
Overexpression of BoLSU1 and BoLSU2 Confers Tolerance to Sulfur Deficiency in Arabidopsis by Manipulating Glucosinolate Metabolism

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
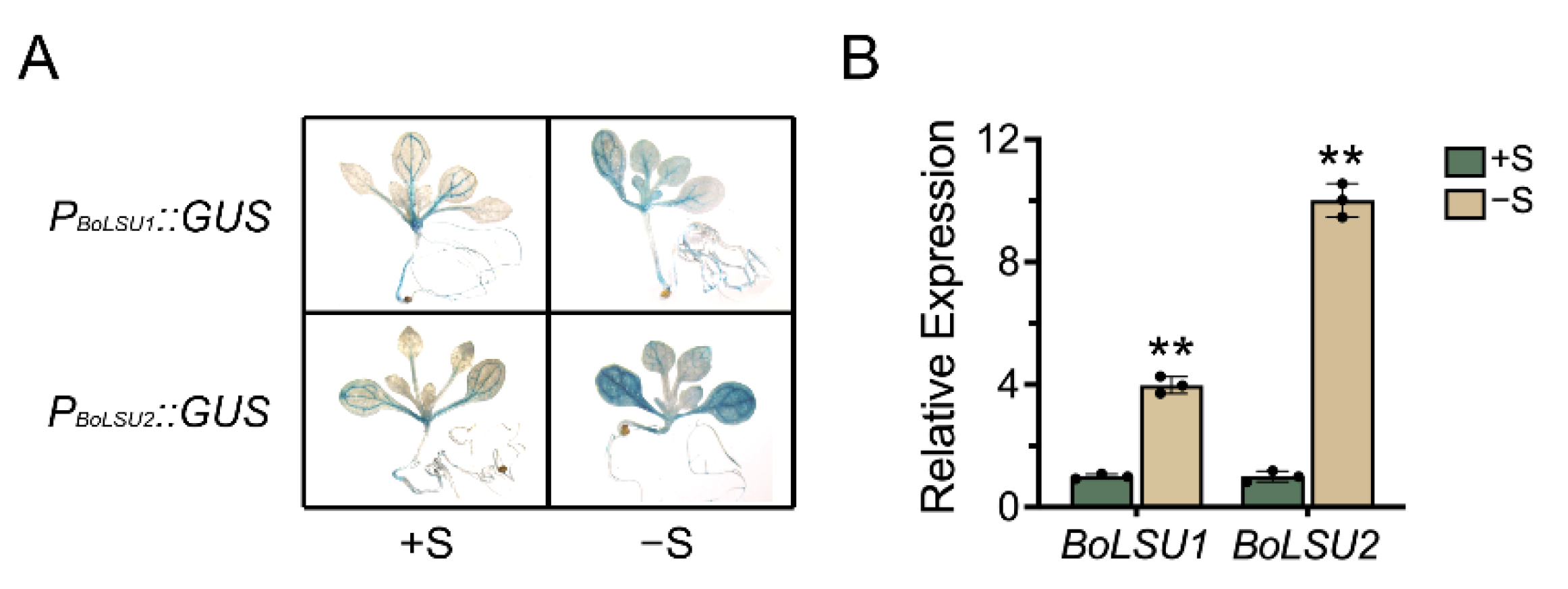
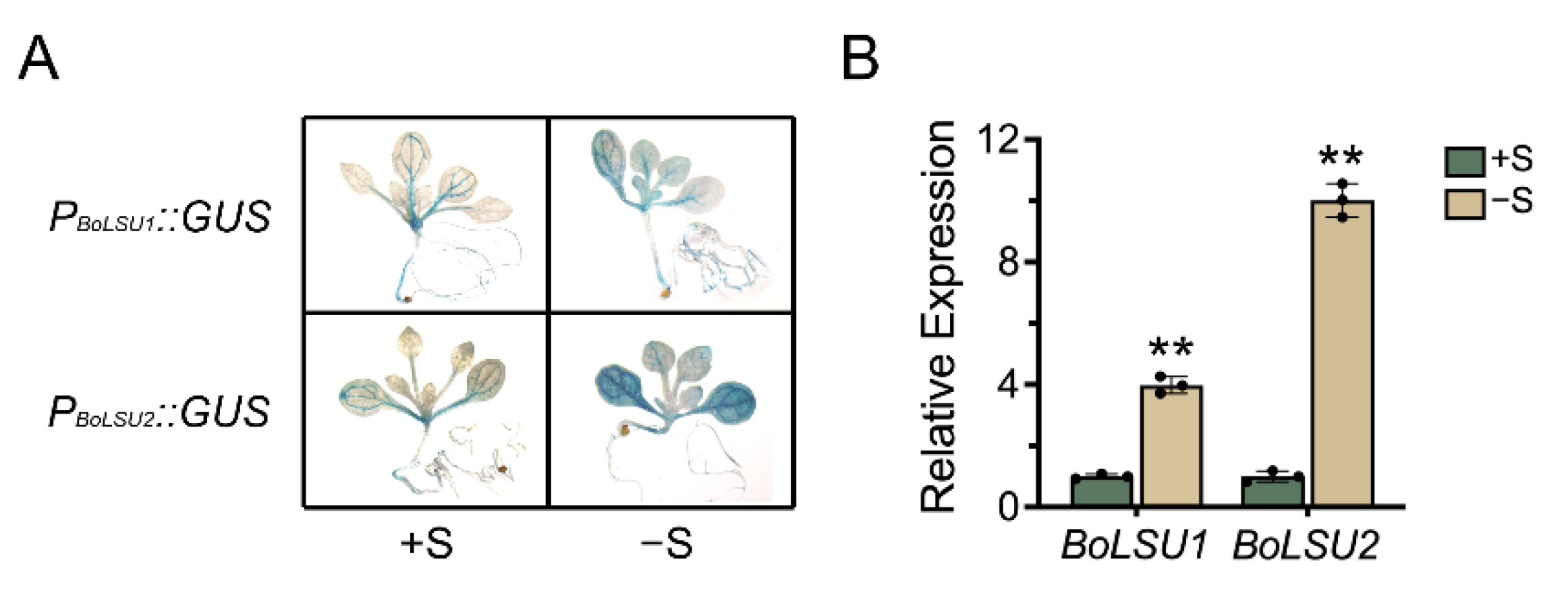
2.1. The Expression of BoLSU1 and BoLSU2 Is Induced by Sulfur Deficiency
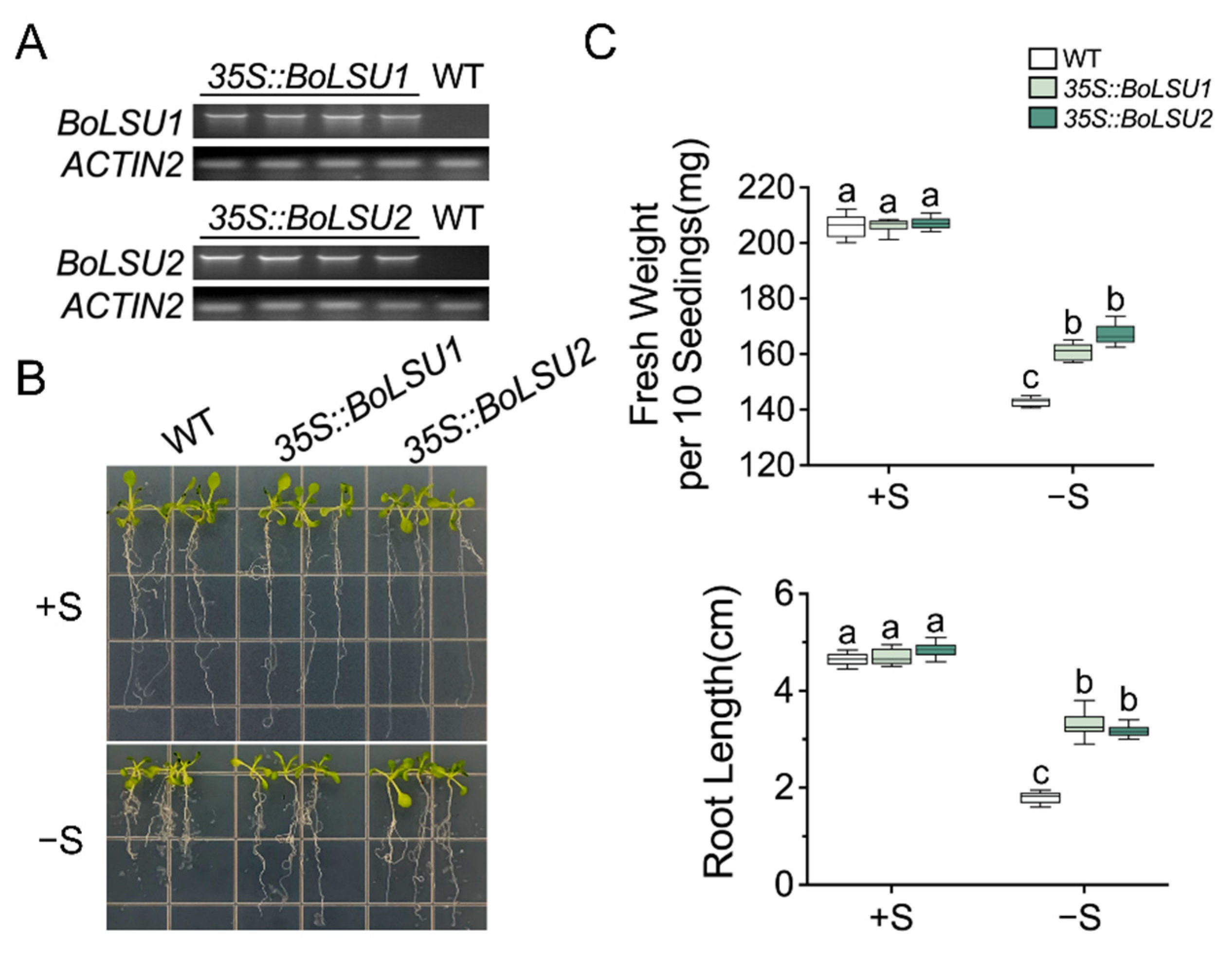
2.2. Overexpression of BoLSU1 and BoLSU2 Enhanced Tolerance to Sulfur Deficiency
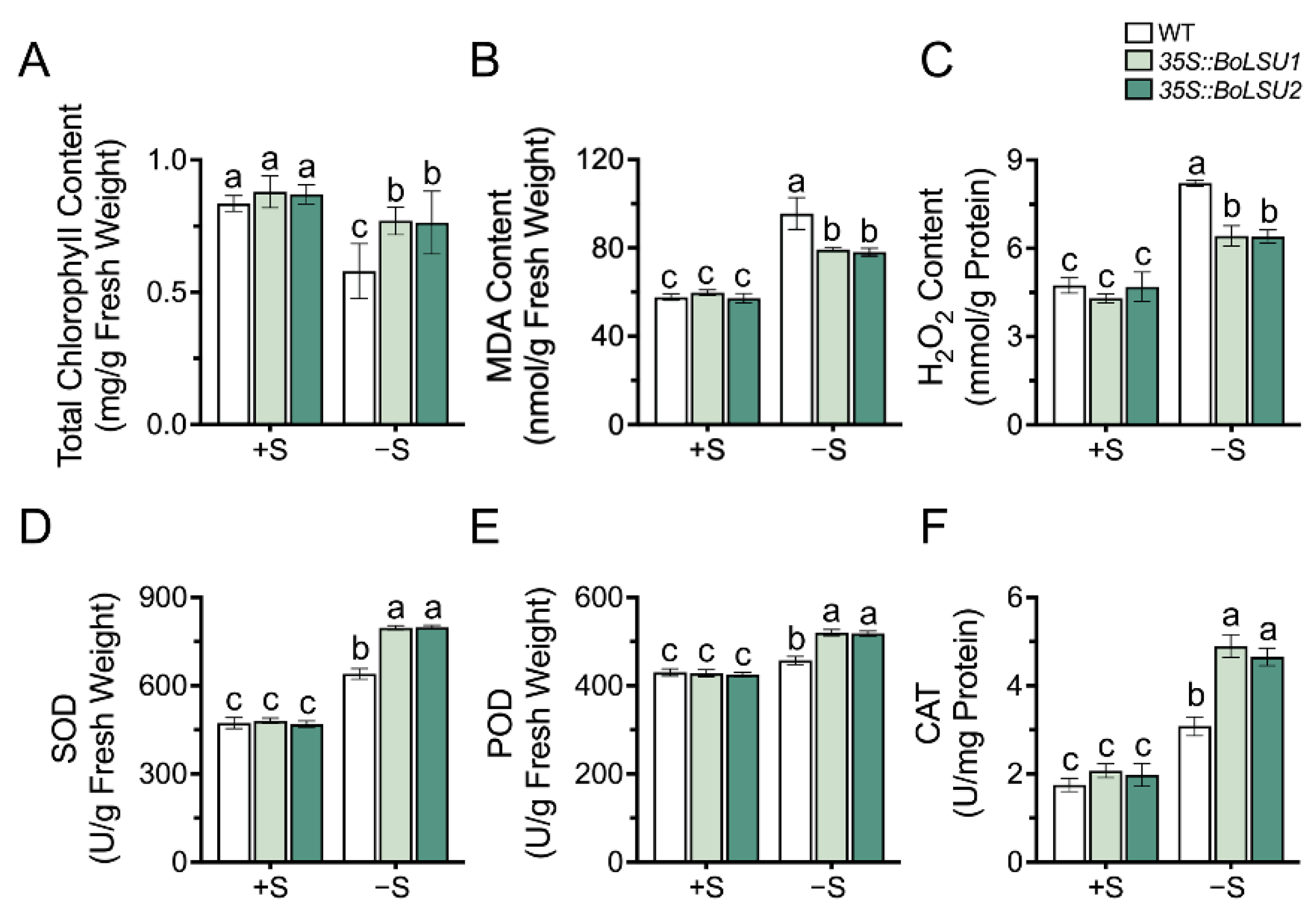
2.3. Overexpression of BoLSU1 and BoLSU2 Alleviated Oxidative Stress Induced by Sulfur Deficiency
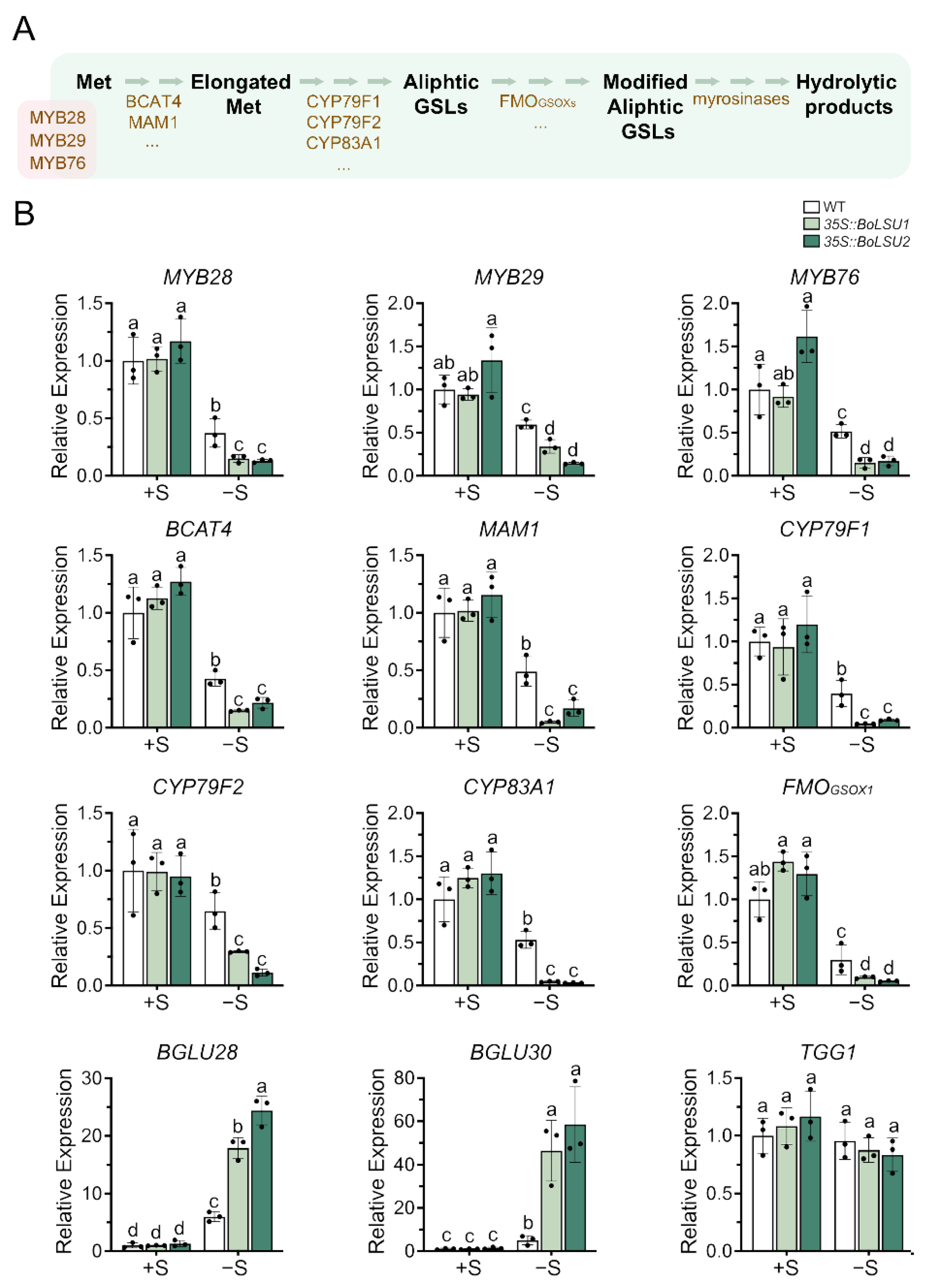
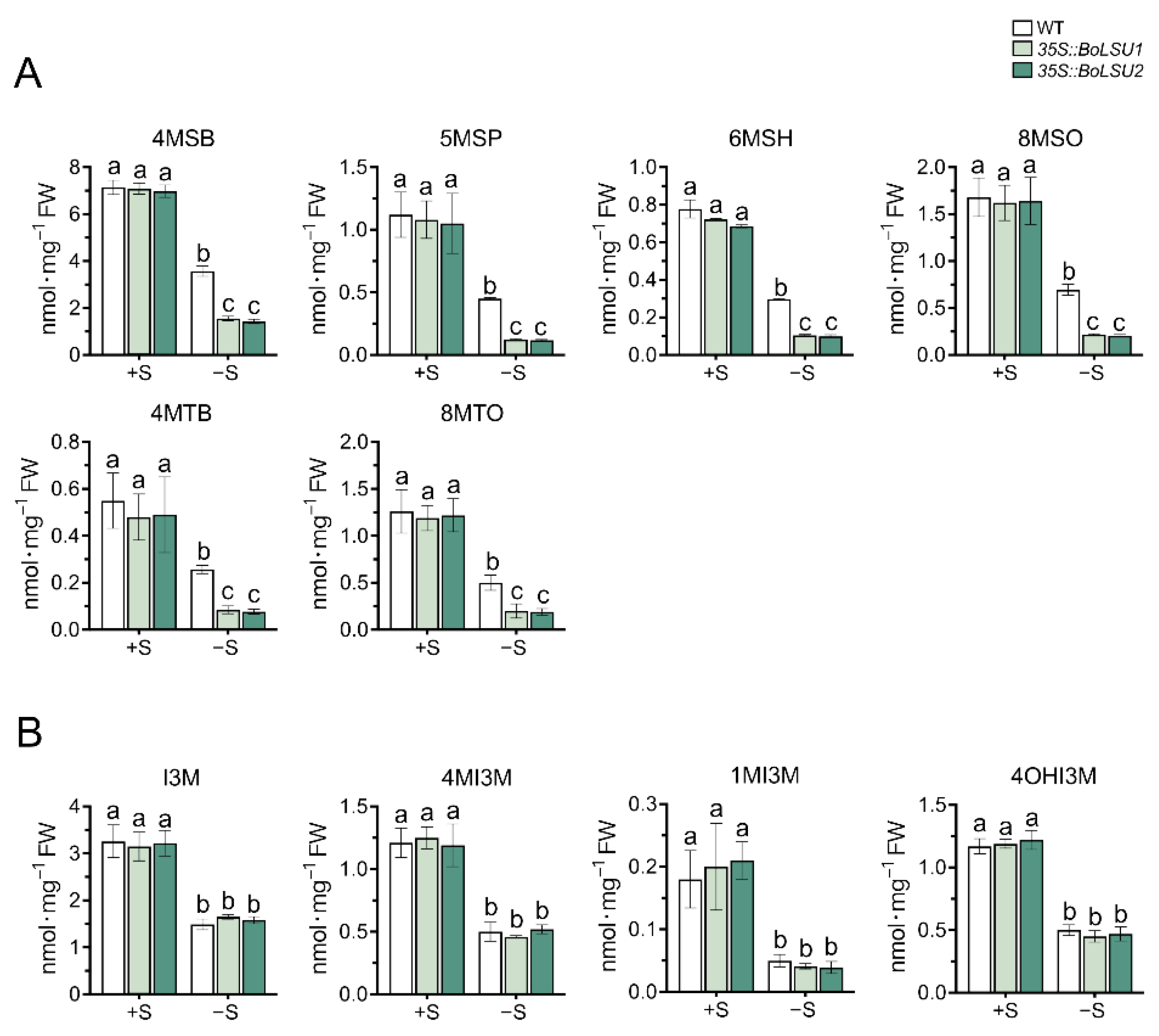
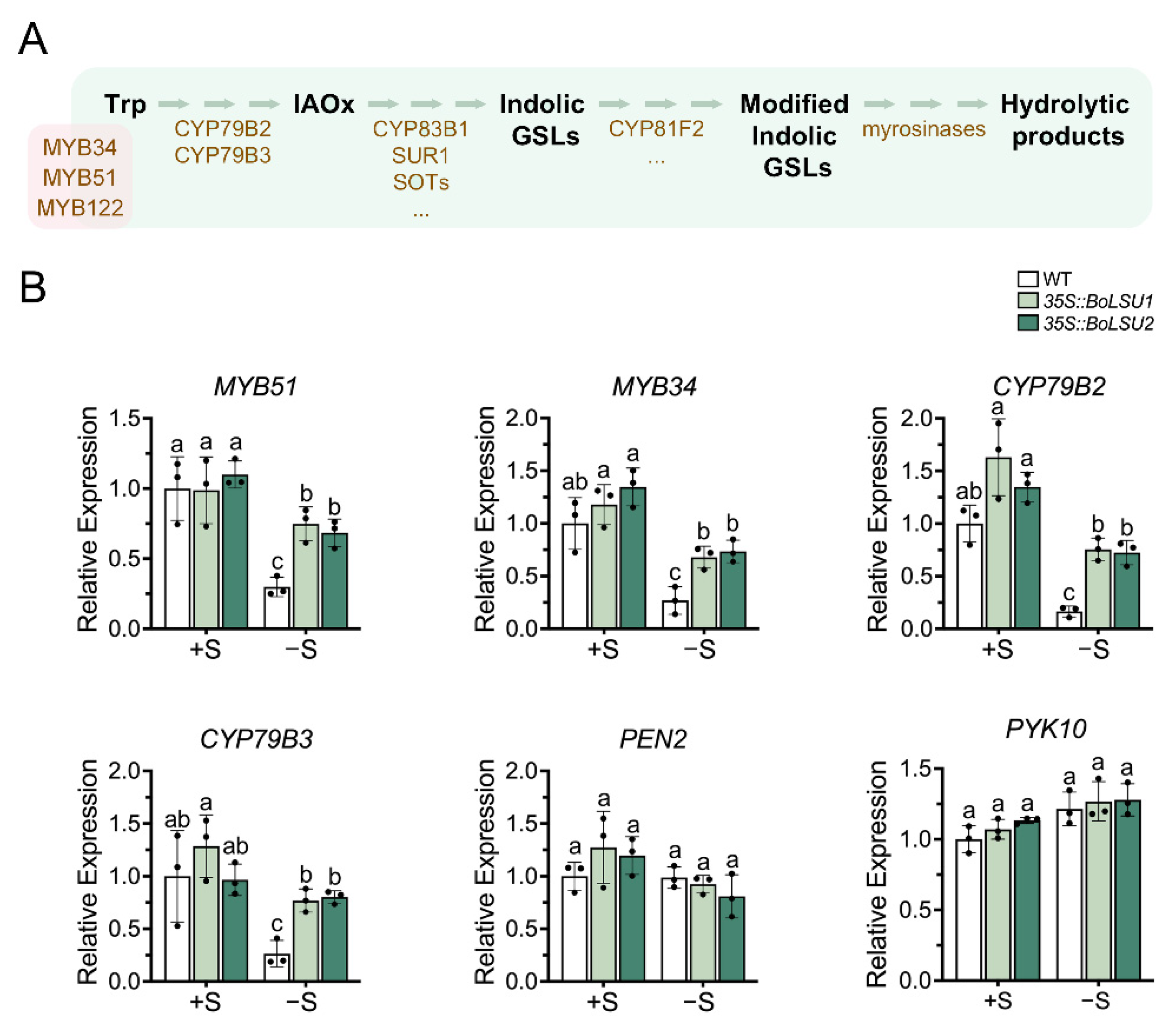
2.4. BoLSU1 and BoLSU2 Manipulated Glucosinolate Metabolism under Sulfur Deficiency
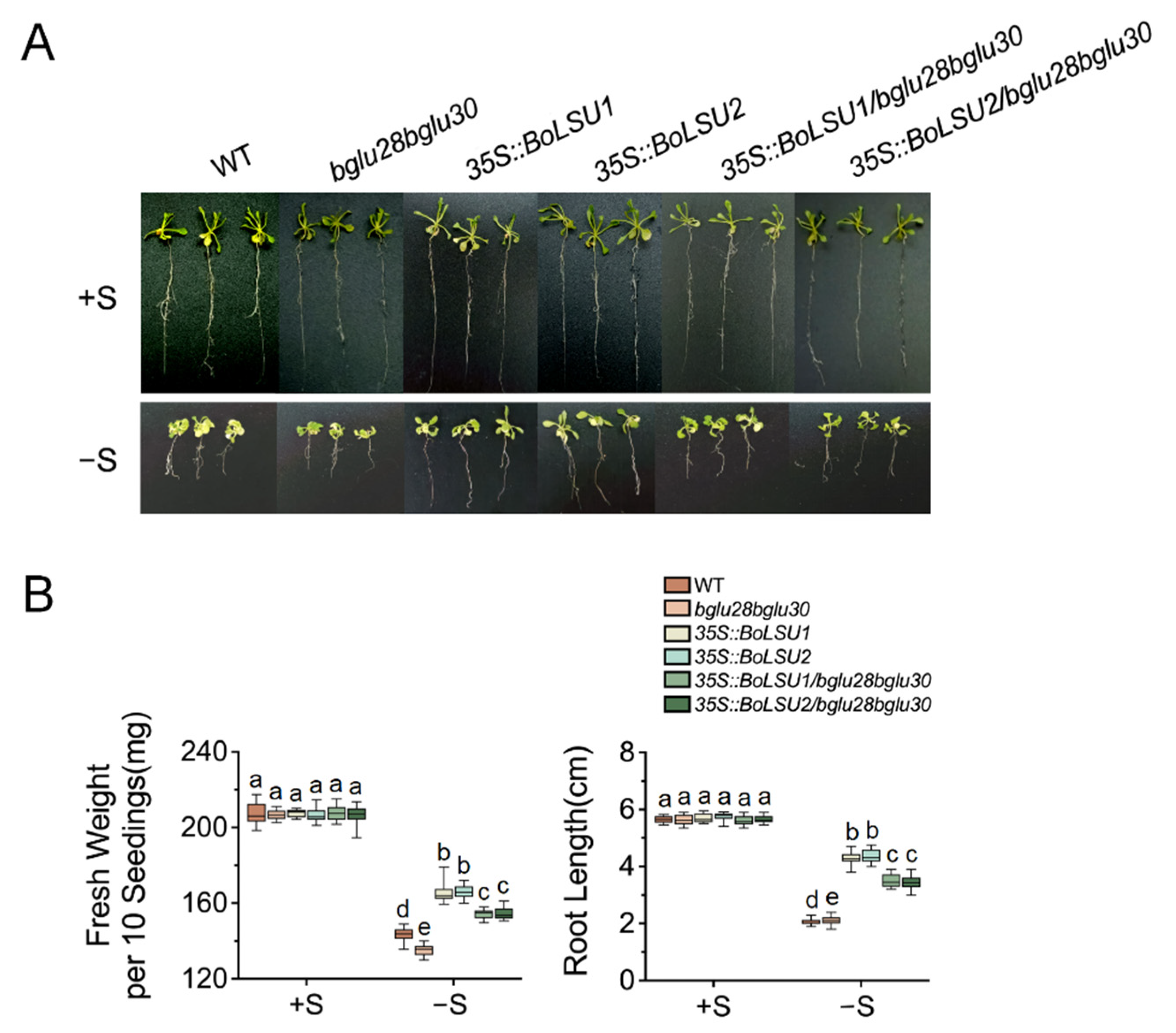
2.5. BoLSU1 and BoLSU2 Mediated Sulfur Deficiency Tolerance Depends on BGLU28 and BGLU30
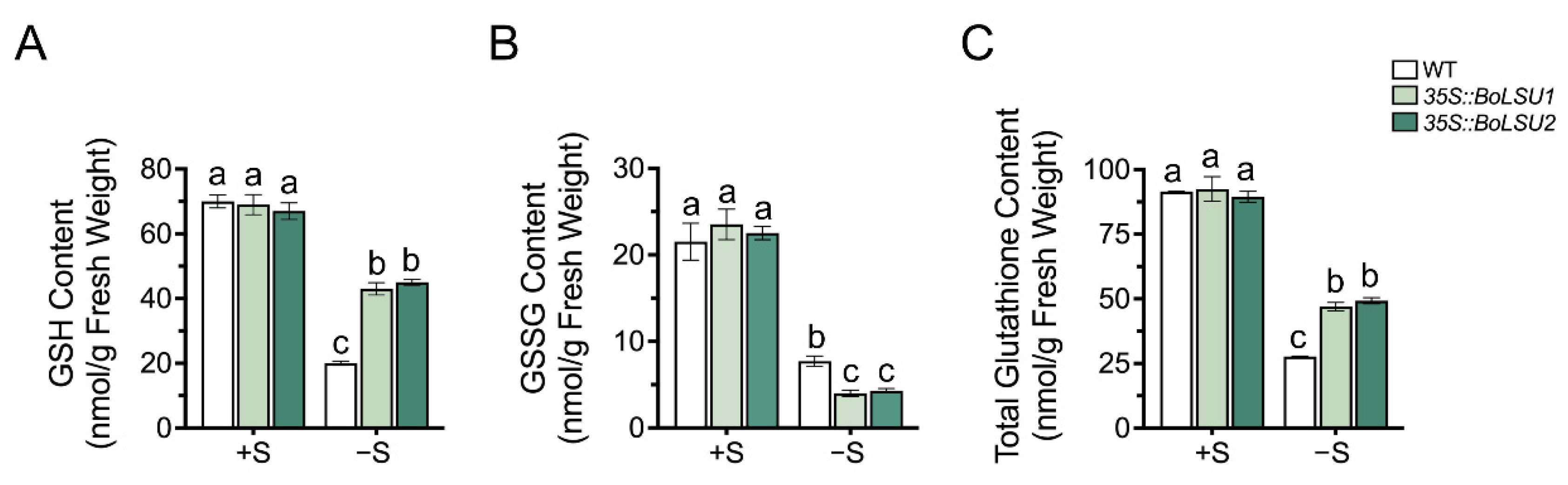
2.6. Overexpression of BoLSU1 and BoLSU2 Increased the Content of Sulfur Metabolite GSH
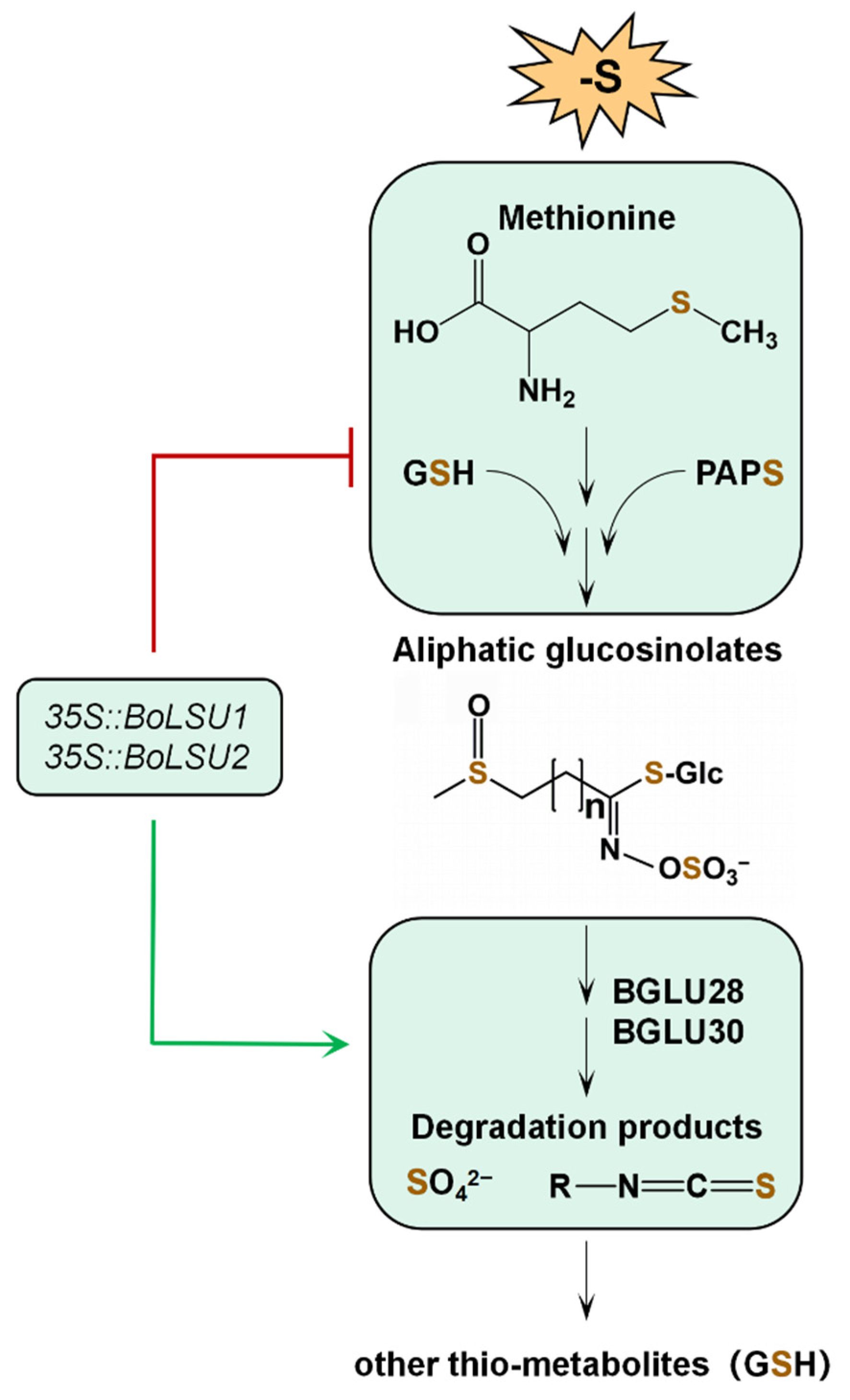
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Culture Conditions
4.2. Sulfur Deficiency Treatment
4.3. Molecular Cloning and Plant Genetic Transformation
4.4. GUS Detection
4.5. Glucosinolate Extraction and Analysis
4.6. Quantitative Real-Time PCR Analyses
