
Identification of a Major QTL and Validation of Related Genes for Tiller Angle in Rice Based on QTL Analysis


 , ,
, ,  ,
,  ,
,  , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
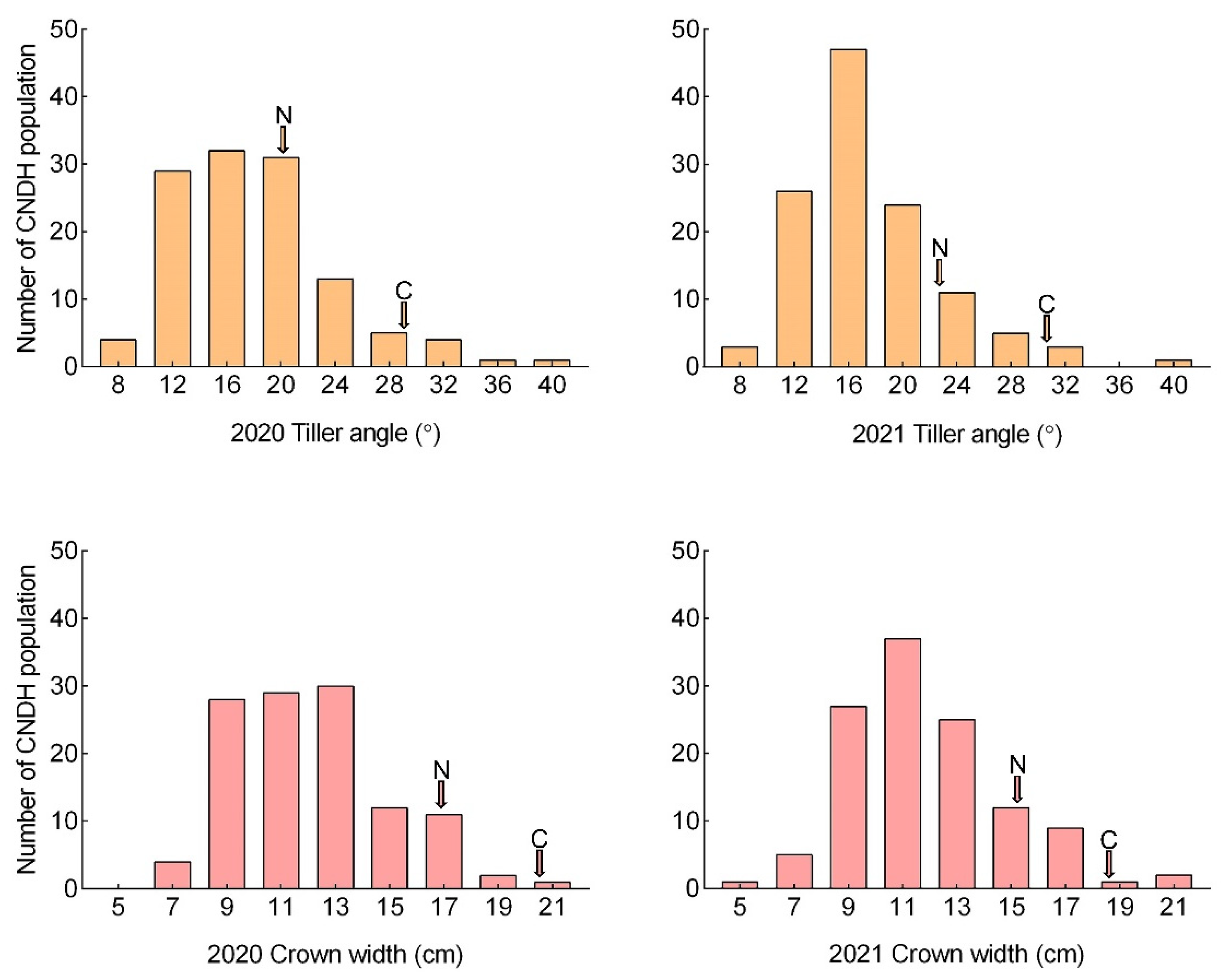
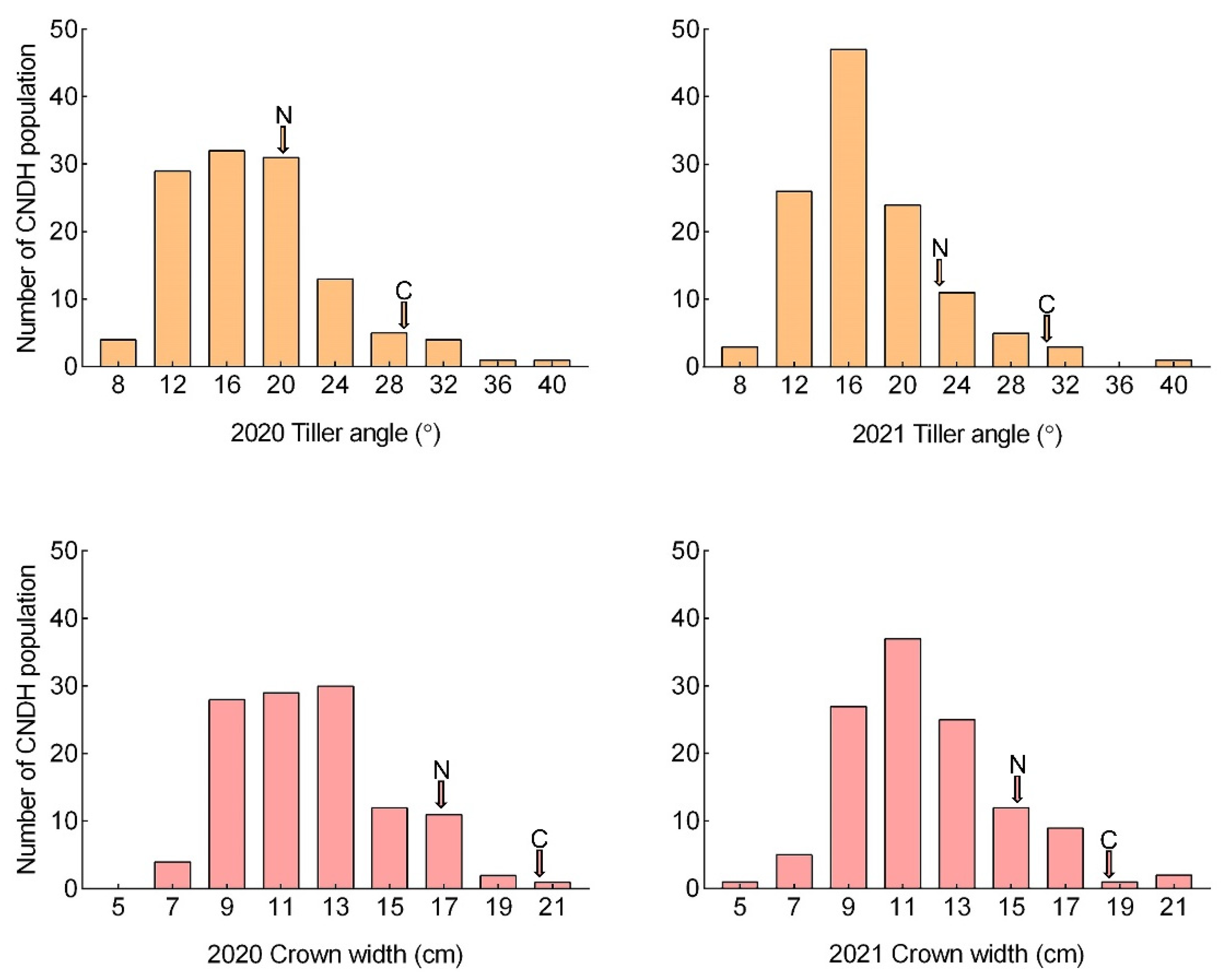
2.1. Phenotype Evaluation for Tiller Angle and Tiller Crown Width in CNDH Population
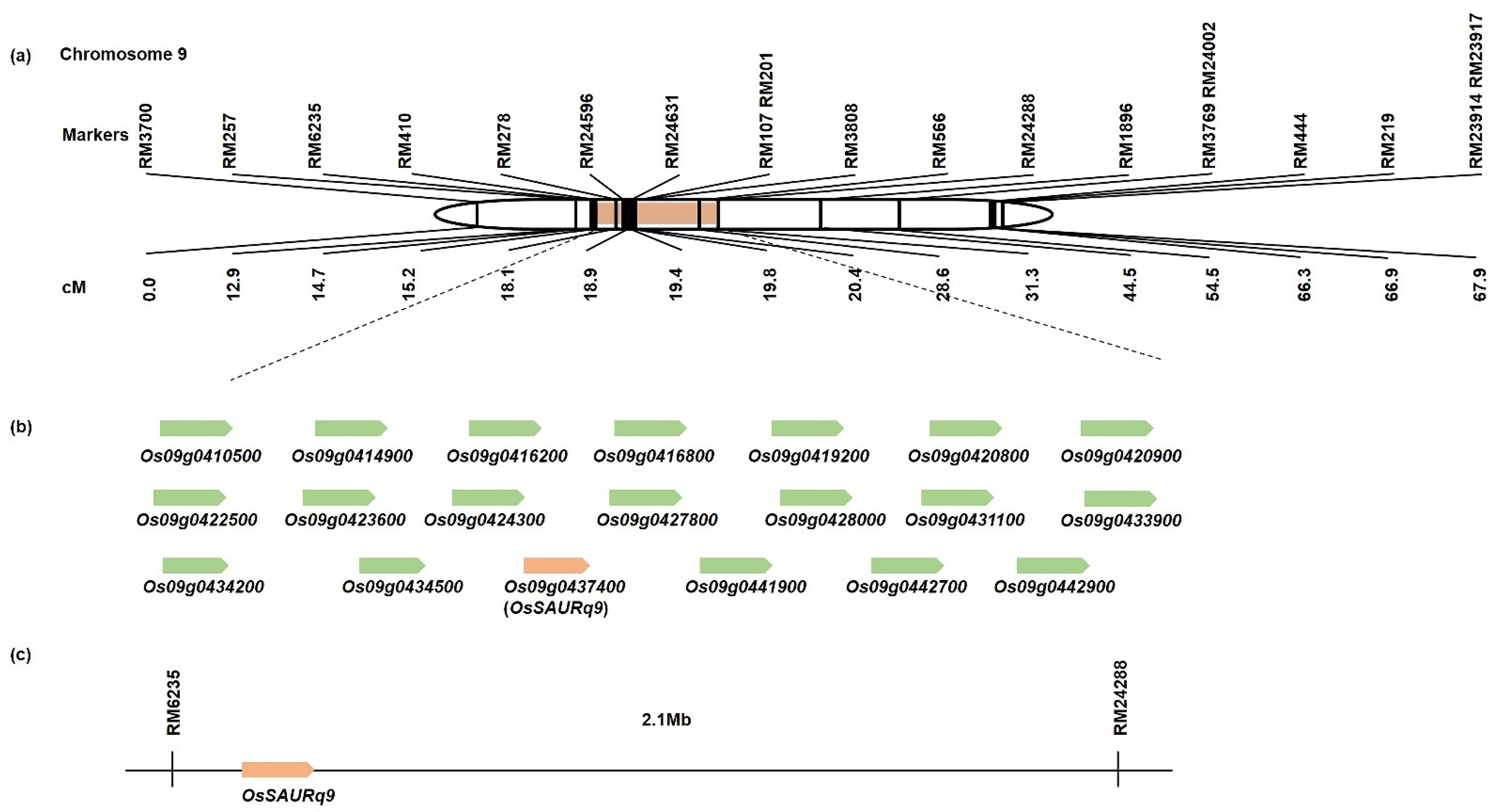
2.2. QTL Analysis Associated with Tiller Angle and Tiller Crown Width
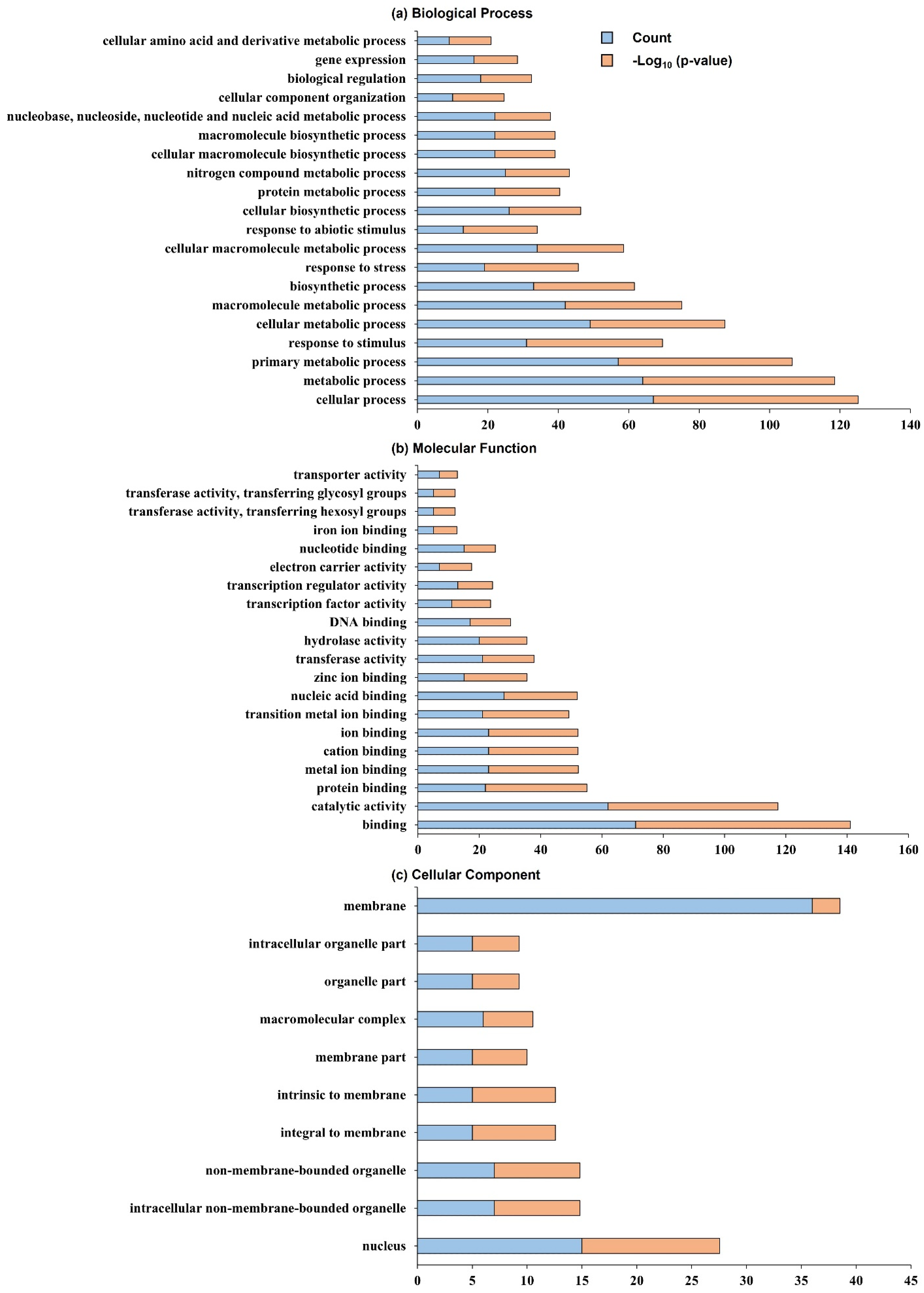
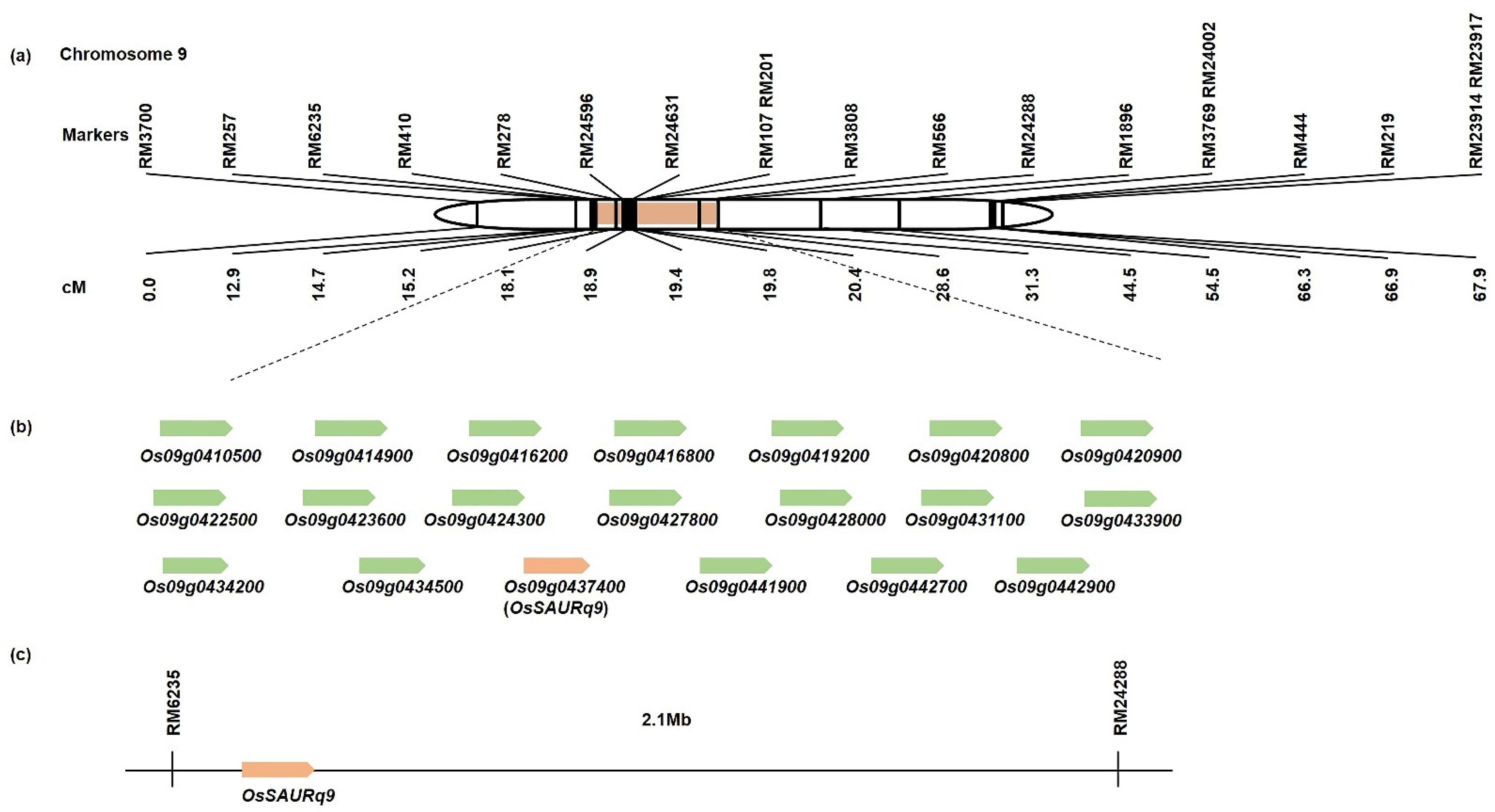
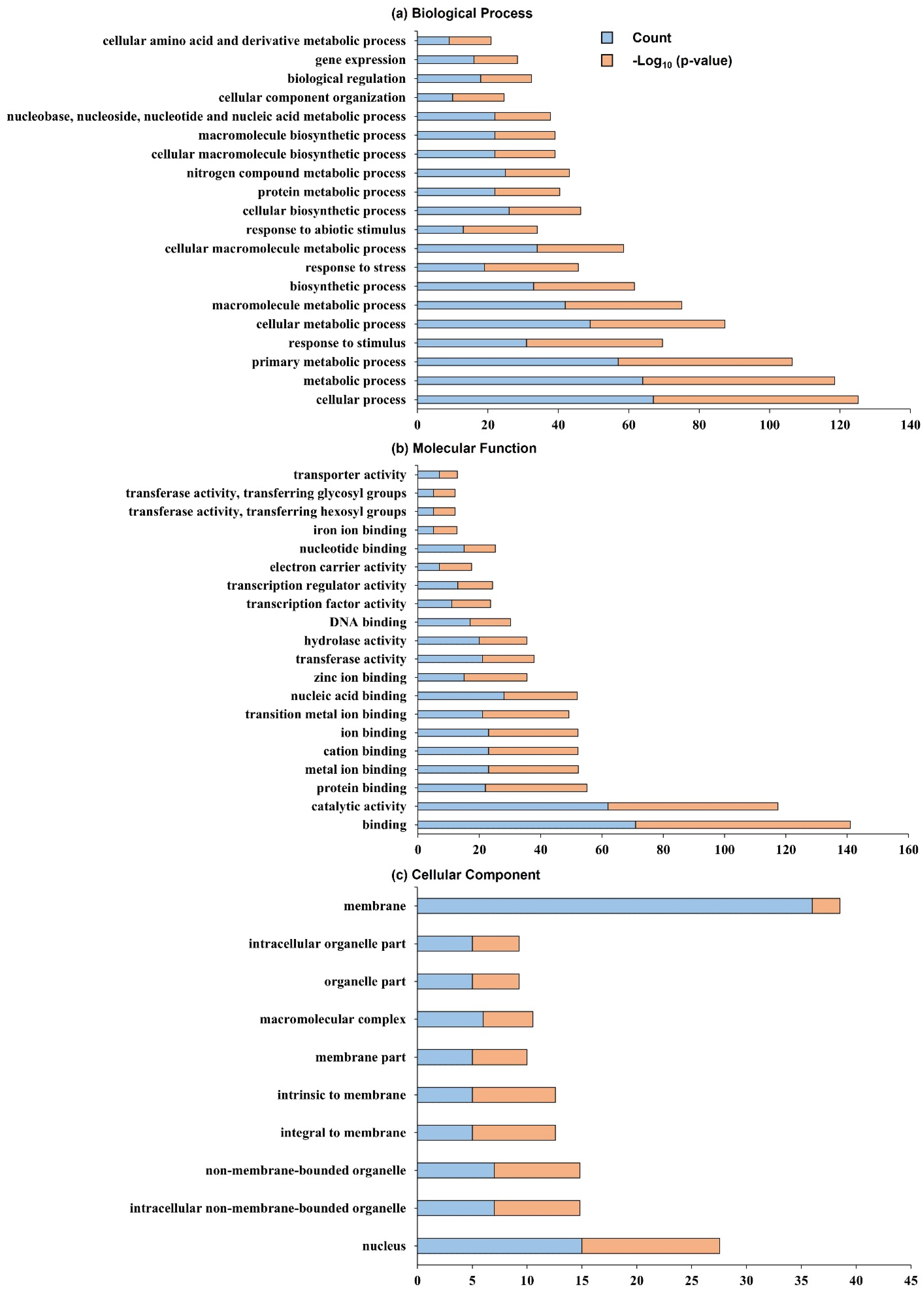
2.3. Search for Related Genes Associated with Tiller Angle and Tiller Crown Width Based on QTL Analysis
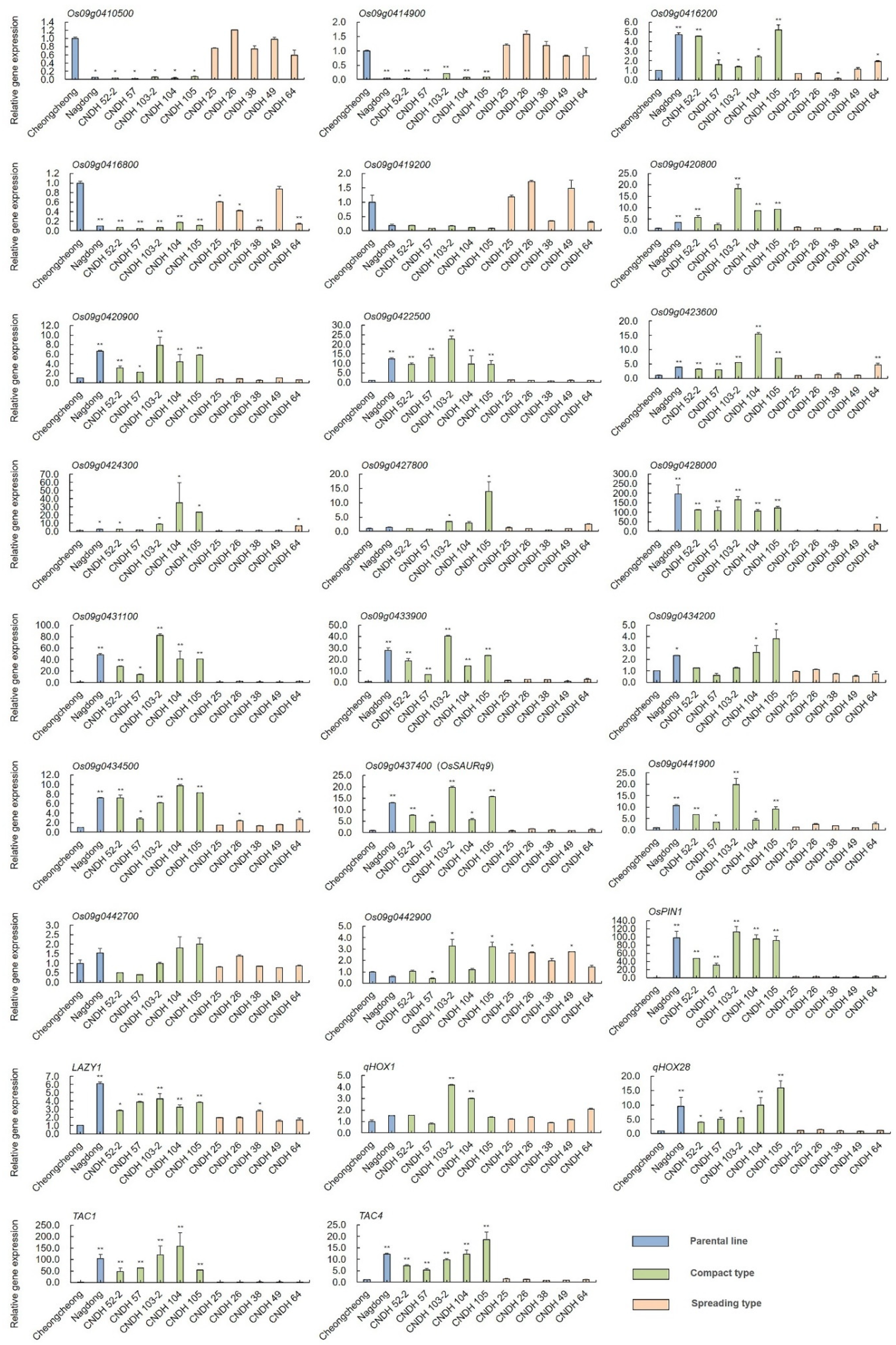
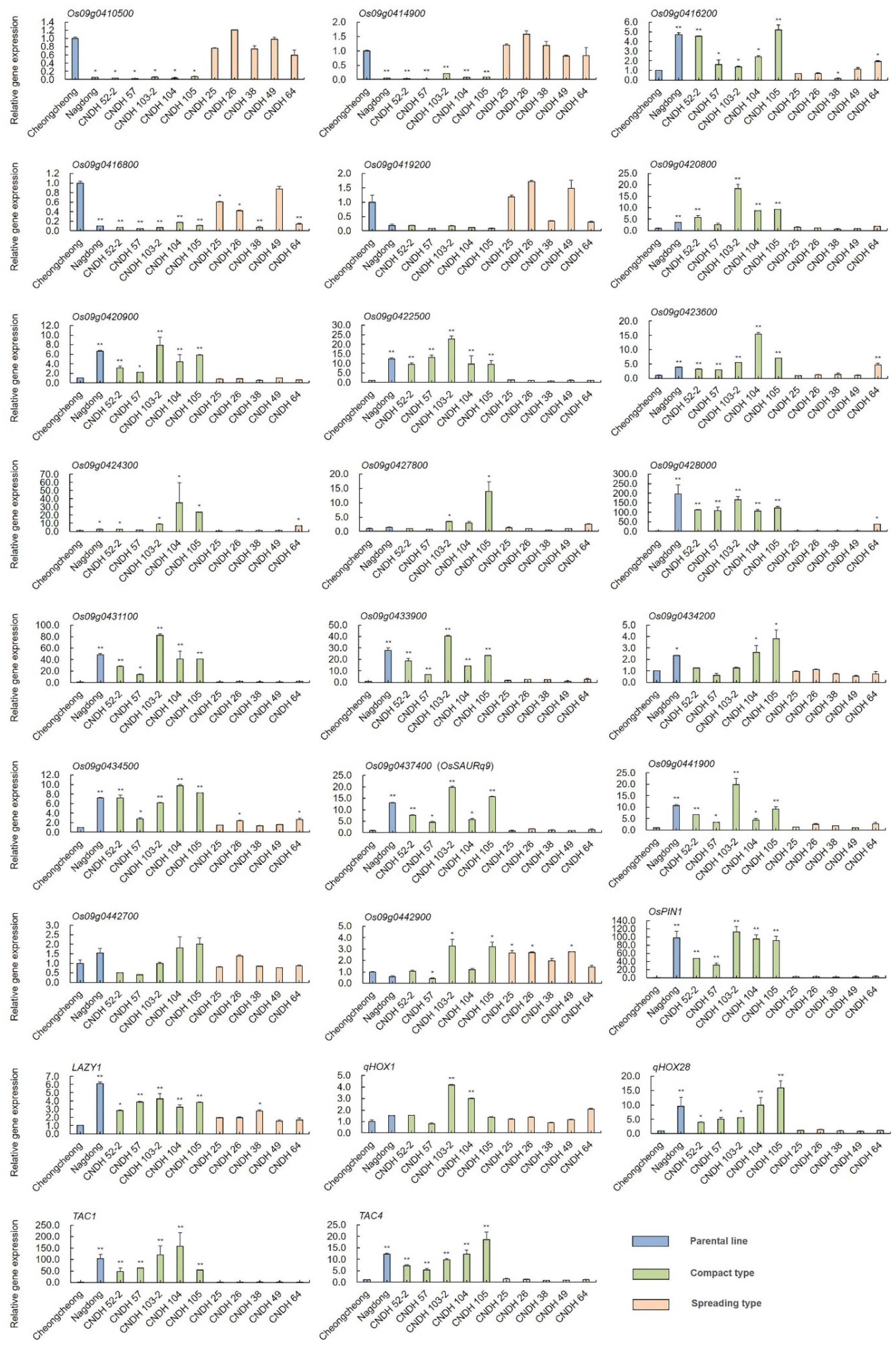
2.4. Selection of Related Genes and Comparison of Relative Expression Level of Other Tiller Angle-Related Genes
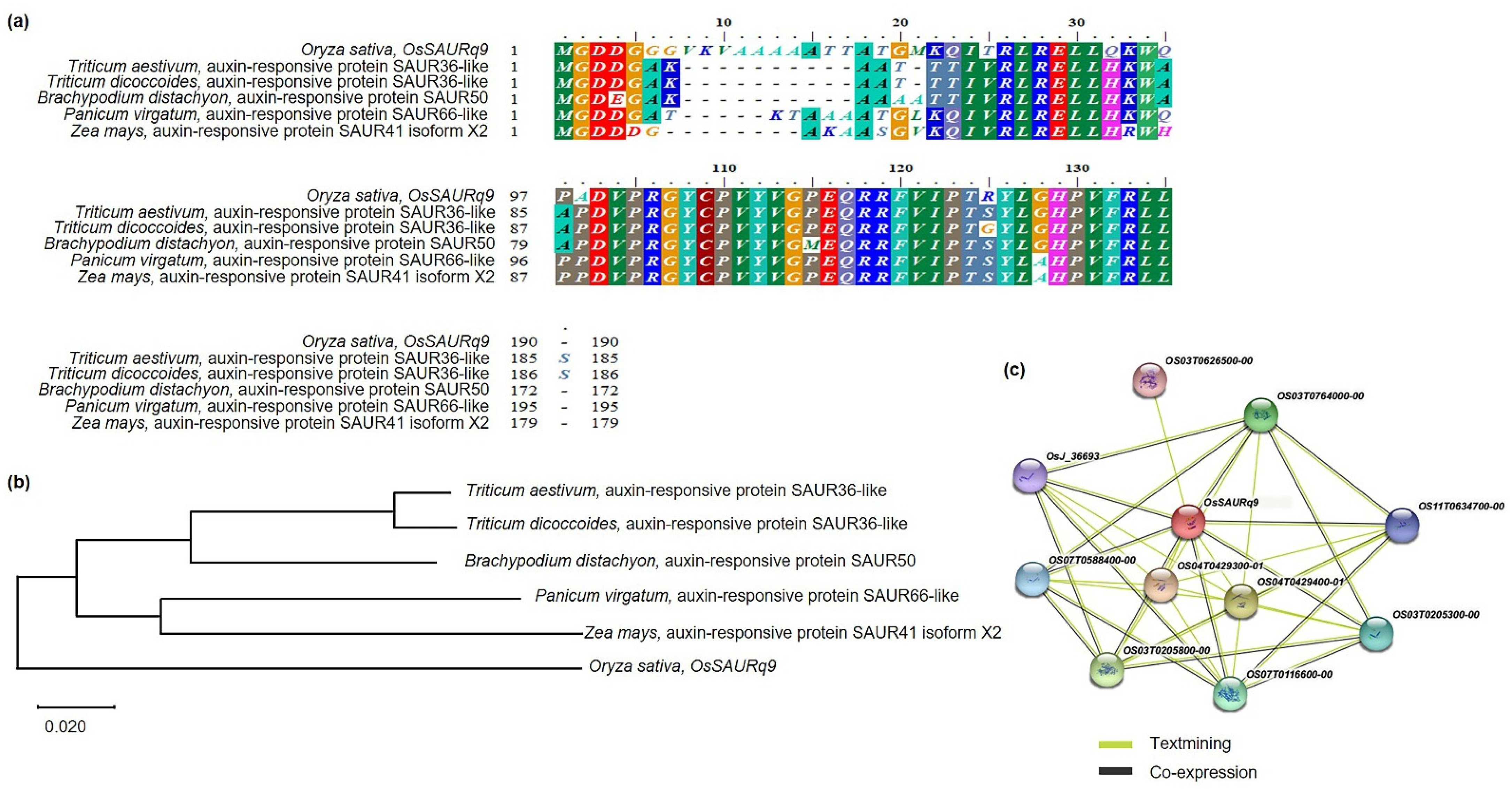
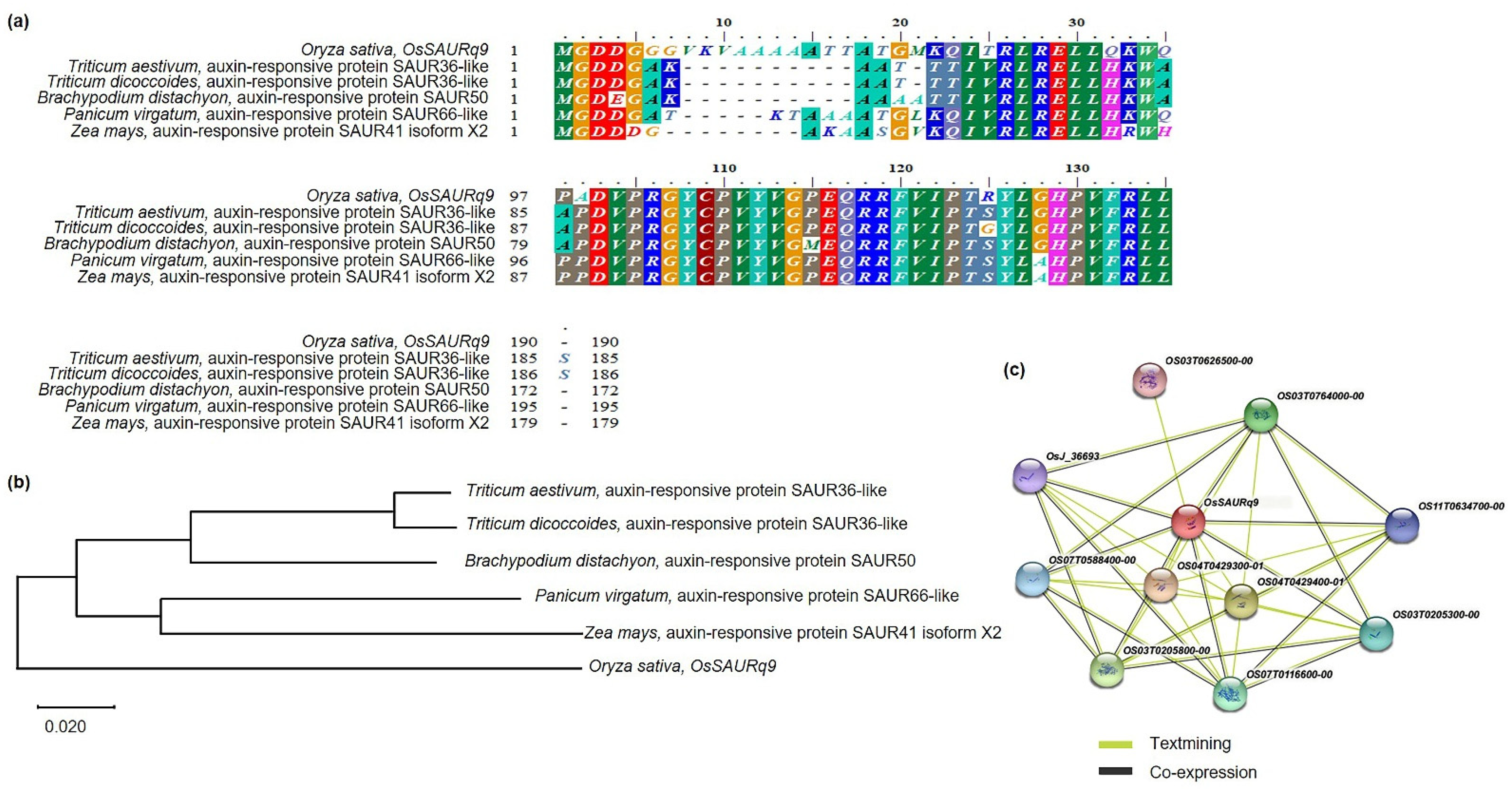
2.5. Phylogenetic Tree and Homology Sequence Analysis of OsSAURq9
3. Discussion
4. Materials and Methods
4.1. Plant Material and Experiment Design
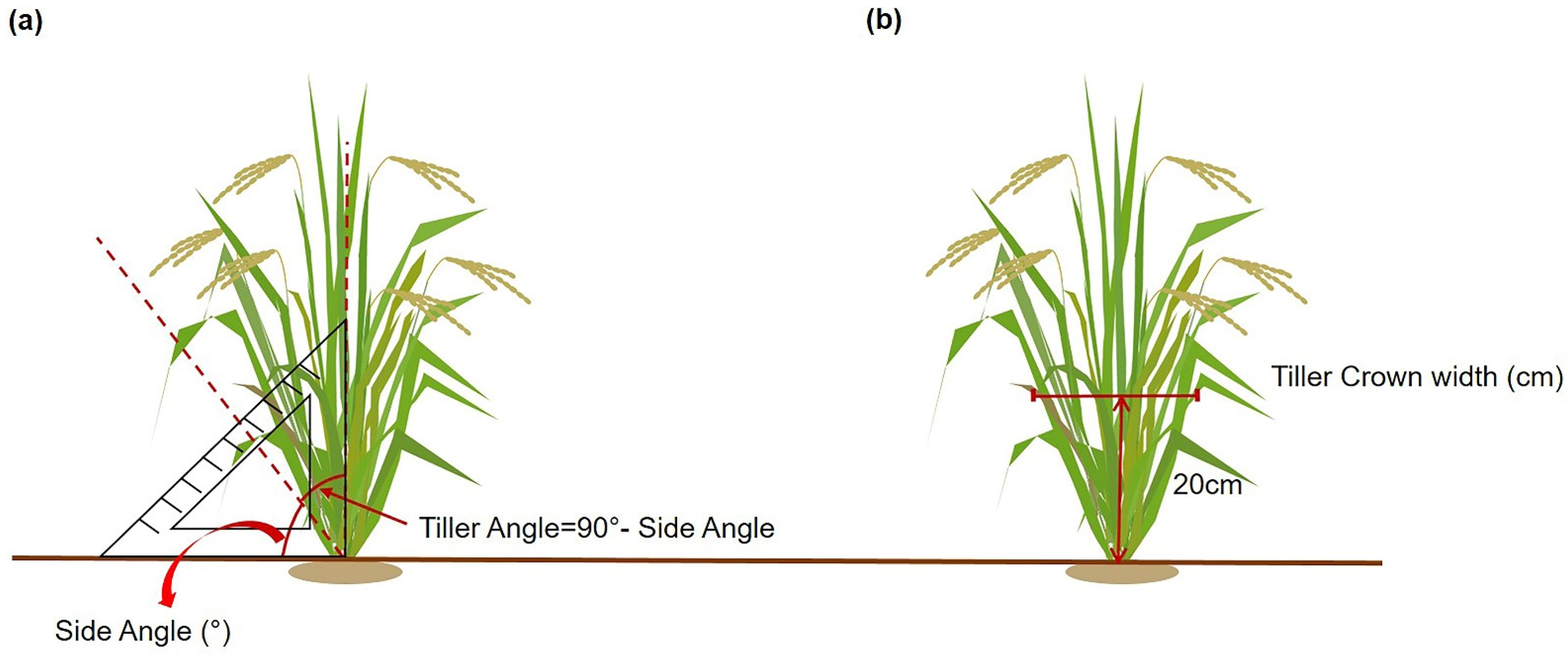
4.2. Measurement of Tiller Angle and Tiller Crown Width
4.3. Mapping Population and QTL Analysis for Tiller Angle and Tiller Crown Width
4.4. Prediction of the Related Genes
4.5. Expression Level of Related Genes
