth Hormone Pituitary Adenoma. J. Clin. Endocrinol. Metab. 2022, 107, e2291–e2300. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Dong, X.; Yap, J.; Hu, J. The MAPK and AMPK signalings: Interplay and implication in targeted cancer therapy. J. Hematol. Oncol. 2020, 13, 113. [Google Scholar] [CrossRef]
- Booth, A.; Trudeau, T.; Gomez, C.; Lucia, M.S.; Gutierrez-Hartmann, A. Persistent ERK/MAPK Activation Promotes Lactotrope Differentiation and Diminishes Tumorigenic Phenotype. Mol. Endocrinol. 2014, 28, 1999–2011. [Google Scholar] [CrossRef]
- Kola, B.; Korbonits, M.; Diaz-Cano, S.; Kaltsas, G.; Morris, D.G.; Jordan, S.; Metherell, L.; Powell, M.; Czirják, S.; Arnaldi, G.; et al. Reduced expression of the growth hormone and type 1 insulin-like growth factor receptors in human somatotroph tumours and an analysis of possible mutations of the growth hormone receptor. Clin. Endocrinol. 2003, 59, 328–338. [Google Scholar] [CrossRef]
- Cuny, T.; Zeiller, C.; Bidlingmaier, M.; Défilles, C.; Roche, C.; Blanchard, M.-P.; Theodoropoulou, M.; Graillon, T.; Pertuit, M.; Figarella-Branger, D.; et al. In vitro impact of pegvisomant on growth hormone-secreting pituitary adenoma cells. Endocr.Relat. Cancer 2016, 23, 509–519. [Google Scholar] [CrossRef]
- Asa, S.L.; DiGiovanni, R.; Jiang, J.; Ward, M.L.; Loesch, K.; Yamada, S.; Sano, T.; Yoshimoto, K.; Frank, S.J.; Ezzat, S. A Growth Hormone Receptor Mutation Impairs Growth Hormone Autofeedback Signaling in Pituitary Tumors. Cancer Res. 2007, 67, 7505–7511. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.S.; Kokay, I.C.; Phillipps, H.R.; Yip, S.H.; Gustafson, P.; Wyatt, A.; Larsen, C.M.; Knowles, P.; Ladyman, S.R.; LeTissier, P.; et al. Conditional Deletion of the Prolactin Receptor Reveals Functional Subpopulations of Dopamine Neurons in the Arcuate Nucleus of the Hypothalamus. J. Neurosci. 2016, 36, 9173–9185. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, A.; Voogt, J.; Carr, L. Plasma Luteinizing Hormone and Prolactin Levels and Hypothalamic Catecholamine Synthesis in Steroid-Treated Ovariectomized Rats. Neuroendocrinology 1977, 23, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, A.; Lenchner, J.R.; Saadabadi, A. Biochemistry, Dopamine Receptors. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Ben-Jonathan, N.; Hnasko, R. Dopamine as a Prolactin (PRL) Inhibitor. Endocr. Rev. 2001, 22, 724–763. [Google Scholar] [CrossRef] [PubMed]
- Scully, K.M.; Rosenfeld, M.G. Pituitary Development: Regulatory Codes in Mammalian Organogenesis. Science 2002, 295, 2231–2235. [Google Scholar] [CrossRef] [PubMed]
- Ferone, D.; de Herder, W.W.; Pivonello, R.; Kros, J.M.; van Koetsveld, P.M.; de Jong, T.; Minuto, F.; Colao, A.; Lamberts, S.W.J.; Hofland, L.J. Correlation of in Vitro and in Vivo Somatotropic Adenoma Responsiveness to Somatostatin Analogs and Dopamine Agonists with Immunohistochemical Evaluation of Somatostatin and Dopamine Receptors and Electron Microscopy. J. Clin. Endocrinol. Metab. 2008, 93, 1412–1417. [Google Scholar] [CrossRef] [PubMed]