4.7. Determination of GSH and GSSG Contents
4.8. Measurement of Chlorophyll Content
4.9. Antioxidant Enzyme Activity Assay
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kopriva, S.; Malagoli, M.; Takahashi, H. Sulfur Nutrition: Impacts on Plant Development, Metabolism, and Stress Responses. J. Exp. Bot. 2019, 70, 4069–4073. [Google Scholar] [CrossRef] [PubMed]
- Saito, K. Sulfur Assimilatory Metabolism. The Long and Smelling Road. Plant Physiol. 2004, 136, 2443–2450. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Gao, Y.; Yang, A. Sulfur Homeostasis in Plants. Int. J. Mol. Sci. 2020, 21, 8926. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Mhmb Bhuyan, J.A.; Mahmud, K.; Nahar, S.M.; Parvin, K.; Fujita, M. Interaction of Sulfur with Phytohormones and Signaling Molecules in Conferring Abiotic Stress Tolerance to Plants. Plant Signal Behav. 2018, 13, e1477905. [Google Scholar] [CrossRef]
- Rausch, T.; Wachter, A. Sulfur Metabolism: A Versatile Platform for Launching Defence Operations. Trends Plant Sci. 2005, 10, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Aarabi, F.; Naake, T.; Fernie, A.R.; Hoefgen, R. Coordinating Sulfur Pools under Sulfate Deprivation. Trends Plant Sci. 2020, 25, 1227–1239. [Google Scholar] [CrossRef] [PubMed]
