
Molecules Inducing Dental Stem Cells Differentiation and Bone Regeneration: State of the Art



Abstract
1. Introduction
1.1. Dental Stem Cells (DSCs)
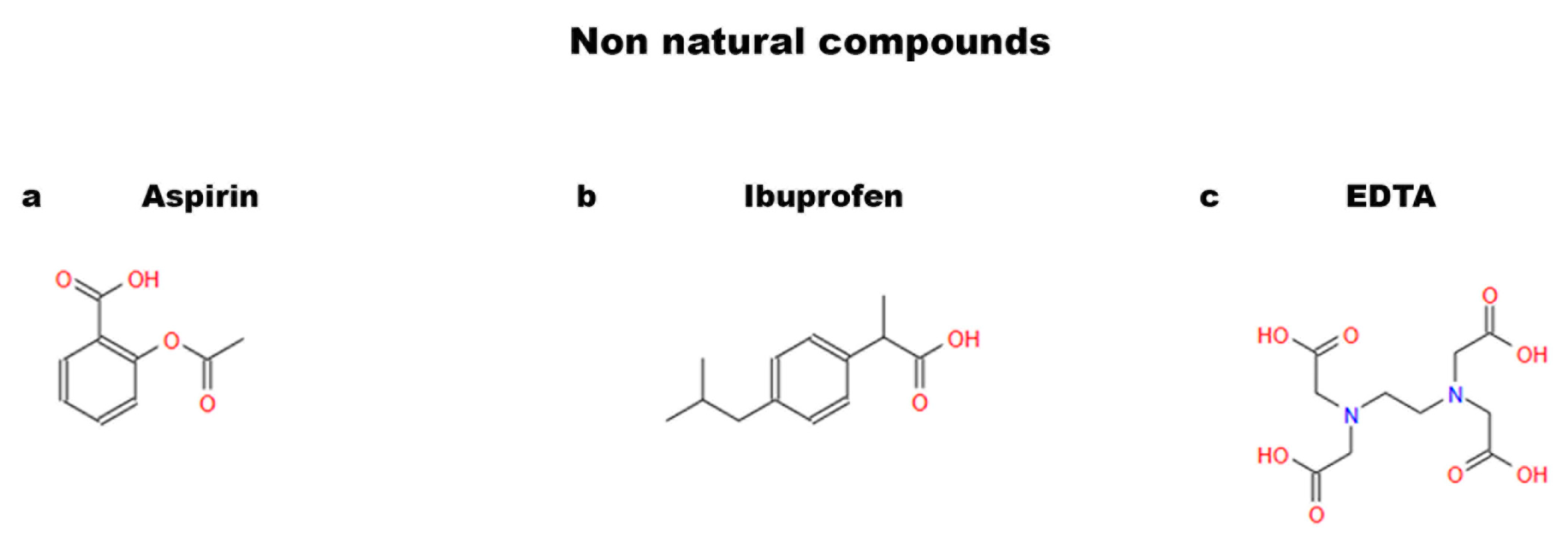
1.2. Natural and Non-Natural Compounds

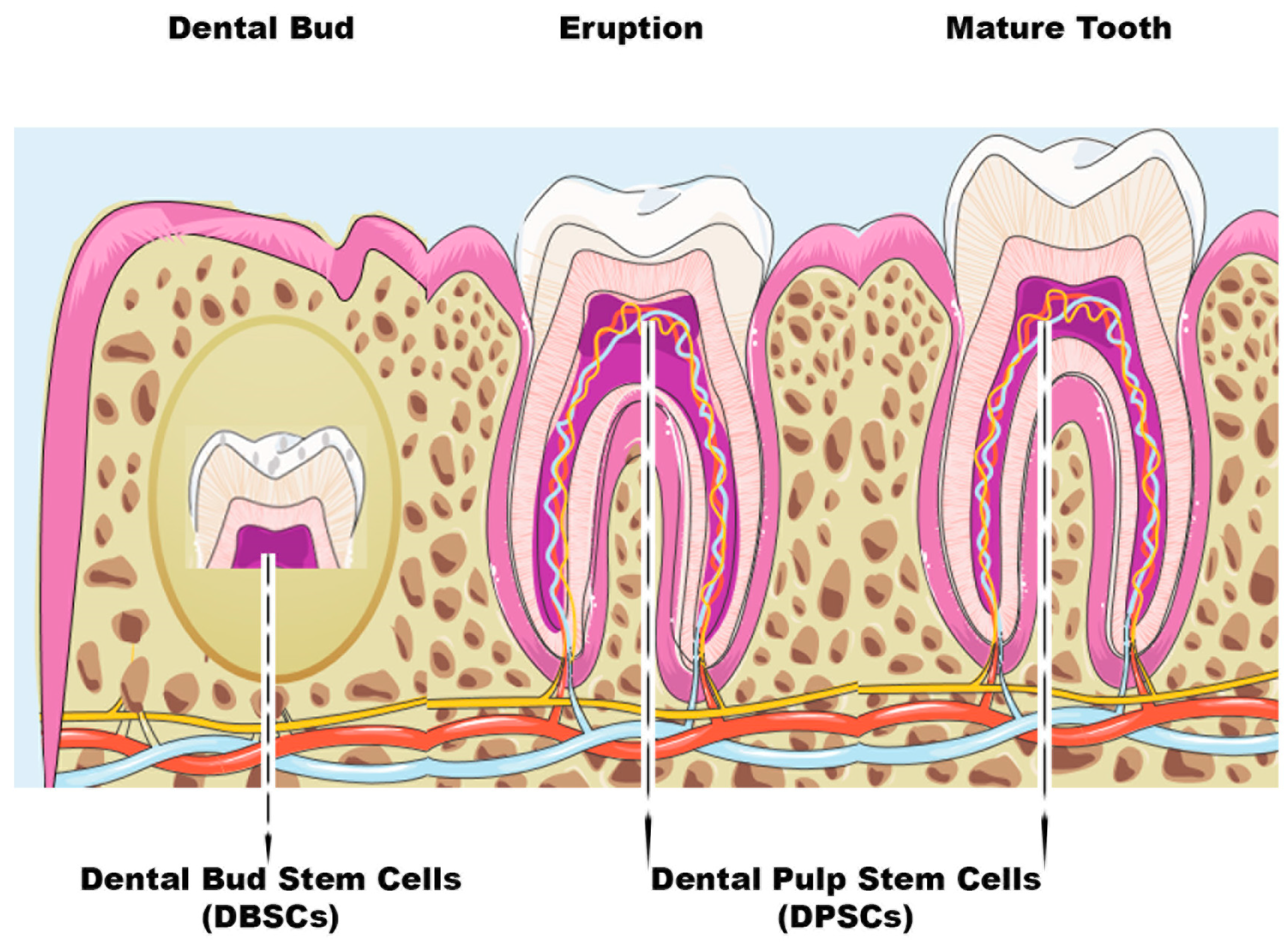{kind=link}
