
Biosynthesis of Vanillin by Rational Design of Enoyl-CoA Hydratase/Lyase
 , ,
, ,
Abstract
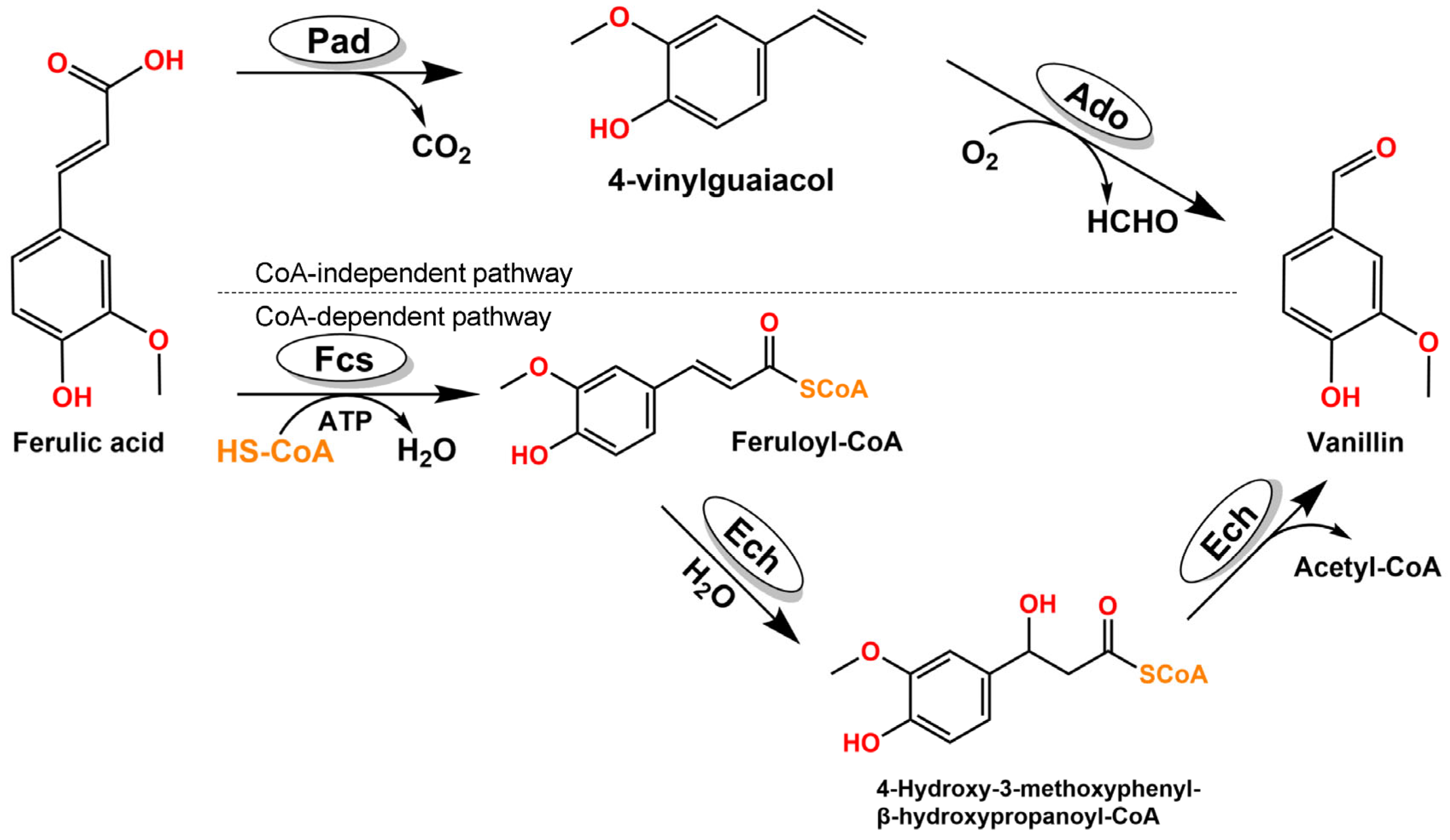
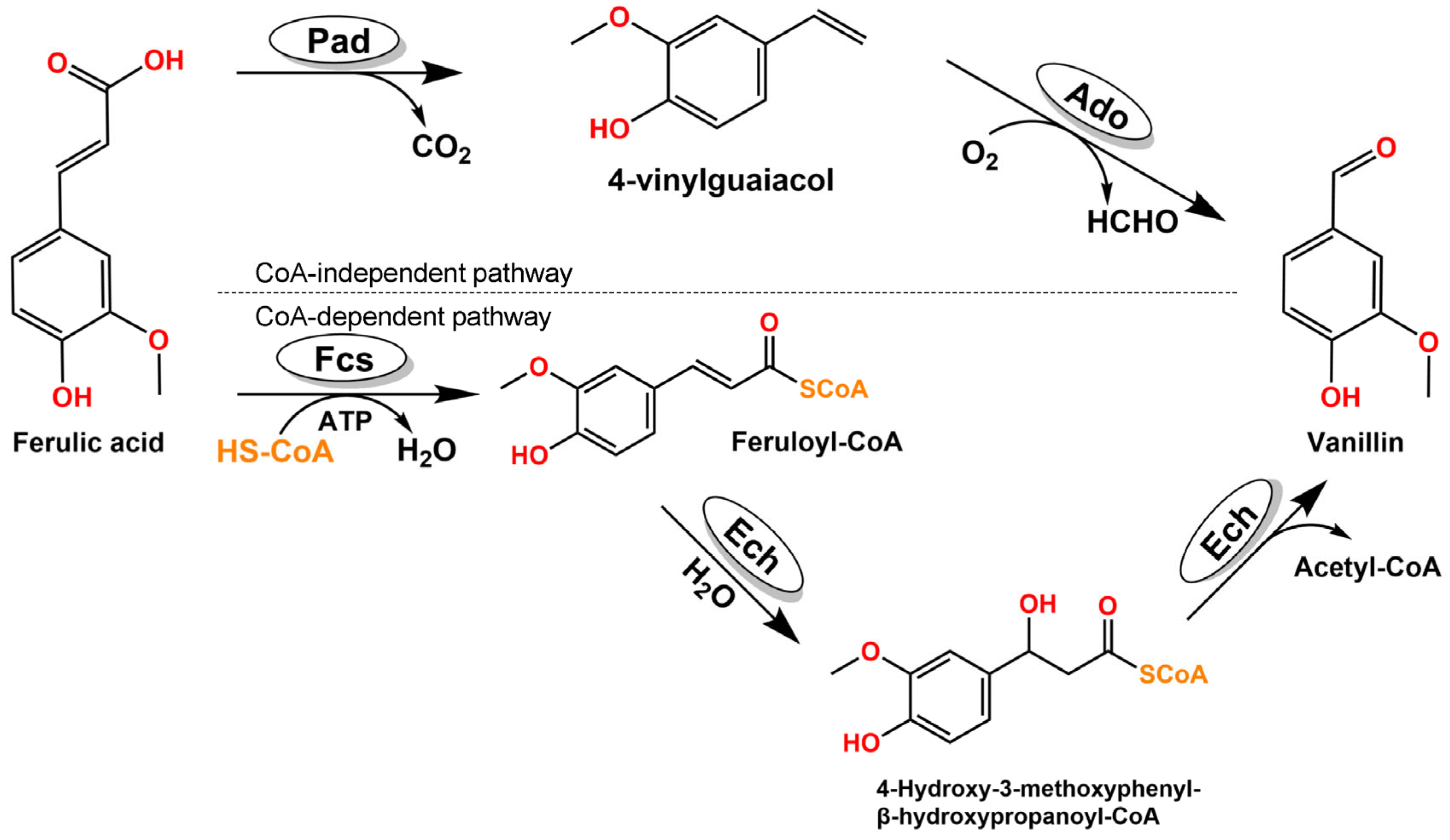
:1. Introduction
2. Results
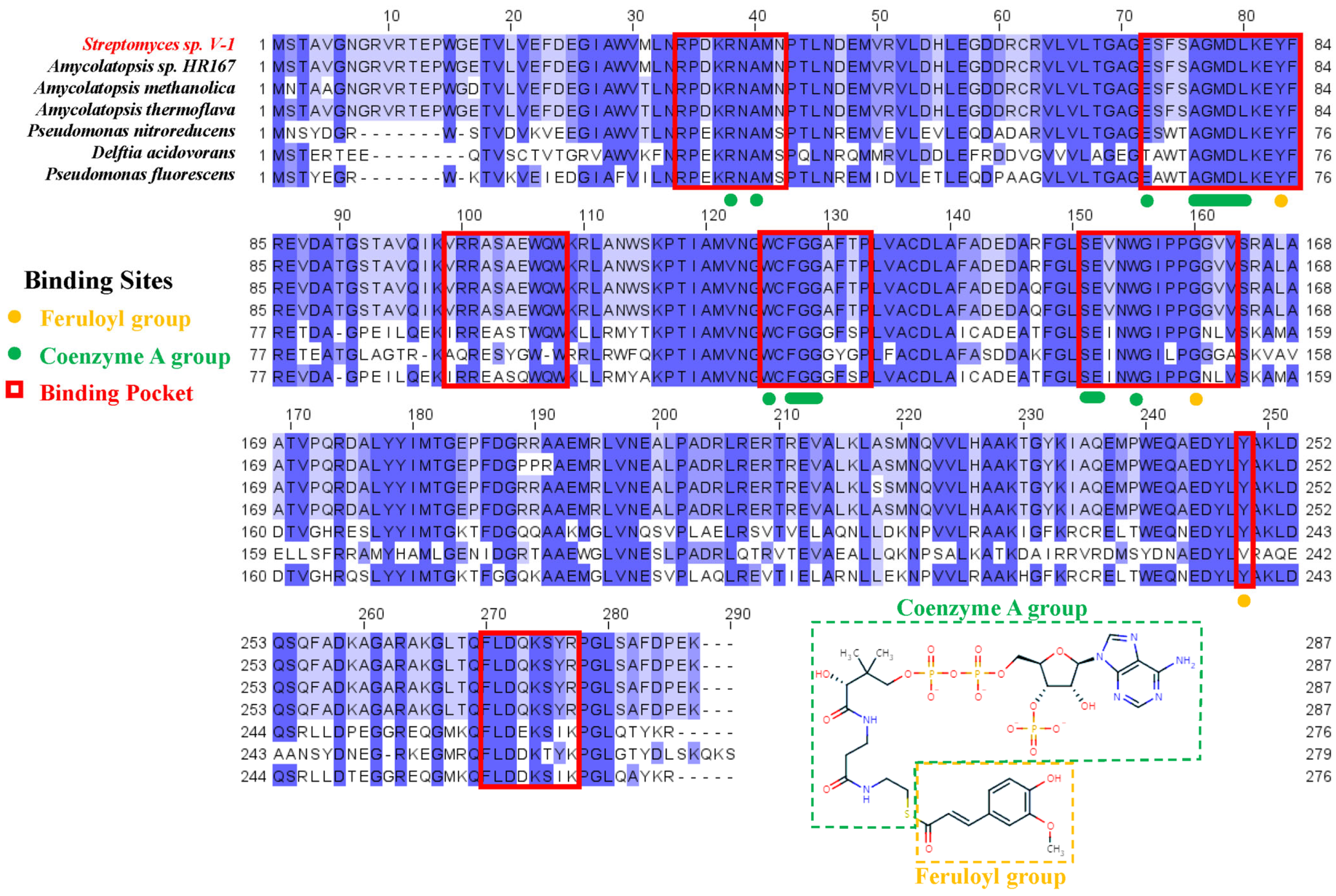
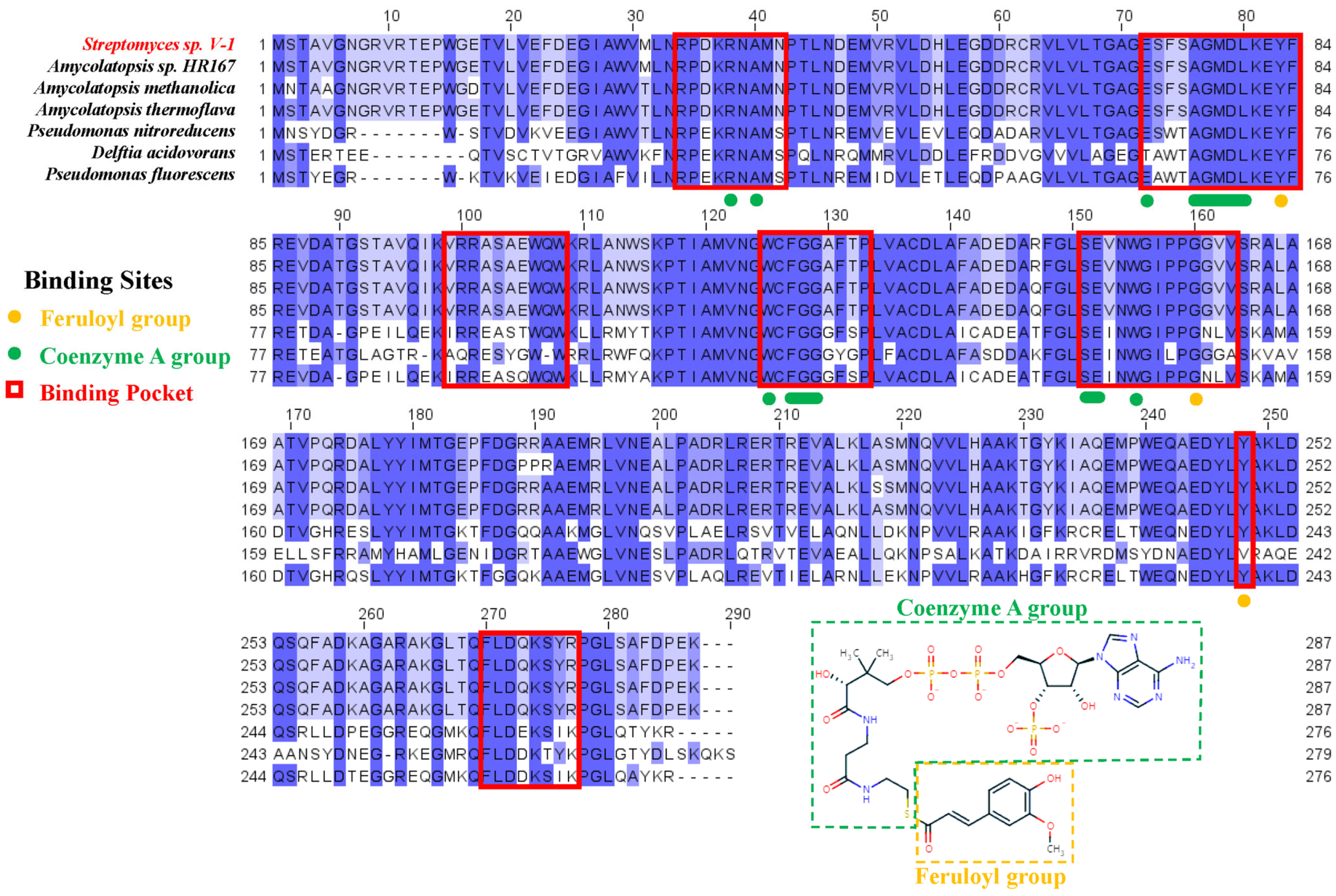
2.1. Sequence and Conservation Analysis of Ech
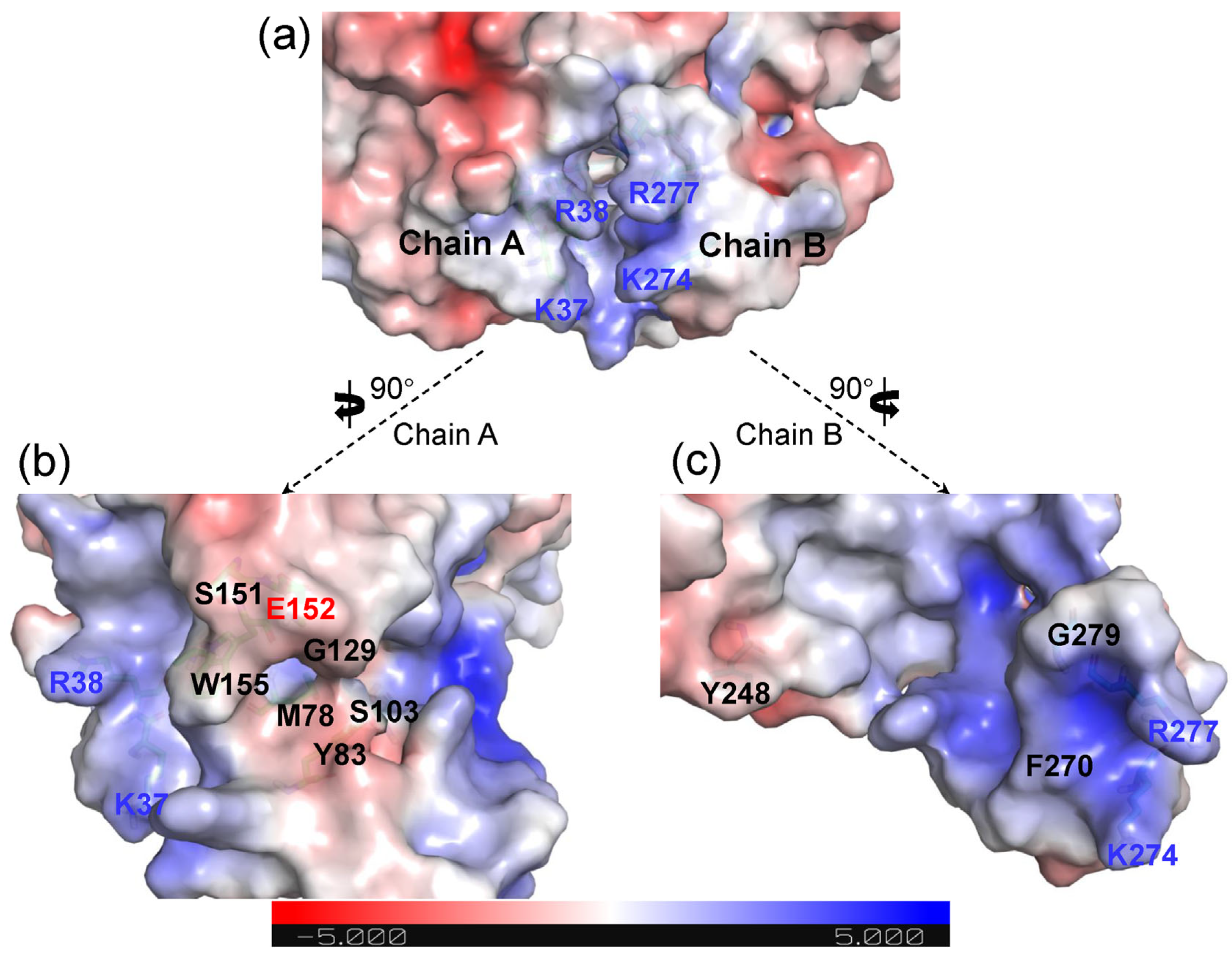
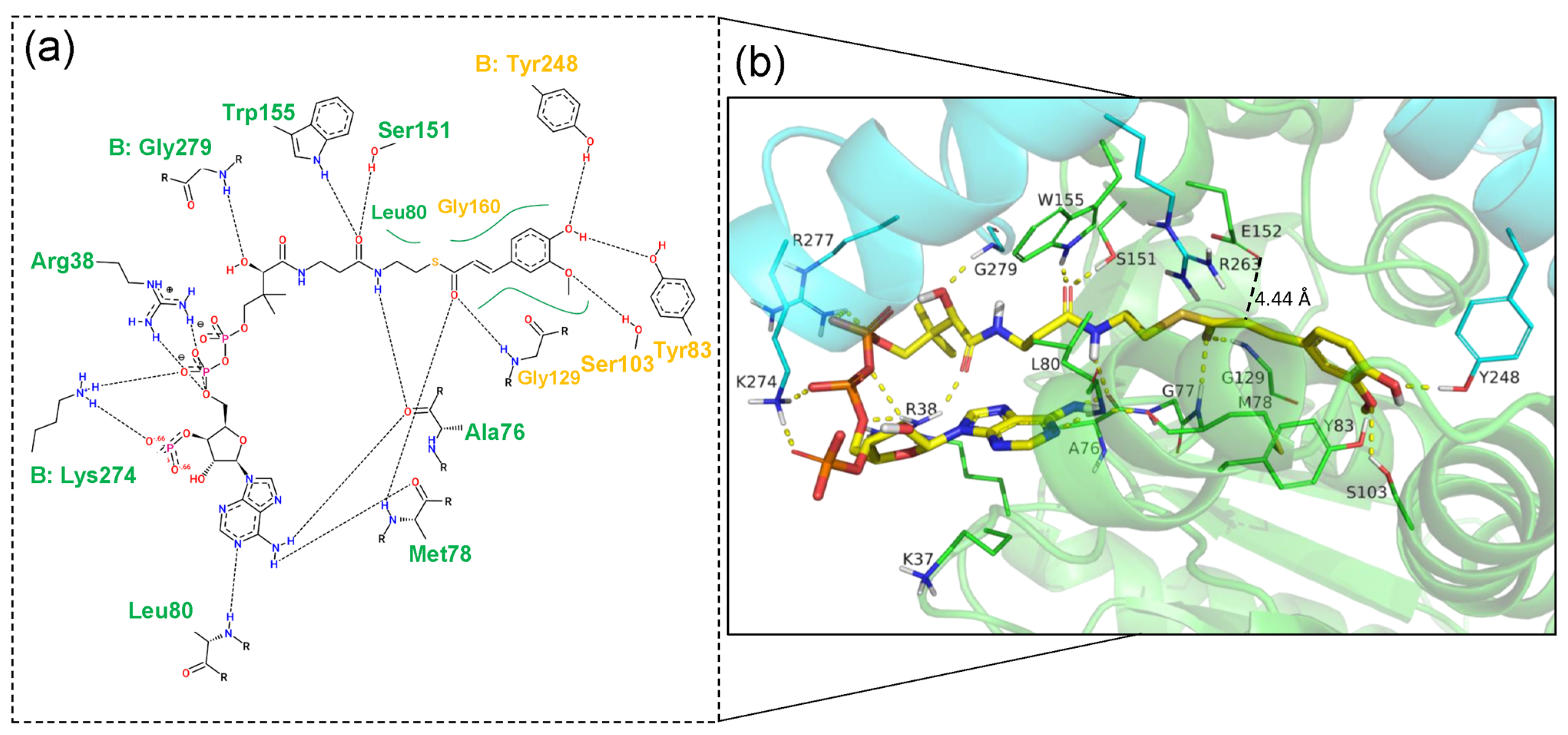
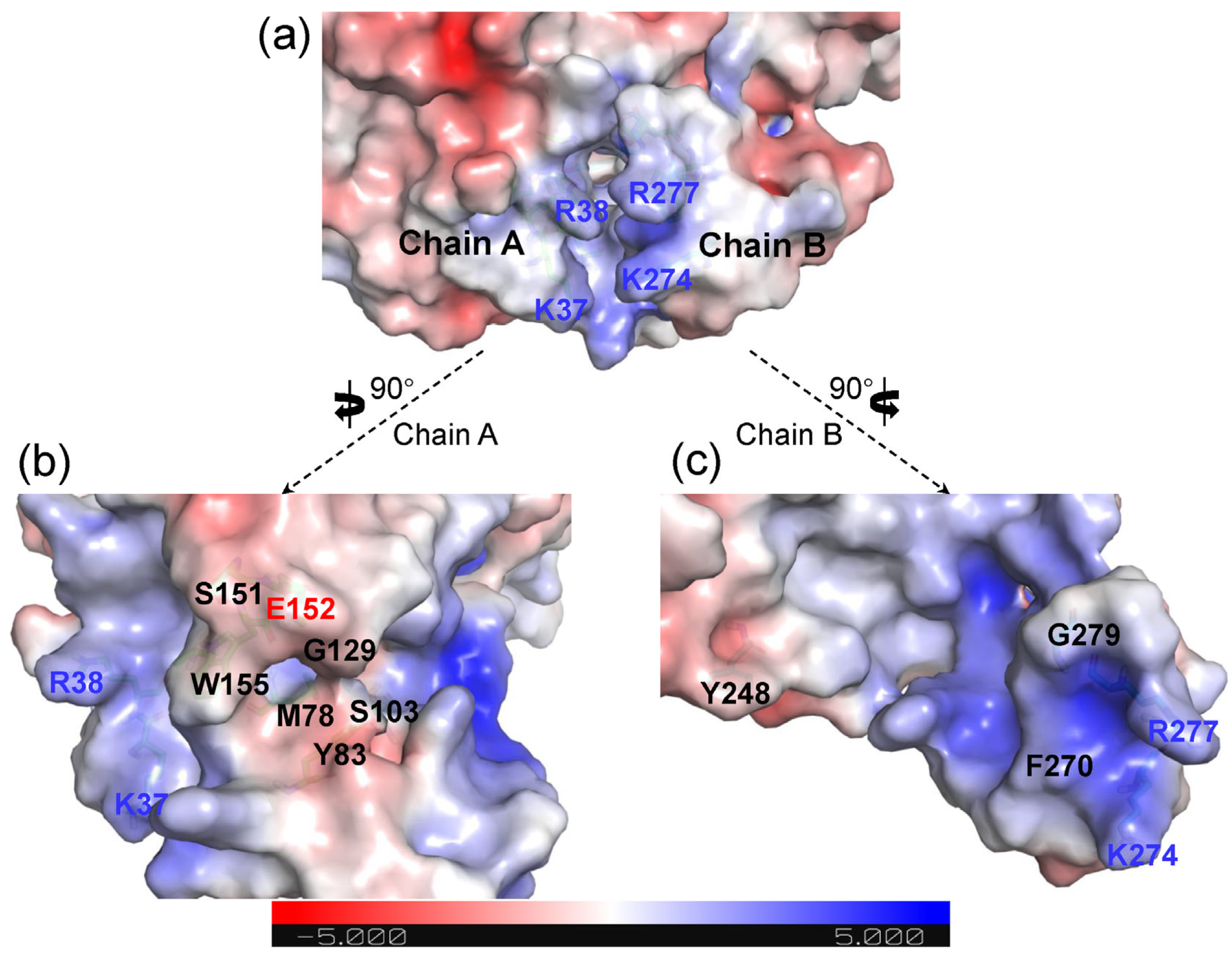
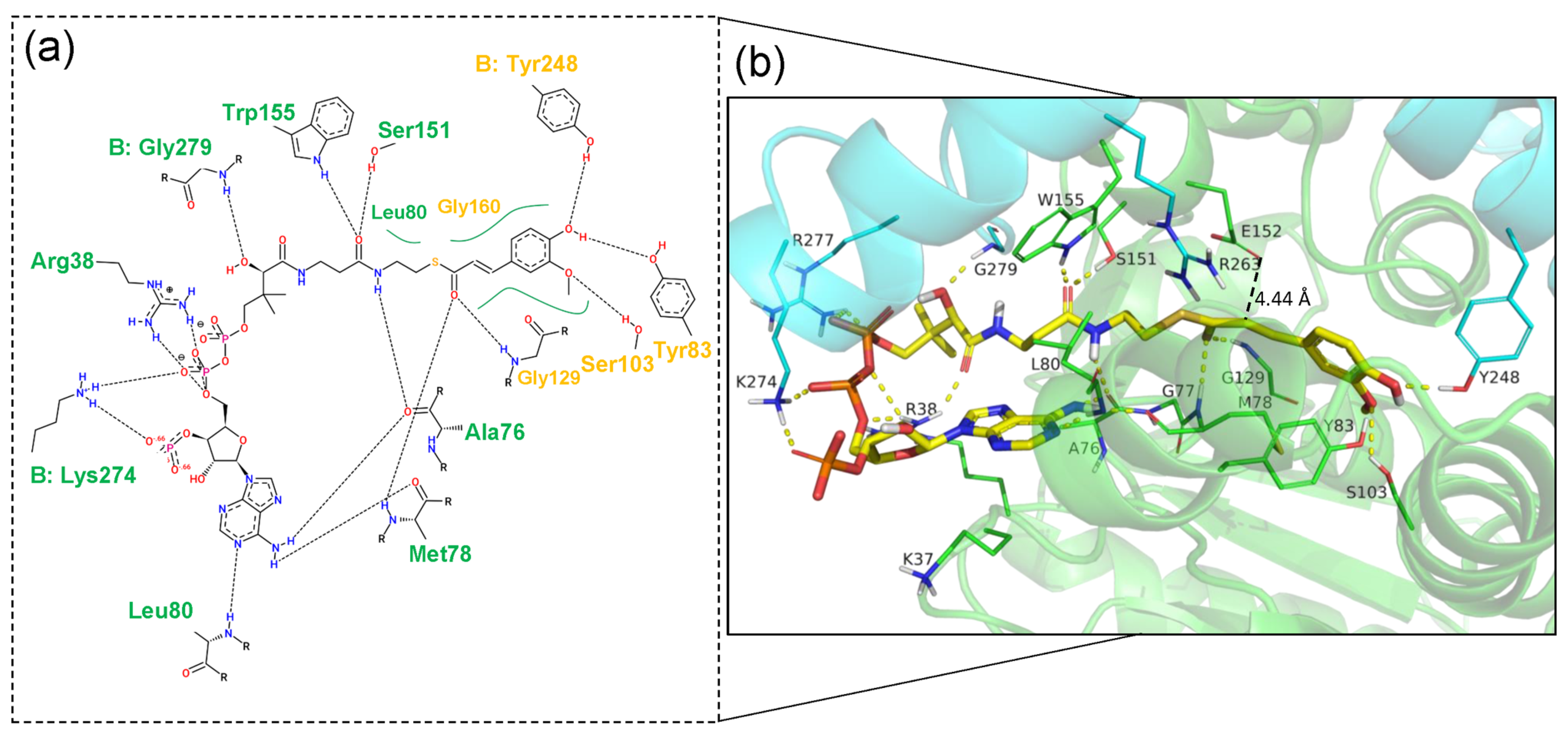
2.2. Structural Features of Ech and Its Binding Mode with FCA
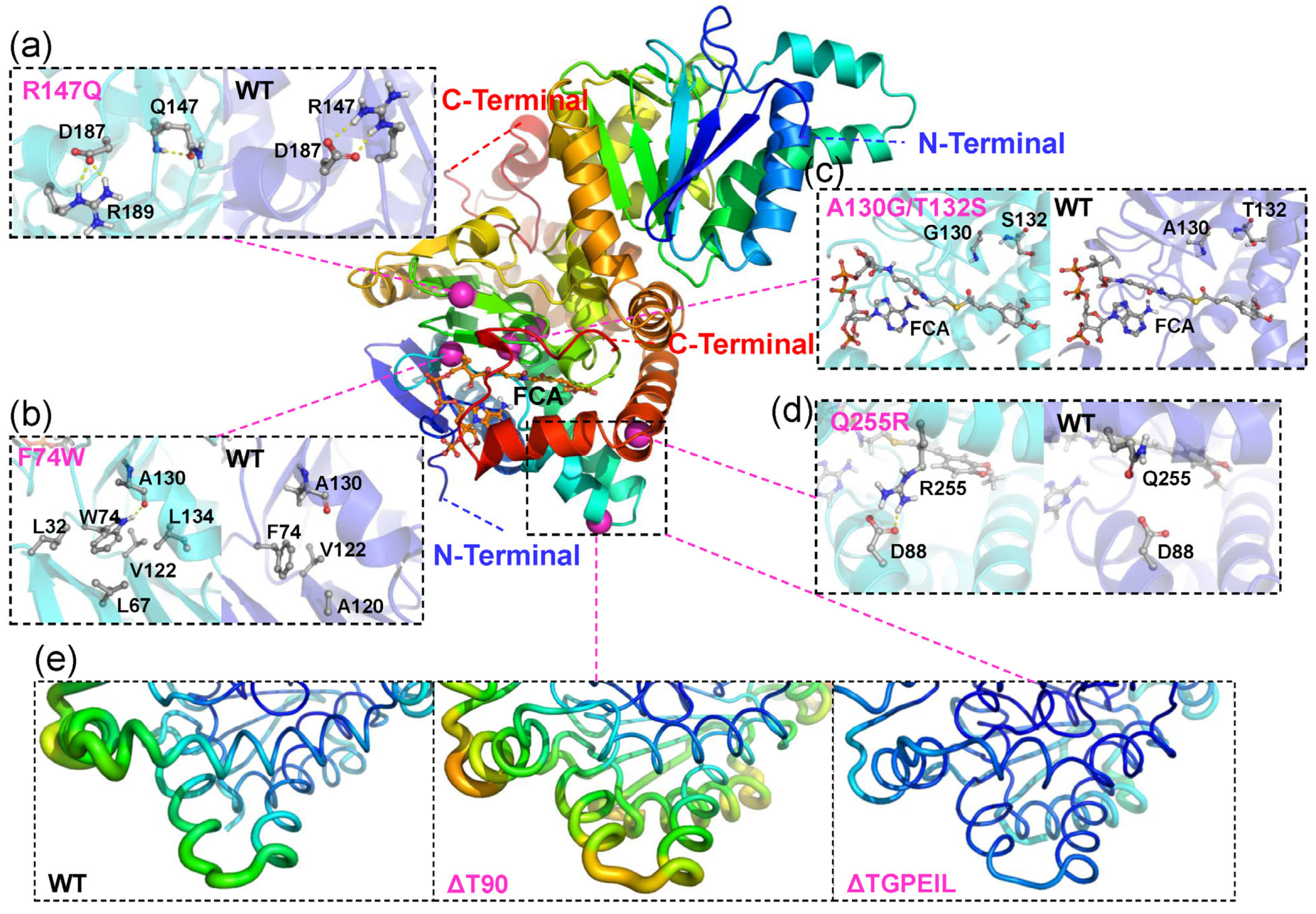
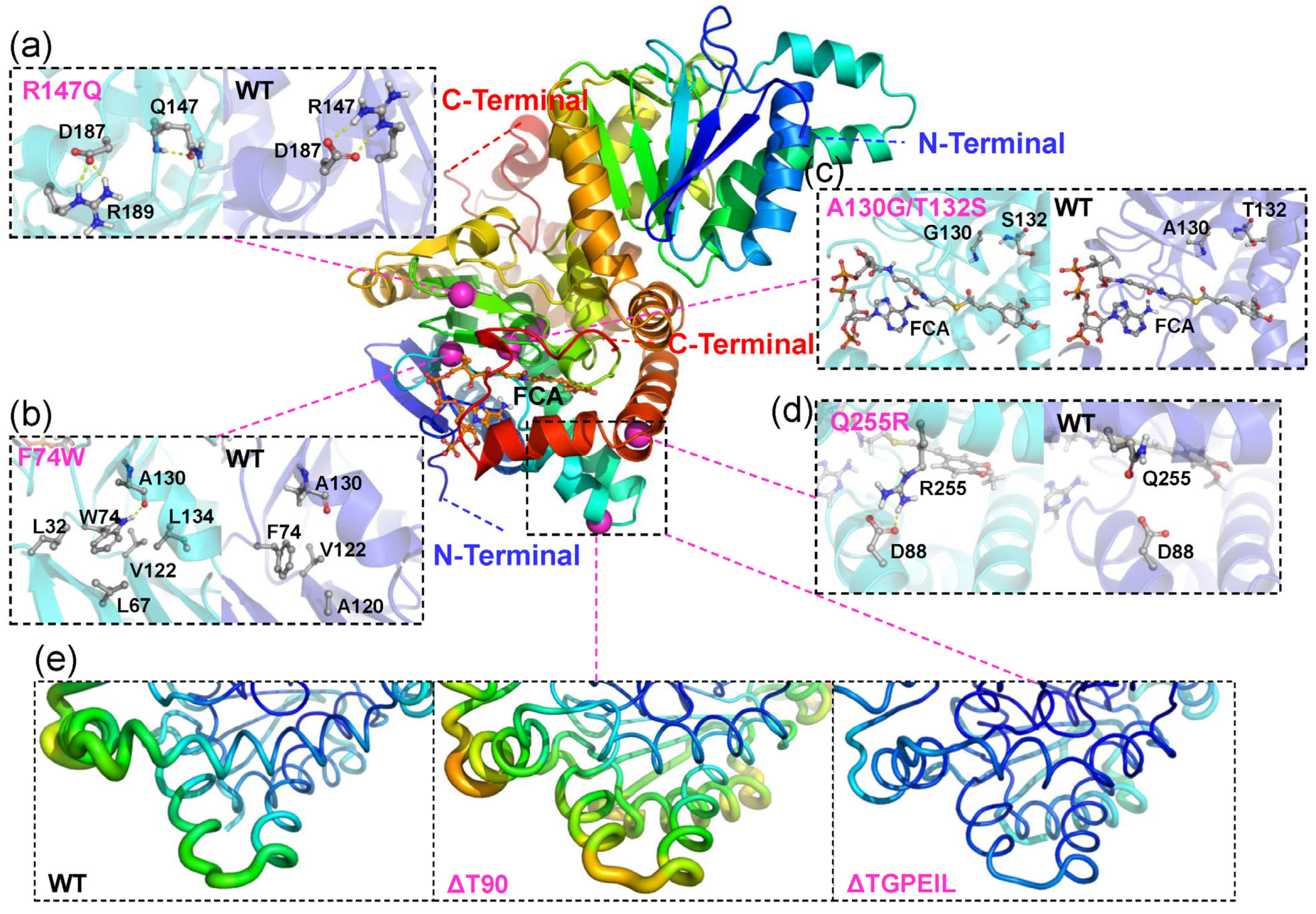
2.3. Rational Design for Ech
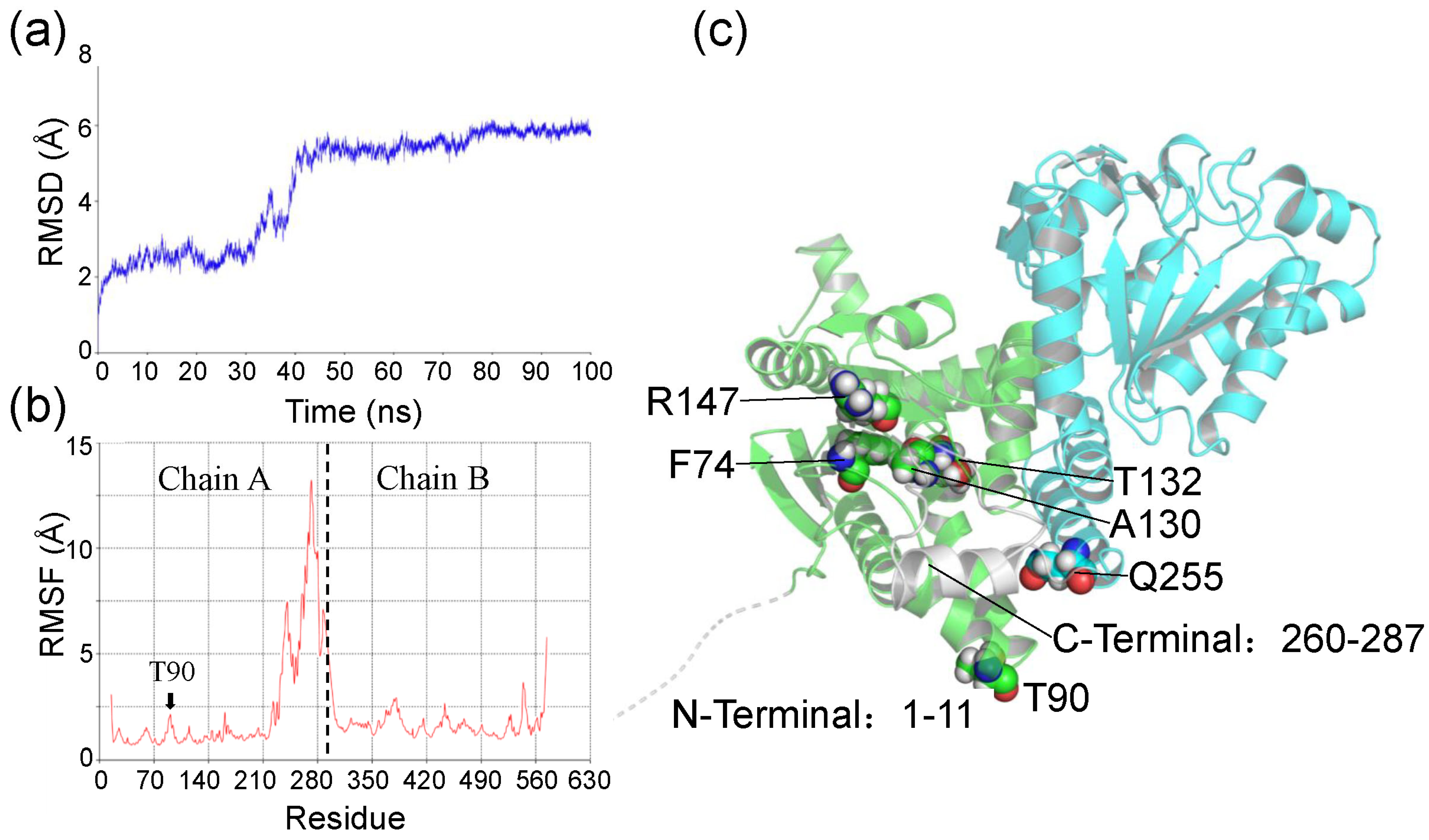
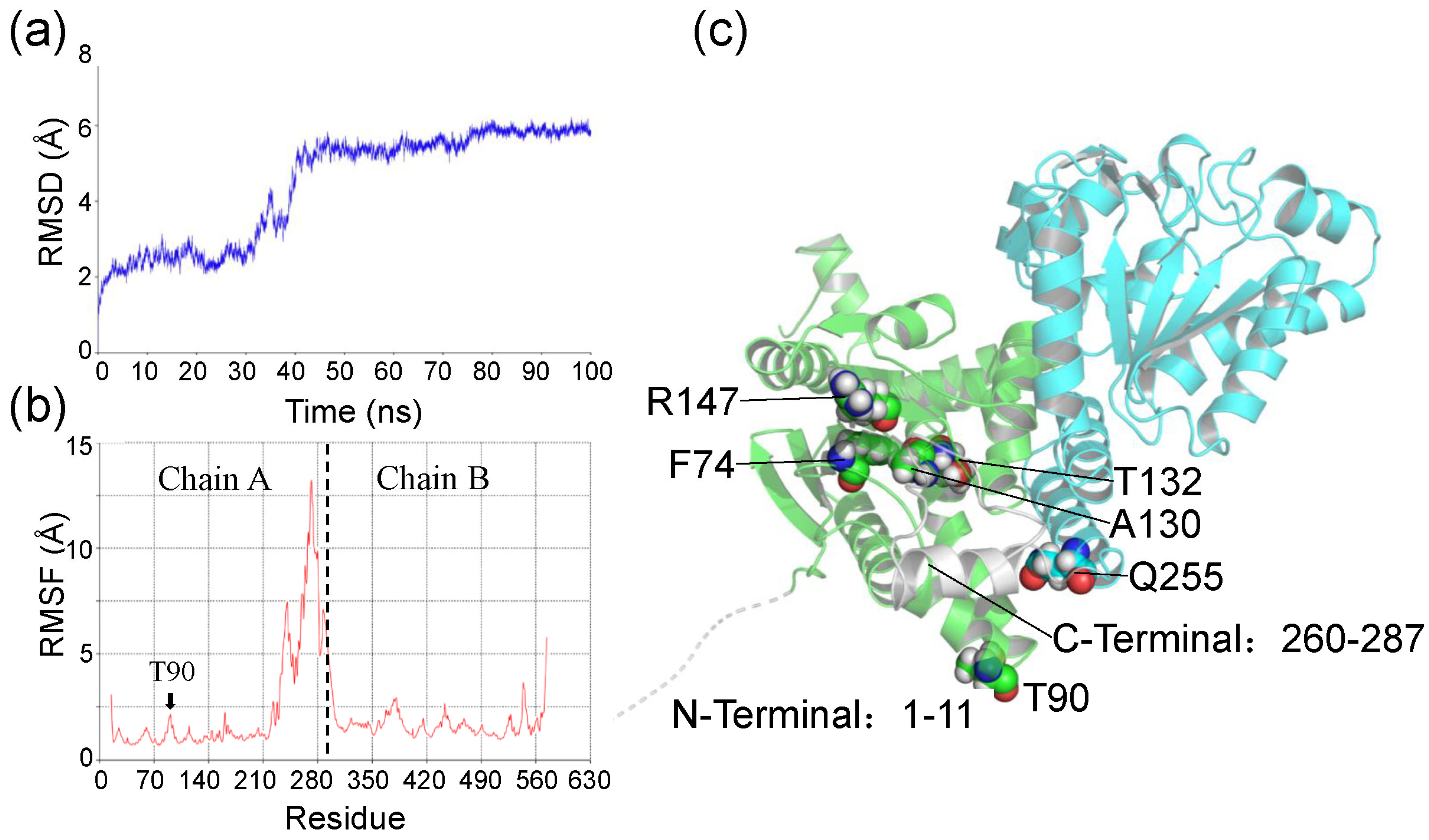
2.4. Molecular Dynamics Simulation and Binding Free Energy Analysis for Ech and Its Mutants
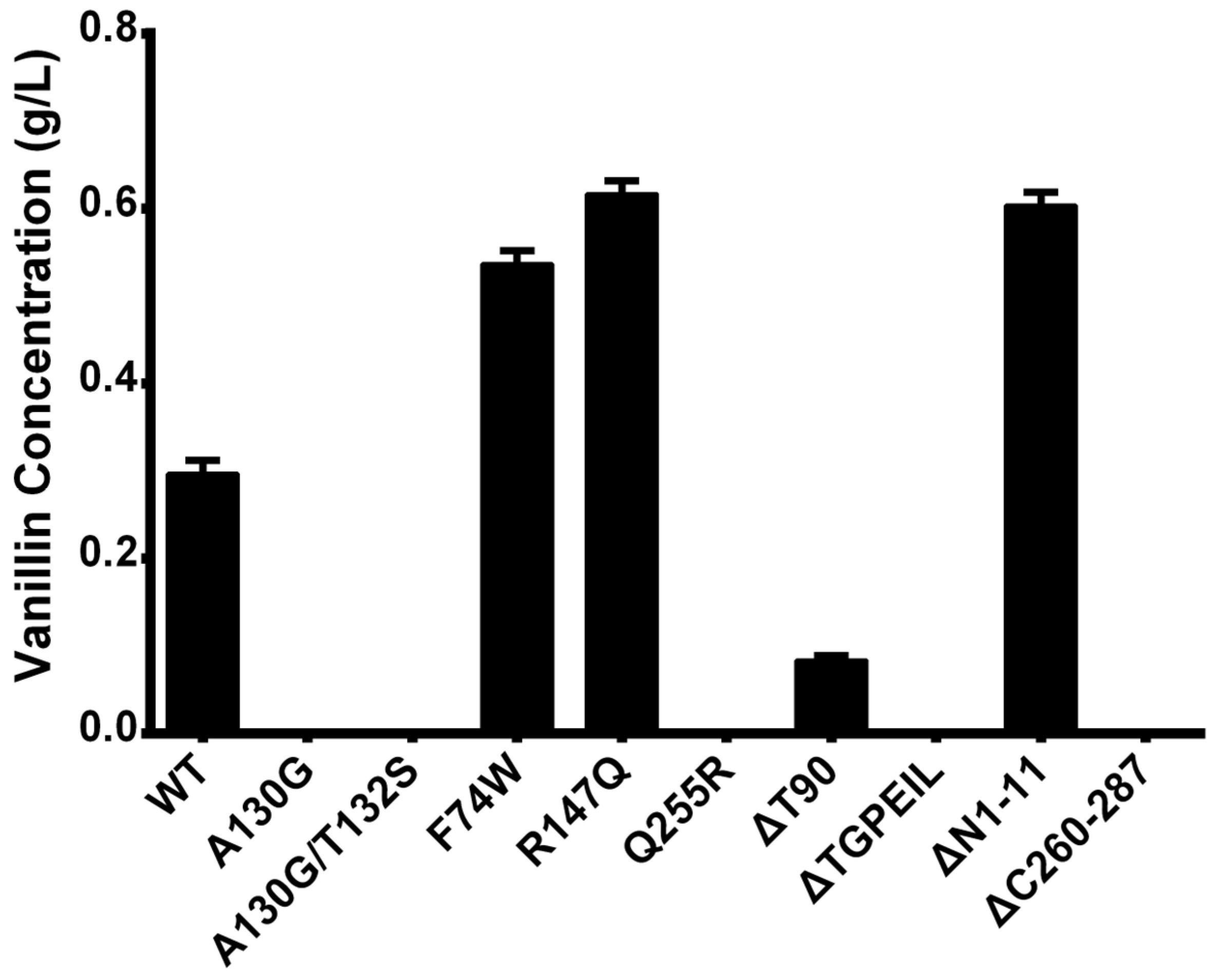
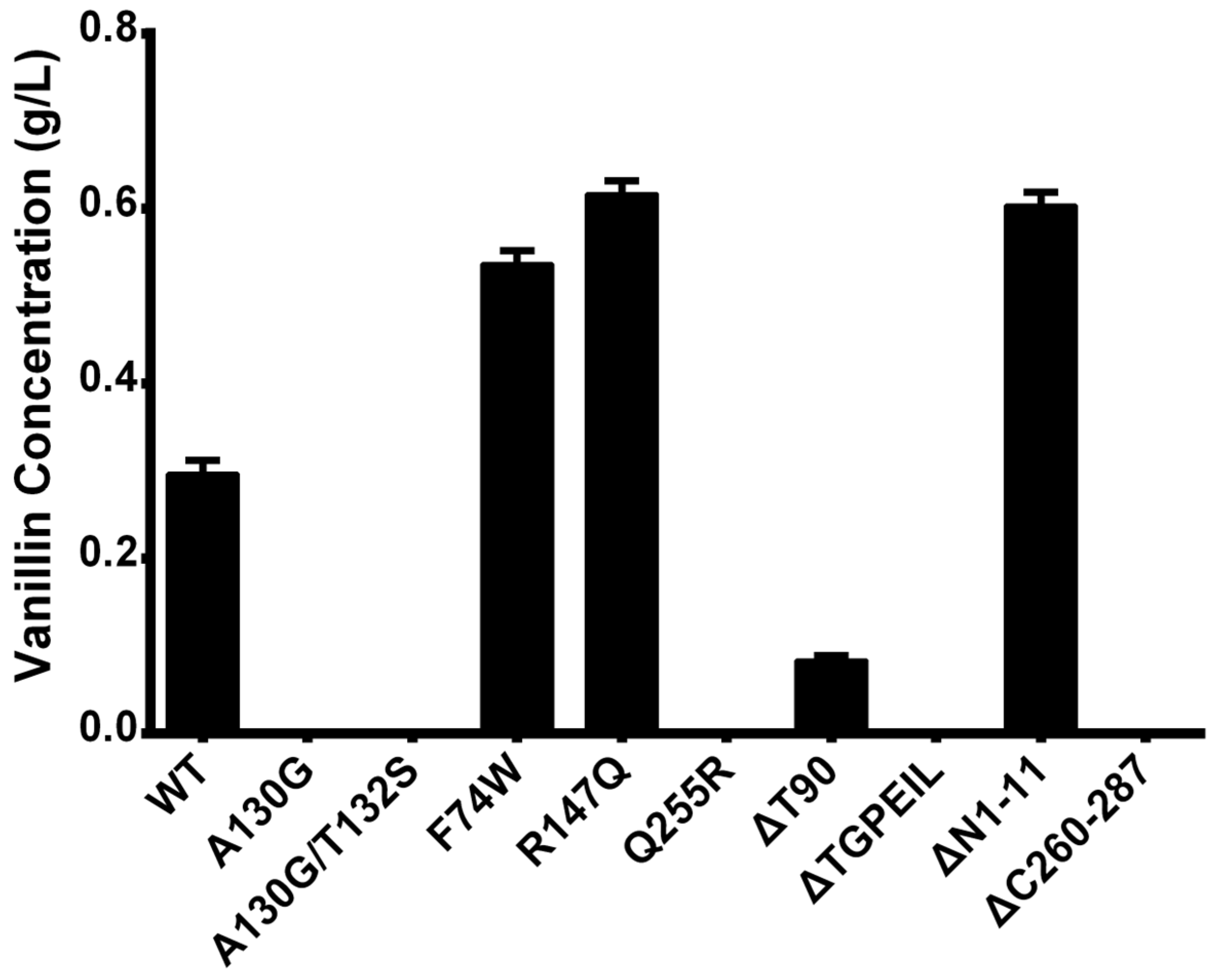
2.5. Experimental Verification of Wild-Type Ech and Its Mutants
3. Discussion
4. Materials and Methods
4.1. Bioinformatics Analysis
4.1.1. Multiple Sequence Alignment and Conservation Analysis
4.1.2. Homologous Modeling and Molecular Docking
4.1.3. Molecular Dynamics (MD) Simulation and Binding Free Energy Calculations
4.2. Experimental Verification
4.2.1. Strains, Plasmids and Reagents
4.2.2. Whole Cell Catalysis
4.2.3. Product Identification and Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Anuradha, K.; Shyamala, B.N.; Naidu, M.M. Vanilla—Its Science of Cultivation, Curing, Chemistry, and Nutraceutical Properties. Crit. Rev. Food Sci. Nutr. 2013, 53, 1250–1276. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Liu, J.; Liao, Y.; Lv, W.; Ma, L.; Wang, C. Degradation of Vanillin During Lignin Valorization Under Alkaline Oxidation. Top. Curr. Chem. 2018, 376, 29. [Google Scholar] [CrossRef]
