
Drosophila Phosphatase of Regenerating Liver Is Critical for Photoreceptor Cell Polarity and Survival during Retinal Development


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. dPRL Is Enriched in the Apical Membrane of Photoreceptor Cells at 50% Pupal Development
2.2. The Prenylation Motif Is Essential for dPRL’s Localization to the Apical Membrane of Photoreceptor Cells
2.3. Knockdown of dPRL Leads to Retinal Degeneration
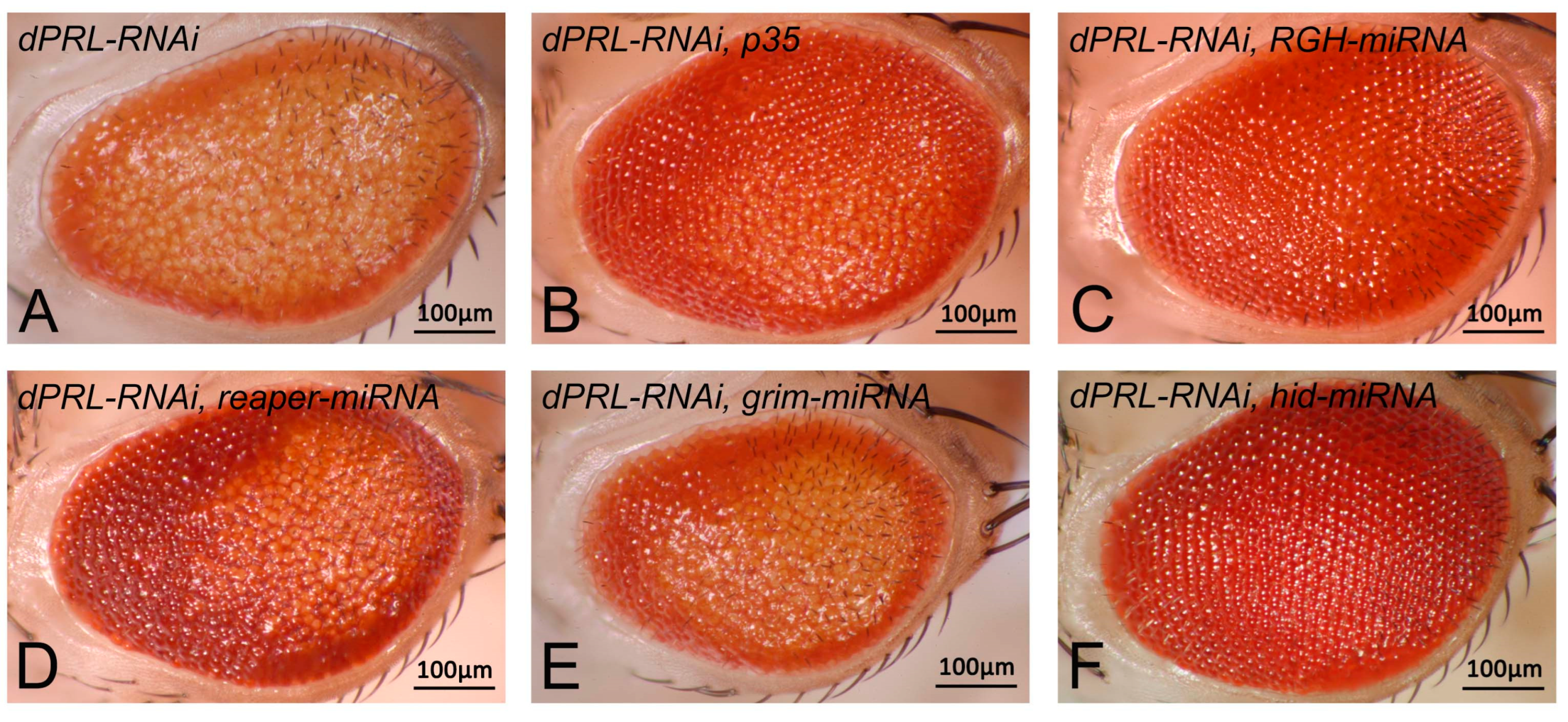
2.4. Knockdown of dPRL Triggers Apoptosis Mediated by the Proapoptotic Gene Hid
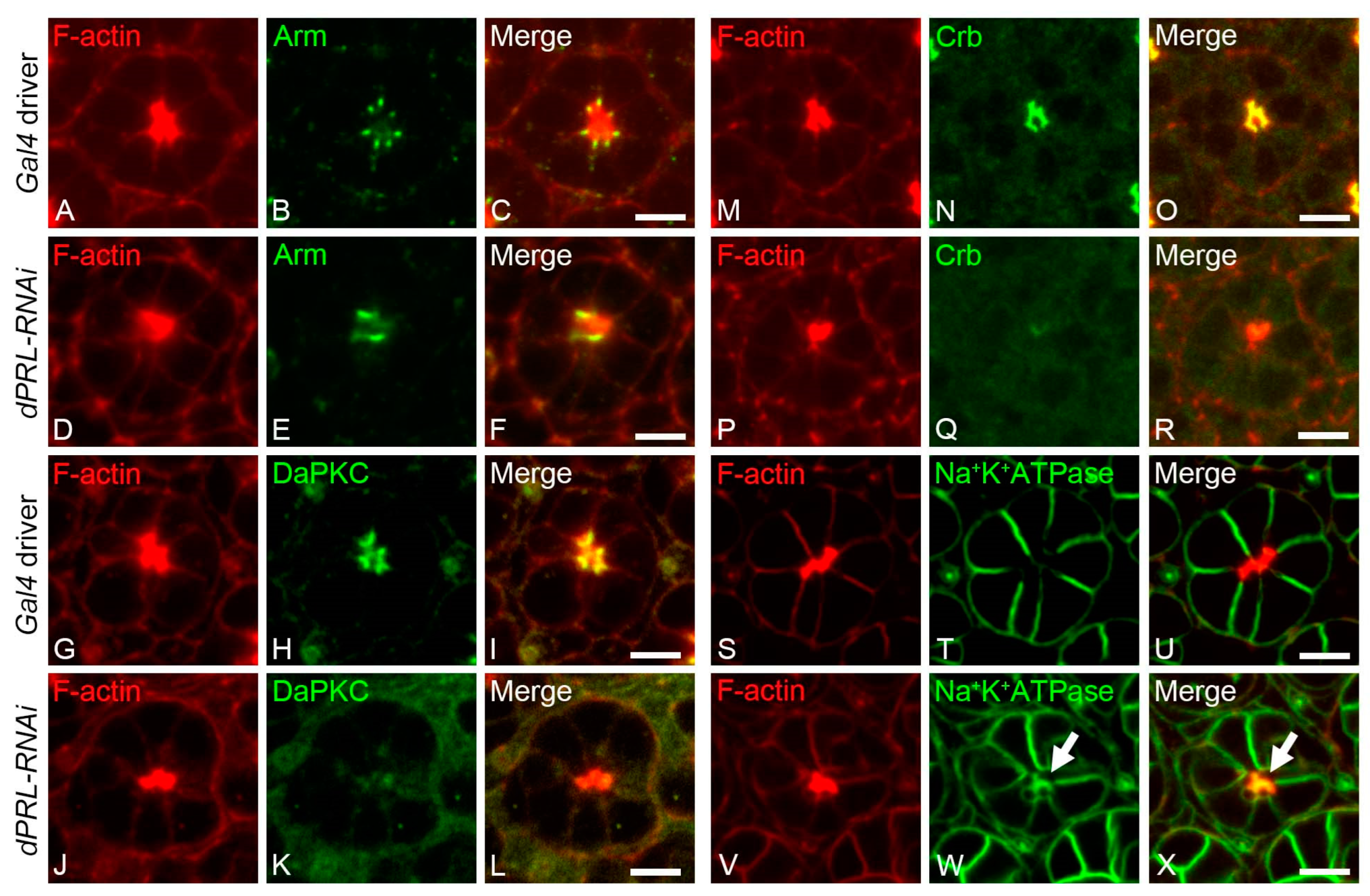
2.5. Polarity of Photoreceptor Cells was Disrupted in the Pupal Retina of dPRL-Knockdowned Flies
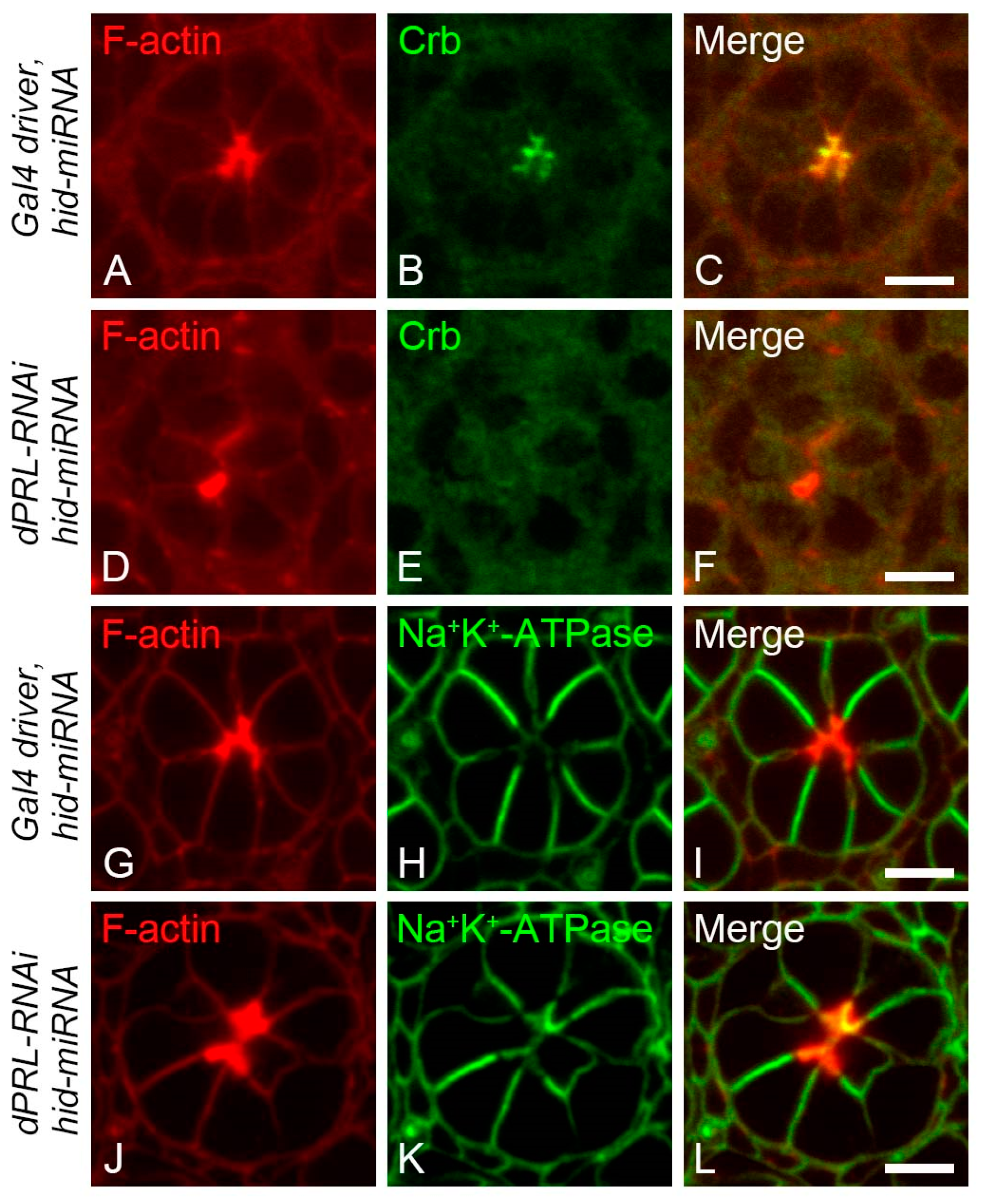
2.6. Polarity Disruption of Photoreceptor Cells in dPRL-Depleted Retina was Not a Secondary Effect of Apoptosis
3. Discussion
4. Materials and Methods
4.1. Drosophila Stocks
4.2. Transmission Electron Microscopy (TEM) for Adult Retina
4.3. Drosophila Whole-Mount Immunostaining of Pupal Retina
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Longley, R.L., Jr.; Ready, D.F. Integrins and the development of three-dimensional structure in the Drosophila compound eye. Dev. Biol. 1995, 171, 415–433. [Google Scholar] [CrossRef]
- Flores-Benitez, D.; Knust, E. Dynamics of epithelial cell polarity in Drosophila: How to regulate the regulators? Curr. Opin. Cell Biol. 2016, 42, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Izaddoost, S.; Nam, S.C.; Bhat, M.A.; Bellen, H.J.; Choi, K.W. Drosophila Crumbs is a positional cue in photoreceptor adherens junctions and rhabdomeres. Nature 2002, 416, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Pellikka, M.; Tanentzapf, G.; Pinto, M.; Smith, C.; McGlade, C.J.; Ready, D.F.; Tepass, U. Crumbs, the Drosophila homologue of human CRB1/RP12, is essential for photoreceptor morphogenesis. Nature 2002, 416, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Atwood, S.X.; Chabu, C.; Penkert, R.R.; Doe, C.Q.; Prehoda, K.E. Cdc42 acts downstream of Bazooka to regulate neuroblast polarity through Par-6 aPKC. J. Cell Sci. 2007, 120 Pt 18, 3200–3206. [Google Scholar] [CrossRef]
