Abstract
It is generally recognized that the root uptake capacity of grafted plants strongly depends on the rootstocks’ well-developed root system. However, we found that grafted plants showed different nitrate uptake capacities when different varieties of oriental melon scion were grafted onto the same squash rootstock, suggesting that the scion regulated the nitrate uptake capacity of the rootstock root. In this study, we estimated the nitrate uptake capacity of grafted plants with the different oriental melon varieties’ seedlings grafted onto the same squash rootstocks. The results indicated a significant difference in the nitrate uptake rate and activity of two heterologous grafting plants. We also showed a significant difference in CmoNRT2.1 expression in the roots of two grafting combinations and verified the positive regulation of nitrate uptake by CmoNRT2.1 expression. In addition, the two varieties of oriental melon scion had highly significant differences in CmHY5 expression, which was transported to the rootstock and positively induced CmoHY5-1 and CmoHY5-2 expression in the rootstock roots. Meanwhile, CmHY5 could positively regulate CmoNRT2.1 expression in the rootstock roots. Furthermore, CmoHY5-1 and CmoHY5-2 also positively regulated CmoNRT2.1 expression, respectively, and CmoHY5-1 dominated the positive regulation of CmoNRT2.1, while CmHY5 could interact with CmoHY5-1 and CmoHY5-2, respectively, to jointly regulate CmoNRT2.1 expression. The oriental melon scion regulated the nitrate uptake capacity of the melon/squash grafting plant roots, and the higher expression of CmHY5 in the oriental melon scion leaves, the more substantial the nitrate uptake capacity of squash rootstock roots.
1. Introduction
The grafting technique is widely used in plant production to overcome continuous cropping obstacles and soil-borne diseases, increase crop yield, and improve resistance to adversity [1,2,3,4,5,6]. In addition, grafting could effectively improve the plant’s ability to absorb water and nutrients. Studies have shown that grafted tomatoes, apples, and other plants have a more robust nutrient absorption capacity than own-root plants [1,7,8,9,10]. Using squash or gourd as the rootstock, grafting watermelon plants had a more substantial absorption capacity of nitrogen, potassium, calcium, magnesium, iron, and manganese [11]. However, the physiological and molecular mechanism of grafts regulating plant growth, development, and morphogenesis becomes more complicated due to the genomes of two or more species in grafted plants.
The various small molecules (small RNA, messenger RNA) involved in the interaction of the genome of grafted plants [12,13,14,15,16,17,18,19,20,21] caused a large number of new gene modifications and expression changes in rootstocks and scions, as well as changes in the phenotype and function of grafted plants [22,23]. Small RNAs such as miRNA and siRNA, the long-distance messengers, move systematically and play an essential role in chromatin modification, mRNA stability, and the downstream regulation of translation [24]. miRNA and siRNA could move from cell to cell through vasculature [16], induce gene silencing, inhibit gene expression [25,26,27], and participate in epigenetic modifications in plants [28,29]. mRNAs’ movement in phloem regulated the plant growth and development and defense responses. However, not all mobile mRNAs performed their functions. The mRNAs as regulators of scion–rootstock interaction could be translated into proteins at the transport location. Moreover, mRNAs transported only occurred in specific transcripts [30]. Many mobile mRNAs have been identified using RNA-Seq in multiple species. mRNAs might be transported from stems and leaves to roots and in reverse [31]. The 138 mobile mRNAs were identified in the heterograft combination of arabidopsis and tobacco [32], and a large number of mobile mRNAs were also detected in grafted grapes by transcriptome detection [33]. In the homograft arabidopsis, proteins in the cells associated with the new buds were transported to the root and participated in the regulation of root growth [34]. It was found that transcription factor SlCyp1 could be transported from scion to rootstock in grafted tomato plants and regulate the root–shoot ratio [35].
ELONGATED HYPOCOTYL5 (HY5) is a long-distance transport transcription factor that regulates the expression of genes containing cis-acting elements G-BOX or Z-BOX and miRNAs [36,37,38,39,40]. Notably, the long-distance transport transcription factor HY5 is essential in regulating plant nutrient uptake and accumulation. HY5 could regulate the expression of sulfite reductase genes, which play an essential role in plant sulfate assimilation [41]. HY5 also interacted with copper signal transcription factor SPL7 and affected copper signal allocation by regulating miRNA408 expression [42]. HY5 was also involved in regulating the expression of nitrite reductase genes and ammonium transporter protein genes to affect nitrogen uptake and utilization in plants [43]. HY5 affected nitrate uptake in plants by regulating the expression of nitrate transporter NRT2.1 [44]. Moreover, in the homograft experiments, HY5 could transport long distances from the scion to the root system, playing an essential role in scion–rootstock interaction [44]. However, how long-distance transcription factor HY5 regulates the nitrate uptake capacity of the heterograft plant, such as the grafted plant of melon scion grafted onto the squash rootstock, is even unclear.
In the protected cultivation of melons and watermelon, the squash and pumpkin are often taken as the rootstock. Grafted melons and watermelons not only improve disease and cold resistance but also enhance the absorption capacity of their root system [2,45]. The increased root absorption capacity of grafted melon and watermelon plants is often thought to be due to the rootstock’s well-developed and vigorous roots. However, our team recently found a significant difference in the root absorption capacity of different melon scions grafted onto the same squash rootstocks, but the underlying mechanism has not yet been reported. Therefore, it is theoretically worth clarifying the underlying molecular mechanism of the differences in nitrate uptake of grafted melon plants and further optimizing the selection of rootstocks and scions for grafting cultivation of melon in practice.
2. Results
2.1. Melon Scions Affect CmoNRT2.1 Expression of Squash Rootstocks to Modulate the Nitrate Uptake Capacity of Grafted Plants
We grafted two different scions of oriental melon (JS-1408 and JS-1429) onto the same squash rootstock (ZM-1507) and obtained two heterologous grafting plants (JS-1408/ZM-1507 and JS-1429/ZM-1507). When the grafted seedlings were grown up with five leaves, we used the 15N stable isotope tracer technique to measure and analyze the difference in the nitrate uptake rate and activity between JS-1408/ZM-1507 and JS-1429/ZM-1507. The nitrate uptake rate and activity of JS-1408/ZM1507 were 1.57 and 1.19 times that of JS-1429/ZM1507, respectively (Figure 1A,B). The results showed a significant difference (p < 0.001), so we suggested that the significant difference in the nitrate uptake capacity of grafted seedlings was due to the different varieties of oriental melon scions.

Figure 1.
Analysis of differences in nitrate uptake capacity and CmoNRT2.1 expression of grafted melon plants with two different varieties of oriental melon scions grafted onto the same squash rootstocks during the five leaves growth stage. CmoNRT2.1 transiently silenced plants: TRV-CmoNRT2.1; Control plants: TRV-CK. Values are means ± SD, n = 3 (biological replicates). Asterisks indicate statistically significant differences (*** p < 0.001, Student’s t-test). (A,B). Nitrate uptake efficiency and nitrate uptake activity by 15N stable isotope tracing assay. (C). Quantitative real-time (qRT) PCR analysis of CmoNRT2.1 expression. (D). Analysis of the silencing efficiency of transient silenced CmoNRT2.1 in the grafted plants. (E,F). Analysis of nitrate uptake rate and uptake activity in the grafted plants’ roots system of transient silenced CmoNRT2.1.
The nitrate transporter gene (NRT) mainly regulates nitrogen uptake and utilization in plants. The molecular mechanism of nitrate uptake in arabidopsis, rice, tomato, barley, and other crops has been well studied. Plants absorb nitrate from the soil through high-affinity and low-affinity systems, and NRT2.1, a member of the nitrate transporter gene families in the high-affinity absorption system, plays an essential role in the absorption of nitrate by roots. Our studies found a significant difference (p < 0.01) by measuring the CmoNRT2.1 expression level in rootstock roots of JS-1408/ZM-1507 and JS-1429/ZM-1507 (Figure 1C). In addition, JS-1408/ZM-1507, with a higher nitrate uptake capacity, also had a higher CmoNRT2.1 expression level than JS-1408/ZM-1507. Therefore, we hypothesized that the oriental melon scion regulates the expression of CmoNRT2.1 in the rootstock to control the nitrate uptake capacity of the grafting seedlings. In order to intensely figure it out, we performed a transient silencing assay of CmoNRT2.1 in the grafted plant roots and measured the nitrate uptake rate and activity using the 15N stable isotope tracer technique. The results of the silencing efficiency assay showed that the CmoNRT2.1 expression of the transient silenced plants was significantly lower than that of the control (Figure 1D). Moreover, the transient silenced plants’ nitrate uptake rate (Figure 1E) and uptake activity (Figure 1F) were significantly lower than those of the control. The results indicated that the oriental melon scion could control the nitrate capacity of the squash rootstocks by regulating CmoNRT2.1 expression in the grafted plants.
