
Cortactin in Lung Cell Function and Disease


Abstract
1. Introduction
1.1. Structure and Function of Cortactin
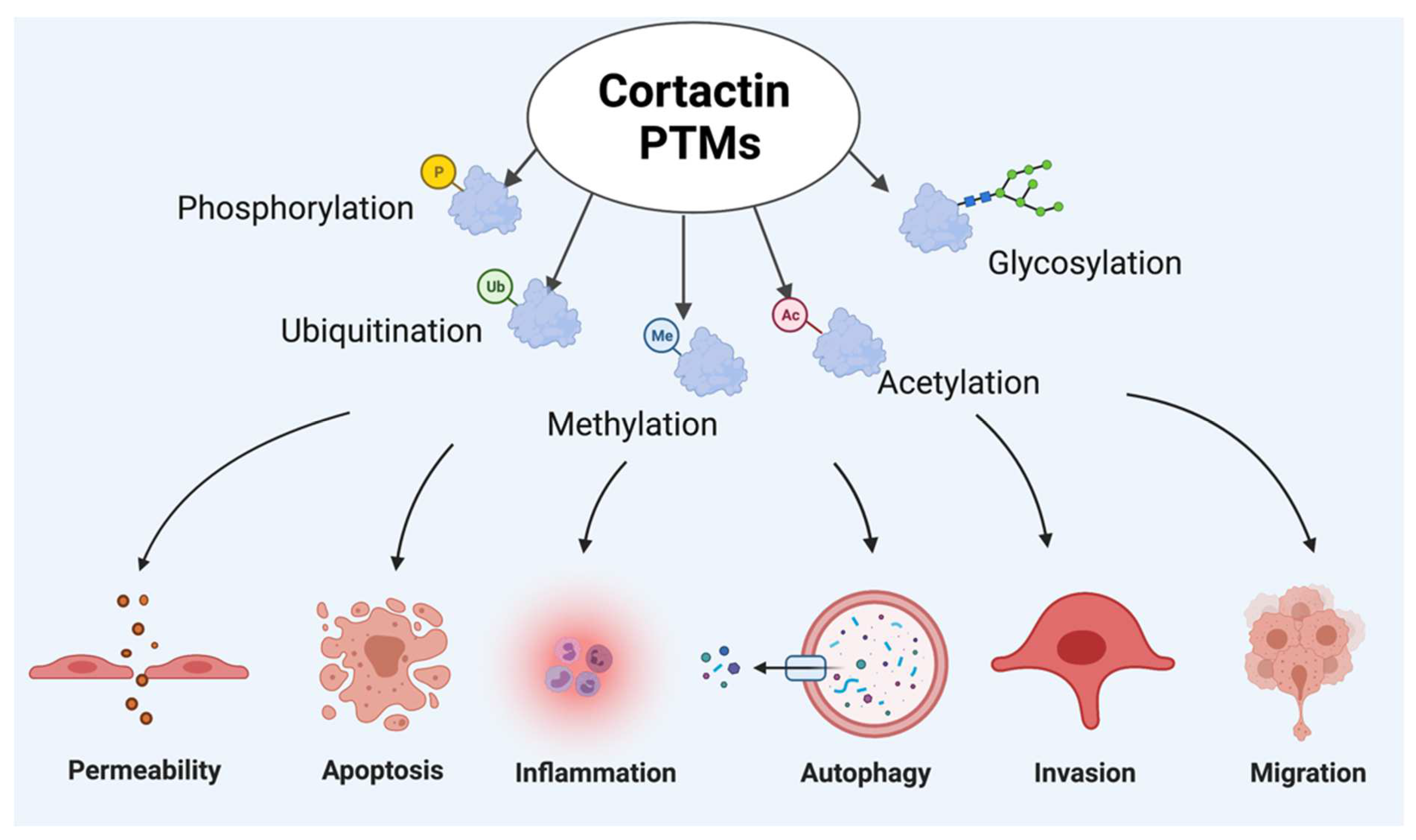
1.2. Post Translational Modifications of Cortactin
1.2.1. Phosphorylation and Dephosphorylation of Cortactin
1.2.2. Ubiquitination and Deubiquitination of Cortactin
1.2.3. Acetylation and Deacetylation of Cortactin
1.2.4. Glycosylation of CTTN
2. Vascular Endothelium and Epithelium
2.1. Vascular Endothelium
2.1.1. Cortactin and Other Cytoskeleton Proteins in Endothelial Barrier Regulation
2.1.2. Cortactin in Cellular Migration
2.1.3. Cortactin and Reactive Oxygen Species
2.1.4. Cortactin in Endothelial Apoptosis
2.2. Airway Epithelium
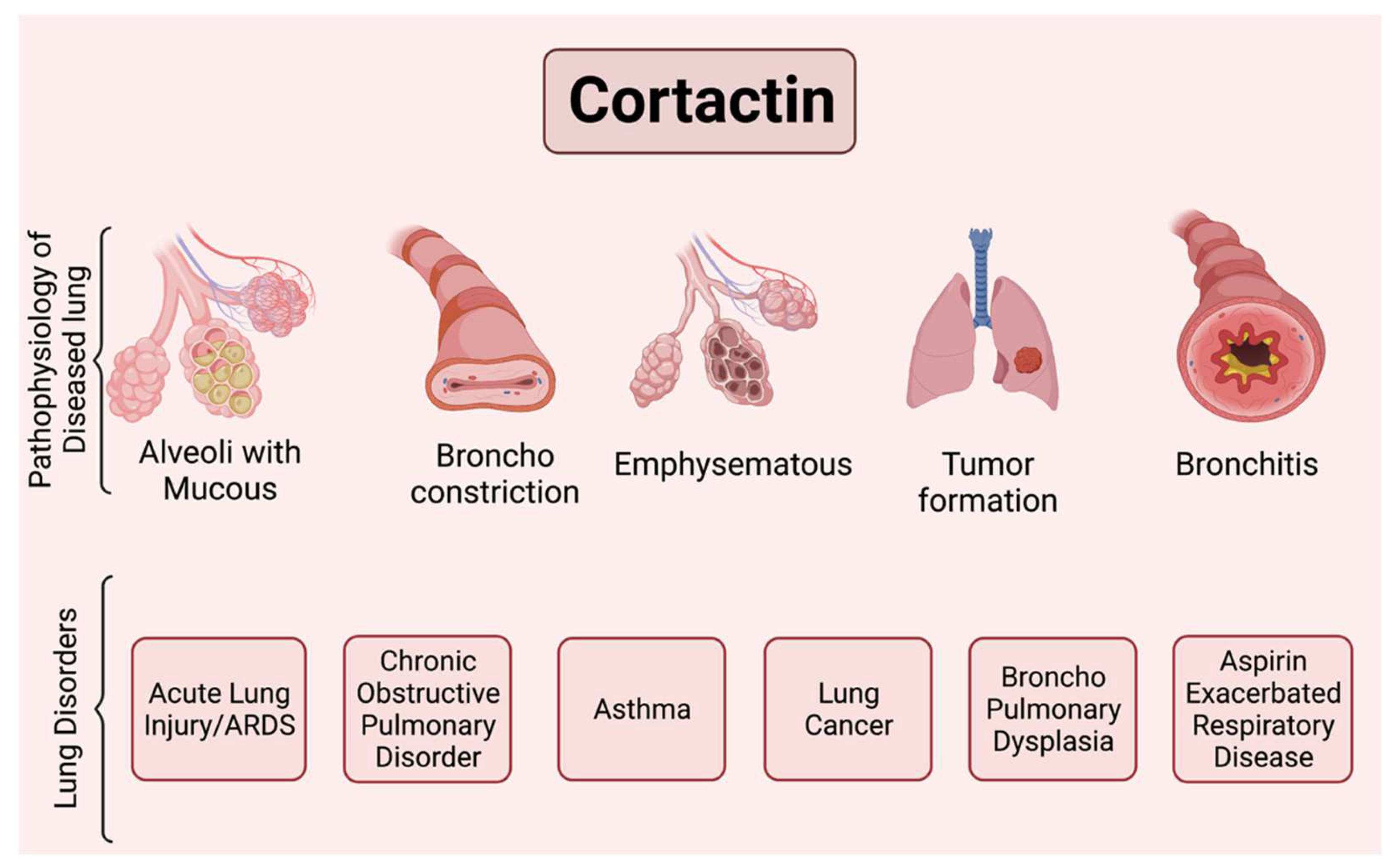
Cortactin in the Airway
3. Acute Lung Injury
3.1. Pathophysiology of Acute Respiratory Distress Syndrome and Acute Lung Injury
3.2. Role of Actin Cytoskeletal Rearrangement in Lung Endothelial Barrier Regulation
3.3. Role of Prostaglandin A2-EP4 in ALI
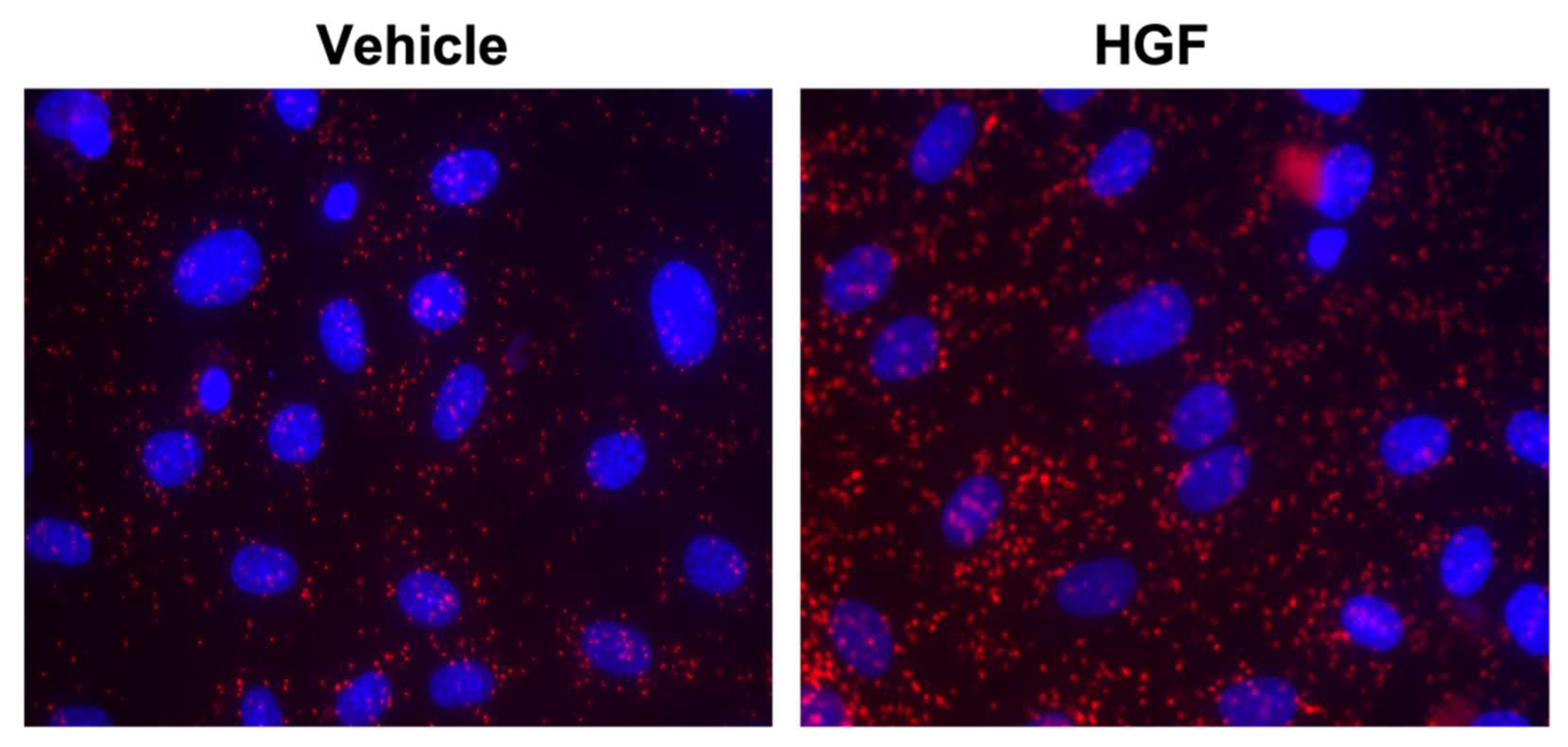
3.4. Role of Asef in Cytoskeleton Remodeling in HGF-Mediated ALI
3.5. Polyethylene Glycol in Actin Cytoskeleton Rearrangement
3.6. Cytoskeletal Activation and Altered Gene Expression Regulated by Simvastatin
3.7. FTY720-S1P1 Receptor in Barrier Enhancement
3.8. Cortactin Genetic Variant in ARDS
4. Chronic Obstructive Pulmonary Disease (COPD)
4.1. Pathophysiology of COPD
4.2. CS-Induced Responses in COPD
4.3. Role of CTTN in Cigarette Smoke-Induced Cell Migration and Invasion
4.4. CTTN Methylation in COPD
5. Asthma and Other Lung Disorders
5.1. Pathophysiology of Asthma and CTTN Gene Variation as a Key Regulator of Cytoskeleton Rearrangements