- Martău, G.A.; Călinoiu, L.-F.; Vodnar, D.C. Bio-vanillin: Towards a sustainable industrial production. Trends Food Sci. Technol. 2021, 109, 579–592. [Google Scholar] [CrossRef]
- García-Bofill, M.; Sutton, P.W.; Straatman, H.; Brummund, J.; Schürmann, M.; Guillén, M.; Álvaro, G. Biocatalytic synthesis of vanillin by an immobilised eugenol oxidase: High biocatalyst yield by enzyme recycling. Appl. Catal. A Gen. 2020, 610, 117934. [Google Scholar] [CrossRef]
- Paul, V.; Rai, D.C.; Ramyaa Lakshmi, T.S.; Srivastava, S.K.; Tripathi, A.D. A comprehensive review on vanillin: Its microbial synthesis, isolation and recovery. Food Biotechnol. 2021, 35, 22–49. [Google Scholar] [CrossRef]
- Tadasa, K. Degradation of Eugenol by a Microorganism. Agric. Biol. Chem. 1977, 41, 925–929. [Google Scholar] [CrossRef]
- Lone, B.A.; Bhushan, A.; Ganjoo, A.; Katoch, M.; Gairola, S.; Gupta, P.; Babu, V. Biotransformation of eugenol by an endophytic fungus Daldinia sp. IIIMF4010 isolated from Rosmarinus officinalis. Nat. Prod. Res. 2023, 37, 535–541. [Google Scholar] [CrossRef]
- Ashengroph, M.; Nahvi, I.; Zarkesh-Esfahani, H.; Momenbeik, F. Conversion of Isoeugenol to Vanillin by Psychrobacter sp. Strain CSW4. Appl. Biochem. Biotechnol. 2012, 166, 1–12. [Google Scholar] [CrossRef]
- Ashengroph, M.; Nahvi, I.; Zarkesh-Esfahani, H.; Momenbeik, F. Pseudomonas resinovorans SPR1, a newly isolated strain with potential of transforming eugenol to vanillin and vanillic acid. New Biotechnol. 2011, 28, 656–664. [Google Scholar] [CrossRef]