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, Y.; Li, J. Genes controlling plant architecture. Curr. Opin. Biotechnol. 2006, 17, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, J.; Chen, Z.; Wei, Y.; Qi, Y.; Wu, C. OsmiR167a-targeted auxin response factors modulate tiller angle via fine-tuning auxin distribution in rice. Plant Biotechnol. J. 2020, 18, 2015–2026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, H.; Zhao, H.; Xie, W.; Han, Z.; Li, G.; Yao, W.; Bai, X.; Hu, Y.; Guo, Z.; Lu, K.; et al. A Novel Tiller Angle Gene, TAC3, together with TAC1 and D2 Largely Determine the Natural Variation of Tiller Angle in Rice Cultivars. PLoS Genet. 2016, 12, e1006412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, P.; Jiang, L.; Yu, C.; Zhang, W.; Wang, J.; Wan, J. The Identification and Mapping of a Tiller Angle QTL on Rice Chromosome 9. Crop Sci. 2008, 48, 1799–1806. [Google Scholar] [CrossRef]
- Xu, Y.; McCouch, S.R.; Shen, Z. Transgressive Segregation of Tiller Angle in Rice Caused by Complementary Gene Action. Crop Sci. 1998, 38, 12–19. [Google Scholar] [CrossRef]
- Wang, Y.; Li, J. The Plant Architecture of Rice (Oryza sativa). Plant Mol. Biol. 2005, 59, 75–84. [Google Scholar] [CrossRef]
- He, J.; Shao, G.; Wei, X.; Huang, F.; Sheng, Z.; Tang, S.; Hu, P. Fine mapping and candidate gene analysis of qTAC8, a major quantitative trait locus controlling tiller angle in rice (Oryza sativa L.). PLoS ONE 2017, 12, e0178177. [Google Scholar] [CrossRef]