- Neto, L.V.; Machado, E.D.O.; Luque, R.M.; Taboada, G.F.; Marcondes, J.B.; Chimelli, L.M.C.; Quintella, L.P.; Niemeyer, P.; de Carvalho, D.P.; Kineman, R.D.; et al. Expression Analysis of Dopamine Receptor Subtypes in Normal Human Pituitaries, Nonfunctioning Pituitary Adenomas and Somatotropinomas, and the Association between Dopamine and Somatostatin Receptors with Clinical Response to Octreotide-LAR in Acromegaly. J. Clin. Endocrinol. Metab. 2009, 94, 1931–1937. [Google Scholar] [CrossRef] [PubMed]
- Katznelson, L.; Laws, E.R.; Melmed, S.; Molitch, M.E.; Murad, M.H.; Utz, A.; Wass, J.A.H. Acromegaly: An Endocrine Society Clinical Practice Guideline. J. Clin. Endocrinol. Metab. 2014, 99, 3933–3951. [Google Scholar] [CrossRef]
- Giustina, A.; Barkhoudarian, G.; Beckers, A.; Ben-Shlomo, A.; Biermasz, N.; Biller, B.; Boguszewski, C.; Bolanowski, M.; Bollerslev, J.; Bonert, V.; et al. Multidisciplinary management of acromegaly: A consensus. Rev. Endocr. Metab. Disord. 2020, 21, 667–678. [Google Scholar] [CrossRef]
- Friedman, E.; Adams, E.F.; Höög, A.; Gejman, P.V.; Carson, E.; Larsson, C.; De Marco, L.; Werner, S.; Fahlbusch, R.; Nordenskjöld, M. Normal structural dopamine type 2 receptor gene in prolactin-secreting and other pituitary tumors. J. Clin. Endocrinol. Metab. 1994, 78, 568–574. [Google Scholar] [CrossRef]
- Bueno, C.; Trarbach, E.; Bronstein, M.D.; Glezer, A. Cabergoline and prolactinomas: Lack of association between DRD2 polymorphisms and response to treatment. Pituitary 2017, 20, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Peillon, F.; Cesselin, F.; Bression, D.; Zygelman, N.; Brandi, A.; Nousbaum, A.; Mauborgne, A. In Vitro Effect of Dopamine and L-Dopa on Prolactin and Growth Hormone Release from Human Pituitary Adenomas. J. Clin. Endocrinol. Metab. 1979, 49, 737–741. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.; Jiang, X.; Wang, C.; Liu, J.; Chen, Y.; Li, Q.; Wu, J.; Zheng, W.; Zhuge, Q.; Jin, K.; et al. Differential effects of nerve growth factor on expression of dopamine 2 receptor subtypes in GH3 rat pituitary tumor cells. Endocrine 2012, 42, 670–675. [Google Scholar] [CrossRef] [PubMed]
- Olarescu, N.C.; Gunawardane, K.; Hansen, T.K.; Møller, N.; Jørgensen, J.O.L. Normal Physiology of Growth Hormone in Adults. In Endotext; Feingold, K.R., Anawalt, B., Boyce, A., Chrousos, G., de Herder, W.W., Dungan, K., Grossman, A., Hershman, J.M., Hofland, H.J., Kaltsas, G., et al., Eds.; MDText.com, Inc.: South Dartmouth, MA, USA, 2000. [Google Scholar]
- Kojima, M.; Hosoda, H.; Date, Y.; Nakazato, M.; Matsuo, H.; Kangawa, K. Ghrelin Is a Growth-Hormone-Releasing Acylated Peptide from Stomach. Nature 1999, 402, 656–660. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Zhang, H.; Cong, Z.; Zhao, L.-H.; Zhou, Q.; Mao, C.; Cheng, X.; Shen, D.-D.; Cai, X.; Ma, C.; et al. Structural basis for activation of the growth hormone-releasing hormone receptor. Nat. Commun. 2020, 11, 5205. [Google Scholar] [CrossRef]