- Blazevic, I.; Montaut, S.; Burcul, F.; Olsen, C.E.; Burow, M.; Rollin, P.; Agerbirk, N. Glucosinolate Structural Diversity, Identification, Chemical Synthesis and Metabolism in Plants. Phytochemistry 2020, 169, 112100. [Google Scholar] [CrossRef]
- Falk, K.L.; Tokuhisa, J.G.; Gershenzon, J. The Effect of Sulfur Nutrition on Plant Glucosinolate Content: Physiology and Molecular Mechanisms. Plant Biol. 2007, 9, 573–581. [Google Scholar] [CrossRef]
- Xu, Z.W.; Escamilla-Trevino, L.L.; Zeng, L.H.; Lalgondar, M.; Bevan, D.R.; Winkel, B.S.J.; Mohamed, A.; Cheng, C.L.; Shih, M.C.; Poulton, J.E.; et al. Functional Genomic Analysis of Arabidopsis Thaliana Glycoside Hydrolase Family 1. Plant Mol. Biol. 2004, 55, 343–367. [Google Scholar] [CrossRef]
- Mewis, I.; Appel, H.M.; Hom, A.; Raina, R.; Schultz, J.C. Major Signaling Pathways Modulate Arabidopsis Glucosinolate Accumulation and Response to Both Phloem-Feeding and Chewing Insects. Plant Physiol. 2005, 138, 1149–1162. [Google Scholar] [CrossRef]
- Halkier, B.A.; Gershenzon, J. Biology and Biochemistry of Glucosinolates. Annu. Rev. Plant Biol. 2006, 57, 303–333. [Google Scholar] [CrossRef] [PubMed]
- Halkier, B.A. General Introduction to Glucosinolates. In Glucosinolates; Kopriva, S., Ed.; Academic Press: Cambridge, MA, USA, 2016; pp. 1–14. [Google Scholar]