2.2. Melon Scion CmHY5 mRNA Undergoes Long-Distance Transport to the Rootstock and Modulates the Nitrate Uptake by Regulating CmoNRT2.1 Expression of Squash Rootstock Roots
Plant roots’ nitrate uptake capacity is closely related to nitric acid transporters (NRTs) and is also affected by light. It has been found that HY5, a long-distance transcription factor, plays an essential regulatory role in plant nitrogen absorption and assimilation [44,46]. HY5 is a bZIP transcription factor that can sense light signals and transmit light signals to photoreceptors, thus regulating plant growth and development [47,48]. In order to clarify whether the different oriental melon scions had a significant effect on the nitrate uptake capacity of grafted plants, we compared the expression level of CmHY5 in the leaves of own-root grafted seedlings (JS-1408 and JS-1429) using the TaqMan technique and found that the expression level of CmHY5 in the leaves of JS-1408 was more than two times that of JS-1429 (Figure 2A). Then, we determined the expression level in the scion leaves, scion stems, rootstock stems, and rootstock roots of two kinds of grafted plants (JS-1429/ZM-1507 and JS-1408/ZM-1507). The results showed that CmHY5 mRNA could be transported over long distances into rootstocks, and the expression level of CmHY5 in four different parts of JS-1408/ZM-1507 was significantly higher than that of JS-1429/ZM-1507 (Figure 2B–E).
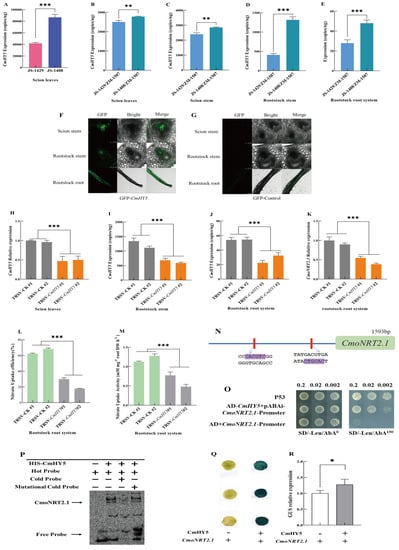
Figure 2.
CmHY5 in the oriental melon scion positively modulates the nitrate uptake of squash rootstock roots by regulating CmoNRT2.1 expression. Values are means ± SD, n = 3 (biological replicates). Asterisks indicate statistically significant differences (*** p < 0.001, ** p < 0.01, * p < 0.05, Student’s t-test). (A). TaqMan analysis of CmHY5 expression in the leaves of two different own-grafted melon plants. (B). TaqMan analysis of CmHY5 expression in the scion leaves of grafted plants with two different varieties of oriental melon scions grafted onto the same squash rootstock. (C). TaqMan analysis of CmHY5 expression in the scion stems of grafted plants with two different varieties of oriental melon scions grafted onto the same squash rootstock. (D). TaqMan analysis of CmHY5 expression in the rootstock stems of grafted plants with two different varieties of oriental melon scions grafted onto the same squash rootstock. (E). TaqMan analysis of CmHY5 expression in the rootstock roots of grafted plants with two different varieties of oriental melon scions grafted onto the same squash rootstock. (F). Analysis of GFP-CmHY5 transport in the grafted plants. (G). Analysis of GFP-Control transport in the grafted plants. (H). qRT-PCR analysis of the relative expression of CmHY5 in the oriental melon scion leaves of grafted plants with transient silenced CmHY5. (I). TaqMan analysis of CmHY5 expression in the squash rootstock stems of grafted plants with transient silenced CmHY5. (J). TaqMan analysis for detecting CmHY5 expression in the squash rootstock roots of grafted plants with transient silenced CmHY5. (K). qRT-PCR analysis of the relative expression of CmoNRT2.1 in the squash rootstocks of grafted plants with transient silenced CmHY5. (L,M). The nitrogen uptake rate and uptake activity assay of grafted plants with transient silenced CmHY5. (N). Analysis of the cis-acting element of the CmoNRT2.1 promoter. (O). Yeast one-hybrid assay to detect CmHY5 binding to the CmoNRT2.1 promoter. (P). EMSA assay to detect CmHY5 binding to the CmoNRT2.1 promoter. (Q,R). GUS staining and GUS activity assay in N. benthamiana leaves after transient co-expression of 35S::CmHY5 with proCmoNRT2.1::GUS and co-expression with pBI101:GUS as a control.
Interestingly, the grafted plants, JS-1408/ZM-1507, with high CmoNRT2.1 expression levels and a strong nitrate uptake capacity, also had high CmHY5 expression levels. To examine whether CmHY5 mRNA was translated and transported into the rootstocks, transient expressions of CmHY5-GFP in melon scion leaves were conducted. We found that the GFP signal could be detected in the phloem of the scion, rootstock stems, and rootstock roots after 72 h (Figure 2F,G). These results suggested that CmHY5 mRNA in melon scion transported into the squash rootstocks might significantly promote nitrate uptake by up-regulating CmoNRT2.1 expression in grafted plants.
In order to further explore the relationship between transcription factor CmHY5 and the nitrate uptake capacity of grafted melon plants, we used the VIGS technique to transient silence CmHY5 in melon scions and measured the transient silencing efficiency (Figure 2H). The results showed that the CmHY5 expression level was significantly reduced in grafted plants’ rootstock stems and roots (Figure 2I,J), and the relative expression level of CmoNRT2.1, the nitrate uptake rate, and the nitrate uptake activity as well (Figure 2K–M). Therefore, it was evidence that transcription factor CmHY5 was transported into the rootstock from the scions to regulate the nitrate uptake of grafted melon plants.
However, the underlying molecular mechanism of the long-distance transcription factor CmHY5 regulating nitrate uptake in the oriental melon scion grafted onto the squash rootstock remains unclear. We analyzed the promoter of CmoNRT2.1 and found it contained the two cis-acting elements G-box that CmHY5 could bind (Figure 2N). Then, we conducted a yeast single hybridization test between CmHY5 and the CmoNRT2.1 promoter. The results showed that CmHY5 could bind to the promoter of CmoNRT2.1 (Figure 2O). Since the two binding elements are the same, we designed the hot probe, cold competitive probe, and mutation cold competitive probe using the G-box element and carried out in vitro EMSA, further proving that CmHY5 could bind to the G-box element on the promoter of CmoNRT2.1 (Figure 2P). We also performed a GUS reporter gene test in tobacco, and the infected leaves were punched and stained (Figure 2Q). Meanwhile, the activity of the GUS protein was also measured (Figure 2R), indicating that CmHY5 could be bound to the promoter of CmoNRT2.1 and positively regulate its expression (p < 0.05). We speculated that the transcription factor CmHY5 might positively regulate the significantly increased expression of CmoNRT2.1 through the other molecular mechanisms.
2.3. CmoHY5 in Rootstocks Modulates the Nitrate Uptake by Regulating CmoNRT2.1 Expression of Squash Rootstock Roots
The above experiments demonstrated that CmHY5 could bind to the CmoNRT2.1 promoter and positively regulate its expression. However, CmoHY5 was present in squash rootstocks in the heterograft plants of oriental melon scion grafted onto squash rootstock, so we suggest that CmoHY5 of squash rootstocks might affect the CmoNRT2.1 expression, and we were uncertain of CmoHY5 like CmHY5 to long-distance transport. Therefore, we measured the expression of CmoHY5-1 and CmoHY5-2 in different parts of JS-1429/ZM-1507 and JS-1408/ZM-1507 (scion leaf, scion stem, rootstock stem, and rootstock root). The results indicated that CmoHY5-1 and CmoHY5-2 expressions were not detected in the melon scion leaves and stems, while CmoHY5-1 and CmoHY5-2 expressions could be detected in the squash rootstock stems and roots of JS-1429/ZM-1507 and JS-1408/ZM-1507 (Figure 3A,B). Remarkably, the expression of CmoHY5-1 and CmoHY5-2 in the stems and roots of JS-1408/ZM-1507 was significantly higher than those of JS-1429/ZM-1507. In addition, the expression of CmoHY5-1 was significantly higher than that of CmoHY5-2 in JS-1429/ZM-1507 and JS-1408/ZM-1507. According to the nitrate uptake capacity of JS-1429/ZM-1507 and JS-1408/ZM-1507 (Figure 1A,B), we concluded that the expression of CmoHY5-1 and CmoHY5-2 in the squash rootstocks was also positively correlated with the nitrate uptake rate and uptake activity of the grafted melon plant.