- Lopes, M.B.; Gaylinn, B.D.; Thorner, M.O.; Stoler, M.H. Growth hormone-releasing hormone receptor mRNA in acromegalic pituitary tumors. Am. J. Pathol. 1997, 150, 1885–1891. [Google Scholar] [PubMed]
- Zeitler, P.; Siriwardana, G. Stimulation of Mitogen-Activated Protein Kinase Pathway in Rat Somatotrophs by Growth Hormone-Releasing Hormone. Endocrine 2000, 12, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, I.; Oka, H.; Naritaka, H.; Sato, Y.; Fujii, K.; Kameya, T. Expression of Pit-1 and growth hormone-releasing hormone receptor mRNA in human pituitary adenomas: Difference among functioning, silent, and other nonfunctioning adenomas. Endocr. Pathol. 2002, 13, 83–98. [Google Scholar] [CrossRef]
- Roche, C.; RasolonJanahary, R.; Thirion, S.; Goddard, I.; Fusco, A.; Figarella-Branger, D.; Dufour, H.; Brue, T.; Franc, J.-L.; Enjalbert, A.; et al. Inactivation of Transcription Factor Pit-1 to Target Tumoral Somatolactotroph Cells. Hum. Gene Ther. 2012, 23, 104–114. [Google Scholar] [CrossRef]
- Spada, A.; Arosio, M.; Bochicchio, D.; Bazzoni, N.; Vallar, L.; Bassetti, M.; Faglia, G. Clinical, Biochemical, and Morphological Correlates in Patients Bearing Growth Hormone-Secreting Pituitary Tumors with or without Constitutively Active Adenylyl Cyclase. J. Clin. Endocrinol. Metab. 1990, 71, 1421–1426. [Google Scholar] [CrossRef]
- Landis, C.A.; Harsh, G.; Lyons, J.; Davis, R.L.; McCormick, F.; Bourne, H.R. Clinical Characteristics of Acromegalic Patients whose Pituitary Tumors Contain Mutant Gs Protein. J. Clin. Endocrinol. Metab. 1990, 71, 1416–1420. [Google Scholar] [CrossRef] [PubMed]
- Landis, C.A.; Masters, S.B.; Spada, A.; Pace, A.M.; Bourne, H.R.; Vallar, L. GTPase inhibiting mutations activate the α chain of Gs and stimulate adenylyl cyclase in human pituitary tumours. Nature 1989, 340, 692–696. [Google Scholar] [CrossRef] [PubMed]
- Lania, A.G.; Ferrero, S.; Pivonello, R.; Mantovani, G.; Peverelli, E.; Di Sarno, A.; Beck-Peccoz, P.; Spada, A.; Colao, A. Evolution of an Aggressive Prolactinoma into a Growth Hormone Secreting Pituitary Tumor Coincident with GNAS Gene Mutation. J. Clin. Endocrinol. Metab. 2010, 95, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Wang, P.; Zheng, H.; Smith, R.G. Ghrelin stimulation of growth hormone release and appetite is mediated through the growth hormone secretagogue receptor. Proc. Natl. Acad. Sci. USA 2004, 101, 4679–4684. [Google Scholar] [CrossRef] [PubMed]
- Reichenbach, A.; Steyn, F.J.; Sleeman, M.W.; Andrews, Z.B. Ghrelin Receptor Expression and Colocalization with Anterior Pituitary Hormones Using a GHSR-GFP Mouse Line. Endocrinology 2012, 153, 5452–5466. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Ye, F.; Wang, B.; Tian, C.; Wang, S.; Shu, K.; Guo, D.; Lei, T. Elevation of growth hormone secretagogue receptor type 1a mRNA expression in human growth hormone-secreting pituitary adenoma harboring G protein alpha subunit mutation. Neuroendocrinol. Lett. 2010, 31, 147–154. [Google Scholar] [PubMed]