- Sugiyama, R.; Li, R.; Kuwahara, A.; Nakabayashi, R.; Sotta, N.; Mori, T.; Ito, T.; Ohkama-Ohtsu, N.; Fujiwara, T.; Saito, K.; et al. Retrograde Sulfur Flow from Glucosinolates to Cysteine in Arabidopsis Thaliana. Proc. Natl. Acad. Sci. USA 2021, 118, e2017890118. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, R.; Hirai, M.Y. Atypical Myrosinase as a Mediator of Glucosinolate Functions in Plants. Front. Plant Sci. 2019, 10, 1008. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Zhou, Z.; Zhang, T.; Su, H.; Li, J. Overexpression of Lsu1 and Lsu2 Confers Cadmium Tolerance by Manipulating Sulfur Metabolism in Arabidopsis. Chemosphere 2023, 334, 139046. [Google Scholar] [CrossRef] [PubMed]
- Sirko, A.; Wawrzynska, A.; Rodriguez, M.C.; Sektas, P. The Family of Lsu-Like Proteins. Front. Plant Sci. 2015, 5, 774. [Google Scholar] [CrossRef]
- Kong, X.; Li, C.; Zhang, F.; Yu, Q.; Gao, S.; Zhang, M.; Tian, H.; Zhang, J.; Yuan, X.; Ding, Z. Ethylene Promotes Cadmium-Induced Root Growth Inhibition through Ein3 Controlled Xth33 and Lsu1 Expression in Arabidopsis. Plant Cell Environ. 2018, 41, 2449–2462. [Google Scholar] [CrossRef]
- Garcia-Molina, A.; Altmann, M.; Alkofer, A.; Epple, P.M.; Dangl, J.L.; Falter-Braun, P. Lsu Network Hubs Integrate Abiotic and Biotic Stress Responses Via Interaction with the Superoxide Dismutase Fsd2. J. Exp. Bot. 2017, 68, 1185–1197. [Google Scholar] [CrossRef]