- Li, Z.; Paterson, A.H.; Pinson, S.R.M.; Stansel, J.W. RFLP facilitated analysis of tiller and leaf angles in rice (Oryza sativa L.). Euphytica 1999, 109, 79–84. [Google Scholar] [CrossRef]
- Yoshihara, T.; Iino, M. Identification of the Gravitropism-Related Rice Gene LAZY1 and Elucidation of LAZY1-Dependent and -Independent Gravity Signaling Pathways. Plant Cell Physiol. 2007, 48, 678–688. [Google Scholar] [CrossRef] [Green Version]
- Tan, L.; Li, X.; Liu, F.; Sun, X.; Li, C.; Zhu, Z.; Fu, Y.; Cai, H.; Wang, X.; Xie, D.; et al. Control of a key transition from prostrate to erect growth in rice domestication. Nat. Genet. 2008, 40, 1360–1364. [Google Scholar] [CrossRef]
- Yu, B.; Lin, Z.; Li, H.; Li, X.; Li, J.; Wang, Y.; Zhang, X.; Zhu, Z.; Zhai, W.; Wang, X.; et al. TAC1, a major quantitative trait locus controlling tiller angle in rice. Plant J. 2007, 52, 891–898. [Google Scholar] [CrossRef]
- Jiang, J.; Tan, L.; Zhu, Z.; Fu, Y.; Liu, F.; Cai, H.; Sun, C. Molecular Evolution of the TAC1 Gene from Rice (Oryza sativa L.). J. Genet. Genom. 2012, 39, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Li, S.; Fan, X.; Song, S.; Zhou, X.; Weng, X.; Xiao, J.; Li, X.; Xiong, L.; You, A.; et al. OsHOX1 and OsHOX28 Redundantly Shape Rice Tiller Angle by Reducing HSFA2D Expression and Auxin Content. Plant Physiol. 2020, 184, 1424–1437. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Sun, H.; Jiang, J.; Sun, X.; Tan, L.; Sun, C. TAC4 controls tiller angle by regulating the endogenous auxin content and distribution in rice. Plant Biotechnol. J. 2021, 19, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Zhu, L.; Shou, H.; Wu, P. A PIN1 Family Gene, OsPIN1, involved in Auxin-dependent Adventitious Root Emergence and Tillering in Rice. Plant Cell Physiol. 2005, 46, 1674–1681. [Google Scholar] [CrossRef]
- He, Y.; Li, L.; Jiang, D. Understanding the Regulatory Mechanisms of Rice Tiller Angle, Then and Now. Plant Mol. Biol. Report. 2021, 39, 640–647. [Google Scholar] [CrossRef]
- Zhang, N.; Yu, H.; Yu, H.; Cai, Y.; Huang, L.; Xu, C.; Xiong, G.; Meng, X.; Wang, J.; Chen, H.; et al. A Core Regulatory Pathway Controlling Rice Tiller Angle Mediated by the LAZY1-Dependent Asymmetric Distribution of Auxin. Plant Cell 2018, 30, 1461–1475. [Google Scholar] [CrossRef] [Green Version]
- Waite, J.M.; Dardick, C. TILLER ANGLE CONTROL 1 modulates plant architecture in response to photosynthetic signals. J. Exp. Bot. 2018, 69, 4935–4944. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Tan, L.; Sun, H.; Zhao, X.; Liu, F.; Cai, H.; Fu, Y.; Sun, X.; Gu, P.; Zhu, Z.; et al. Natural Variations at TIG1 Encoding a TCP Transcription Factor Contribute to Plant Architecture Domestication in Rice. Mol. Plant 2019, 12, 1075–1089. [Google Scholar] [CrossRef]