- Plaggenborg, R.; Overhage, J.; Loos, A.; Archer, J.A.C.; Lessard, P.; Sinskey, A.J.; Steinbüchel, A.; Priefert, H. Potential of Rhodococcus strains for biotechnological vanillin production from ferulic acid and eugenol. Appl. Microbiol. Biotechnol. 2006, 72, 745–755. [Google Scholar] [CrossRef]
- Karakaya, H.; Yilmaztekin, M. Natural Vanillin Production from Isoeugenol by Using Pseudomonas putida in Biphasic Bioconversion Medium. Tarım Bilim. Derg. 2021, 28, 423–429. [Google Scholar] [CrossRef]
- Ashengroph, M.; Nahvi, I.; Zarkesh-Esfahani, H.; Momenbeik, F. Use of Growing Cells of Pseudomonas aeruginosa for Synthesis of the Natural Vanillin via Conversion of Isoeugenol. Iran. J. Pharm. Res. 2011, 10, 749–757. [Google Scholar]
- García-Bofill, M.; Sutton, P.W.; Guillén, M.; Álvaro, G. Enzymatic synthesis of vanillin catalysed by an eugenol oxidase. Appl. Catal. A Gen. 2019, 582, 117117. [Google Scholar] [CrossRef]
- Liu, H.; Zou, Y.; Yao, C.; Yang, Z. Enzymatic synthesis of vanillin and related catalytic mechanism. Flavour Fragr. J. 2020, 35, 51–58. [Google Scholar] [CrossRef]
- Ni, J.; Wu, Y.-T.; Tao, F.; Peng, Y.; Xu, P. A Coenzyme-Free Biocatalyst for the Value-Added Utilization of Lignin-Derived Aromatics. J. Am. Chem. Soc. 2018, 140, 16001–16005. [Google Scholar] [CrossRef]
- Gasson, M.J.; Kitamura, Y.; McLauchlan, W.R.; Narbad, A.; Parr, A.J.; Parsons, E.L.H.; Payne, J.; Rhodes, M.J.C.; Walton, N.J. Metabolism of Ferulic Acid to Vanillin. J. Biol. Chem. 1998, 273, 4163–4170. [Google Scholar] [CrossRef] [PubMed]