Figure 3.
CmoHY5 regulates the expression of CmoNRT2.1 in the squash rootstock roots. Values are means ± SD, n = 3 (biological replicates). Asterisks indicate statistically significant differences (*** p < 0.001, ** p < 0.01, Student’s t-test). (A). TaqMan analysis of CmoHY5 expression in the squash rootstock stems of grafted plants with two different varieties of oriental melon scions grafted onto the same squash rootstock. (B). TaqMan analysis of CmoHY5 expression in the rootstock roots of grafted plants with two different varieties of oriental melon scions grafted onto the same squash rootstock. (C). Yeast one-hybrid assay detecting CmoHY5 binding to the CmoNRT2.1 promoter. (D). EMSA assay to detect CmoHY5-1 binding to the CmoNRT2.1 promoter. (E). EMSA assay to detect CmoHY5-2 binding to the CmoNRT2.1 promoter. (F,G). GUS staining and GUS activity assay in N. benthamiana leaves after transient co-expression of 35S::CmoHY5-1 with proCmoNRT2.1::GUS and co-expression with pBI101:GUS as a control. (H,I). GUS staining and GUS activity assay in N. benthamiana leaves after transient co-expression of 35S::CmoHY5-2 with proCmoNRT2.1::GUS and co-expression with pBI101:GUS as a control.
Therefore, we speculate that CmoHY5-1 and CmoHY5-2 also regulated the expression of CmoNRT2.1. Then, the yeast one-hybrid assay demonstrated that both CmoHY5-1 and CmoHY5-2 could bind to the promoter of CmoNRT2.1 (Figure 3C). The EMSA assay also showed that CmoHY5-1 and CmoHY5-2 could bind to the G-BOX on the CmoNRT2.1 promoter (Figure 3D,E). In addition, we also performed the tobacco GUS reporter gene assay and stained the infested leaves by punching, and GUS protein activity was measured. The results indicated that both CmoHY5-1 and CmoHY5-2 positively regulated the expression of CmoNRT2.1. However, the promoter activity of CmoHY5-2 on CmoNRT2.1 was weak compared to the promoter activity of CmoHY5-1 on CmoNRT2.1(Figure 3F–I). Thus, during this process, CmoHY5-1 has a significant regulatory role in the expression of CmoNRT2.1.
2.4. CmHY5 in Melon Scion Positively Regulates CmoHY5 Expression in Squash Rootstock
Given the experimental results, CmHY5, CmoHY5-1, and CmoHY5-2 all positively regulated the expression of CmoNRT2.1. In addition, there were also highly significant differences in the expression of CmoHY5-1 and CmoHY5-2 in rootstock stems and the roots of the grafted plants with two different melon scions grafted onto one type of squash rootstock (JS-1429/ZM-1507, JS-1408/ZM-1507) (Figure 3A,B). Furthermore, the grafted plants with the higher expression of CmHY5 owned the higher expression of CmoHY5-1 and CmoHY5-2. However, the direct promotion of CmoNRT2.1 expression by CmHY5 was insignificant, so we speculated that CmoHY5 mediated the regulation of the squash rootstock CmoNRT2.1 by the melon scion CmHY5. To confirm the relationship between CmHY5 and CmoHY5, we first measured the expression of CmoHY5-1 and CmoHY5-2 in the rootstock stems and roots of grafted melon plants with transient silenced CmHY5 in melon scions and found that grafted plants with a lower expression of CmHY5 had a lower expression of CmoHY5-1 and CmoHY5-2 in the rootstock stems and roots (Figure 4A–D). Moreover, we analyzed the promoters of CmoHY5-1 and CmoHY5-2 and found that they both contained the CmHY5 binding element G-BOX in their promoters (Figure 4E). A yeast one-hybrid assay was performed using CmHY5 with the promoters of CmoHY5-1 and CmoHY5-2, respectively. The results showed that CmHY5 could bind to the promoters of CmoHY5-1 and CmoHY5-2, respectively (Figure 4F). An EMSA assay was also carried out with these two components, and the results showed that CmHY5 could bind to both components (Figure 4G,H). In addition, we carried out a tobacco GUS reporter gene assay, in which GUS staining and GUS protein activity were measured on infested tobacco leaves. The results demonstrated that CmHY5 positively regulated the expression of CmoHY5-1 and CmoHY5-2, and the expression of CmoHY5-1 was more effectively promoted (Figure 4I–L).
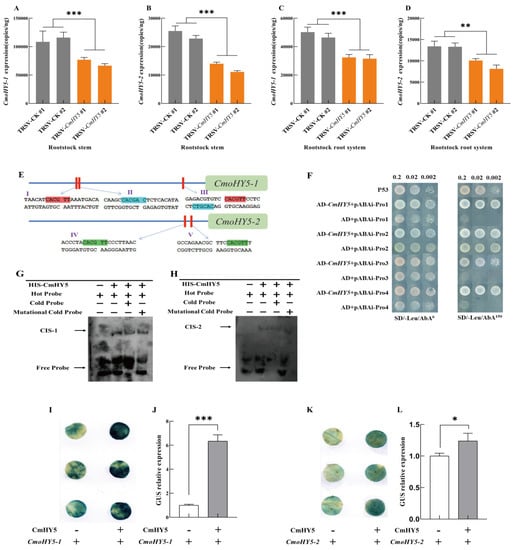
Figure 4.
CmHY5 in the oriental melon scion regulates the expression of CmoHY5 in the squash rootstock. Values are means ± SD, n = 3 (biological replicates). Asterisks indicate statistically significant differences (*** p < 0.001, ** p < 0.01, * p < 0.05, Student’s t-test). (A). TaqMan analysis of CmoHY5-1 expression in the squash rootstock stems of grafted oriental melon plants with transient silenced CmHY5 in the oriental melon scions. (B). TaqMan analysis of CmoHY5-2 expression in the squash rootstock stems of grafted melon plants with transient silenced CmHY5 in the oriental melon scions. (C). TaqMan analysis of CmoHY5-1 expression in the squash rootstock roots of grafted oriental melon plants with transient silenced CmHY5 in the oriental melon scions. (D). TaqMan analysis of CmoHY5-2 expression in the squash rootstock roots of grafted plants with transient silenced CmHY5 in the oriental melon scions. (E). Analysis of the promoter cis-acting elements of CmoHY5-1 and CmoHY5-2. (F). Yeast one-hybrid assay to detect CmHY5 binding to the promoters of CmoHY5-1 and CmoHY5-2, respectively. (G). EMSA assay to detect promoter binding of CmHY5 to CmoHY5-1. (H). EMSA assay to detect promoter binding of CmHY5 to CmoHY5-2. (I,J). GUS staining and GUS activity assay in N. benthamiana leaves after transient co-expression of 35S::CmHY5 with proCmoHY5-1::GUS and co-expression with pBI101:GUS as a control. (K,L). GUS staining and GUS activity assay in N. benthamiana leaves after transient co-expression of 35S::CmHY5 with proCmoHY5-2::GUS and co-expression with pBI101:GUS as a control.
2.5. CmHY5 Protein of Melon Scion Interacts with CmoHY5 Protein of Squash Rootstock
Previous studies have shown that transcription factor HY5 not only forms dimers on its own to function but also binds to its promoter to accelerate its expression under light [49,50]. Therefore, we suggested that the CmHY5 protein of melon scion might interact with the CmoHY5 protein of squash rootstock. On the one hand, the yeast two-hybrid assay showed that CmHY5 protein could interact with CmoHY5-1 and CmoHY5-2, respectively (Figure 5A). We carried out further fluorophore complementation assays and bimolecular fluorescence complementation assays, which also showed the same results (Figure 5B,C).

Figure 5.
Analysis of protein interaction between CmHY5 and CmoHY5. (A). Y2H assay analysis of CmHY5 interacting with CmoHY5-1 and CmoHY5-2 proteins, respectively. (B). Fluorophore enzyme complementation assay analysis of CmHY5 interacting with CmoHY5-1 and CmoHY5-2 proteins, respectively. (C). Bimolecular fluorescence complementation assay analysis of CmHY5 interacting with CmoHY5-1 and CmoHY5-2 proteins, respectively.