- Maruyama-Nakashita, A.; Nakamura, Y.; Watanabe-Takahashi, A.; Inoue, E.; Yamaya, T.; Takahashi, H. Identification of a Novel Cis-Acting Element Conferring Sulfur Deficiency Response in Arabidopsis Roots. Plant J. 2005, 42, 305–314. [Google Scholar] [CrossRef]
- Niemiro, A.; Cysewski, D.; Brzywczy, J.; Wawrzyńska, A.; Sieńko, M.; Poznański, J.; Sirko, A. Similar but Not Identical-Binding Properties of Lsu (Response to Low Sulfur) Proteins from Arabidopsis Thaliana. Front. Plant Sci. 2020, 11, 1246. [Google Scholar] [CrossRef]
- Wang, H.; Seo, J.K.; Gao, S.; Cui, X.; Jin, H. Silencing of Atrap, a Target Gene of a Bacteria-Induced Small Rna, Triggers Antibacterial Defense Responses through Activation of Lsu2 and Down-Regulation of Glk1. New Phytol. 2017, 215, 1144–1155. [Google Scholar] [CrossRef]
- Gao, J.; Yu, X.; Ma, F.; Li, J. Rna-Seq Analysis of Transcriptome and Glucosinolate Metabolism in Seeds and Sprouts of Broccoli (Brassica Oleracea Var. Italic). PLoS ONE 2014, 9, e88804. [Google Scholar] [CrossRef]
- Xiang, C.; Werner, B.L.; Christensen, E.M.; Oliver, D.J. The Biological Functions of Glutathione Revisited in Arabidopsis Transgenic Plants with Altered Glutathione Levels. Plant Physiol. 2001, 126, 564–574. [Google Scholar] [CrossRef] [PubMed]
- Serafin-Andrzejewska, M.; Kozak, M.; Kotecki, A. Effect of Different Sulfur Fertilizer Doses on the Glucosinolate Content and Profile of White Mustard Seeds. J. Elem. 2020, 25, 1413–1422. [Google Scholar] [CrossRef]