5.2. CTTN PTMs in Pathogenesis of Asthma
5.2.1. Cortactin in Association with Profilin in Pathogenesis of Asthma
5.2.2. Cortactin in Aspirin-Exacerbated Respiratory Disease
5.2.3. Deacetylation of Cortactin
5.2.4. Shear Stress in Cortactin and Actin Polymerization
5.3. Cortactin in Broncho Pulmonary Dysplasia
5.4. Cortactin in Lung Cancer
5.5. Role of Host Cytoskeleton in Coronavirus Infection
6. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schliwa, M.; Pryzwansky, K.B.; van Blerkom, J. Implications of cytoskeletal interactions for cellular architecture and behavior. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1982, 299, 199–205. [Google Scholar] [PubMed]
- Schnoor, M.; Stradal, T.E.; Rottner, K. Cortactin: Cell Functions of A Multifaceted Actin-Binding Protein. Trends Cell Biol. 2018, 28, 79–98. [Google Scholar] [CrossRef] [PubMed]
- Schaks, M.; Giannone, G.; Rottner, K. Actin dynamics in cell migration. Essays Biochem. 2019, 63, 483–495. [Google Scholar] [CrossRef] [PubMed]
- Weed, S.A.; Parsons, J.T. Cortactin: Coupling membrane dynamics to cortical actin assembly. Oncogene 2001, 20, 6418–6434. [Google Scholar] [CrossRef] [PubMed]
- Machesky, L.; Hall, A. Role of Actin Polymerization and Adhesion to Extracellular Matrix in Rac- and Rho-induced Cytoskeletal Reorganization. J. Cell Biol. 1997, 138, 913–926. [Google Scholar] [CrossRef] [PubMed]
- Delaguillaumie, A.; Lagaudrière-Gesbert, C.; Popoff, M.R.; Conjeaud, H. Rho GTPases link cytoskeletal rearrangements and activation processes induced via the tetraspanin CD82 in T lymphocytes. J. Cell Sci. 2002, 115, 433–443. [Google Scholar] [CrossRef] [PubMed]