- Jin, J.; Huang, W.; Gao, J.-P.; Yang, J.; Shi, M.; Zhu, M.-Z.; Luo, D.; Lin, H.-X. Genetic control of rice plant architecture under domestication. Nat. Genet. 2008, 40, 1365–1369. [Google Scholar] [CrossRef]
- Wang, Y.; Li, J. Rice, rising. Nat. Genet. 2008, 40, 1273–1275. [Google Scholar] [CrossRef] [PubMed]
- Xu, S. Mapping Quantitative Trait Loci by Controlling Polygenic Background Effects. Genetics 2013, 195, 1209–1222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, A.; Courtois, B. Mapping QTLs associated with drought resistance in rice: Progress, problems and prospects. Plant Growth Regul. 1999, 29, 123–133. [Google Scholar] [CrossRef]
- Hagen, G.; Guilfoyle, T. Auxin-responsive gene expression: Genes, promoters and regulatory factors. Plant Mol. Biol. 2002, 49, 373–385. [Google Scholar] [CrossRef]
- Kant, S.; Bi, Y.-M.; Zhu, T.; Rothstein, S.J. SAUR39, a Small Auxin-Up RNA Gene, Acts as a Negative Regulator of Auxin Synthesis and Transport in Rice. Plant Physiol. 2009, 151, 691–701. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.-X.; Xiao, M.-Z.; Liu, Y.; Fu, J.-L.; He, Y.; Jiang, D.-A. The small auxin-up RNA OsSAUR45 affects auxin synthesis and transport in rice. Plant Mol. Biol. 2017, 94, 97–107. [Google Scholar] [CrossRef]
- Li, P.; Wang, Y.; Qian, Q.; Fu, Z.; Wang, M.; Zeng, D.; Li, B.; Wang, X.; Li, J. LAZY1 controls rice shoot gravitropism through regulating polar auxin transport. Cell Res. 2007, 17, 402–410. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Xu, Z.