- Overhage, J.; Priefert, H.; Steinbüchel, A. Biochemical and Genetic Analyses of Ferulic Acid Catabolism in Pseudomonas sp. Strain HR199. Appl. Environ. Microbiol. 1999, 65, 4837–4847. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.-H.; Li, C.; Lee, Y.-M.; Lee, S.-H.; Kim, S.-H.; Choi, M.-S.; Seo, W.-T.; Yang, J.-K.; Kim, J.-Y.; Kim, S.-W. Production of vanillin from ferulic acid using recombinant strains of Escherichia coli. Biotechnol. Bioprocess Eng. 2005, 10, 378–384. [Google Scholar] [CrossRef]
- Graf, N.; Altenbuchner, J. Genetic engineering of Pseudomonas putida KT2440 for rapid and high-yield production of vanillin from ferulic acid. Appl. Microbiol. Biotechnol. 2013, 98, 137–149. [Google Scholar] [CrossRef]
- Di Gioia, D.; Luziatelli, F.; Negroni, A.; Ficca, A.G.; Fava, F.; Ruzzi, M. Metabolic engineering of Pseudomonas fluorescens for the production of vanillin from ferulic acid. J. Biotechnol. 2011, 156, 309–316. [Google Scholar] [CrossRef]
- Kaur, B.; Chakraborty, D.; Kumar, B. Metabolic engineering of Pediococcus acidilactici BD16 for production of vanillin through ferulic acid catabolic pathway and process optimization using response surface methodology. Appl. Microbiol. Biotechnol. 2014, 98, 8539–8551. [Google Scholar] [CrossRef]
- Liu, X.; Kong, J.-Q. Steroids hydroxylation catalyzed by the monooxygenase mutant 139-3 from Bacillus megaterium BM3. Acta Pharm. Sin. B 2017, 7, 510–516. [Google Scholar] [CrossRef]
- Chen, P.; Yan, L.; Wu, Z.; Li, S.; Bai, Z.; Yan, X.; Wang, N.; Liang, N.; Li, H. A microbial transformation using Bacillus subtilis B7-S to produce natural vanillin from ferulic acid. Sci. Rep. 2016, 6, 20400. [Google Scholar] [CrossRef]
- Yang, W.; Tang, H.; Ni, J.; Wu, Q.; Hua, D.; Tao, F.; Xu, P. Characterization of Two Streptomyces Enzymes that Convert Ferulic Acid to Vanillin. PLoS ONE 2013, 8, e67339. [Google Scholar] [CrossRef] [PubMed]