- Walther, R.F.; Pichaud, F. Crumbs/DaPKC-dependent apical exclusion of Bazooka promotes photoreceptor polarity remodeling. Curr. Biol. 2010, 20, 1065–1074. [Google Scholar] [CrossRef]
- Harris, T.J.; Peifer, M. Adherens junction-dependent and -independent steps in the establishment of epithelial cell polarity in Drosophila. J. Cell Biol. 2004, 167, 135–147. [Google Scholar] [CrossRef]
- McGill, M.A.; McKinley, R.F.; Harris, T.J. Independent cadherin-catenin and Bazooka clusters interact to assemble adherens junctions. J. Cell Biol. 2009, 185, 787–796. [Google Scholar] [CrossRef]
- Kozlov, G.; Cheng, J.; Ziomek, E.; Banville, D.; Gehring, K.; Ekiel, I. Structural insights into molecular function of the metastasis-associated phosphatase PRL-3. J. Biol. Chem. 2004, 279, 11882–11889. [Google Scholar] [CrossRef]
- Jeong, D.G.; Kim, S.J.; Kim, J.H.; Son, J.H.; Park, M.R.; Lim, S.M.; Yoon, T.S.; Ryu, S.E. Trimeric structure of PRL-1 phosphatase reveals an active enzyme conformation and regulation mechanisms. J. Mol. Biol. 2005, 345, 401–413. [Google Scholar] [CrossRef]
- Lin, M.D.; Lee, H.T.; Wang, S.C.; Li, H.R.; Hsien, H.L.; Cheng, K.W.; Chang, Y.D.; Huang, M.L.; Yu, J.K.; Chen, Y.H. Expression of phosphatase of regenerating liver family genes during embryogenesis: An evolutionary developmental analysis among Drosophila, amphioxus, and zebrafish. BMC Dev. Biol. 2013, 13, 18. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.; Si, X.; Horstmann, H.; Xu, Y.; Hong, W.; Pallen, C.J. Prenylation-dependent association of protein-tyrosine phosphatases PRL-1, -2, and -3 with the plasma membrane and the early endosome. J. Biol. Chem. 2000, 275, 21444–21452. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Kirby, C.E.; Herbst, R. The tyrosine phosphatase PRL-1 localizes to the endoplasmic reticulum and the mitotic spindle and is required for normal mitosis. J. Biol. Chem. 2002, 277, 46659–46668. [Google Scholar] [CrossRef]