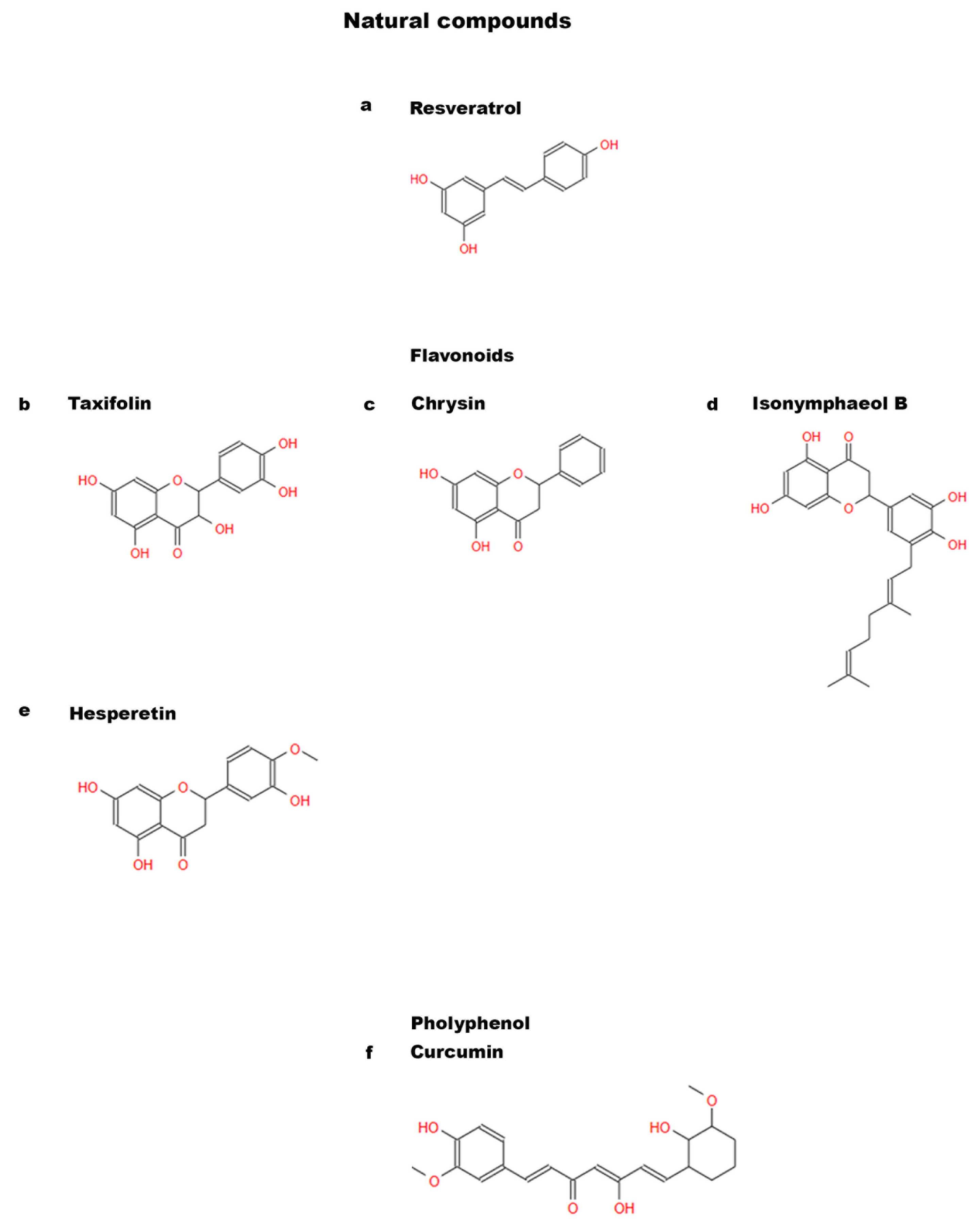{kind=link}
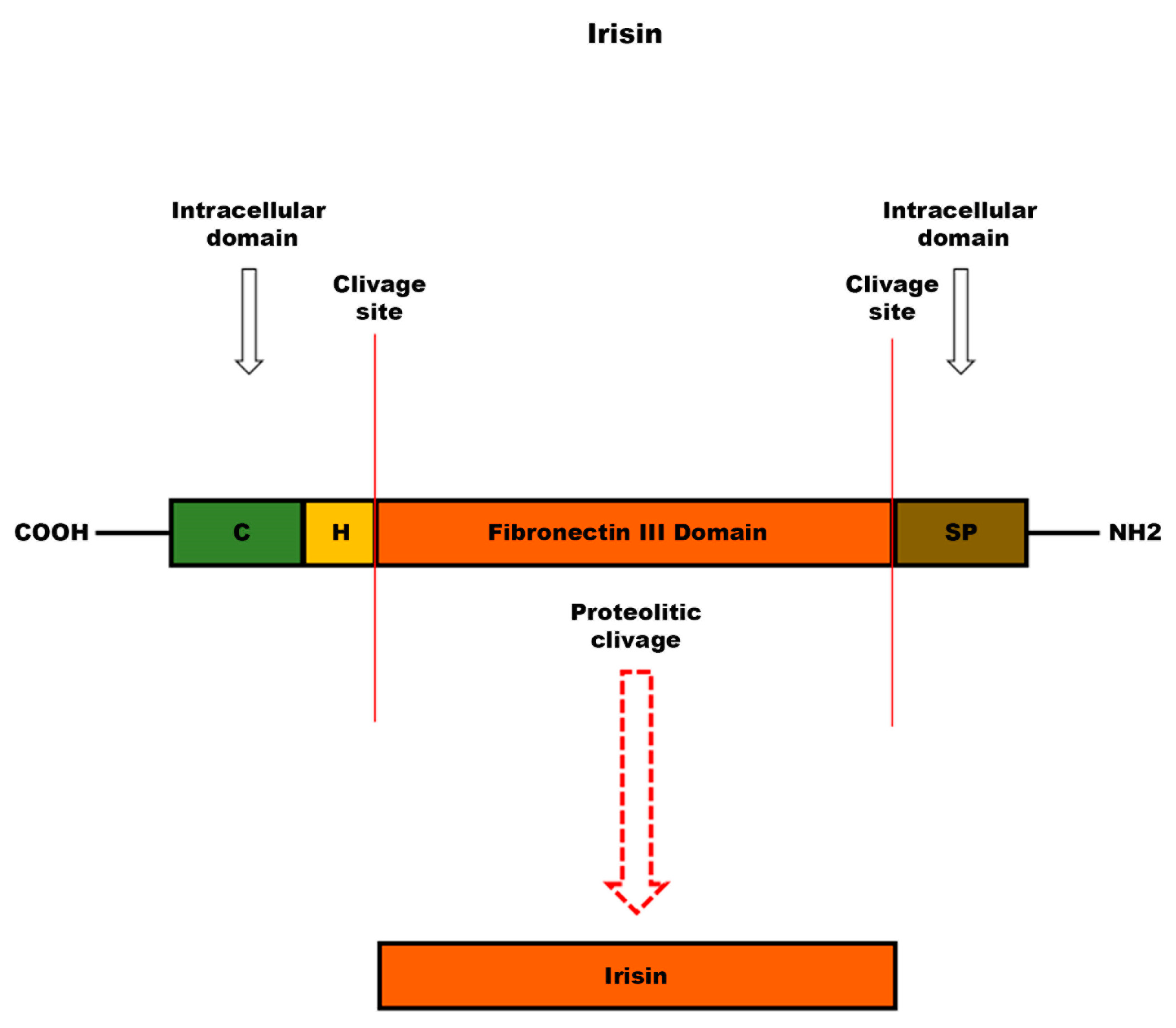{kind=link}
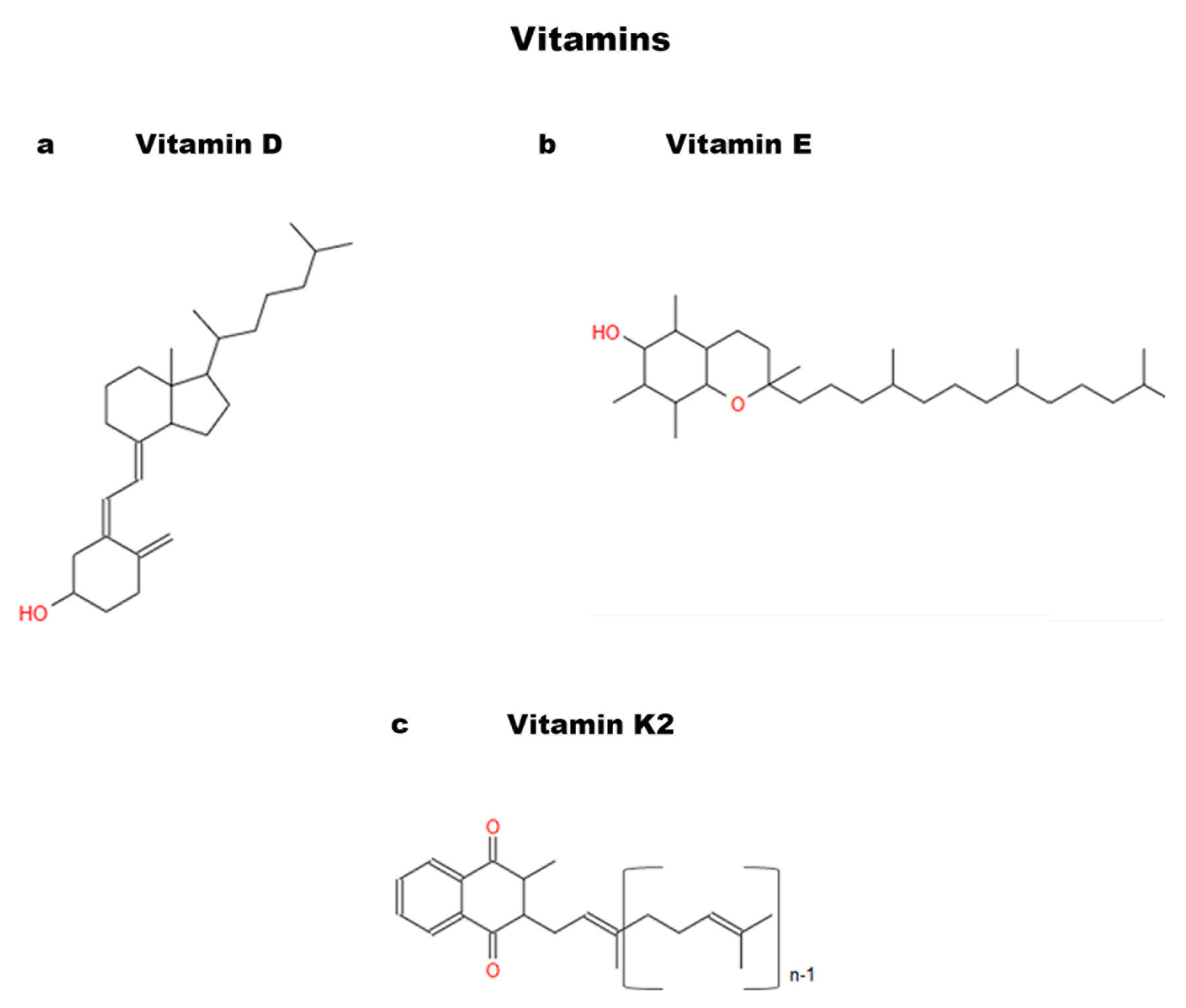{kind=link}
{kind=link}
{kind=link}
{kind=link}
 increase,
increase,  decrease,
decrease,  absence).
increase, decrease, absence).
absence).