- Fleige, C.; Meyer, F.; Steinbüchel, A. Metabolic Engineering of the Actinomycete Amycolatopsis sp. Strain ATCC 39116 towards Enhanced Production of Natural Vanillin. Appl. Environ. Microbiol. 2016, 82, 3410–3419. [Google Scholar] [CrossRef]
- Arnold, F.H.; Volkov, A.A. Directed evolution of biocatalysts. Curr. Opin. Chem. Biol. 1999, 3, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Kuchner, O.; Arnold, F.H. Directed evolution of enzyme catalysts. Trends Biotechnol. 1997, 15, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xue, P.; Cao, M.; Yu, T.; Lane, S.T.; Zhao, H. Directed Evolution: Methodologies and Applications. Chem. Rev. 2021, 121, 12384–12444. [Google Scholar] [CrossRef]
- Yadav, S.; Pandey, A.K.; Dubey, S.K. Molecular modeling, docking and simulation dynamics of β-glucosidase reveals high-efficiency, thermo-stable, glucose tolerant enzyme in Paenibacillus lautus BHU3 strain. Int. J. Biol. Macromol. 2021, 168, 371–382. [Google Scholar] [CrossRef]
- Huang, P.; Lv, A.; Yan, Q.; Jiang, Z.; Yang, S. The structure and molecular dynamics of prolyl oligopeptidase from Microbulbifer arenaceous provide insights into catalytic and regulatory mechanisms. Acta Crystallogr. D Struct. Biol. 2022, 78, 735–751. [Google Scholar] [CrossRef] [PubMed]
- Taidi, L.; Maurady, A.; Britel, M.R. Molecular docking study and molecular dynamic simulation of human cyclooxygenase-2 (COX-2) with selected eutypoids. J. Biomol. Struct. Dyn. 2022, 40, 1189–1204. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Gibson, T.J.; Higgins, D.G. Multiple Sequence Alignment Using ClustalW and ClustalX. Curr. Protoc. Bioinform. 2003, 2, 2–3. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.A.; Clamp, M.; Barton, G.J. Jalview Version 2—A multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef]