- Larkin, S.; Reddy, R.; Karavitaki, N.; Cudlip, S.; Wass, J.; Ansorge, O. Granulation pattern, but not GSP or GHR mutation, is associated with clinical characteristics in somatostatin-naïve patients with somatotroph adenomas. Eur. J. Endocrinol. 2013, 168, 491–499. [Google Scholar] [CrossRef]
- Taboada, G.F.; Luque, R.M.; Bastos, W.; Guimarães, R.F.C.; Marcondes, J.B.; Chimelli, L.M.C.; Fontes, R.; Mata, P.J.P.; Filho, P.N.; Carvalho, D.P.; et al. Quantitative analysis of somatostatin receptor subtype (SSTR1–5) gene expression levels in somatotropinomas and non-functioning pituitary adenomas. Eur. J. Endocrinol. 2007, 156, 65–74. [Google Scholar] [CrossRef]
- Taboada, G.F.; Luque, R.M.; Neto, L.V.; Machado, E.D.O.; Sbaffi, B.C.; Domingues, R.C.; Marcondes, J.B.; Chimelli, L.M.C.; Fontes, R.; Niemeyer, P.; et al. Quantitative analysis of somatostatin receptor subtypes (1–5) gene expression levels in somatotropinomas and correlation to in vivo hormonal and tumor volume responses to treatment with octreotide LAR. Eur. J. Endocrinol. 2008, 158, 295–303. [Google Scholar] [CrossRef]
- Casar-Borota, O.; Heck, A.; Schulz, S.; Nesland, J.M.; Ramm-Pettersen, J.; Lekva, T.; Alafuzoff, I.; Bollerslev, J. Expression of SSTR2a, but not of SSTRs 1, 3, or 5 in Somatotroph Adenomas Assessed by Monoclonal Antibodies Was Reduced by Octreotide and Correlated with the Acute and Long-Term Effects of Octreotide. J. Clin. Endocrinol. Metab. 2013, 98, E1730–E1739. [Google Scholar] [CrossRef]
- Iacovazzo, D.; Carlsen, E.; Lugli, F.; Chiloiro, S.; Piacentini, S.; Bianchi, A.; Giampietro, A.; Mormando, M.; Clear, A.J.; Doglietto, F.; et al. Factors predicting pasireotide responsiveness in somatotroph pituitary adenomas resistant to first-generation somatostatin analogues: An immunohistochemical study. Eur. J. Endocrinol. 2016, 174, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Venegas-Moreno, E.; Vazquez-Borrego, M.C.; Dios, E.; Gros-Herguido, N.; Flores-Martinez, A.; Rivero-Cortés, E.; Madrazo-Atutxa, A.; Japón, M.A.; Luque, R.M.; Castaño, J.P.; et al. Association between dopamine and somatostatin receptor expression and pharmacological response to somatostatin analogues in acromegaly. J. Cell. Mol. Med. 2018, 22, 1640–1649. [Google Scholar] [CrossRef] [PubMed]
- Puig-Domingo, M.; Gil, J.; Sampedro-Nuñez, M.; Jordà, M.; Webb, S.M.; Serra, G.; Pons, L.; Salinas, I.; Blanco, A.; Marques-Pamies, M.; et al. Molecular profiling for acromegaly treatment: A validation study. Endocr.-Relat. Cancer 2020, 27, 375–389. [Google Scholar] [CrossRef] [PubMed]