increase, decrease, absence).| DPSCs | DBSCs | ||||
|---|---|---|---|---|---|
| Study | Natural Compounds | Effects | Study | Natural Compounds | Effects |
| Zhang et al., 2022 [14] Feng et al., 2016 [15] | Resveratrol | osteogenic differentiation antioxRunx-2, BMP2, Col I, OCN, ALP | Di Benedetto et al., 2018 [16] | Polydatin | osteogenic differentiation mineral matrix depositionATF-4, OPN, ALP |
| Flavonoids: | Di Benedetto et al., 2018 [16] | Resveratrol | osteogenic differentiationOPN, ALP | ||
| Fu et al., 2021 [17] | Taxifolin | apoptosisproliferation | |||
| Huo et al., 2021 [18] | Chrysin | osteogenic differentiationproliferationRunx-2, Col I, OCN | Posa et al., 2016 [19] Posa et al., 2018 [20] | Vitamin D | osteogenic differentiationRunx-2, Col Imineral matrix deposition αVβ3 integrin expression |
| Prenylflavonoids | |||||
| Nam et al., 2021 [21] | Isonymphaeol B (INB) | mineral matrix depositionodontogenic markers | |||
| Alipour et al., 2023 [22] | Hesperetin 0.5 and 1 μM | proliferationcytotoxic effectCol I, BSP, Runx-2, OCN, ALPmineral matrix depositioninflammation | |||
| Curcumin | |||||
| Samiei et al., 2021 [23] | 0.5–1 μM | proliferationcytotoxic effectALP | |||
| Samiei et al., 2022 [24] | 25 μM | proliferation | |||
| Son et al., 2021 [25] | Irisin | proliferationodontogenic differentiationodontogenic markersALPmineralized nodule formation | |||
| Vitamins: | |||||
| Escobar et al., 2020 [26] | Vit D/ Vit E | osteogenic differentiationRunx-2, OSXproliferation | |||
| Rasouli-Ghahroudi et al., 2017 [27] | Vitamin K2 (MK-4) | osteogenic differentiationmineral matrix deposition | |||
| Khanna-Jain et al., 2012 [28] | Vitamin D3 | osteogenic differentiationOCN | |||
| Study | Non-Natural Compounds | Effects | Study | Non-Natural Compounds | Effects |
| Aspirin | Di Benedetto et al., 2020 [29] | T-LysYal | osteogenic differentiation Runx-2, Col I αVβ3 integrin expression | ||
| Khampatee et al., 2022 [30] | 25–50 μg/mL | odontogenesisproliferation | |||
| Salkın et al., 2023 [13] | Ibuprofen | proliferation | |||
| Pang et al, 2014 [31] Galler et al., 2011 [32] Zand et al.,2023 [33] | EDTA | osteogenic differentiationmineral matrix depositionproliferationDSPP, ALP, OCN, DMP-1 | |||
2. Dental Pulp Stem Cells (DPSCs)
3. Differentiative Role of Natural Molecules in DPSCs
3.1. Resveratrol
3.2. Flavonoids
3.2.1. Taxifolin
3.2.2. Chrysin
3.2.3. Prenylflavonoids
3.2.4. Hesperetin
3.3. Curcumin
3.4. Irisin
3.5. Vitamins
4. Non-Natural Compounds
4.1. Aspirin
4.2. Ibuprofen
4.3. EDTA
5. Dental Bud Stem Cells (DBSCs)
6. Differentiative Role of Natural Molecules in DBSCs
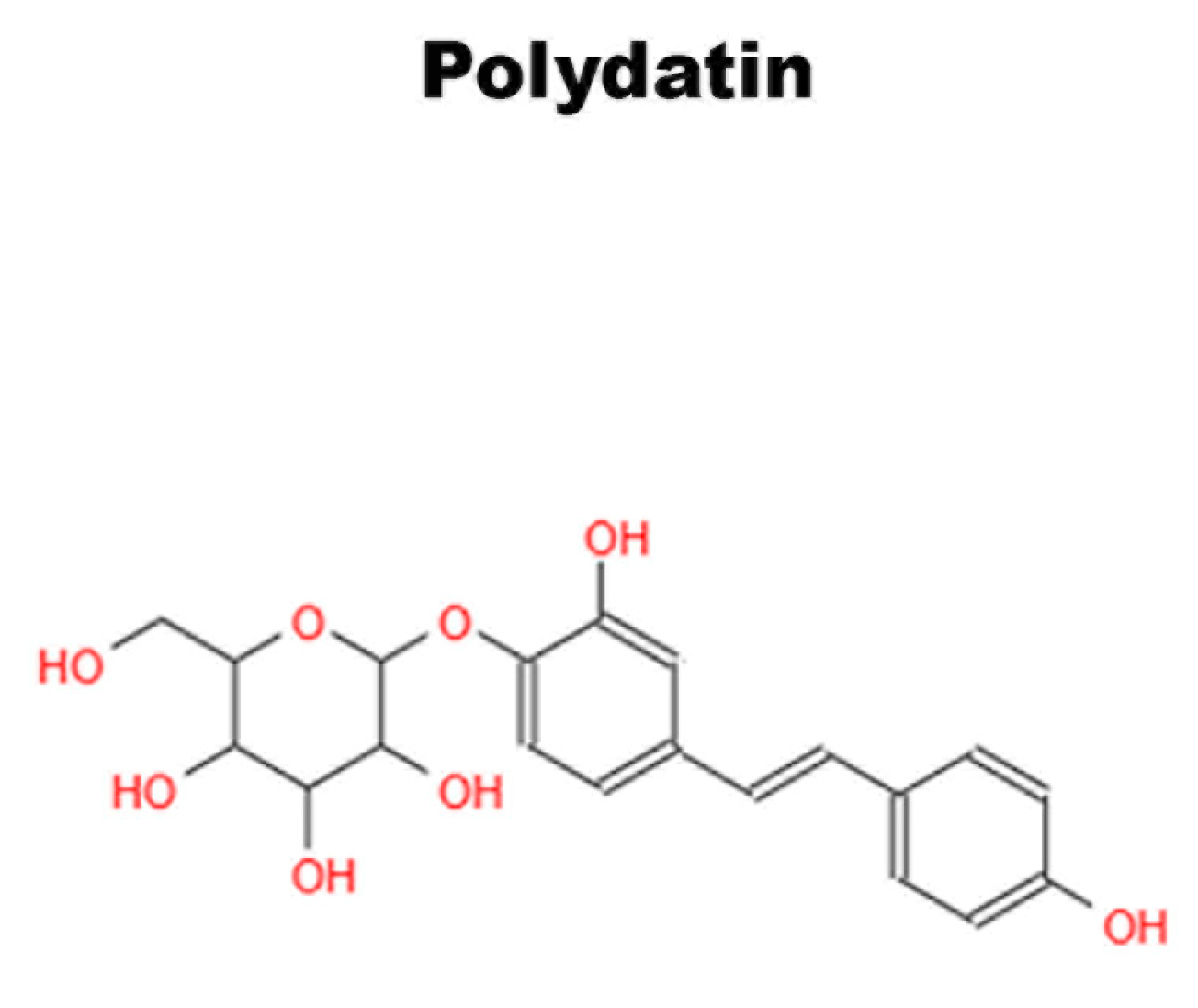
6.1. Polydatin
6.2. Vitamin D
6.3. Irisin
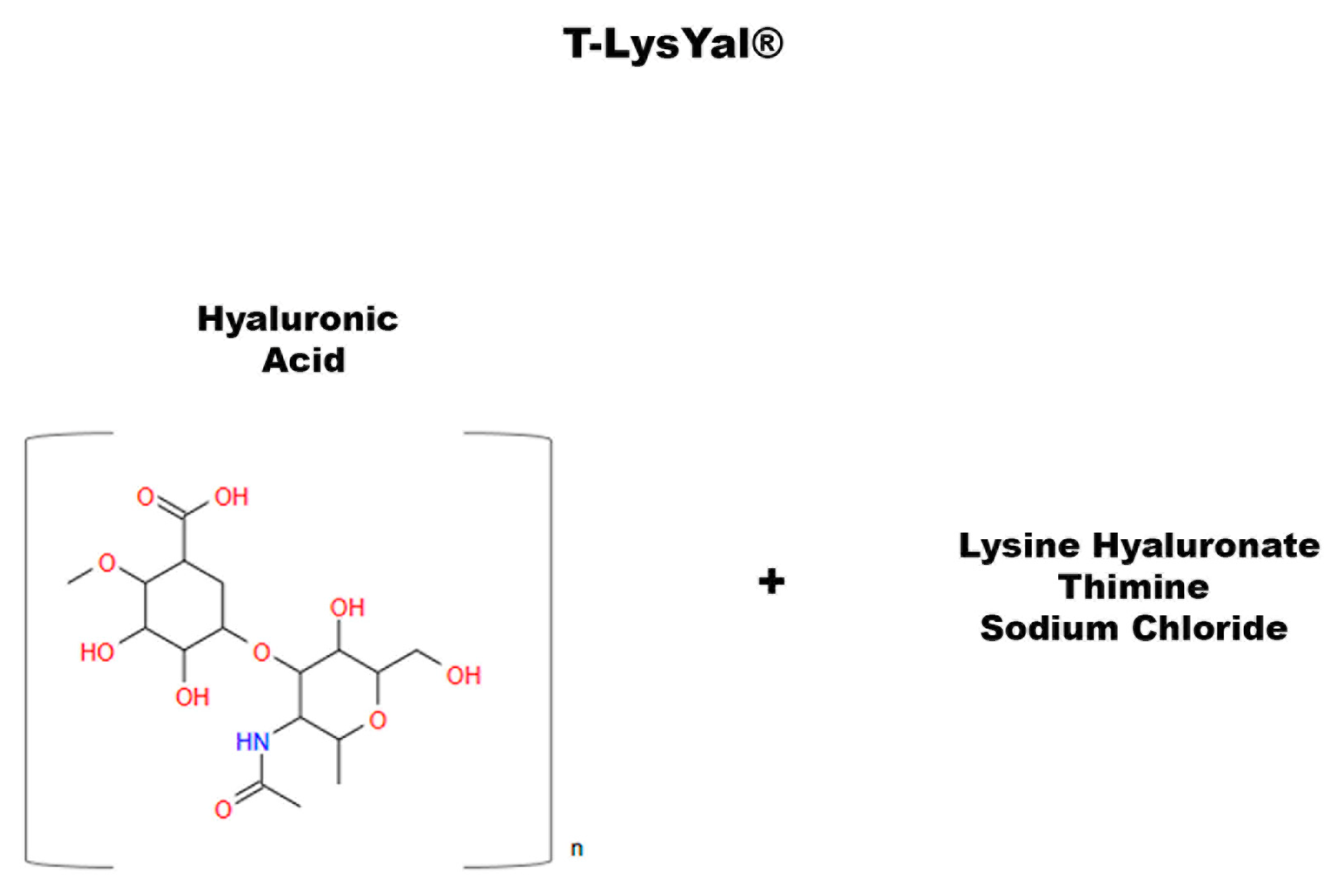
7. T-LysYal: A Non-Natural Compound That Promotes Osteogenic Differentiation
8. Perspectives and Limitations
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Uccelli, A.; Moretta, L.; Pistoia, V. Mesenchymal stem cells in health and disease. Nat. Rev. Immunol. 2008, 8, 726–736. [Google Scholar] [CrossRef]
- Rodgers, K.; Jadhav, S.S. The application of mesenchymal stem cells to treat thermal and radiation burns. Adv. Drug Deliv. Rev. 2018, 123, 75–81. [Google Scholar] [CrossRef]
- Rodeo, S.A. Cell therapy in orthopaedics: Where are we in 2019? Bone Jt. J. 2019, 101, 361–364. [Google Scholar] [CrossRef]
- Kebriaei, P.; Isola, L.; Bahceci, E.; Holland, K.; Rowley, S.; McGuirk, J.; Devetten, M.; Jansen, J.; Herzig, R.; Schuster, M.; et al. Adult human mesenchymal stem cells added to corticosteroid therapy for the treatment of acute graft-versus-host disease. Biol. Blood Marrow Transplant. 2009, 15, 804–811. [Google Scholar] [CrossRef]
- Madonna, R.; Geng, Y.J.; De Caterina, R. Adipose tissue-derived stem cells: Characterization and potential for cardiovascular repair. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 1723–1729. [Google Scholar] [CrossRef]
- Řehořová, M.; Vargová, I.; Forostyak, S.; Vacková, I.; Turnovcová, K.; Kupcová Skalníková, H.; Vodička, P.; Kubinová, Š.; Syková, E.; Jendelová, P. A Combination of Intrathecal and Intramuscular Application of Human Mesenchymal Stem Cells Partly Reduces the Activation of Necroptosis in the Spinal Cord of SOD1(G93A) Rats. Stem Cells Transl. Med. 2019, 8, 535–547. [Google Scholar] [CrossRef]
- Gronthos, S.; Mankani, M.; Brahim, J.; Robey, P.G.; Shi, S. Postnatal human dental pulp stem cells (DPSCs) in vitro and in vivo. Proc. Natl. Acad. Sci. USA 2000, 97, 13625–13630. [Google Scholar] [CrossRef]
- Rodas-Junco, B.A.; Villicaña, C. Dental Pulp Stem Cells: Current Advances in Isolation, Expansion and Preservation. Tissue Eng. Regen. Med. 2017, 14, 333–347. [Google Scholar] [CrossRef] [PubMed]
- Fabbrini, M.; D’Amico, F.; Barone, M.; Conti, G.; Mengoli, M.; Brigidi, P.; Turroni, S. Polyphenol and Tannin Nutraceuticals and Their Metabolites: How the Human Gut Microbiota Influences Their Properties. Biomolecules 2022, 12, 875. [Google Scholar] [CrossRef]
- Mora, J.R.; Iwata, M.; von Andrian, U.H. Vitamin effects on the immune system: Vitamins A and D take centre stage. Nat. Rev. Immunol. 2008, 8, 685–698. [Google Scholar] [CrossRef]
- Greenwald, P.; Anderson, D.; Nelson, S.A.; Taylor, P.R. Clinical trials of vitamin and mineral supplements for cancer prevention. Am. J. Clin. Nutr. 2007, 85, 314S–317S. [Google Scholar] [CrossRef] [PubMed]
- Di Lorenzo, C.; Colombo, F.; Biella, S.; Stockley, C.; Restani, P. Polyphenols and Human Health: The Role of Bioavailability. Nutrients 2021, 13, 273. [Google Scholar] [CrossRef] [PubMed]
- Salkın, H.; Basaran, K.E. Effects of non-steroidal anti-inflammatory drug (ibuprofen) in low and high dose on stemness and biological characteristics of human dental pulp-derived mesenchymal stem cells. Connect. Tissue Res. 2023, 64, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, R.; Man, K.; Yang, X.B. Enhancing osteogenic potential of hDPSCs by resveratrol through reducing oxidative stress via the Sirt1/Nrf2 pathway. Pharm. Biol. 2022, 60, 501–508. [Google Scholar] [CrossRef]
- Feng, G.; Zheng, K.; Song, D.; Xu, K.; Huang, D.; Zhang, Y.; Cao, P.; Shen, S.; Zhang, J.; Feng, X.; et al. SIRT1 was involved in TNF-α-promoted osteogenic differentiation of human DPSCs through Wnt/β-catenin signal. In Vitro Cell. Dev. Biology. Anim. 2016, 52, 1001–1011. [Google Scholar] [CrossRef] [PubMed]