- Diamond, R.H.; Cressman, D.E.; Laz, T.M.; Abrams, C.S.; Taub, R. PRL-1, a unique nuclear protein tyrosine phosphatase, affects cell growth. Mol. Cell. Biol. 1994, 14, 3752–3762. [Google Scholar] [CrossRef] [PubMed]
- Mohn, K.L.; Laz, T.M.; Hsu, J.C.; Melby, A.E.; Bravo, R.; Taub, R. The immediate-early growth response in regenerating liver and insulin-stimulated H-35 cells: Comparison with serum-stimulated 3T3 cells and identification of 41 novel immediate-early genes. Mol. Cell. Biol. 1991, 11, 381–390. [Google Scholar] [CrossRef]
- Bessette, D.C.; Qiu, D.; Pallen, C.J. PRL PTPs: Mediators and markers of cancer progression. Cancer Metastasis Rev. 2008, 27, 231–252. [Google Scholar] [CrossRef] [PubMed]
- Funato, Y.; Hashizume, O.; Miki, H. Phosphatase-independent role of phosphatase of regenerating liver in cancer progression. Cancer Sci. 2023, 114, 25–33. [Google Scholar] [CrossRef]
- Rubio, T.; Kohn, M. Regulatory mechanisms of phosphatase of regenerating liver (PRL)-3. Biochem. Soc. Trans. 2016, 44, 1305–1312. [Google Scholar] [CrossRef][Green Version]
- Jiao, Y.; Ye, D.Z.; Li, Z.; Teta-Bissett, M.; Peng, Y.; Taub, R.; Greenbaum, L.E.; Kaestner, K.H. Protein tyrosine phosphatase of liver regeneration-1 is required for normal timing of cell cycle progression during liver regeneration. Am. J. Physiol. Gastrointest Liver Physiol. 2015, 308, G85–G91. [Google Scholar] [CrossRef]
- Yan, H.; Kong, D.; Ge, X.; Gao, X.; Han, X. Generation of conditional knockout alleles for PRL-3. J. Biomed. Res. 2011, 25, 438–443. [Google Scholar] [CrossRef]
- Zimmerman, M.W.; Homanics, G.E.; Lazo, J.S. Targeted deletion of the metastasis-associated phosphatase Ptp4a3 (PRL-3) suppresses murine colon cancer. PLoS ONE 2013, 8, e58300. [Google Scholar] [CrossRef]