- Jiang, J.; Wang, Z.; Kong, X.; Chen, Y.; Li, J. Exogenous Tryptophan Application Improves Cadmium Tolerance and Inhibits Cadmium Upward Transport in Broccoli (Brassica Oleracea Var. Italica). Front. Plant Sci. 2022, 13, 969675. [Google Scholar] [CrossRef]
- Mitreiter, S.; Gigolashvili, T. Regulation of Glucosinolate Biosynthesis. J. Exp. Bot. 2021, 72, 70–91. [Google Scholar] [CrossRef] [PubMed]
- Uribe, F.; Henríquez-Valencia, C.; Arenas, M.A.; Medina, J.; Vidal, E.A.; Canales, J. Evolutionary and Gene Expression Analyses Reveal New Insights into the Role of Lsu Gene-Family in Plant Responses to Sulfate-Deficiency. Plants 2022, 11, 1526. [Google Scholar] [CrossRef]
- Yang, R.Q.; Guo, L.P.; Jin, X.L.; Shen, C.; Zhou, Y.L.; Gu, Z.X. Enhancement of Glucosinolate and Sulforaphane Formation of Broccoli Sprouts by Zinc Sulphate Via Its Stress Effect. J. Funct. Foods 2015, 13, 345–349. [Google Scholar] [CrossRef]
- Kim, Y.C.; Hussain, M.; Anarjan, M.B.; Lee, S. Examination of Glucoraphanin Content in Broccoli Seedlings over Growth and the Impact of Hormones and Sulfur-Containing Compounds. Plant Biotechnol. Rep. 2020, 14, 491–496. [Google Scholar] [CrossRef]
- Yang, Q.X.; Luo, M.L.; Zhou, Q.; Zhao, Y.B.; Chen, J.Y.; Ji, S.J. Insights into the Loss of Glucoraphanin in Post-Harvested Broccoli--Possible Involvement of the Declined Supply Capacity of Sulfur Donor. Plant Sci. 2023, 328, 111580. [Google Scholar] [CrossRef] [PubMed]