- Di Benedetto, A.; Posa, F.; De Maria, S.; Ravagnan, G.; Ballini, A.; Porro, C.; Trotta, T.; Grano, M.; Muzio, L.L.; Mori, G. Polydatin, Natural Precursor of Resveratrol, Promotes Osteogenic Differentiation of Mesenchymal Stem Cells. Int. J. Med. Sci. 2018, 15, 944–952. [Google Scholar] [CrossRef]
- Fu, X.; Feng, Y.; Shao, B.; Zhang, Y. Taxifolin Protects Dental Pulp Stem Cells under Hypoxia and Inflammation Conditions. Cell Transplant. 2021, 30, 9636897211034452. [Google Scholar] [CrossRef]
- Huo, J.F.; Zhang, M.L.; Wang, X.X.; Zou, D.H. Chrysin induces osteogenic differentiation of human dental pulp stem cells. Exp. Cell Res. 2021, 400, 112466. [Google Scholar] [CrossRef]
- Posa, F.; Di Benedetto, A.; Colaianni, G.; Cavalcanti-Adam, E.A.; Brunetti, G.; Porro, C.; Trotta, T.; Grano, M.; Mori, G. Vitamin D Effects on Osteoblastic Differentiation of Mesenchymal Stem Cells from Dental Tissues. Stem Cells Int. 2016, 2016, 9150819. [Google Scholar] [CrossRef] [PubMed]
- Posa, F.; Di Benedetto, A.; Cavalcanti-Adam, E.A.; Colaianni, G.; Porro, C.; Trotta, T.; Brunetti, G.; Lo Muzio, L.; Grano, M.; Mori, G. Vitamin D Promotes MSC Osteogenic Differentiation Stimulating Cell Adhesion and αVβ3 Expression. Stem Cells Int. 2018, 2018, 6958713. [Google Scholar] [CrossRef]
- Nam, S.H.; Yamano, A.; Kim, J.A.; Lim, J.; Baek, S.H.; Kim, J.E.; Kwon, T.G.; Saito, Y.; Teruya, T.; Choi, S.Y.; et al. Prenylflavonoids isolated from Macaranga tanarius stimulate odontoblast differentiation of human dental pulp stem cells and tooth root formation via the mitogen-activated protein kinase and protein kinase B pathways. Int. Endod. J. 2021, 54, 1142–1154. [Google Scholar] [CrossRef]
- Alipour, M.; Sharifi, S.; Samiei, M.; Shahi, S.; Aghazadeh, M.; Dizaj, S.M. Synthesis, characterization, and evaluation of Hesperetin nanocrystals for regenerative dentistry. Sci. Rep. 2023, 13, 2076. [Google Scholar] [CrossRef] [PubMed]
- Samiei, M.; Abedi, A.; Sharifi, S.; Maleki Dizaj, S. Early Osteogenic Differentiation Stimulation of Dental Pulp Stem Cells by Calcitriol and Curcumin. Stem Cells Int. 2021, 2021, 9980137. [Google Scholar] [CrossRef] [PubMed]
- Samiei, M.; Arablouye Moghaddam, F.; Dalir Abdolahinia, E.; Ahmadian, E.; Sharifi, S.; Maleki Dizaj, S. Influence of Curcumin Nanocrystals on the Early Osteogenic Differentiation and Proliferation of Dental Pulp Stem Cells. J. Nanomater. 2022, 2022, 8517543. [Google Scholar] [CrossRef]
- Son, J.W.; Choi, S.H.; Jang, J.H.; Koh, J.T.; Oh, W.M.; Hwang, Y.C.; Lee, B.N. Irisin promotes odontogenic differentiation and angiogenic potential in human dental pulp cells. Int. Endod. J. 2021, 54, 399–412. [Google Scholar] [CrossRef] [PubMed]
- Escobar, L.M.; Bendahan, Z.; Bayona, A.; Castellanos, J.E.; González, M.C. Effect of Vitamins D and E on the Proliferation, Viability, and Differentiation of Human Dental Pulp Stem Cells: An In Vitro Study. Int. J. Dent. 2020, 2020, 8860840. [Google Scholar] [CrossRef] [PubMed]
- Rasouli-Ghahroudi, A.A.; Akbari, S.; Najafi-Alishah, M.; Bohloli, M. The Effect of Vitamin K2 on Osteogenic Differentiation of Dental Pulp Stem Cells: An In Vitro Study. J. Regen. Reconstr. Restor. (Triple R) 2017, 2, 26–29. [Google Scholar] [CrossRef]
- Khanna-Jain, R.; Mannerström, B.; Vuorinen, A.; Sándor, G.K.; Suuronen, R.; Miettinen, S. Osteogenic differentiation of human dental pulp stem cells on β-tricalcium phosphate/poly (l-lactic acid/caprolactone) three-dimensional scaffolds. J. Tissue Eng. 2012, 3, 2041731412467998. [Google Scholar] [CrossRef]
- Di Benedetto, A.; Posa, F.; Marazzi, M.; Kalemaj, Z.; Grassi, R.; Lo Muzio, L.; Comite, M.D.; Cavalcanti-Adam, E.A.; Grassi, F.R.; Mori, G. Osteogenic and Chondrogenic Potential of the Supramolecular Aggregate T-LysYal®. Front. Endocrinol. 2020, 11, 285. [Google Scholar] [CrossRef]
- Khampatee, V.; Zhang, C.; Chou, L. Effects of Aspirin on Odontogenesis of Human Dental Pulp Cells and TGF-β1 Liberation from Dentin In Vitro. Int. J. Dent. 2022, 2022, 3246811. [Google Scholar] [CrossRef]
- Pang, N.S.; Lee, S.J.; Kim, E.; Shin, D.M.; Cho, S.W.; Park, W.; Zhang, X.; Jung, I.Y. Effect of EDTA on attachment and differentiation of dental pulp stem cells. J. Endod. 2014, 40, 811–817. [Google Scholar] [CrossRef] [PubMed]
- Galler, K.M.; D’Souza, R.N.; Federlin, M.; Cavender, A.C.; Hartgerink, J.D.; Hecker, S.; Schmalz, G. Dentin conditioning codetermines cell fate in regenerative endodontics. J. Endod. 2011, 37, 1536–1541. [Google Scholar] [CrossRef]
- Zand, V.; Salem Milani, A.; Primus, C.; Aghazade, M.; Mokhtari, H.; Bagheri Sabzevar, S.; Tehranchi, P. Comparison of the effect of NaOCL, curcumin, and EDTA on differentiation, proliferation, and adhesion of dental pulp stem cells. J. Oral Biol. Craniofacial Res. 2023, 13, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Ballini, A.; Mastrangelo, F.; Gastaldi, G.; Tettamanti, L.; Bukvic, N.; Cantore, S.; Cocco, T.; Saini, R.; Desiate, A.; Gherlone, E.; et al. Osteogenic differentiation and gene expression of dental pulp stem cells under low-level laser irradiation: A good promise for tissue engineering. J. Biol. Regul. Homeost. Agents 2015, 29, 813–822. [Google Scholar] [PubMed]
- Kawashima, N. Characterisation of dental pulp stem cells: A new horizon for tissue regeneration? Arch. Oral Biol. 2012, 57, 1439–1458. [Google Scholar] [CrossRef]
- Liu, H.; Gronthos, S.; Shi, S. Dental pulp stem cells. Methods Enzymol. 2006, 419, 99–113. [Google Scholar] [CrossRef]
- Yang, X.; Li, L.; Xiao, L.; Zhang, D. Recycle the dental fairy’s package: Overview of dental pulp stem cells. Stem Cell Res. Ther. 2018, 9, 347. [Google Scholar] [CrossRef]
- Yang, X.; van den Dolder, J.; Walboomers, X.F.; Zhang, W.; Bian, Z.; Fan, M.; Jansen, J.A. The odontogenic potential of STRO-1 sorted rat dental pulp stem cells in vitro. J. Tissue Eng. Regen. Med. 2007, 1, 66–73. [Google Scholar] [CrossRef]
- Gronthos, S.; Brahim, J.; Li, W.; Fisher, L.W.; Cherman, N.; Boyde, A.; DenBesten, P.; Robey, P.G.; Shi, S. Stem cell properties of human dental pulp stem cells. J. Dent. Res. 2002, 81, 531–535. [Google Scholar] [CrossRef]
- Grottkau, B.E.; Purudappa, P.P.; Lin, Y.F. Multilineage differentiation of dental pulp stem cells from green fluorescent protein transgenic mice. Int. J. Oral Sci. 2010, 2, 21–27. [Google Scholar] [CrossRef]