- Dong, Y.; Zhang, L.; Bai, Y.; Zhou, H.M.; Campbell, A.M.; Chen, H.; Yong, W.; Zhang, W.; Zeng, Q.; Shou, W.; et al. Phosphatase of regenerating liver 2 (PRL2) deficiency impairs Kit signaling and spermatogenesis. J. Biol. Chem. 2014, 289, 3799–3810. [Google Scholar] [CrossRef]
- Dong, Y.; Zhang, L.; Zhang, S.; Bai, Y.; Chen, H.; Sun, X.; Yong, W.; Li, W.; Colvin, S.C.; Rhodes, S.J.; et al. Phosphatase of regenerating liver 2 (PRL2) is essential for placental development by down-regulating PTEN (Phosphatase and Tensin Homologue Deleted on Chromosome 10) and activating Akt protein. J. Biol. Chem. 2012, 287, 32172–32179. [Google Scholar] [CrossRef] [PubMed]
- Maacha, S.; Planque, N.; Laurent, C.; Pegoraro, C.; Anezo, O.; Maczkowiak, F.; Monsoro-Burq, A.H.; Saule, S. Protein tyrosine phosphatase 4A3 (PTP4A3) is required for Xenopus laevis cranial neural crest migration in vivo. PLoS ONE 2013, 8, e84717. [Google Scholar] [CrossRef] [PubMed]
- Deichmann, C.; Link, M.; Seyfang, M.; Knotz, V.; Gradl, D.; Wedlich, D. Neural crest specification by Prohibitin1 depends on transcriptional regulation of prl3 and vangl1. Genesis 2015, 53, 627–639. [Google Scholar] [CrossRef] [PubMed]
- Pagarigan, K.T.; Bunn, B.W.; Goodchild, J.; Rahe, T.K.; Weis, J.F.; Saucedo, L.J. Drosophila PRL-1 is a growth inhibitor that counteracts the function of the Src oncogene. PLoS ONE 2013, 8, e61084. [Google Scholar] [CrossRef] [PubMed]
- Phelps, C.B.; Brand, A.H. Ectopic gene expression in Drosophila using GAL4 system. Methods 1998, 14, 367–379. [Google Scholar] [CrossRef]
- Cates, C.A.; Michael, R.L.; Stayrook, K.R.; Harvey, K.A.; Burke, Y.D.; Randall, S.K.; Crowell, P.L.; Crowell, D.N. Prenylation of oncogenic human PTP(CAAX) protein tyrosine phosphatases. Cancer Lett. 1996, 110, 49–55. [Google Scholar] [CrossRef]