- Schonhof, I.; Blankenburg, D.; Muller, S.; Krumbein, A. Sulfur and Nitrogen Supply Influence Growth, Product Appearance, and Glucosinolate Concentration of Broccoli. J. Plant Nutr. Soil Sci. 2007, 170, 65–72. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The Water-Culture Method for Growing Plants without Soil. In California Agricultural Experiment Station Circular; University of Michigan Library: Ann Arbor, MI, USA, 1950. [Google Scholar]
- Nour-Eldin, H.H.; Hansen, B.G.; Nørholm, M.H.; Jensen, J.K.; Halkier, B.A. Advancing Uracil-Excision Based Cloning Towards an Ideal Technique for Cloning Pcr Fragments. Nucleic Acids Res. 2006, 34, e122. [Google Scholar] [CrossRef]
- Zhang, X.; Henriques, R.; Lin, S.S.; Niu, Q.W.; Chua, N.H. Agrobacterium-Mediated Transformation of Arabidopsis Thaliana Using the Floral Dip Method. Nat. Protoc. 2006, 1, 641–646. [Google Scholar] [CrossRef] [PubMed]
- Jefferson, R.A.; Kavanagh, T.A.; Bevan, M.W. Gus Fusions: Beta-Glucuronidase as a Sensitive and Versatile Gene Fusion Marker in Higher Plants. EMBO J. 1987, 6, 3901–3907. [Google Scholar] [CrossRef] [PubMed]