- Shi, S.; Gronthos, S. Perivascular niche of postnatal mesenchymal stem cells in human bone marrow and dental pulp. J. Bone Miner. Res. 2003, 18, 696–704. [Google Scholar] [CrossRef]
- Sloan, A.J.; Smith, A.J. Stem cells and the dental pulp: Potential roles in dentine regeneration and repair. Oral Dis. 2007, 13, 151–157. [Google Scholar] [CrossRef]
- Baolin, L.; Inami, Y.; Tanaka, H.; Inagaki, N.; Iinuma, M.; Nagai, H. Resveratrol inhibits the release of mediators from bone marrow-derived mouse mast cells in vitro. Planta Med. 2004, 70, 305–309. [Google Scholar] [CrossRef]
- Knutson, M.D.; Leeuwenburgh, C. Resveratrol and novel potent activators of SIRT1: Effects on aging and age-related diseases. Nutr. Rev. 2008, 66, 591–596. [Google Scholar] [CrossRef]
- Biswas, P.; Dellanoce, C.; Vezzoli, A.; Mrakic-Sposta, S.; Malnati, M.; Beretta, A.; Accinni, R. Antioxidant Activity with Increased Endogenous Levels of Vitamin C, E and A Following Dietary Supplementation with a Combination of Glutathione and Resveratrol Precursors. Nutrients 2020, 12, 3224. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Li, F.; Zhao, C.; Ming, Y.; Zheng, C.; Li, Y.; Lei, S.; Chen, C. Effects and Mechanisms of Traditional Chinese Herbal Medicine in the Treatment of Ischemic Cardiomyopathy. Pharmacol. Res. 2020, 151, 104488. [Google Scholar] [CrossRef] [PubMed]
- Poulsen, M.M.; Ornstrup, M.J.; Harsløf, T.; Jessen, N.; Langdahl, B.L.; Richelsen, B.; Jørgensen, J.O.L.; Pedersen, S.B. Short-term resveratrol supplementation stimulates serum levels of bone-specific alkaline phosphatase in obese non-diabetic men. J. Funct. Foods 2014, 6, 305–310. [Google Scholar] [CrossRef]
- Poornima, B.; Korrapati, P.S. Fabrication of chitosan-polycaprolactone composite nanofibrous scaffold for simultaneous delivery of ferulic acid and resveratrol. Carbohydr. Polym. 2017, 157, 1741–1749. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wu, Y.; Wang, J.; Zhang, C.; Yan, H.; Zhu, M.; Wang, K.; Li, C.; Xu, Q.; Kong, D. Effect of Resveratrol on Modulation of Endothelial Cells and Macrophages for Rapid Vascular Regeneration from Electrospun Poly(ε-caprolactone) Scaffolds. ACS Appl. Mater. Interfaces 2017, 9, 19541–19551. [Google Scholar] [CrossRef]
- Riccitiello, F.; De Luise, A.; Conte, R.; D’Aniello, S.; Vittoria, V.; Di Salle, A.; Calarco, A.; Peluso, G. Effect of resveratrol release kinetic from electrospun nanofibers on osteoblast and osteoclast differentiation. Eur. Polym. J. 2018, 99, 289–297. [Google Scholar] [CrossRef]
- Chen, J.P.; Chang, Y.S. Preparation and characterization of composite nanofibers of polycaprolactone and nanohydroxyapatite for osteogenic differentiation of mesenchymal stem cells. Colloids Surf. B Biointerfaces 2011, 86, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Serafini, M.; Peluso, I.; Raguzzini, A. Flavonoids as anti-inflammatory agents. Proc. Nutr. Soc. 2010, 69, 273–278. [Google Scholar] [CrossRef]
- Yi, Y.S. Regulatory Roles of Flavonoids on Inflammasome Activation during Inflammatory Responses. Mol. Nutr. Food Res. 2018, 62, e1800147. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, T.P.; Sheu, S.Y.; Sun, J.S.; Chen, M.H. Icariin inhibits osteoclast differentiation and bone resorption by suppression of MAPKs/NF-κB regulated HIF-1α and PGE(2) synthesis. Phytomedicine 2011, 18, 176–185. [Google Scholar] [CrossRef]
- Sak, K. Characteristic features of cytotoxic activity of flavonoids on human cervical cancer cells. Asian Pac. J. Cancer Prev. APJCP 2014, 15, 8007–8019. [Google Scholar] [CrossRef]
- Swarnkar, G.; Sharan, K.; Siddiqui, J.A.; Mishra, J.S.; Khan, K.; Khan, M.P.; Gupta, V.; Rawat, P.; Maurya, R.; Dwivedi, A.K.; et al. A naturally occurring naringenin derivative exerts potent bone anabolic effects by mimicking oestrogen action on osteoblasts. Br. J. Pharmacol. 2012, 165, 1526–1542. [Google Scholar] [CrossRef] [PubMed]
- Sunil, C.; Xu, B. An insight into the health-promoting effects of taxifolin (dihydroquercetin). Phytochemistry 2019, 166, 112066. [Google Scholar] [CrossRef]
- Chen, X.; Gu, N.; Xue, C.; Li, B.R. Plant flavonoid taxifolin inhibits the growth, migration and invasion of human osteosarcoma cells. Mol. Med. Rep. 2018, 17, 3239–3245. [Google Scholar] [CrossRef] [PubMed]
- Anthony, K.P.; Saleh, M.A. Free Radical Scavenging and Antioxidant Activities of Silymarin Components. Antioxidants 2013, 2, 398–407. [Google Scholar] [CrossRef]
- Topal, F.; Nar, M.; Gocer, H.; Kalin, P.; Kocyigit, U.M.; Gülçin, İ.; Alwasel, S.H. Antioxidant activity of taxifolin: An activity-structure relationship. J. Enzym. Inhib. Med. Chem. 2016, 31, 674–683. [Google Scholar] [CrossRef]
- Liu, X.; Ma, Y.; Luo, L.; Zong, D.; Li, H.; Zeng, Z.; Cui, Y.; Meng, W.; Chen, Y. Dihydroquercetin suppresses cigarette smoke induced ferroptosis in the pathogenesis of chronic obstructive pulmonary disease by activating Nrf2-mediated pathway. Phytomedicine 2022, 96, 153894. [Google Scholar] [CrossRef]
- Feng, X.; Qin, H.; Shi, Q.; Zhang, Y.; Zhou, F.; Wu, H.; Ding, S.; Niu, Z.; Lu, Y.; Shen, P. Chrysin attenuates inflammation by regulating M1/M2 status via activating PPARγ. Biochem. Pharmacol. 2014, 89, 503–514. [Google Scholar] [CrossRef]
- Ha, S.K.; Moon, E.; Kim, S.Y. Chrysin suppresses LPS-stimulated proinflammatory responses by blocking NF-κB and JNK activations in microglia cells. Neurosci. Lett. 2010, 485, 143–147. [Google Scholar] [CrossRef]
- Shin, E.K.; Kwon, H.S.; Kim, Y.H.; Shin, H.K.; Kim, J.K. Chrysin, a natural flavone, improves murine inflammatory bowel diseases. Biochem. Biophys. Res. Commun. 2009, 381, 502–507. [Google Scholar] [CrossRef] [PubMed]
- Weng, M.S.; Ho, Y.S.; Lin, J.K. Chrysin induces G1 phase cell cycle arrest in C6 glioma cells through inducing p21Waf1/Cip1 expression: Involvement of p38 mitogen-activated protein kinase. Biochem. Pharmacol. 2005, 69, 1815–1827. [Google Scholar] [CrossRef] [PubMed]
- Woo, K.J.; Jeong, Y.J.; Park, J.W.; Kwon, T.K. Chrysin-induced apoptosis is mediated through caspase activation and Akt inactivation in U937 leukemia cells. Biochem. Biophys. Res. Commun. 2004, 325, 1215–1222. [Google Scholar] [CrossRef]
- Zeng, W.; Yan, Y.; Zhang, F.; Zhang, C.; Liang, W. Chrysin promotes osteogenic differentiation via ERK/MAPK activation. Protein Cell 2013, 4, 539–547. [Google Scholar] [CrossRef] [PubMed]