- Bustelo, X.R.; Sauzeau, V.; Berenjeno, I.M. GTP-binding proteins of the Rho/Rac family: Regulation, effectors and functions in vivo. BioEssays 2007, 29, 356–370. [Google Scholar] [CrossRef]
- Jeannot, P.; Besson, A. Cortactin function in invadopodia. Small GTPases 2017, 11, 256–270. [Google Scholar] [CrossRef]
- Weed, S.A.; Karginov, A.V.; Schafer, D.A.; Weaver, A.M.; Kinley, A.W.; Cooper, J.A.; Parsons, J.T. Cortactin localization to sites of actin assembly in lamellipodia requires interactions with F-actin and the Arp2/3 complex. J. Cell Biol. 2000, 151, 29–40. [Google Scholar] [CrossRef]
- Van Rossum, A.G.S.H.; Schuuring-Scholtes, E.; Seggelen, V.V.B.-V.; Kluin, P.M.; Schuuring, E. Comparative genome analysis of cortactin and HS1: The significance of the F-actin binding repeat domain. BMC Genom. 2005, 6, 15. [Google Scholar] [CrossRef]
- Ammer, A.G.; Weed, S.A. Cortactin branches out: Roles in regulating protrusive actin dynamics. Cell Motil. Cytoskelet. 2008, 65, 687–707. [Google Scholar] [CrossRef] [PubMed]
- Usatyuk, P.V.; Fu, P.; Mohan, V.; Epshtein, Y.; Jacobson, J.R.; Gomez-Cambronero, J.; Wary, K.K.; Bindokas, V.; Dudek, S.M.; Salgia, R.; et al. Role of c-Met/phosphatidylinositol 3-kinase (PI3k)/Akt signaling in hepatocyte growth factor (HGF)-mediated lamellipodia formation, reactive oxygen species (ROS) generation, and motility of lung endothelial cells. J. Biol. Chem. 2014, 289, 13476–13491. [Google Scholar] [CrossRef] [PubMed]