-F. Ectopic Overexpression of an AUXIN/INDOLE-3-ACETIC ACID (Aux/IAA) Gene OsIAA4 in Rice Induces Morphological Changes and Reduces Responsiveness to Auxin. Int. J. Mol. Sci. 2013, 14, 13645–13656. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.; Yang, Y.; Xu, E.; Ge, X.; Xiang, Y.; Li, Z. Novel and major QTL for branch angle detected by using DH population from an exotic introgression in rapeseed (Brassica napus L.). Theor. Appl. Genet. 2018, 131, 67–78. [Google Scholar] [CrossRef]
- Yun, B.-W.; Kim, M.-G.; Handoyo, T.; Kim, K.-M. Analysis of rice grain quality-associated quantitative trait loci by using genetic mapping. Am. J. Plant Sci. 2014, 5, 1125–1132. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Gao, H.; Liang, Y.; Li, J.; Wang, Y. Molecular basis underlying rice tiller angle: Current progress and future perspectives. Mol. Plant 2022, 15, 125–137. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.; Kang, Y.S.; Lee, Y.K.; Koh, H.-J. Evaluating Multiple Allelic Combination to Determine Tiller Angle Variation in Rice. Agriculture 2020, 10, 428. [Google Scholar] [CrossRef]
- Kashiwagi, T.; Togawa, E.; Hirotsu, N.; Ishimaru, K. Improvement of lodging resistance with QTLs for stem diameter in rice (Oryza sativa L.). Theor. Appl. Genet. 2008, 117, 749–757. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.; Kim, K.; Manigbas, N.L.; Yi, G.; Sohn, J. Identification of quantitative trait loci for resistance to white-backed planthopper (Sogatella furcifera) in rice with Milyang 46 (Cheongcheongbyeo) background. Philipp. J. Crop Sci. 2013, 38, 30–36. [Google Scholar]