- Crooks, G.E.; Hon, G.; Chandonia, J.-M.; Brenner, S.E. WebLogo: A Sequence Logo Generator. Genome Res. 2004, 14, 1188–1190. [Google Scholar] [CrossRef] [PubMed]
- Webb, B.; Sali, A. Comparative Protein Structure Modeling Using MODELLER. Curr. Protoc. Bioinform. 2016, 54, 5.6.1–5.6.37. [Google Scholar] [CrossRef] [PubMed]
- Eberhardt, J.; Santos-Martins, D.; Tillack, A.F.; Forli, S. AutoDock Vina 1.2.0: New Docking Methods, Expanded Force Field, and Python Bindings. J. Chem. Inf. Model. 2021, 61, 3891–3898. [Google Scholar] [CrossRef]
- Salomon-Ferrer, R.; Case, D.A.; Walker, R.C. An overview of the Amber Biomolecular Simulation Package: Amber Biomolecular Simulation Package. WIREs Comput. Mol. Sci. 2013, 3, 198–210. [Google Scholar] [CrossRef]
- Maier, J.A.; Martinez, C.; Kasavajhala, K.; Wickstrom, L.; Hauser, K.E.; Simmerling, C. ff14SB: Improving the accuracy of protein side chain and backbone parameters from ff99SB. J. Chem. Theory Comput. 2015, 11, 3696–3713. [Google Scholar] [CrossRef]
- Sprenger, K.G.; Jaeger, V.W.; Pfaendtner, J. The General AMBER Force Field (GAFF) Can Accurately Predict Thermodynamic and Transport Properties of Many Ionic Liquids. J. Phys. Chem. B 2015, 119, 5882–5895. [Google Scholar] [CrossRef]
- Frisch, M.J.; Trucks, G.W.; Schlegel, H.B.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R.; Scalmani, G.; Barone, V.; Mennucci, B.; Petersson, G.A.; et al. Gaussian 09; Revision E.01; Gaussian, Inc.: Wallingford, CT, USA, 2009. [Google Scholar]
- Darden, T.; York, D.; Pedersen, L. Particle mesh Ewald: An N⋅log(N) method for Ewald sums in large systems. J. Chem. Phys. 1993, 98, 10089–10092. [Google Scholar] [CrossRef]