- Gadelha, M.R.; Bronstein, M.D.; Brue, T.; Coculescu, M.; Fleseriu, M.; Guitelman, M.; Pronin, V.; Raverot, G.; Shimon, I.; Lievre, K.K.; et al. Pasireotide versus continued treatment with octreotide or lanreotide in patients with inadequately controlled acromegaly (PAOLA): A randomised, phase 3 trial. Lancet Diabetes Endocrinol. 2014, 2, 875–884. [Google Scholar] [CrossRef] [PubMed]
- Colao, A.; Bronstein, M.D.; Freda, P.; Gu, F.; Shen, C.-C.; Gadelha, M.; Fleseriu, M.; van der Lely, A.J.; Farrall, A.J.; Ruffin, M.; et al. Pasireotide Versus Octreotide in Acromegaly: A Head-to-Head Superiority Study. J. Clin. Endocrinol. Metab. 2014, 99, 791–799. [Google Scholar] [CrossRef] [PubMed]
- Gatto, F.; Feelders, R.A.; Franck, S.E.; Van Koetsveld, P.M.; Dogan, F.; Kros, J.M.; Neggers, S.J.C.M.M.; Van Der Lely, A.-J.; Lamberts, S.W.J.; Ferone, D.; et al. In Vitro Head-to-Head Comparison Between Octreotide and Pasireotide in GH-Secreting Pituitary Adenomas. J. Clin. Endocrinol. Metab. 2017, 102, 2009–2018. [Google Scholar] [CrossRef] [PubMed]
- Chiloiro, S.; Giampietro, A.; Mirra, F.; Donfrancesco, F.; Tartaglione, T.; Mattogno, P.P.; Angelini, F.; Liverana, L.; Gessi, M.; Carmelo, A.; et al. Pegvisomant and Pasireotide LAR as second line therapy in acromegaly: Clinical effectiveness and predictors of response. Eur. J. Endocrinol. 2021, 184, 217–229. [Google Scholar] [CrossRef]
- Hofland, L.J.; van der Hoek, J.; van Koetsveld, P.M.; de Herder, W.W.; Waaijers, M.; Sprij-Mooij, D.; Bruns, C.; Weckbecker, G.; Feelders, R.; van der Lely, A.-J.; et al. The Novel Somatostatin Analog SOM230 Is a Potent Inhibitor of Hormone Release by Growth Hormone- and Prolactin-Secreting Pituitary Adenomas In Vitro. J. Clin. Endocrinol. Metab. 2004, 89, 1577–1585. [Google Scholar] [CrossRef]
- Waser, B.; Cescato, R.; Tamma, M.-L.; Maecke, H.R.; Reubi, J.C. Absence of somatostatin SST2 receptor internalization in vivo after intravenous SOM230 application in the AR42J animal tumor model. Eur. J. Pharmacol. 2010, 644, 257–262. [Google Scholar] [CrossRef]
- Muhammad, A.; Coopmans, E.C.; Gatto, F.; Franck, S.E.; Janssen, J.A.M.J.L.; van der Lely, A.J.; Hofland, L.J.; Neggers, S.J.C.M.M. Pasireotide Responsiveness in Acromegaly Is Mainly Driven by Somatostatin Receptor Subtype 2 Expression. J. Clin. Endocrinol. Metab. 2019, 104, 915–924. [Google Scholar] [CrossRef]
- Shimon, I.; Yan, X.; Taylor, J.E.; Weiss, M.H.; Culler, M.D.; Melmed, S. Somatostatin receptor (SSTR) subtype-selective analogues differentially suppress in vitro growth hormone and prolactin in human pituitary adenomas. Novel potential therapy for functional pituitary tumors. J. Clin. Investig. 1997, 100, 2386–2392. [Google Scholar] [CrossRef] [PubMed]
- Coopmans, E.C.; van Meyel, S.W.F.; Pieterman, K.J.; van Ipenburg, J.A.; Hofland, L.J.; Donga, E.; Daly, A.F.; Beckers, A.; van der Lely, A.-J.; Neggers, S.J.C.M.M. Excellent response to pasireotide therapy in an aggressive and dopamine-resistant prolactinoma. Eur. J. Endocrinol. 2019, 181, K21–K27. [Google Scholar] [CrossRef] [PubMed]