- Lincoln, S.E.; Daly, M.J.; Lander, E.S. Constructing Genetic Linkage Maps with MAPMAKER/EXP Version 3.0: A Tutorial and Reference Manual. A Whitehead Institute for Biomedical Research Technical Report, 3rd ed.; Whitehead Institute: Cambridge, MA, USA, 1993. [Google Scholar]
- Zeng, Z.B. Precision mapping of quantitative trait loci. Genetics 1994, 136, 1457–1468. [Google Scholar] [CrossRef] [PubMed]
- McCough, S.R.; Doerge, R.W. QTL mapping in rice. Trends Genet. 1995, 11, 482–487. [Google Scholar] [CrossRef]
- Sato, Y.; Takehisa, H.; Kamatsuki, K.; Minami, H.; Namiki, N.; Ikawa, H.; Ohyanagi, H.; Sugimoto, K.; Antonio, B.A.; Nagamura, Y. RiceXPro Version 3.0: Expanding the informatics resource for rice transcriptome. Nucleic Acids Res. 2013, 41, D1206–D1213. [Google Scholar] [CrossRef] [Green Version]
- Du, Z.; Zhou, X.; Ling, Y.; Zhang, Z.; Su, Z. agriGO: A GO analysis toolkit for the agricultural community. Nucleic Acids Res. 2010, 38, W64–W70. [Google Scholar] [CrossRef] [Green Version]
- Hall, T. BioEdit, version 7.0.; Informer Technologies Inc.: Los Angeles, CA, USA, 2004; Available online: http://www.mbio.ncsu.edu/BioEdit/bioedit.html (accessed on 10 April 2022).