- Pileggi, R.; Antony, K.; Johnson, K.; Zuo, J.; Holliday, L.S. Propolis inhibits osteoclast maturation. Dent. Traumatol. 2009, 25, 584–588. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, X. Chrysin Attenuates High Glucose-Induced BMSC Dysfunction via the Activation of the PI3K/AKT/Nrf2 Signaling Pathway. Drug Des. Dev. Ther. 2022, 16, 165–182. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, Z.; Luo, Y.; Li, X.; Huang, G.; Chen, H.; Li, A.; Qin, S. The Role of Flavonoids in the Osteogenic Differentiation of Mesenchymal Stem Cells. Front. Pharmacol. 2022, 13, 849513. [Google Scholar] [CrossRef]
- Santos, C.M.M.; Silva, A.M.S. The Antioxidant Activity of Prenylflavonoids. Molecules 2020, 25, 696. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Yuan, L.; Wang, X.; Zhang, T.L.; Wang, K. Icaritin and its glycosides enhance osteoblastic, but suppress osteoclastic, differentiation and activity in vitro. Life Sci. 2007, 81, 832–840. [Google Scholar] [CrossRef]
- Lim, R.Z.L.; Li, L.; Chew, N.; Yong, E.L. The prenylflavonoid Icaritin enhances osteoblast proliferation and function by signal transducer and activator of transcription factor 3 (STAT-3) regulation of C-X-C chemokine receptor type 4 (CXCR4) expression. Bone 2017, 105, 122–133. [Google Scholar] [CrossRef] [PubMed]
- Lim, R.Z.L.; Li, L.; Yong, E.L.; Chew, N. STAT-3 regulation of CXCR4 is necessary for the prenylflavonoid Icaritin to enhance mesenchymal stem cell proliferation, migration and osteogenic differentiation. Biochim. Et Biophys. Acta (BBA)-Gen. Subj. 2018, 1862, 1680–1692. [Google Scholar] [CrossRef]
- Ming, L.G.; Ge, B.F.; Wang, M.G.; Chen, K.M. Comparison between 8-prenylnarigenin and narigenin concerning their activities on promotion of rat bone marrow stromal cells’ osteogenic differentiation in vitro. Cell Prolif. 2012, 45, 508–515. [Google Scholar] [CrossRef] [PubMed]
- Galati, E.M.; Monforte, M.T.; Kirjavainen, S.; Forestieri, A.M.; Trovato, A.; Tripodo, M.M. Biological effects of hesperidin, a citrus flavonoid. (Note I): Antiinflammatory and analgesic activity. Farmaco 1994, 40, 709–712. [Google Scholar]
- Xue, D.; Chen, E.; Zhang, W.; Gao, X.; Wang, S.; Zheng, Q.; Pan, Z.; Li, H.; Liu, L. The role of hesperetin on osteogenesis of human mesenchymal stem cells and its function in bone regeneration. Oncotarget 2017, 8, 21031–21043. [Google Scholar] [CrossRef] [PubMed]
- Xiao, S.; Liu, W.; Bi, J.; Liu, S.; Zhao, H.; Gong, N.; Xing, D.; Gao, H.; Gong, M. Anti-inflammatory effect of hesperidin enhances chondrogenesis of human mesenchymal stem cells for cartilage tissue repair. J. Inflamm. 2018, 15, 14. [Google Scholar] [CrossRef] [PubMed]
- Farokhzad, O.C.; Langer, R. Impact of nanotechnology on drug delivery. ACS Nano 2009, 3, 16–20. [Google Scholar] [CrossRef] [PubMed]
- Kasprzak-Drozd, K.; Oniszczuk, T.; Gancarz, M.; Kondracka, A.; Rusinek, R.; Oniszczuk, A. Curcumin and Weight Loss: Does It Work? Int. J. Mol. Sci. 2022, 23, 639. [Google Scholar] [CrossRef] [PubMed]
- Hatcher, H.; Planalp, R.; Cho, J.; Torti, F.M.; Torti, S.V. Curcumin: From ancient medicine to current clinical trials. Cell. Mol. Life Sci. 2008, 65, 1631–1652. [Google Scholar] [CrossRef]
- Sharifi, S.; Zununi Vahed, S.; Ahmadian, E.; Maleki Dizaj, S.; Abedi, A.; Hosseiniyan Khatibi, S.M.; Samiei, M. Stem Cell Therapy: Curcumin Does the Trick. Phytother. Res. 2019, 33, 2927–2937. [Google Scholar] [CrossRef]
- Khanizadeh, F.; Rahmani, A.; Asadollahi, K.; Ahmadi, M.R.H. Combination therapy of curcumin and alendronate modulates bone turnover markers and enhances bone mineral density in postmenopausal women with osteoporosis. Arch. Endocrinol. Metab. 2018, 62, 438–445. [Google Scholar] [CrossRef]
- Son, H.E.; Kim, E.J.; Jang, W.G. Curcumin induces osteoblast differentiation through mild-endoplasmic reticulum stress-mediated such as BMP2 on osteoblast cells. Life Sci. 2018, 193, 34–39. [Google Scholar] [CrossRef]
- Tahmasebi Birgani, M.; Erfani-Moghadam, V.; Babaei, E.; Najafi, F.; Zamani, M.; Shariati, M.; Nazem, S.; Farhangi, B.; Motahari, P.; Sadeghizadeh, M. Dendrosomal nano-curcumin; The novel formulation to improve the anticancer properties of curcumin. Prog. Biol. Sci. 2015, 5, 143–158. [Google Scholar] [CrossRef]
- Rajalakshmi, N.; Dhivya, N. A Review on the preparation methods of Curcumin Nanoparticles. PharmaTutor 2018, 6, 6–10. [Google Scholar] [CrossRef]
- Colaianni, G.; Cinti, S.; Colucci, S.; Grano, M. Irisin and musculoskeletal health. Ann. N. Y. Acad. Sci. 2017, 1402, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Curtis, K.M.; Aenlle, K.K.; Roos, B.A.; Howard, G.A. 24R,25-dihydroxyvitamin D3 promotes the osteoblastic differentiation of human mesenchymal stem cells. Mol. Endocrinol. 2014, 28, 644–658. [Google Scholar] [CrossRef]
- De Kok, I.J.; Hicok, K.C.; Padilla, R.J.; Young, R.G.; Cooper, L.F. Effect of vitamin D pretreatment of human mesenchymal stem cells on ectopic bone formation. J. Oral Implantol. 2006, 32, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Oakes, B.; Bolia, I.K.; Weber, A.E.; Petrigliano, F.A. Vitamin C in orthopedic practices: Current concepts, novel ideas, and future perspectives. J. Orthop. Res. 2021, 39, 698–706. [Google Scholar] [CrossRef] [PubMed]
- Atkins, G.J.; Welldon, K.J.; Wijenayaka, A.R.; Bonewald, L.F.; Findlay, D.M. Vitamin K promotes mineralization, osteoblast-to-osteocyte transition, and an anticatabolic phenotype by γ-carboxylation-dependent and -independent mechanisms. Am. J. Physiol. Cell Physiol. 2009, 297, C1358–C1367. [Google Scholar] [CrossRef]
- Koshihara, Y.; Hoshi, K.; Okawara, R.; Ishibashi, H.; Yamamoto, S. Vitamin K stimulates osteoblastogenesis and inhibits osteoclastogenesis in human bone marrow cell culture. J. Endocrinol. 2003, 176, 339–348. [Google Scholar] [CrossRef]
- Khodabandehloo, F.; Taleahmad, S.; Aflatoonian, R.; Rajaei, F.; Zandieh, Z.; Nassiri-Asl, M.; Eslaminejad, M.B. Microarray analysis identification of key pathways and interaction network of differential gene expressions during osteogenic differentiation. Hum. Genom. 2020, 14, 43. [Google Scholar] [CrossRef]
- Khanna-Jain, R.; Vuorinen, A.; Sándor, G.K.; Suuronen, R.; Miettinen, S. Vitamin D(3) metabolites induce osteogenic differentiation in human dental pulp and human dental follicle cells. J. Steroid Biochem. Mol. Biol. 2010, 122, 133–141. [Google Scholar] [CrossRef]