- Dudek, S.M.; Jacobson, J.R.; Chiang, E.T.; Birukov, K.G.; Wang, P.; Zhan, X.; Garcia, J.G. Pulmonary endothelial cell barrier enhancement by sphingosine 1-phosphate: Roles for cortactin and myosin light chain kinase. J. Biol. Chem. 2004, 279, 24692–24700. [Google Scholar] [CrossRef]
- Dudek, S.M.; Birukov, K.G.; Zhan, X.; Garcia, J.G. Novel interaction of cortactin with endothelial cell myosin light chain kinase. Biochem. Biophys. Res. Commun. 2002, 298, 511–519. [Google Scholar] [CrossRef]
- MacGillavry, H.D.; Kerr, J.M.; Kassner, J.; Frost, N.; Blanpied, T.A. Shank-cortactin interactions control actin dynamics to maintain flexibility of neuronal spines and synapses. Eur. J. Neurosci. 2015, 43, 179–193. [Google Scholar] [CrossRef] [PubMed]
- Hou, P.; Estrada, L.; Kinley, A.W.; Parsons, J.T.; Vojtek, A.B.; Gorski, J.L. Fgd1, the Cdc42 GEF responsible for Faciogenital Dysplasia, directly interacts with cortactin and mAbp1 to modulate cell shape. Hum. Mol. Genet. 2003, 12, 1981–1993. [Google Scholar] [CrossRef]
- Gifford, S.M.; Liu, W.; Mader, C.C.; Halo, T.L.; Machida, K.; Boggon, T.J.; Koleske, A.J. Two Amino Acid Residues Confer Different Binding Affinities of Abelson Family Kinase Src Homology 2 Domains for Phosphorylated Cortactin. J. Biol. Chem. 2014, 289, 19704–19713. [Google Scholar] [CrossRef]
- Hattan, D.; Nesti, E.; Cachero, T.G.; Morielli, A.D. Tyrosine Phosphorylation of Kv1.2 Modulates Its Interaction with the Actin-binding Protein Cortactin. J. Biol. Chem. 2002, 277, 38596–38606. [Google Scholar] [CrossRef]
- Lazzarino, G.; Corsico, N.; Tavazzi, B.; Di Pierro, D.; Arrigoni-Martelli, E.; Giardina, B. L-propionylcarnitine taurine amide induces the metabolic recovery of the isolated postischemic rat heart. Cardiologia 1992, 37, 715–717. [Google Scholar]