3. Discussion
Grafting uses the rootstock root to replace the scion root, thus improving the grafting plants’ resistance and nutrient uptake. Many studies have shown that grafting could improve the uptake of mineral nutrition such as nitrogen, potassium, calcium, magnesium, iron, and manganese [7,9,10,11,45,51,52]. In the grafted plant, exchanging information between rootstock and scion is a mutual process. It has been shown that the rootstock controls the accumulation and distribution of salt ions in the scion [53,54], and scion growth and yield are also regulated by the rootstock [55]. In addition, the rootstock affects the biomass of the scion, while the scion also affects the biomass of the rootstock [56,57,58]. However, our studies found that two grafting combinations of the two different varieties of oriental melon scion grafted onto the same squash rootstock showed highly significant differences in nitrate uptake rate and activity (Figure 1A,B). In further investigation, we found that CmoNRT2.1 regulated the nitrate uptake capacity of squash rootstock roots and the higher the expression of CmoNRT2.1, the better the nitrate uptake capacity of the grafted plants (Figure 1C–F). We therefore suggested that the oriental melon scion would have a regulatory effect on the nitrate uptake of the squash rootstock, a process involving long-distance transport and regulation of signaling substances in the grafted plant.
The regulation of scion to rootstock in grafted plants is regulated by various long-distance transport signals such as hormones, RNA, DNA, and proteins [24,59,60,61,62,63]. Studies in apples have shown that more growth hormone is transported downwards when multi-branched apple scions are grafted on the same rootstock compared to controls, resulting in higher growth hormone and cytokinin levels in the roots and suppressed root growth [64]. In addition, gibberellins and cytokinins are mobile and transported in grafted plants and are involved in regulating graft healing and plant growth and development [65,66]. Besides the plant hormones, long-distance transport of RNA, DNA, and proteins occurs in plants, with relatively few processes involving proteins and DNA compared to RNA, and various studies have shown that proteins can accompany mRNA transport [59,67,68]. It can also be long-distance transported to the rootstock in grafted tomato plants. SlCyp1 was transported from the scion to the rootstock and involved in regulating the root-to-shoot ratio [35]. In the arabidopsis homologous grafting combination, proteins from the companion cells of the new shoot can be transported to the root and regulate root growth [34]. DNA interchange provides the basis for graft-induced genetic variation, and grafting provides a channel for forming asexual species [69,70]. In the grafting complex, RNA has a critical role in substances that are transported over long distances, including various types of small RNAs as well as mRNAs [61,62,71,72,73]. It has been shown that mRNA transport is involved in stress signaling and response and fruit quality regulation [74,75], and mobile mRNA CAX3 regulates iron and zinc uptake [76]. It has also been shown that the transport of non-cell autonomous mRNAs is regulated by specific RNA mobile elements and is also related to the structure of the RNA [30]. Therefore, studying the role of mRNAs in rootstock is essential for the future regulation of growth and development, morphogenesis, and fruit quality in grafted plants. In this study, we examined CmHY5 mRNA in all tissues (oriental melon scion leaves and stems, squash rootstock stems and roots) and found that RNA was detected in all tissues, but the mRNA content of CmHY5 was lower in the rootstock stems and roots compared to the scion leaves and stems, especially in the rootstock root system, which was 58–89 times lower compared to the scion leaves. The results indicated that CmHY5 mRNA could be transported as mRNA (Figure 2A–E). In addition, we attached CmHY5 to GFP fluorescence and then infiltrated the leaves of oriental melon scion in grafted plants (Figure 2F,G). After 72 h, GFP signals were detected in the oriental melon scion stem, squash rootstock stem, and root, further indicating that CmHY5 of the oriental melon scion could be transported to the squash rootstock, either in the form of protein or possibly translated after transport to function. Several studies have been conducted on HY5 as a signaling substance for long-distance transport. HY5 can function through branches sensing FR light to the root system to regulate root growth [77] and transported from leaves to roots to activate the expression of FER transcription factors and regulate iron uptake in plants [78]. In addition, HY5 can be transported from the silique to the root system to regulate lateral root growth and orientation [77,79,80,81]. We also measured the CmoHY5 expression in various tissues and showed that no expression of CmoHY5-1 and CmoHY5-2 was detected in the oriental melon scions and that expression was higher in both the stems and roots of the squash rootstock, indicating that CmoHY5 produced by the rootstock is not transported upwards. Furthermore, it has been shown that HY5 can form dimers on its own to function and bind its promoter to promote its expression in the light [49,50]. Therefore, we hypothesized that CmHY5 produced by the scion would promote CmoHY5 expression in the rootstock, so we next demonstrated that CmHY5 produced by the oriental melon scion would indeed positively regulate both CmoHY5-1 and CmoHY5-2 in the squash rootstock using a Y1H assay, an EMSA assay, a tobacco GUS reporter gene assay, and a VIGS transient silencing assay (Figure 4). Among them, the promotion of CmoHY5-1 was highly significant, whereas the promotion of CmoHY5-2 was insignificant, and our results demonstrated that inter-binding of HY5 could also occur in different species (Figure 5). Moreover, they all positively regulated the CmoNRT2.1 expression to modulate the nitrate uptake capacity of the grafted plants (Figure 2 and Figure 3).
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
Oriental melon ‘JS-1408’ and ‘JS-1429’ cultivars (Cucumis melo var. makuwa Makino) were, respectively, grafted onto squash, ShengZhen No.1 cultivar (C. moschata), with the one-cotyledon graft method [82]. Grafted seedlings were transplanted into the nutritional bowl (12 cm × 12 cm) in the glasshouse at Shenyang Agriculture University, Liaoning, China (41°49′ N 123°32′ E). When the grafted plants, JS-1408/ZM-1507 and JS-1429/ZM-1507, grow to five leaves, we took the samples for subsequent qRT-PCR analysis, TaqMan analysis, and 15N stable isotope tracer test.
4.2. 15N Stable Isotope Tracing Assay
When the grafted seedlings grew to five leaves and one heart, each grafted seedling was fed with 20 mL (5 mM) K15NO3 solution (abundance 10.16%), and the grafted seedlings were divided into aerial parts from the grafting interface until the 48th hour of tracing. Take samples from the underground part, fix the green at 105 °C for 30 min, bake in an oven at 70 °C for 3–5 days to entirely remove moisture, then grind the sample and pass it through a 100-mesh sieve: ratiometric mass spectrometer (Iso Prime100 Isotope Ratio Mass Spectrometer, Germany) determination, three biological replicates. The δ15N in the samples was determined by the elementary analysis–isotope ratio mass spectrometers (EA-IRMS) method [83]. The calculation formula for 15N absorption rate and absorption activity is as follows:
15N absorption rate = (SN + RN) × (15N feeding amount)−1 × 100%;
15N absorption activity = (SN + RN) × (DWR × h)−1.
(Note: SN, 15N content in rootstock roots; RN, 15N content in scion shoots; DWR, dry rootstock root weight; h, hours.)
4.3. RNA Extraction and Reverse Transcription
TRIzol RNA isolation reagent (Kangwei Century, Taizhou, China) extracted scion leaves and stems and rootstock stems and roots of two grafting combinations (JS-1408/ZM-1507 and JS-1429/ZM-1507), as well as transiently silenced plants and silenced control plants total RNA from roots. The cDNA was obtained using a reverse transcription kit (TaKaRa, Beijing, China).
4.4. qRT-PCR
qRT-PCR was used to detect the silencing efficiency of the transient silencing test and to determine the relative expression of CmoNRT2.1 in squash rootstock roots. qRT-PCR was performed with a PCR machine (Bio-Rad, CFX96) using the Pro Taq HS SYBR Green premix qPCR kit (AG) in a 20 μL reaction system (2× SYBR Green Pro Taq HS Premix 10 μL, cDNA 2 μL, forward and reverse primer 0.4 μL each, ddH2O 7.2μL) was used for qRT-PCR analysis, with thermal cycling as follows: 95 °C for 2 min, 40 cycles for 10 s at 95 °C, and 30 s at 60 °C.