- Mojarad, F.; Amiri, I.; Rafatjou, R.; Janeshin, A.; Farhadian, M. The Effect of 1α,25(OH)2D3 on Osteogenic Differentiation of Stem Cells from Dental Pulp of Exfoliated Deciduous Teeth. J. Dent. 2016, 17, 348–353. [Google Scholar]
- Urayama, S.; Kawakami, A.; Nakashima, T.; Tsuboi, M.; Yamasaki, S.; Hida, A.; Ichinose, Y.; Nakamura, H.; Ejima, E.; Aoyagi, T.; et al. Effect of vitamin K2 on osteoblast apoptosis: Vitamin K2 inhibits apoptotic cell death of human osteoblasts induced by Fas, proteasome inhibitor, etoposide, and staurosporine. J. Lab. Clin. Med. 2000, 136, 181–193. [Google Scholar] [CrossRef] [PubMed]
- Rickard, D.J.; Sullivan, T.A.; Shenker, B.J.; Leboy, P.S.; Kazhdan, I. Induction of rapid osteoblast differentiation in rat bone marrow stromal cell cultures by dexamethasone and BMP-2. Dev. Biol. 1994, 161, 218–228. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.B.; Willis, A.L. Aspirin selectively inhibits prostaglandin production in human platelets. Nat. New Biol. 1971, 231, 235–237. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, C.; Liu, S.; Liu, D.; Xu, X.; Chen, X.; Shi, S. Acetylsalicylic acid treatment improves differentiation and immunomodulation of SHED. J. Dent. Res. 2015, 94, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Yamaza, T.; Miura, Y.; Bi, Y.; Liu, Y.; Akiyama, K.; Sonoyama, W.; Patel, V.; Gutkind, S.; Young, M.; Gronthos, S.; et al. Pharmacologic stem cell based intervention as a new approach to osteoporosis treatment in rodents. PLoS ONE 2008, 3, e2615. [Google Scholar] [CrossRef]
- Abd Rahman, F.; Mohd Ali, J.; Abdullah, M.; Abu Kasim, N.H.; Musa, S. Aspirin Enhances Osteogenic Potential of Periodontal Ligament Stem Cells (PDLSCs) and Modulates the Expression Profile of Growth Factor-Associated Genes in PDLSCs. J. Periodontol. 2016, 87, 837–847. [Google Scholar] [CrossRef] [PubMed]
- Sculean, A.; Chiantella, G.C.; Windisch, P.; Gera, I.; Reich, E. Clinical evaluation of an enamel matrix protein derivative (Emdogain) combined with a bovine-derived xenograft (Bio-Oss) for the treatment of intrabony periodontal defects in humans. Int. J. Periodontics Restor. Dent. 2002, 22, 259–267. [Google Scholar]
- Yuan, M.; Zhan, Y.; Hu, W.; Li, Y.; Xie, X.; Miao, N.; Jin, H.; Zhang, B. Aspirin promotes osteogenic differentiation of human dental pulp stem cells. Int. J. Mol. Med. 2018, 42, 1967–1976. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.F.; Cheng, S.L.; Mbalaviele, G.; Donsante, C.; Watkins, M.; Radice, G.L.; Civitelli, R. Accentuated ovariectomy-induced bone loss and altered osteogenesis in heterozygous N-cadherin null mice. J. Bone Miner. Res. 2006, 21, 1897–1906. [Google Scholar] [CrossRef] [PubMed]
- Rainsford, K.D. Ibuprofen: Pharmacology, efficacy and safety. Inflammopharmacology 2009, 17, 275–342. [Google Scholar] [CrossRef]
- Akrami, H.; Moradi, B.; Borzabadi Farahani, D.; Mehdizadeh, K. Ibuprofen reduces cell proliferation through inhibiting Wnt/β catenin signaling pathway in gastric cancer stem cells. Cell Biol. Int. 2018, 42, 949–958. [Google Scholar] [CrossRef]
- Becker, C.; Wilson, J.C.; Jick, S.S.; Meier, C.R. Non-steroidal anti-inflammatory drugs and the risk of head and neck cancer: A case-control analysis. Int. J. Cancer 2015, 137, 2424–2431. [Google Scholar] [CrossRef]
- Schack, A.; Fransgaard, T.; Klein, M.F.; Gögenur, I. Perioperative Use of Nonsteroidal Anti-inflammatory Drugs Decreases the Risk of Recurrence of Cancer After Colorectal Resection: A Cohort Study Based on Prospective Data. Ann. Surg. Oncol. 2019, 26, 3826–3837. [Google Scholar] [CrossRef]
- Shen, W.; Zhang, X.; Du, R.; Gao, W.; Wang, J.; Bao, Y.; Yang, W.; Luo, N.; Li, J. Ibuprofen mediates histone modification to diminish cancer cell stemness properties via a COX2-dependent manner. Br. J. Cancer 2020, 123, 730–741. [Google Scholar] [CrossRef] [PubMed]
- Qiu, B.; Xu, X.; Yi, P.; Hao, Y. Curcumin reinforces MSC-derived exosomes in attenuating osteoarthritis via modulating the miR-124/NF-kB and miR-143/ROCK1/TLR9 signalling pathways. J. Cell. Mol. Med. 2020, 24, 10855–10865. [Google Scholar] [CrossRef] [PubMed]
- Lanigan, R.S.; Yamarik, T.A. Final report on the safety assessment of EDTA, calcium disodium EDTA, diammonium EDTA, dipotassium EDTA, disodium EDTA, TEA-EDTA, tetrasodium EDTA, tripotassium EDTA, trisodium EDTA, HEDTA, and trisodium HEDTA. Int. J. Toxicol. 2002, 21 (Suppl. S2), 95–142. [Google Scholar] [CrossRef] [PubMed]
- Trevino, E.G.; Patwardhan, A.N.; Henry, M.A.; Perry, G.; Dybdal-Hargreaves, N.; Hargreaves, K.M.; Diogenes, A. Effect of irrigants on the survival of human stem cells of the apical papilla in a platelet-rich plasma scaffold in human root tips. J. Endod. 2011, 37, 1109–1115. [Google Scholar] [CrossRef]
- Huang, G.T.; Yamaza, T.; Shea, L.D.; Djouad, F.; Kuhn, N.Z.; Tuan, R.S.; Shi, S. Stem/progenitor cell-mediated de novo regeneration of dental pulp with newly deposited continuous layer of dentin in an in vivo model. Tissue Eng. Part A 2010, 16, 605–615. [Google Scholar] [CrossRef]
- Galler, K.M.; Widbiller, M.; Buchalla, W.; Eidt, A.; Hiller, K.A.; Hoffer, P.C.; Schmalz, G. EDTA conditioning of dentine promotes adhesion, migration and differentiation of dental pulp stem cells. Int. Endod. J. 2016, 49, 581–590. [Google Scholar] [CrossRef] [PubMed]
- Di Benedetto, A.; Brunetti, G.; Posa, F.; Ballini, A.; Grassi, F.R.; Colaianni, G.; Colucci, S.; Rossi, E.; Cavalcanti-Adam, E.A.; Lo Muzio, L.; et al. Osteogenic differentiation of mesenchymal stem cells from dental bud: Role of integrins and cadherins. Stem Cell Res. 2015, 15, 618–628. [Google Scholar] [CrossRef]
- Greenbaum, A.M.; Revollo, L.D.; Woloszynek, J.R.; Civitelli, R.; Link, D.C. N-cadherin in osteolineage cells is not required for maintenance of hematopoietic stem cells. Blood 2012, 120, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Conti, V.; Izzo, V.; Corbi, G.; Russomanno, G.; Manzo, V.; De Lise, F.; Di Donato, A.; Filippelli, A. Antioxidant Supplementation in the Treatment of Aging-Associated Diseases. Front. Pharmacol. 2016, 7, 24. [Google Scholar] [CrossRef] [PubMed]
- Levis, S.; Theodore, G. Summary of AHRQ’s comparative effectiveness review of treatment to prevent fractures in men and women with low bone density or osteoporosis: Update of the 2007 report. J. Manag. Care Pharm. 2012, 18, S1–S15. [Google Scholar] [CrossRef]