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein–protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, D.-D.; Jang, Y.-H.; Farooq, M.; Park, J.-R.; Kim, E.-G.; Du, X.-X.; Jan, R.; Kim, K.-H.; Lee, S.I.; Lee, G.-S.; et al. Identification of a Major QTL and Validation of Related Genes for Tiller Angle in Rice Based on QTL Analysis. Int. J. Mol. Sci. 2022, 23, 5192. https://doi.org/10.3390/ijms23095192
Zhao D-D, Jang Y-H, Farooq M, Park J-R, Kim E-G, Du X-X, Jan R, Kim K-H, Lee SI, Lee G-S, et al. Identification of a Major QTL and Validation of Related Genes for Tiller Angle in Rice Based on QTL Analysis. International Journal of Molecular Sciences. 2022; 23(9):5192. https://doi.org/10.3390/ijms23095192
Chicago/Turabian StyleZhao, Dan-Dan, Yoon-Hee Jang, Muhammad Farooq, Jae-Ryoung Park, Eun-Gyeong Kim, Xiao-Xuan Du, Rahmatullah Jan, Kyung-Hwan Kim, Soo In Lee, Gang-Seob Lee, and et al. 2022. "Identification of a Major QTL and Validation of Related Genes for Tiller Angle in Rice Based on QTL Analysis" International Journal of Molecular Sciences 23, no. 9: 5192. https://doi.org/10.3390/ijms23095192
APA StyleZhao, D.-D., Jang, Y.-H., Farooq, M., Park, J.-R., Kim, E.-G., Du, X.-X., Jan, R., Kim, K.-H., Lee, S. I., Lee, G.-S., & Kim, K.-M. (2022). Identification of a Major QTL and Validation of Related Genes for Tiller Angle in Rice Based on QTL Analysis. International Journal of Molecular Sciences, 23(9), 5192. https://doi.org/10.3390/ijms23095192