- Lasolle, H.; Vasiljevic, A.; Borson-Chazot, F.; Raverot, G. Pasireotide: A potential therapeutic alternative for resistant prolactinoma. Ann. d’Endocrinol. 2019, 80, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Raverot, G.; Vasiljevic, A.; Jouanneau, E.; Lasolle, H. Confirmation of a new therapeutic option for aggressive or dopamine agonist-resistant prolactin pituitary neuroendocrine tumors. Eur. J. Endocrinol. 2019, 181, C1–C3. [Google Scholar] [CrossRef] [PubMed]
- Zatelli, M.C.; Piccin, D.; Tagliati, F.; Ambrosio, M.R.; Margutti, A.; Padovani, R.; Scanarini, M.; Culler, M.D.; degli Uberti, E.C. Somatostatin Receptor Subtype 1 Selective Activation in Human Growth Hormone (GH)- and Prolactin (PRL)-Secreting Pituitary Adenomas: Effects on Cell Viability, GH, and PRL Secretion. J. Clin. Endocrinol. Metab. 2003, 88, 2797–2802. [Google Scholar] [CrossRef] [PubMed]
- Rocheville, M.; Lange, D.C.; Kumar, U.; Sasi, R.; Patel, R.C.; Patel, Y.C. Subtypes of the Somatostatin Receptor Assemble as Functional Homo- and Heterodimers. J. Biol. Chem. 2000, 275, 7862–7869. [Google Scholar] [CrossRef] [PubMed]
- Ashton, C.; Rhie, S.K.; Carmichael, J.D.; Zada, G. Role of KCNAB2 expression in modulating hormone secretion in somatotroph pituitary adenoma. J. Neurosurg. 2020, 1, 787–793. [Google Scholar] [CrossRef]
- Lopes, M.B.S. Growth hormone-secreting adenomas: Pathology and cell biology. Neurosurg. Focus 2010, 29, E2. [Google Scholar] [CrossRef]
- Mete, O.; Alshaikh, O.M.; Cintosun, A.; Ezzat, S.; Asa, S.L. Synchronous Multiple Pituitary Neuroendocrine Tumors of Different Cell Lineages. Endocr. Pathol. 2018, 29, 332–338. [Google Scholar] [CrossRef]
- Asa, S.L.; Mete, O.; Riddle, N.D.; Perry, A. Multilineage Pituitary Neuroendocrine Tumors (PitNETs) Expressing PIT1 and SF1. Endocr. Pathol. 2023, 1–6. [Google Scholar] [CrossRef]
- von Schöning, J.; Flitsch, J.; Lüdecke, D.K.; Fahlbusch, R.; Buchfelder, M.; Buslei, R.; Knappe, U.J.; Bergmann, M.; Schulz-Schaeffer, W.J.; Herms, J.; et al. Multiple tumorous lesions of the pituitary gland. Hormones 2022, 21, 653–663. [Google Scholar] [CrossRef] [PubMed]
- Rick, J.; Jahangiri, A.; Flanigan, P.M.; Chandra, A.; Kunwar, S.; Blevins, L.; Aghi, M.K. Growth hormone and prolactin-staining tumors causing acromegaly: A retrospective review of clinical presentations and surgical outcomes. J. Neurosurg. 2019, 131, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Zhang, R.; Zhang, D.; Wang, Z.; Gao, L.; Yao, Y.; Deng, K.; Bao, X.; Feng, M.; Xu, Z.; et al. Hyperprolactinemia and Hypopituitarism in Acromegaly and Effect of Pituitary Surgery: Long-Term Follow-up on 529 Patients. Front. Endocrinol. 2022, 12, 807054. [Google Scholar] [CrossRef] [PubMed]
- Halmi, N.S. Occurrence of both growth hormone- and prolactin-immunoreactive material in the cells of human somatotropic pituitary adenomas containing Mammotropic elements. Virchows Arch. 1982, 398, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Robert, F.; Pelletier, G.; Serri, O.; Hardy, J. Mixed growth hormone and prolactin-secreting human pituitary adenomas: A pathologic, immunocytochemical, ultrastructural, and immunoelectron microscopic study. Hum. Pathol. 1988, 19, 1327–1334. [Google Scholar] [CrossRef] [PubMed]