4.5. TaqMan
TaqMan was used to detect the expression levels of CmHY5, CmoHY5-1, and CmoHY5-2 in various tissues. It was performed with a PCR machine (Bio-Rad, CFX96) using SuperReal fluorescence quantitative premix reagent (probe method) kit (TIANGEN) in a 20 μL reaction system (2× Super Real Premix (Probe) 10 μL, cDNA 2 μL, forward and reverse 0.6 μL each, primer 0.4 μL, ddH2O 6.4 μL)) for TaqMan analysis, with thermal cycling as follows: 95 °C for 15 min, 40 cycles for 3 s at 95 °C, and 25 s at 60 °C. The probe was modified by 5’-FAM. The Ct values obtained by the analysis were substituted into the standard curves of the three genes to calculate the absolute expression levels. The standard curves were made as follows: the mRNA sequences of CmHY5, CmoHY5-1, and CmoHY5-2 were connected to the TA cloning vector, and the recombinant plasmid was amplified by 10-fold gradient dilution. The diluted plasmid was used as a standard using the above system and method. TaqMan assay was performed, and a standard curve was drawn based on the natural logarithm of the Ct values and the plasmid concentrations. The standard curves of CmHY5, CmoHY5-1, and CmoHY5-2 were prepared, respectively.
4.6. Virus-Induced Gene Silencing (VIGS)
The transient silencing test mediated by tobacco ringspot virus was used [84], the coding sequence (CDS) sequence of CmHY5 (477 bp) was inserted into the SnaBI restriction site of the pTRSV2 RNA2S vector. Then, the pTRSV2-CmHY5, P19, pTRSV RNA1, and pTRSV2-CuPDS were transformed into Agrobacterium-competent EHA105. P19, pTRSV RNA1, and pTRSV2-CmHY5were suspended to OD600 = 1.0 and injected into N. benthamiana leaves at a ratio of 1:1:1. pTRSV RNA1+ pTRSV2-CuPDS+P19 was used as positive disease control, and pTRSV RNA1+ pTRSV2+P19 was used as a silent control. After the diseased tobacco leaves were collected, the diseased leaves were collected for friction inoculation of fully healed thin-skinned muskmelon/Chinese pumpkin grafted scion leaves. Ten to fifteen days for sampling and 15N stable isotope tracer assay.
4.7. Subcellular Localization of CmHY5, CmoHY5-1, CmoHY5-2, and CmoNRT2.1
Full-length CDs of CmHY5, CmoHY5-1, CmoHY5-2, and CmoNRT2.1 were cloned into the 35S::GFP vector. Subsequently, the recombinant vectors were transformed into Agrobacterium-competent EHA105 and injected into N. benthamiana leaves. The leaves were stained with DAPI for 5 min to clarify CmHY5, CmoHY5-1, and CmoHY5-2 subcellular localization. As previously described [85], confocal imaging was performed with an FV3000 microscope (Olympus).
4.8. Y1H Assay
Full-length CDs of CmHY5, CmoHY5-1, and CmoHY5-2 were cloned into the pGADT7 vector to generate prey. The full-length promoter of CmoNRT2.1 and four promoter fragments, Pro1 (291 bp with two G-box), Pro2 (228 bp with a G-box), Pro3 (217 bp with a G-box), and Pro4 (111 bp with a G-box) of CmoHY5-1 and CmoHY5-2 were amplified and cloned into the pAbAi vector, producing five baits. All bait vectors require linearization and purification before transforming yeast Y1H-Gold. The bait vector was transformed into yeast Y1H-Gold separately and plated on SD/-Ura +50/100/150/200/300 ng/mL Aureobasidin A (AbA) medium and incubated at 30 °C for 3 days to screen AbA concentration. The prey vector pGADT7-CmHY5 was transformed into yeast with baits vector pAbAi-Pro1, pAbAi-Pro2, pAbAi-Pro3, and pAbAi-Pro4, separately. The prey vectors pGADT7-CmHY5, pGADT7-CmoHY5-1, and pGADT7-CmoHY5-2 were transformed into yeast with baits vector pAbAi-ProCmoNRT2.1, separately, along with positive (pGAD-p53+p53-AbAi) and negative (pGADT7-AD+bait-pAbAi) controls, were plated on SD /-Leu+150 ng/mL AbA and incubated at 30 °C for 3 days.
4.9. Electrophoretic Mobility Shift Assay (EMSA)
Full-length CDs of CmHY5, CmoHY5-1, and CmoHY5-2 were cloned into the pD2P_1.06eTM vector. Standard Mini Kit(Kangmaxin Intelligent Technology Co., Ltd. Shanghai, China) was used for protein induction following the manufacturer’s instructions. EMSA was performed using the Light Shift Chemiluminescent EMSA Kit (Beyotime, Shanghai, China), following the manufacturer’s instructions. Probes synthesized from yeast one-hybrid results. Two oligonucleotide probes with G-box were synthesized based on the sequences of the CmoHY5-1 promoter and CmoHY5-2 promoter, and one oligonucleotide probe with G-box was synthesized based on the sequences of the promoter of CmoNRT2.1 and biotin-labeled by Saibaisheng (Beijing, China). The unlabeled probes were used as competition probes. The unlabeled mutant probe was used as a mutant cold competition probe. The fusion protein was incubated at 25 °C with the probes, with or without the competition probe and mutant probe, in a total reaction volume of 10 µL. Protein–DNA complexes were separated on a 6% polyacrylamide gel and electroblotted onto a Hybondnylon membrane (Biosharp, Shanghai, China), which was then subjected to UV crosslinking and chemiluminescence detection.
4.10. GUS Assay
Full-length CDs of CmHY5, CmoHY5-1, and CmoHY5-2 were cloned into the pRI101 vector to generate an effector vector. Promoter fragments with G-box of CmoNRT2.1, CmoHY5-1, and CmoHY5-2 were amplified and cloned into the pBI101 vector to generate the reporter vector. The recombinant vectors were transformed into Agrobacterium-competent EHA105. The reporter and effector co-infect N. benthamiana leaves, and GUS activity and staining measurements were performed as previously described [64,86], along with the negative (pRI101+ reporter) controls.
4.11. Y2H Assay
Full-length CDs of CmHY5 were cloned into the pGBKT7 vector to generate bait. Full-length CDs of CmoHY5-1 and CmoHY5-2 were cloned into the pGBKT7 vector to generate prey. Recombinant plasmids were introduced into yeast strain Y2H-Gold, and the transformed yeast cells were plated on SD/-Trp/-Leu medium and incubated at 30 °C for 3 days. Then, the colonies were transferred to SD/-Trp/-Leu/-Ade/-His/AbA150/X-α-Gal medium incubated at 30 °C for 3 days.
4.12. Luciferase Complementation (LCA) Assay
Full-length CDs of CmHY5 were cloned into the NLuc vector. Full-length CDs of CmoHY5-1 and CmoHY5-2 were cloned into the CLuc vector. Recombinant plasmids were introduced into Agrobacterium-competent EHA105 (pSoup) and expressed in tobacco leaves. Fluorescence was observed using an imaging system (Berthold, Lb985, Bad Wildbad, Germany).
4.13. Bimolecular Fluorescence Complementation (BiFC) Assay
Full-length CDs of CmHY5 were cloned into the pCAMBIA1300-35s-N-YFPC vector. Full-length CDs of CmoHY5-1 and CmoHY5-2 were cloned into the pCAMBIA1300-35s-C-YFPC vector. Recombinant plasmids were introduced into Agrobacterium-competent EHA105 (pSoup) and expressed in tobacco leaves. The leaves were stained with DAPI for 5 min, and confocal imaging was performed with an FV3000 microscope (Olympus) as previously described [85].
5. Conclusions
In conclusion, the CmHY5 expression level of the scion played a vital role in the nitrate uptake capacity of oriental melon scion grafted on squash rootstock. The scion-to-rootstock mobile transcription factor CmHY5 induced the CmoHY5-1 and CmoHY5-2 expression of squash rootstock and interacted with each other. Moreover, they positively regulated CmoNRT2.1 expression jointly and individually to modulate the nitrate uptake of grafted plants (Figure 6). The grafted plants with the higher CmHY5 mRNA level of the oriental melon scion possessed a stronger nitrate uptake capacity.

Figure 6.
The model of scion-to-rootstock mobile transcription factor CmHY5 positively modulates the nitrate uptake capacity of oriental melon scion grafted onto squash rootstock. (A). The grafted plants of lower CmHY5 mRNA level. (B). The grafted plant of higher CmHY5 mRNA level.