- Sun, J.P.; Luo, Y.; Yu, X.; Wang, W.Q.; Zhou, B.; Liang, F.; Zhang, Z.Y. Phosphatase activity, trimerization, and the C-terminal polybasic region are all required for PRL1-mediated cell growth and migration. J. Biol. Chem. 2007, 282, 29043–29051. [Google Scholar] [CrossRef]
- Dietzl, G.; Chen, D.; Schnorrer, F.; Su, K.C.; Barinova, Y.; Fellner, M.; Gasser, B.; Kinsey, K.; Oppel, S.; Scheiblauer, S.; et al. A genome-wide transgenic RNAi library for conditional gene inactivation in Drosophila. Nature 2007, 448, 151–156. [Google Scholar] [CrossRef]
- Chen, C.H.; Huang, H.; Ward, C.M.; Su, J.T.; Schaeffer, L.V.; Guo, M.; Hay, B.A. A synthetic maternal-effect selfish genetic element drives population replacement in Drosophila. Science 2007, 316, 597–600. [Google Scholar] [CrossRef] [PubMed]
- Yasuhara, J.C.; Baumann, O.; Takeyasu, K. Localization of Na/K-ATPase in developing and adult Drosophila melanogaster photoreceptors. Cell Tissue Res. 2000, 300, 239–249. [Google Scholar] [CrossRef] [PubMed]
- Lujan, P.; Varsano, G.; Rubio, T.; Hennrich, M.L.; Sachsenheimer, T.; Galvez-Santisteban, M.; Martin-Belmonte, F.; Gavin, A.C.; Brugger, B.; Kohn, M. PRL-3 disrupts epithelial architecture by altering the post-mitotic midbody position. J. Cell Sci. 2016, 129, 4130–4142. [Google Scholar] [CrossRef] [PubMed]
- Colosimo, P.F.; Liu, X.; Kaplan, N.A.; Tolwinski, N.S. GSK3beta affects apical-basal polarity and cell-cell adhesion by regulating aPKC levels. Dev. Dyn. 2010, 239, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, N.A.; Colosimo, P.F.; Liu, X.; Tolwinski, N.S. Complex interactions between GSK3 and aPKC in Drosophila embryonic epithelial morphogenesis. PLoS ONE 2011, 6, e18616. [Google Scholar] [CrossRef]
- Frame, S.; Cohen, P.; Biondi, R.M. A common phosphate binding site explains the unique substrate specificity of GSK3 and its inactivation by phosphorylation. Mol. Cell 2001, 7, 1321–1327. [Google Scholar] [CrossRef]