- Andersen, M.; Hagen, C.; Frystyk, J.; Schroeder, H. Development of acromegaly in patients with prolactinomas. Eur. J. Endocrinol. 2003, 149, 17–22. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Akirov, A.; Greenman, Y.; Glaser, B.; S’chigol, I.; Mansiterski, Y.; Eizenberg, Y.; Shraga-Slutzky, I.; Shimon, I. IGF-1 levels may increase paradoxically with dopamine agonist treatment for prolactinomas. Pituitary 2018, 21, 406–413. [Google Scholar] [CrossRef]
- Iglesias, P.; Villabona, C.; Díez, J.J. Effects of cabergoline therapy on serum IGF-1 concentrations in prolactinoma patients. Pituitary 2020, 23, 200–202. [Google Scholar] [CrossRef]
- Rosário, P.W.; Purisch, S. Biochemical acromegaly in patients with prolactinoma during treatment with dopaminergic agonists. Arq. Bras. Endocrinol. Metabol. 2010, 54, 546–549. [Google Scholar] [CrossRef]
- Arihara, Z.; Sakurai, K.; Yamashita, R.; Niitsuma, S.; Ueno, T.; Yamamura, N.; Yamada, S.; Inoshita, N.; Takahashi, K. Bromocriptine, a Dopamine Agonist, Increases Growth Hormone Secretion in a Patient with Acromegaly. Tohoku J. Exp. Med. 2014, 234, 129–135. [Google Scholar] [CrossRef][Green Version]
- Wang, M.; Mou, C.; Jiang, M.; Han, L.; Fan, S.; Huan, C.; Qu, X.; Han, T.; Qu, Y.; Xu, G. The characteristics of acromegalic patients with hyperprolactinemia and the differences in patients with merely GH-secreting adenomas: Clinical analysis of 279 cases. Eur. J. Endocrinol. 2012, 166, 797–802. [Google Scholar] [CrossRef] [PubMed]
- Van Laethem, D.; Michotte, A.; Cools, W.; Velkeniers, B.; Unuane, D.; Andreescu, C.E.; Bravenboer, B. Hyperprolactinemia in Acromegaly is Related to Prolactin Secretion by Somatolactotroph Tumours. Horm. Metab. Res. 2020, 52, 647–653. [Google Scholar] [CrossRef]
- Kerbel, J.; Cano-Zaragoza, A.; Espinosa-Dorado, R.; de la Torre, K.-E.G.; Mercado, M. Real World Data on the Epidemiology, Diagnosis, and Treatment of Acromegaly: A Registries-based Approach. Arch. Med. Res. 2023, 54, 102856. [Google Scholar] [CrossRef] [PubMed]
- Mete, O.; Lopes, M.B. Overview of the 2017 WHO Classification of Pituitary Tumors. Endocr. Pathol. 2017, 28, 228–243. [Google Scholar] [CrossRef] [PubMed]
- Asa, S.L.; Ezzat, S. The Pathogenesis of Pituitary Tumors. Annu. Rev. Pathol. Mech. Dis. 2009, 4, 97–126. [Google Scholar] [CrossRef] [PubMed]
- Varlamov, E.V.; Wood, M.D.; Netto, J.P.; Thiessen, J.; Kim, J.; Lim, D.S.T.; Yedinak, C.G.; Banskota, S.; Cetas, J.S.; Fleseriu, M. Cystic appearance on magnetic resonance imaging in bihormonal growth hormone and prolactin tumors in acromegaly. Pituitary 2020, 23, 672–680. [Google Scholar] [CrossRef] [PubMed]