- Gunn, C.A.; Weber, J.L.; McGill, A.T.; Kruger, M.C. Increased intake of selected vegetables, herbs and fruit may reduce bone turnover in post-menopausal women. Nutrients 2015, 7, 2499–2517. [Google Scholar] [CrossRef]
- Şöhretoğlu, D.; Baran, M.Y.; Arroo, R.; Kuruüzüm-Uz, A. Recent advances in chemistry, therapeutic properties and sources of polydatin. Phytochem. Rev. 2018, 17, 973–1005. [Google Scholar] [CrossRef]
- Chen, Z.; Wei, Q.; Hong, G.; Chen, D.; Liang, J.; He, W.; Chen, M.H. Polydatin induces bone marrow stromal cells migration by activation of ERK1/2. Biomed. Pharmacother. 2016, 82, 49–53. [Google Scholar] [CrossRef]
- Shen, Y.S.; Chen, X.J.; Wuri, S.N.; Yang, F.; Pang, F.X.; Xu, L.L.; He, W.; Wei, Q.S. Polydatin improves osteogenic differentiation of human bone mesenchymal stem cells by stimulating TAZ expression via BMP2-Wnt/β-catenin signaling pathway. Stem Cell Res. Ther. 2020, 11, 204. [Google Scholar] [CrossRef] [PubMed]
- Lanzilli, G.; Cottarelli, A.; Nicotera, G.; Guida, S.; Ravagnan, G.; Fuggetta, M.P. Anti-inflammatory effect of resveratrol and polydatin by in vitro IL-17 modulation. Inflammation 2012, 35, 240–248. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.L.; Gao, J.P.; Han, Y.L.; Xu, X.; Wu, R.; Gao, Y.; Cui, X.H. Comparative studies of polydatin and resveratrol on mutual transformation and antioxidative effect in vivo. Phytomedicine 2015, 22, 553–559. [Google Scholar] [CrossRef]
- Pullisaar, H.; Colaianni, G.; Lian, A.M.; Vandevska-Radunovic, V.; Grano, M.; Reseland, J.E. Irisin promotes growth, migration and matrix formation in human periodontal ligament cells. Arch. Oral Biol. 2020, 111, 104635. [Google Scholar] [CrossRef] [PubMed]
- Colaianni, G.; Cuscito, C.; Mongelli, T.; Pignataro, P.; Buccoliero, C.; Liu, P.; Lu, P.; Sartini, L.; Di Comite, M.; Mori, G.; et al. The myokine irisin increases cortical bone mass. Proc. Natl. Acad. Sci. USA 2015, 112, 12157–12162. [Google Scholar] [CrossRef]
- Colucci, S.C.; Buccoliero, C.; Sanesi, L.; Errede, M.; Colaianni, G.; Annese, T.; Khan, M.P.; Zerlotin, R.; Dicarlo, M.; Schipani, E.; et al. Systemic Administration of Recombinant Irisin Accelerates Fracture Healing in Mice. Int. J. Mol. Sci. 2021, 22, 10863. [Google Scholar] [CrossRef]
- Luo, Y.; Ma, Y.; Qiao, X.; Zeng, R.; Cheng, R.; Nie, Y.; Li, S.; Shen, X.; Yang, M.; Xu, C.C.; et al. Irisin ameliorates bone loss in ovariectomized mice. Climacteric 2020, 23, 496–504. [Google Scholar] [CrossRef]
- Oranger, A.; Zerlotin, R.; Buccoliero, C.; Sanesi, L.; Storlino, G.; Schipani, E.; Kozloff, K.M.; Mori, G.; Colaianni, G.; Colucci, S.; et al. Irisin Modulates Inflammatory, Angiogenic, and Osteogenic Factors during Fracture Healing. Int. J. Mol. Sci. 2023, 24, 1809. [Google Scholar] [CrossRef]
- Kim, H.; Wrann, C.D.; Jedrychowski, M.; Vidoni, S.; Kitase, Y.; Nagano, K.; Zhou, C.; Chou, J.; Parkman, V.A.; Novick, S.J.; et al. Irisin Mediates Effects on Bone and Fat via αV Integrin Receptors. Cell 2018, 175, 1756–1768.e17. [Google Scholar] [CrossRef]
- Storlino, G.; Colaianni, G.; Sanesi, L.; Lippo, L.; Brunetti, G.; Errede, M.; Colucci, S.; Passeri, G.; Grano, M. Irisin Prevents Disuse-Induced Osteocyte Apoptosis. J. Bone Miner. Res. 2020, 35, 766–775. [Google Scholar] [CrossRef]
- Posa, F.; Colaianni, G.; Di Cosola, M.; Dicarlo, M.; Gaccione, F.; Colucci, S.; Grano, M.; Mori, G. The Myokine Irisin Promotes Osteogenic Differentiation of Dental Bud-Derived MSCs. Biology 2021, 10, 295. [Google Scholar] [CrossRef] [PubMed]
- Barthes, J.; Özçelik, H.; Hindié, M.; Ndreu-Halili, A.; Hasan, A.; Vrana, N.E. Cell microenvironment engineering and monitoring for tissue engineering and regenerative medicine: The recent advances. BioMed Res. Int. 2014, 2014, 921905. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, D.T. “Ins” and “Outs” of mesenchymal stem cell osteogenesis in regenerative medicine. World J. Stem Cells 2014, 6, 94–110. [Google Scholar] [CrossRef] [PubMed]
- Cantone, E.; Castagna, G.; Sicignano, S.; Ferranti, I.; Rega, F.; Di Rubbo, V.; Iengo, M. Impact of intranasal sodium hyaluronate on the short-term quality of life of patients undergoing functional endoscopic sinus surgery for chronic rhinosinusitis. Int. Forum Allergy Rhinol. 2014, 4, 484–487. [Google Scholar] [CrossRef] [PubMed]
- Barabino, S.; De Servi, B.; Aragona, S.; Manenti, D.; Meloni, M. Efficacy of a New Ocular Surface Modulator in Restoring Epithelial Changes in an In Vitro Model of Dry Eye Syndrome. Curr. Eye Res. 2017, 42, 358–363. [Google Scholar] [CrossRef] [PubMed]
- Felzani, G.; Spoletini, I.; Convento, A.; Di Lorenzo, B.; Rossi, P.; Miceli, M.; Rosano, G. Effect of lysine hyaluronate on the healing of decubitus ulcers in rehabilitation patients. Adv. Ther. 2011, 28, 439–445. [Google Scholar] [CrossRef]
- Docheva, D.; Popov, C.; Mutschler, W.; Schieker, M. Human mesenchymal stem cells in contact with their environment: Surface characteristics and the integrin system. J. Cell. Mol. Med. 2007, 11, 21–38. [Google Scholar] [CrossRef]
- Wang, Y.K.; Chen, C.S. Cell adhesion and mechanical stimulation in the regulation of mesenchymal stem cell differentiation. J. Cell. Mol. Med. 2013, 17, 823–832. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ariano, A.; Posa, F.; Storlino, G.; Mori, G. Molecules Inducing Dental Stem Cells Differentiation and Bone Regeneration: State of the Art. Int. J. Mol. Sci. 2023, 24, 9897. https://doi.org/10.3390/ijms24129897
Ariano A, Posa F, Storlino G, Mori G. Molecules Inducing Dental Stem Cells Differentiation and Bone Regeneration: State of the Art. International Journal of Molecular Sciences. 2023; 24(12):9897. https://doi.org/10.3390/ijms24129897
Chicago/Turabian StyleAriano, Anastasia, Francesca Posa, Giuseppina Storlino, and Giorgio Mori. 2023. "Molecules Inducing Dental Stem Cells Differentiation and Bone Regeneration: State of the Art" International Journal of Molecular Sciences 24, no. 12: 9897. https://doi.org/10.3390/ijms24129897
APA StyleAriano, A., Posa, F., Storlino, G., & Mori, G. (2023). Molecules Inducing Dental Stem Cells Differentiation and Bone Regeneration: State of the Art. International Journal of Molecular Sciences, 24(12), 9897. https://doi.org/10.3390/ijms24129897