Author Contributions
S.H. and C.X. conceived and designed the research. S.H., Y.Z., C.X., X.W. and Y.X. conducted the experiments. J.G., F.W., H.Y. and Z.S. contributed reagents and analytical tools. S.H. and C.X. wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (32272696), the China Agriculture Research System of Watermelon and Melon (CARS-25), and the Basic Research Project of Liaoning Province (LSNJC202005).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
We thank Chao Geng (Shandong Agricultural University) for kindly providing the pTRSV1 and pTRSV2 vector.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lee, J.M.; Kubota, C.; Tsao, S.J.; Bie, Z.; Echevarria, P.H.; Morra, L.; Oda, M. Current status of vegetable grafting: Diffusion, grafting techniques, automation. Sci. Hortic. 2010, 127, 93–105. [Google Scholar] [CrossRef]
- Schwarz, D.; Rouphael, Y.; Colla, G.; Venema, J.H. Grafting as a tool to improve tolerance of vegetables to abiotic stresses: Thermal stress, water stress and organic pollutants. Sci. Hortic. 2010, 127, 162–171. [Google Scholar] [CrossRef]
- López-Marín, J.; González, A.; Pérez-Alfocea, F.; Egea-Gilabert, C.; Fernández, J.A. Grafting is an efficient alternative to shading screens to alleviate thermal stress in greenhouse grown sweet pepper. Sci. Hortic. 2013, 149, 39–46. [Google Scholar] [CrossRef]
- Tsaballa, A.; Athanasiadis, C.; Pasentsis, K.; Ganopoulos, I.; Nianiou-Obeidat, I.; Tsaftaris, A. Molecular studies of inheritable grafting induced changes in pepper (Capsicum annuum) fruit shape. Sci. Hortic. 2013, 149, 2–8. [Google Scholar] [CrossRef]
- Bhatt, R.M.; Upreti, K.K.; Divya, M.H.; Bhat, S.; Pavithra, C.B.; Sadashiva, A.T. Interspecific grafting to enhance physiological resilience to flooding stress in tomato (Solanum lycopersicum L.). Sci. Hortic. 2015, 182, 8–17. [Google Scholar] [CrossRef]
- Nawaz, M.A.; Imtiaz, M.; Kong, Q.; Cheng, F.; Ahmed, W.; Huang, Y.; Bie, Z. Grafting: A technique to modify ion accumulation in horticultural crops. Front. Plant Sci. 2016, 7, 1457. [Google Scholar] [CrossRef]
- Rivero, R.M.; Ruiz, J.M.; Romero, L. Iron metabolism in tomato and watermelon plants: Influence of grafting. J. Plant Nutr. 2005, 27, 2221–2234. [Google Scholar] [CrossRef]
- Schwarz, D.; Öztekin, G.B.; Tüzel, Y.; Brückner, B.; Krumbein, A. Rootstocks can enhance tomato growth and quality characteristics at low potassium supply. Sci. Hortic. 2013, 149, 70–79. [Google Scholar] [CrossRef]
- Amiri, M.E.; Fallahi, E.; Safi-Songhorabad, M. Influence of rootstock on mineral uptake and scion growth of ‘Golden delicious’ and ‘Royal Gala’apples. J. Plant Nutr. 2014, 37, 16–29. [Google Scholar] [CrossRef]
- Chang, C.; Chao, L.; LI, C.Y.; KANG, X.Y.; ZOU, Y.J. Differences in the efficiency of potassium (K) uptake and use in five apple rootstock genotypes. J. Integr. Agric. 2014, 13, 1934–1942. [Google Scholar] [CrossRef]
- Uygur, V.; Yetisir, H. Effects of rootstocks on some growth parameters, phosphorous and nitrogen uptake watermelon under salt stress. J. Plant Nutr. 2009, 32, 629–643. [Google Scholar] [CrossRef]
- Pant, B.D.; Buhtz, A.; Kehr, J.; Scheible, W.R. MicroRNA399 is a long-distance signal for the regulation of plant phosphate homeostasis. Plant J. 2008, 53, 731–738. [Google Scholar] [CrossRef]
- Martin, A.; Adam, H.; Díaz-Mendoza, M.; Żurczak, M.; González-Schain, N.D.; Suárez-López, P. Graft-transmissible induction of potato tuberization by the microRNA miR172. Development 2009, 136, 2873–2881. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Sun, L.; Kragler, F. The phloem-delivered RNA pool contains small noncoding RNAs and interferes with translation. Plant Physiol. 2009, 150, 378–387. [Google Scholar] [CrossRef]
- Buhtz, A.; Pieritz, J.; Springer, F.; Kehr, J. Phloem small RNAs, nutrient stress responses, and systemic mobility. BMC Plant Biol. 2010, 10, 64–77. [Google Scholar] [CrossRef]
- Chitwood, D.H.; Timmermans, M.C.P. Small RNAs are on the move. Nature 2010, 467, 415–419. [Google Scholar] [CrossRef] [PubMed]
- Dunoyer, P.; Schott, G.; Himber, C.; Meyer, D.; Takeda, A.; Carrington, J.C.; Voinnet, O. Small RNA duplexes function as mobile silencing signals between plant cells. Science 2010, 328, 912–916. [Google Scholar] [CrossRef]
- Molnar, A.; Melnyk, C.W.; Bassett, A.; Hardcastle, T.J.; Dunn, R.; Baulcombe, D.C. Small silencing RNAs in plants are mobile and direct epigenetic modification in recipient cells. Science 2010, 328, 872–875. [Google Scholar] [CrossRef]
- Fragoso, V.; Goddard, H.; Baldwin, I.T.; Kim, S.G. A simple and efficient micrografting method for stably transformed Nicotiana attenuata plants to examine shoot-root signaling. Plant Methods 2011, 7, 34. [Google Scholar] [CrossRef]
- Bhogale, S.; Mahajan, A.S.; Natarajan, B.; Rajabhoj, M.; Thulasiram, H.V.; Banerjee, A.K. MicroRNA156: A potential graft-transmissible microRNA that modulates plant architecture and tuberization in Solanum tuberosum ssp. andigena. Plant Physiol. 2014, 164, 1011–1027. [Google Scholar] [CrossRef]
- Wang, J.; Jiang, L.; Wu, R. Plant grafting: How genetic exchange promotes vascular reconnection. New Phytol. 2017, 214, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Cookson, S.J.; Clemente Moreno, M.J.; Hevin, C.; Nyamba Mendome, L.Z.; Delrot, S.; Trossat-Magnin, C.; Ollat, N. Graft union formation in grapevine induces transcriptional changes related to cell wall modification, wounding, hormone signalling, and secondary metabolism. J. Exp. Bot. 2013, 64, 2997–3008. [Google Scholar] [CrossRef] [PubMed]
- Gaut, B.S.; Miller, A.J.; Seymour, D.K. Living with two genomes: Grafting and its implications for plant genome-to-genome interactions, phenotypic variation, and evolution. Annu. Rev. Genet. 2019, 53, 195–215. [Google Scholar] [CrossRef] [PubMed]
- Gouil, Q.; Lewsey, M.G. Small RNAs shoot for the root. Nat. Plants 2021, 7, 2–3. [Google Scholar] [CrossRef]
- Palauqui, J.C.; Elmayan, T.; Pollien, J.M.; Vaucheret, H. Systemic acquired silencing: Transgene-specific post-transcriptional silencing is transmitted by grafting from silenced stocks to non-silenced scions. EMBO J. 1997, 16, 4738–4745. [Google Scholar] [CrossRef]
- Voinnet, O.; Baulcombe, D.C. Systemic signalling in gene silencing. Nature 1997, 389, 553. [Google Scholar] [CrossRef]
- Melnyk, C.W.; Molnar, A.; Bassett, A.; Baulcombe, D.C. Mobile 24 nt small RNAs direct transcriptional gene silencing in the root meristems of Arabidopsis thaliana. Curr. Biol. 2011, 21, 1678–1683. [Google Scholar] [CrossRef]
- Albacete, A.; Martínez-Andújar, C.; Martínez-Pérez, A.; Thompson, A.J.; Dodd, I.C.; Pérez-Alfocea, F. Unravelling rootstock× scion interactions to improve food security. J. Exp. Bot. 2015, 66, 2211–2226. [Google Scholar] [CrossRef]
- Berger, M.M.; Gallusci, P.; Teyssier, E. Roles of epigenetic mechanisms in grafting and possible applications. Adv. Bot. Res. 2018, 88, 203–246. [Google Scholar]