- Ryckaert, J.-P.; Ciccotti, G.; Berendsen, H.J.C. Numerical integration of the cartesian equations of motion of a system with constraints: Molecular dynamics of n-alkanes. J. Comput. Phys. 1977, 23, 327–341. [Google Scholar] [CrossRef]
- Izaguirre, J.A.; Catarello, D.P.; Wozniak, J.M.; Skeel, R.D. Langevin stabilization of molecular dynamics. J. Chem. Phys. 2001, 114, 2090–2098. [Google Scholar] [CrossRef]
- Hou, T.; Wang, J.; Li, Y.; Wang, W. Assessing the Performance of the MM/PBSA and MM/GBSA Methods. 1. The Accuracy of Binding Free Energy Calculations Based on Molecular Dynamics Simulations. J. Chem. Inf. Model. 2011, 51, 69–82. [Google Scholar] [CrossRef] [PubMed]
- Shao, J.; Tanner, S.W.; Thompson, N.; Cheatham, T.E. Clustering Molecular Dynamics Trajectories: 1. Characterizing the Performance of Different Clustering Algorithms. J. Chem. Theory Comput. 2007, 3, 2312–2334. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mutation Type | Mutant | Description |
|---|---|---|
| Single | A130G | Changing the size and flexibility of the active cavity |
| Double | A130G/T132S | Changing the size and flexibility of the active cavity |
| Single | R147Q | Substitution from Amycolatopsis thermoflava |
| Single | F74W | Substitution from Pseudomonas fluorescens |
| Single | Q255R | Substitution from Pseudomonas fluorescens |
| Deletion | ΔT90 | Reducing the flexibility of the structure |
| Deletion & Multiple | ΔTGPEIL | Reducing the flexibility of the structure |
| Truncation | ΔN1-11 | N-Terminal truncation |
| Truncation | ΔC260-287 | C-Terminal truncation |
| System | ΔEVDW | ΔEEL | ΔEGB | ΔESURF | ΔETotal |