- Hansen, B.G.; Kliebenstein, D.J.; Halkier, B.A. Identification of a Flavin-Monooxygenase as the S-Oxygenating Enzyme in Aliphatic Glucosinolate Biosynthesis in Arabidopsis. Plant J. 2007, 50, 902–910. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative Pcr and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, S.; Zhou, Z.; Zhang, T.; Zhang, Q.; Li, R.; Li, J. Overexpression of BoLSU1 and BoLSU2 Confers Tolerance to Sulfur Deficiency in Arabidopsis by Manipulating Glucosinolate Metabolism. Int. J. Mol. Sci. 2023, 24, 13520. https://doi.org/10.3390/ijms241713520
Yang S, Zhou Z, Zhang T, Zhang Q, Li R, Li J. Overexpression of BoLSU1 and BoLSU2 Confers Tolerance to Sulfur Deficiency in Arabidopsis by Manipulating Glucosinolate Metabolism. International Journal of Molecular Sciences. 2023; 24(17):13520. https://doi.org/10.3390/ijms241713520
Chicago/Turabian StyleYang, Shuang, Zihuan Zhou, Tianqi Zhang, Qianqian Zhang, Rui Li, and Jing Li. 2023. "Overexpression of BoLSU1 and BoLSU2 Confers Tolerance to Sulfur Deficiency in Arabidopsis by Manipulating Glucosinolate Metabolism" International Journal of Molecular Sciences 24, no. 17: 13520. https://doi.org/10.3390/ijms241713520
APA StyleYang, S., Zhou, Z., Zhang, T., Zhang, Q., Li, R., & Li, J. (2023). Overexpression of BoLSU1 and BoLSU2 Confers Tolerance to Sulfur Deficiency in Arabidopsis by Manipulating Glucosinolate Metabolism. International Journal of Molecular Sciences, 24(17), 13520. https://doi.org/10.3390/ijms241713520