- Huang, N.C.; Yu, T.S. The sequences of Arabidopsis GA-INSENSITIVE RNA constitute the motifs that are necessary and sufficient for RNA long-distance trafficking. Plant J. 2009, 59, 921–929. [Google Scholar] [CrossRef]
- Thieme, C.J.; Rojas-Triana, M.; Stecyk, E.; Schudoma, C.; Zhang, W.; Yang, L.; Miñambres, M.; Walther, D.; Schulze, W.X.; Paz-Ares, J.; et al. Endogenous Arabidopsis messenger RNAs transported to distant tissues. Nat. Plants 2015, 1, 15025. [Google Scholar] [CrossRef]
- Notaguchi, M.; Higashiyama, T.; Suzuki, T. Identification of mRNAs that move over long distances using an RNA-Seq analysis of Arabidopsis/Nicotiana benthamiana heterografts. Plant Cell Physiol. 2015, 56, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Mao, L.; Jittayasothorn, Y.; Kang, Y.; Jiao, C.; Fei, Z.; Zhong, G.Y. Messenger RNA exchange between scions and rootstocks in grafted grapevines. BMC Plant Biol. 2015, 15, 251. [Google Scholar] [CrossRef] [PubMed]
- Paultre, D.S.G.; Gustin, M.P.; Molnar, A.; Oparka, K.J. Lost in transit: Long-distance trafficking and phloem unloading of protein signals in Arabidopsis homografts. Plant Cell 2016, 28, 2016–2025. [Google Scholar] [CrossRef] [PubMed]
- Spiegelman, Z.; Ham, B.K.; Zhang, Z.; Toal, T.W.; Brady, S.M.; Zheng, Y.; Fei, Z.; Lucas, W.J.; Wolf, S. A tomato phloem-mobile protein regulates the shoot-to-root ratio by mediating the auxin response in distant organs. Plant J. 2015, 83, 853–863. [Google Scholar] [CrossRef]
- Koornneef, M.; Rolff, E.; Spruit, C.J.P. Genetic control of light-inhibited hypocotyl elongation in Arabidopsis thaliana (L.) Heynh. Z. Pflanzenphysiol. 1980, 100, 147–160. [Google Scholar] [CrossRef]
- Ang, L.H.; Chattopadhyay, S.; Wei, N.; Oyama, T.; Okada, K.; Batschauer, A.; Deng, X.W. Molecular interaction between COP1 and HY5 defines a regulatory switch for light control of Arabidopsis development. Mol. Cell 1998, 1, 213–222. [Google Scholar] [CrossRef]
- Cluis, C.P.; Mouchel, C.F.; Hardtke, C.S. The Arabidopsis transcription factor HY5 integrates light and hormone signaling pathways. Plant J. 2004, 38, 332–347. [Google Scholar] [CrossRef]
- Ulm, R.; Baumann, A.; Oravecz, A.; Máté, Z.; Ádám, É.; Oakeley, E.J.; Schäfer, E.; Nag, F. Genome-wide analysis of gene expression reveals function of the bZIP transcription factor HY5 in the UV-B response of Arabidopsis. Proc. Natl. Acad. Sci. USA 2004, 101, 1397–1402. [Google Scholar] [CrossRef]
- Lee, J.; He, K.; Stolc, V.; Lee, H.; Figueroa, P.; Gao, Y.; Tongprasit, W.; Zhao, H.; Lee, I.; Deng, X.W. Analysis of transcription factor HY5 genomic binding sites revealed its hierarchical role in light regulation of development. Plant Cell 2007, 19, 731–749. [Google Scholar] [CrossRef]
- Lee, B.R.; Koprivova, A.; Kopriva, S. The key enzyme of sulfate assimilation, adenosine 5′-phosphosulfate reductase, is regulated by HY5 in Arabidopsis. Plant J. 2011, 67, 1042–1054. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhao, X.; Li, J.; Cai, H.; Deng, X.W.; Li, L. MicroRNA408 is critical for the HY5-SPL7 gene network that mediates the coordinated response to light and copper. Plant Cell 2014, 26, 4933–4953. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Zhang, H.; Zhang, H.; Deng, X.W.; Wei, N. HY5 regulates nitrite reductase 1 (NIR1) and ammonium transporter1; 2 (AMT1; 2) in Arabidopsis seedlings. Plant Sci. 2015, 238, 330–339. [Google Scholar] [CrossRef]
- Chen, X.; Yao, Q.; Gao, X.; Jiang, C.; Harberd, N.P.; Fu, X. Shoot-to-root mobile transcription factor HY5 coordinates plant carbon and nitrogen acquisition. Curr. Biol. 2016, 26, 640–646. [Google Scholar] [CrossRef] [PubMed]
- Colla, G.; Suarez, C.M.C.; Cardarelli, M.; Rouphael, Y. Improving nitrogen use efficiency in melon by grafting. HortScience 2010, 45, 559–565. [Google Scholar] [CrossRef]
- Gaudinier, A.; Rodriguez-Medina, J.; Zhang, L.; Olson, A.; Liseron-Monfils, C.; Bågman, A.M.; Foret, J.; Abbitt, S.; Tang, M.; Li, B.; et al. Transcriptional regulation of nitrogen-associated metabolism and growth. Nature 2018, 563, 259–264. [Google Scholar] [CrossRef]
- Oyama, T.; Shimura, Y.; Okada, K. The Arabidopsis HY5 gene encodes a bZIP protein that regulates stimulus-induced development of root and hypocotyl. Genes Dev. 1997, 11, 2983–2995. [Google Scholar] [CrossRef]
- Yoon, M.K.; Shin, J.; Choi, G.; Choi, B.S. Intrinsically unstructured N-terminal domain of bZIP transcription factor HY5. Proteins Struct. Funct. Bioinform. 2006, 65, 856–866. [Google Scholar] [CrossRef]
- Holm, M.; Ma, L.G.; Qu, L.J.; Deng, X.W. Two interacting bZIP proteins are direct targets of COP1-mediated control of light-dependent gene expression in Arabidopsis. Genes Dev. 2002, 16, 1247–1259. [Google Scholar] [CrossRef]
- Abbas, N.; Maurya, J.P.; Senapati, D.; Gangappa, S.N.; Chattopadhyay, S. Arabidopsis CAM7 and HY5 physically interact and directly bind to the HY5 promoter to regulate its expression and thereby promote photomorphogenesis. Plant Cell 2014, 26, 1036–1052. [Google Scholar] [CrossRef]
- Pulgar, G.; Villora, G.; Moreno, D.A.; Romero, L. Improving the mineral nutrition in grafted watermelon plants: Nitrogen metabolism. Biol. Plant. 2000, 43, 607–609. [Google Scholar] [CrossRef]
- Huang, Y.; Zhao, L.; Kong, Q.; Cheng, F.; Niu, M.; Xie, J.; Nawaz, M.A.; Bie, Z. Comprehensive mineral nutrition analysis of watermelon grafted onto two different rootstocks. Hortic. Plant J. 2016, 2, 105–113. [Google Scholar] [CrossRef]
- Santa-Cruz, A.; Martinez-Rodriguez, M.M.; Perez-Alfocea, F.; Romero-Aranda, R.; Bolarin, M.C. The rootstock effect on the tomato salinity response depends on the shoot genotype. Plant Sci. 2002, 162, 825–831. [Google Scholar] [CrossRef]
- Sanwal, S.K.; Mann, A.; Kumar, A.; Kesh, H.; Kaur, G.; Rai, A.K.; Kumar, R.; Sharma, P.C.; Kumar, A.; Bahadur, A.; et al. Salt Tolerant Eggplant Rootstocks Modulate Sodium Partitioning in Tomato Scion and Improve Performance under Saline Conditions. Agriculture 2022, 12, 183. [Google Scholar] [CrossRef]
- Zijlstra, S.; Groot, S.P.C.; Jansen, J. Genotypic variation of rootstocks for growth and production in cucumber; possibilities for improving the root system by plant breeding. Sci. Hortic. 1994, 56, 185–196. [Google Scholar] [CrossRef]
- Martínez-Ballesta, M.C.; Alcaraz-López, C.; Muries, B.; Mota-Cadenas, C.; Carvajal, M. Physiological aspects of rootstock-scion interactions. Sci Hortic. 2010, 127, 112–118. [Google Scholar] [CrossRef]
- Tandonnet, J.P.; Cookson, S.J.; Vivin, P.; Ollat, N. Scion genotype controls biomass allocation and root development in grafted grapevine. Aust. J. Grape Wine Res. 2010, 16, 290–300. [Google Scholar] [CrossRef]
- Cookson, S.J.; Hevin, C.; Donnart, M.; Ollat, N. Grapevine rootstock effects on scion biomass are not associated with large modifications of primary shoot growth under nonlimiting conditions in the first year of growth. Funct. Plant Biol. 2012, 39, 650–660. [Google Scholar] [CrossRef]