- Wang, H.; Quah, S.Y.; Dong, J.M.; Manser, E.; Tang, J.P.; Zeng, Q. PRL-3 down-regulates PTEN expression and signals through PI3K to promote epithelial-mesenchymal transition. Cancer Res. 2007, 67, 2922–2926. [Google Scholar] [CrossRef]
- Chong PS, Y.; Zhou, J.; Lim JS, L.; Hee, Y.T.; Chooi, J.Y.; Chung, T.H.; Tan, Z.T.; Zeng, Q.; Waller, D.D.; Sebag, M.; et al. IL6 Promotes a STAT3-PRL3 Feedforward Loop via SHP2 Repression in Multiple Myeloma. Cancer Res. 2019, 79, 4679–4688. [Google Scholar] [CrossRef]
- Lazo, J.S.; Isbell, K.N.; Vasa, S.A.; Llaneza, D.C.; Rastelli, E.J.; Wipf, P.; Sharlow, E.R. Disruption of Ovarian Cancer STAT3 and p38 Signaling with a Small-Molecule Inhibitor of PTP4A3 Phosphatase. J. Pharmacol. Exp. Ther. 2023, 384, 429–438. [Google Scholar] [CrossRef]
- Zhang, S.Q.; Tsiaras, W.G.; Araki, T.; Wen, G.; Minichiello, L.; Klein, R.; Neel, B.G. Receptor-specific regulation of phosphatidylinositol 3′-kinase activation by the protein tyrosine phosphatase Shp2. Mol. Cell. Biol. 2002, 22, 4062–4072. [Google Scholar] [CrossRef]
- Liu, Y.; Zhou, J.; Chen, J.; Gao, W.; Le, Y.; Ding, Y.; Li, J. PRL-3 promotes epithelial mesenchymal transition by regulating cadherin directly. Cancer Biol. Ther. 2009, 8, 1352–1359. [Google Scholar] [CrossRef] [PubMed]
- Brodsky, M.H.; Weinert, B.T.; Tsang, G.; Rong, Y.S.; McGinnis, N.M.; Golic, K.G.; Rio, D.C.; Rubin, G.M. Drosophila melanogaster MNK/Chk2 and p53 regulate multiple DNA repair and apoptotic pathways following DNA damage. Mol. Cell. Biol. 2004, 24, 1219–1231. [Google Scholar] [CrossRef]
- Moon, N.S.; Di Stefano, L.; Morris, E.J.; Patel, R.; White, K.; Dyson, N.J. E2F and p53 induce apoptosis independently during Drosophila development but intersect in the context of DNA damage. PLoS Genet. 2008, 4, e1000153. [Google Scholar] [CrossRef] [PubMed]