|---|---|---|---|---|---|
| WT | −113.41 | 76.80 | −51.39 | −14.34 | −102.34 |
| A130G | −102.45 | −84.87 | 93.89 | −13.28 | −106.71 |
| A130G/T132S | −105.73 | −104.10 | 112.78 | −13.27 | −110.32 |
| F74W | −108.98 | 1.46 | 27.15 | −13.70 | −94.07 |
| R147Q | −98.27 | 35.37 | −27.28 | −12.81 | −102.99 |
| Q255R | −98.50 | −181.67 | 186.42 | −13.27 | −107.02 |
| ΔT90 | −101.89 | −42.82 | 59.27 | −13.49 | −98.93 |
| ΔTGPEIL | −108.30 | 67.65 | −43.93 | −13.79 | −98.37 |
| ΔC260-287 | −77.13 | 298.71 | −287.68 | −9.94 | −76.04 |
| Primers | Primers Sequence (5′→3′) |
|---|---|
| Bgl II-ech-F | GGAAGATCTATGAGCACAGCGGTCGGCAACGG |
| Xho I-ech-R | CCGCTCGAGCTACTTCTCCGGGTCGAAGGCGCTCAGG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ye, Q.; Xu, W.; He, Y.; Li, H.; Zhao, F.; Zhang, J.; Song, Y. Biosynthesis of Vanillin by Rational Design of Enoyl-CoA Hydratase/Lyase. Int. J. Mol. Sci. 2023, 24, 13631. https://doi.org/10.3390/ijms241713631
Ye Q, Xu W, He Y, Li H, Zhao F, Zhang J, Song Y. Biosynthesis of Vanillin by Rational Design of Enoyl-CoA Hydratase/Lyase. International Journal of Molecular Sciences. 2023; 24(17):13631. https://doi.org/10.3390/ijms241713631
Chicago/Turabian StyleYe, Qi, Weizhuo Xu, Yanan He, Hao Li, Fan Zhao, Jinghai Zhang, and Yongbo Song. 2023. "Biosynthesis of Vanillin by Rational Design of Enoyl-CoA Hydratase/Lyase" International Journal of Molecular Sciences 24, no. 17: 13631. https://doi.org/10.3390/ijms241713631
APA StyleYe, Q., Xu, W., He, Y., Li, H., Zhao, F., Zhang, J., & Song, Y. (2023). Biosynthesis of Vanillin by Rational Design of Enoyl-CoA Hydratase/Lyase. International Journal of Molecular Sciences, 24(17), 13631. https://doi.org/10.3390/ijms241713631