- Xoconostle-Cázares, B.; Xiang, Y.; Ruiz-Medrano, R.; Wang, H.L.; Monzer, J.; Yoo, B.C.; Mcfarland, K.C.; Franceschi, V.R.; Lucas, W.J. Plant paralog to viral movement protein that potentiates transport of mRNA into the phloem. Science 1999, 283, 94–98. [Google Scholar] [CrossRef]
- Wu, R.; Wang, X.; Lin, Y.; Ma, Y.; Liu, G.; Yu, X.; Zhong, S.; Liu, B. Inter-species grafting caused extensive and heritable alterations of DNA methylation in Solanaceae plants. PLoS ONE 2013, 8, e61995. [Google Scholar] [CrossRef]
- Zhang, Z.; Zheng, Y.; Ham, B.K.; Chen, J.; Yoshida, A.; Kochian, L.V.; Fei, Z.; Lucas, W.J. Vascular-mediated signalling involved in early phosphate stress response in plants. Nat. Plants 2016, 2, 16033. [Google Scholar] [CrossRef] [PubMed]
- Ham, B.K.; Lucas, W.J. Phloem-mobile RNAs as systemic signaling agents. Annu. Rev. Plant Biol. 2017, 68, 173–195. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Zheng, B. Molecular responses during plant grafting and its regulation by auxins, cytokinins, and gibberellins. Biomolecules 2019, 9, 397. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Ma, J.; Tan, M.; Mao, J.; An, N.; Sha, G.; Zhang, D.; Zhao, C.; Han, M. Transcriptome analysis reveals the effects of sugar metabolism and auxin and cytokinin signaling pathways on root growth and development of grafted apple. BMC Genom. 2016, 17, 150. [Google Scholar] [CrossRef] [PubMed]
- Werner, T.; Motyka, V.; Strnad, M.; Schmülling, T. Regulation of plant growth by cytokinin. Proc. Natl. Acad. Sci. USA 2001, 98, 10487–10492. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, S.; Böhlenius, H.; Moritz, T.; Nilsson, O. GA4 is the active gibberellin in the regulation of LEAFY transcription and Arabidopsis floral initiation. Plant Cell 2006, 18, 2172–2181. [Google Scholar] [CrossRef] [PubMed]
- Duan, X.; Zhang, W.; Huang, J.; Zhao, L.; Ma, C.; Hao, L.; Yuan, H.; Harada, T.; Li, T. KNOTTED1 mRNA undergoes long-distance transport and interacts with movement protein binding protein 2C in pear (Pyrus betulaefolia). Plant Cell Tissue Organ Cult. (PCTOC) 2015, 121, 109–119. [Google Scholar] [CrossRef]
- Duan, X.; Zhang, W.; Huang, J.; Hao, L.; Wang, S.; Wang, A.; Meng, D.; Zhang, Q.; Chen, Q.; Li, T. PbWoxT1 mRNA from pear (Pyrus betulaefolia) undergoes long-distance transport assisted by a polypyrimidine tract binding protein. New Phytol. 2016, 210, 511–524. [Google Scholar] [CrossRef]
- Taller, J.; Hirata, Y.; Yagishita, N.; Kita, M.; Ogata, S. Graft-induced genetic changes and the inheritance of several characteristics in pepper (Capsicum annuum L.). Theor. Appl. Genet. 1998, 97, 705–713. [Google Scholar] [CrossRef]
- Fuentes, I.; Stegemann, S.; Golczyk, H.; Karcher, D.; Bock, R. Horizontal genome transfer as an asexual path to the formation of new species. Nature 2014, 511, 232–235. [Google Scholar] [CrossRef]
- Forde, B.G. The role of long-distance signalling in plant responses to nitrate and other nutrients. J. Exp. Bot. 2002, 53, 39–43. [Google Scholar] [PubMed]
- Atkins, C.A.; Smith, P.M.C. Translocation in legumes: Assimilates, nutrients, and signaling molecules. Plant Physiol. 2007, 144, 550–561. [Google Scholar] [CrossRef]
- Lin, S.I.; Chiang, S.F.; Lin, W.Y.; Chen, J.W.; Tseng, C.Y.; Wu, P.C.; Chiou, T.J. Regulatory network of microRNA399 and PHO2 by systemic signaling. Plant Physiol. 2008, 147, 732–746. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Lozano, M.; Dutta, S.K.; Natarajan, P.; Tomason, Y.R.; Lopez, C.; Katam, R.; Levi, A.; Nimmakayala, P.; Reddy, U.K. Transcriptome changes in reciprocal grafts involving watermelon and bottle gourd reveal molecular mechanisms involved in increase of the fruit size, rind toughness and soluble solids. Plant Mol. Biol. 2020, 102, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, L.; Xing, N.; Wu, X.; Wu, X.; Wang, B.; Lu, Z.; Xu, P.; Tao, Y.; Li, G.; et al. A universal pipeline for mobile mRNA detection and insights into heterografting advantages under chilling stress. Hortic. Res. 2020, 7, 10. [Google Scholar]
- Hao, P.; Lv, X.; Fu, M.; Xu, Z.; Tian, J.; Wang, Y.; Zhang, X.; Xu, X.; Wu, T.; Han, Z. Long-distance mobile mRNA CAX3 modulates iron uptake and zinc compartmentalization. EMBO Rep. 2022, 23, e53698. [Google Scholar] [CrossRef]
- Van Gelderen, K.; Kang, C.; Paalman, R.; Keuskamp, D.; Hayes, S.; Pierik, R. Far-red light detection in the shoot regulates lateral root development through the HY5 transcription factor. Plant Cell 2018, 30, 101–116. [Google Scholar] [CrossRef]
- Guo, Z.; Xu, J.; Wang, Y.; Hu, C.; Shi, K.; Zhou, J.; Xia, X.; Zhou, Y.; Foyer, C.H.; Yu, J. The phyB-dependent induction of HY5 promotes iron uptake by systemically activating FER expression. EMBO Rep. 2021, 22, e51944. [Google Scholar] [CrossRef]
- Lee, H.J.; Ha, J.H.; Kim, S.G.; Choi, H.K.; Kim, Z.H.; Han, Y.J.; Kim, J.; Oh, I.; Fragoso, V.; Shin, K.; et al. Stem-piped light activates phytochrome B to trigger light responses in Arabidopsis thaliana roots. Sci. Signal. 2016, 9, ra106. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, C.; Xu, H.; Shi, X.; Zhen, W.; Hu, Z.; Huang, J.; Zheng, Y.; Huang, P.; Zhang, K.; et al. HY5 contributes to light-regulated root system architecture under a root-covered culture system. Front. Plant Sci. 2019, 10, 1490. [Google Scholar] [CrossRef]
- Burko, Y.; Gaillochet, C.; Seluzicki, A.; Chory, J.; Busch, W. Local HY5 activity mediates hypocotyl growth and shoot-to-root communication. Plant Commun. 2020, 1, 100078. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.R.; Perkins-Veazie, P.; Sakata, Y.; Lopez-Galarza, S.; Maroto, J.V.; Lee, S.G.; Huh, Y.C.; Sun, Z.Y.; Migule, A.; King, S.R.; et al. Cucurbit grafting. Crit. Rev. Plant Sci. 2008, 27, 50–74. [Google Scholar] [CrossRef]
- Liu, Y.; Hu, B.; Chu, C. 15N-nitrate uptake activity and root-to-shoot transport assay in rice. Bio-Protoc. 2016, 6, e1897. [Google Scholar] [CrossRef]
- Fang, L.; Wei, X.Y.; Liu, L.Z.; Zhou, L.X.; Tian, Y.P.; Geng, C.; Li, X.D. A tobacco ringspot virus-based vector system for gene and microRNA function studies in cucurbits. Plant Physiol. 2021, 186, 853–864. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.Q.; Hao, P.B.; Lv, X.M.; Tian, J.; Wang, Y.; Zhang, X.Z.; Xu, X.F.; Han, Z.H.; Wu, T. A long non-coding apple RNA, MSTRG.85814.11, acts as a transcriptional enhancer of SAUR32 and contributes to the Fe-deficiency response. Plant J. 2020, 103, 53–67. [Google Scholar] [CrossRef] [PubMed]
- Spartz, A.K.; Lee, S.H.; Wenger, J.P.; Gonzalez, N.; Itoh, H.; Inzé, D.; Peer, W.A.; Murphy, A.S.; Overvoorde, P.J.; Gray, W.M. The SAUR19 subfamily of SMALL AUXIN UP RNA genes promote cell expansion. Plant J. 2012, 70, 978–990. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).