- Basak, S.; Jacobs, S.B.; Krieg, A.J.; Pathak, N.; Zeng, Q.; Kaldis, P.; Giaccia, A.J.; Attardi, L.D. The metastasis-associated gene Prl-3 is a p53 target involved in cell-cycle regulation. Mol. Cell 2008, 30, 303–314. [Google Scholar] [CrossRef] [PubMed]
- Brumby, A.M.; Richardson, H.E. scribble mutants cooperate with oncogenic Ras or Notch to cause neoplastic overgrowth in Drosophila. EMBO J. 2003, 22, 5769–5779. [Google Scholar] [CrossRef] [PubMed]
- Igaki, T.; Pagliarini, R.A.; Xu, T. Loss of cell polarity drives tumor growth and invasion through JNK activation in Drosophila. Curr. Biol. 2006, 16, 1139–1146. [Google Scholar] [CrossRef]
- Uhlirova, M.; Jasper, H.; Bohmann, D. Non-cell-autonomous induction of tissue overgrowth by JNK/Ras cooperation in a Drosophila tumor model. Proc. Natl. Acad. Sci. USA 2005, 102, 13123–13128. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, S.-F.; Hsien, H.-L.; Wang, T.-F.; Lin, M.-D. Drosophila Phosphatase of Regenerating Liver Is Critical for Photoreceptor Cell Polarity and Survival during Retinal Development. Int. J. Mol. Sci. 2023, 24, 11501. https://doi.org/10.3390/ijms241411501
Chen S-F, Hsien H-L, Wang T-F, Lin M-D. Drosophila Phosphatase of Regenerating Liver Is Critical for Photoreceptor Cell Polarity and Survival during Retinal Development. International Journal of Molecular Sciences. 2023; 24(14):11501. https://doi.org/10.3390/ijms241411501
Chicago/Turabian StyleChen, Shu-Fen, Hsin-Lun Hsien, Ting-Fang Wang, and Ming-Der Lin. 2023. "Drosophila Phosphatase of Regenerating Liver Is Critical for Photoreceptor Cell Polarity and Survival during Retinal Development" International Journal of Molecular Sciences 24, no. 14: 11501. https://doi.org/10.3390/ijms241411501
APA StyleChen, S.-F., Hsien, H.-L., Wang, T.-F., & Lin, M.-D. (2023). Drosophila Phosphatase of Regenerating Liver Is Critical for Photoreceptor Cell Polarity and Survival during Retinal Development. International Journal of Molecular Sciences, 24(14), 11501. https://doi.org/10.3390/ijms241411501