- Gatto, F.; Feelders, R.A.; van Koetsveld, P.M.; Dogan, F.; Neggers, S.J.C.C.M.M.; van der Lelij, A.-J.; Amarù, J.; Ferone, D.; Hofland, L.J. Dissecting the In Vitro Efficacy of Octreotide and Cabergoline in GH- and GH/PRL-Secreting Pituitary Tumors. J. Clin. Endocrinol. Metab. 2022, 108, e98–e109. [Google Scholar] [CrossRef]
- Hagiwara, A.; Inoue, Y.; Wakasa, K.; Haba, T.; Tashiro, T.; Miyamoto, T. Comparison of Growth Hormone–producing and Non–Growth Hormone–producing Pituitary Adenomas: Imaging Characteristics and Pathologic Correlation. Radiology 2003, 228, 533–538. [Google Scholar] [CrossRef]
- Sandret, L.; Maison, P.; Chanson, P. Place of Cabergoline in Acromegaly: A Meta-Analysis. J. Clin. Endocrinol. Metab. 2011, 96, 1327–1335. [Google Scholar] [CrossRef]
- Peacey, S.R.; Shalet, S.M. Insulin-like growth factor 1 measurement in diagnosis and management of acromegaly. Ann. Clin. Biochem. Int. J. Biochem. Lab. Med. 2001, 38, 297–303. [Google Scholar] [CrossRef]



| Search Terms | Total of Results |
|---|---|
| pituitary [TI] AND ontogeny [TI] | 119 |
| mammosomatotroph [TIAB] | 90 |
| molecular [TIAB] AND acromegaly [TI] | 87 |
| molecular [TIAB] AND prolactinoma [TIAB] | 72 |
| prolactin receptor [TI] AND molecular [TIAB] | 102 |
| growth hormone receptor [TI] AND molecular [TIAB] | 161 |
| somatostatin receptor [TI] AND molecular [TIAB] AND pituitary [TIAB] | 47 |
| dopamine receptor [TI] AND molecular [TIAB] AND pituitary [TIAB] | 18 |
| GHRH receptor [TI] | 71 |
| Ghrelin receptor [TI] AND pituitary [TI] | 36 |
| plurihormonal [TIAB] AND pituitary [TI] | 189 |
| GH [TI] AND prolactin pituitary [TI] | 3 |
| mixed pituitary adenomas [TI] | 4 |
| acromegaly [TI] AND plurihormonal [TI] | 5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Araujo-Castro, M.; Marazuela, M.; Puig-Domingo, M.; Biagetti, B. Prolactin and Growth Hormone Signaling and Interlink Focused on the Mammosomatotroph Paradigm: A Comprehensive Review of the Literature. Int. J. Mol. Sci. 2023, 24, 14002. https://doi.org/10.3390/ijms241814002
Araujo-Castro M, Marazuela M, Puig-Domingo M, Biagetti B. Prolactin and Growth Hormone Signaling and Interlink Focused on the Mammosomatotroph Paradigm: A Comprehensive Review of the Literature. International Journal of Molecular Sciences. 2023; 24(18):14002. https://doi.org/10.3390/ijms241814002
Chicago/Turabian StyleAraujo-Castro, Marta, Mónica Marazuela, Manel Puig-Domingo, and Betina Biagetti. 2023. "Prolactin and Growth Hormone Signaling and Interlink Focused on the Mammosomatotroph Paradigm: A Comprehensive Review of the Literature" International Journal of Molecular Sciences 24, no. 18: 14002. https://doi.org/10.3390/ijms241814002
APA StyleAraujo-Castro, M., Marazuela, M., Puig-Domingo, M., & Biagetti, B. (2023). Prolactin and Growth Hormone Signaling and Interlink Focused on the Mammosomatotroph Paradigm: A Comprehensive Review of the Literature. International Journal of Molecular Sciences, 24(18), 14002. https://doi.org/10.3390/ijms241814002