- Brown, M.T.; Andrade, J.; Radhakrishna, H.; Donaldson, J.G.; Cooper, J.A.; Randazzo, P.A. ASAP1, a Phospholipid-Dependent Arf GTPase-Activating Protein That Associates with and Is Phosphorylated by Src. Mol. Cell. Biol. 1998, 18, 7038–7051. [Google Scholar] [CrossRef]
- Weigand, D.A. Transfollicular Extrusion of Sebaceous Glands: Natural Phenomenon or Artifact? A Case Report. J. Cutan. Pathol. 1976, 3, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Hirooka, S.; Akashi, T.; Ando, N.; Suzuki, Y.; Ishida, N.; Kurata, M.; Takizawa, T.; Kayamori, K.; Sakamoto, K.; Fujiwara, N.; et al. Localization of the Invadopodia-Related Proteins Actinin-1 and Cortactin to Matrix-Contact-Side Cytoplasm of Cancer Cells in Surgically Resected Lung Adenocarcinomas. Pathobiology 2011, 78, 10–23. [Google Scholar] [CrossRef] [PubMed]
- Weaver, A.M.; Heuser, J.E.; Karginov, A.V.; Lee, W.L.; Parsons, J.T.; Cooper, J.A. Interaction of cortactin and N-WASp with Arp2/3 complex. Curr. Biol. 2002, 12, 1270–1278. [Google Scholar] [CrossRef]
- Li, Y.; Uruno, T.; Haudenschild, C.; Dudek, S.M.; Garcia, J.G.; Zhan, X. Interaction of cortactin and Arp2/3 complex is required for sphingosine-1-phosphate-induced endothelial cell remodeling. Exp. Cell Res. 2004, 298, 107–121. [Google Scholar] [CrossRef]
- Hajdu, P.; Martin, G.V.; Chimote, A.A.; Szilagyi, O.; Takimoto, K.; Conforti, L. The C-terminus SH3-binding domain of Kv1.3 is required for the actin-mediated immobilization of the channel via cortactin. Mol. Biol. Cell 2015, 26, 1640–1651. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-R.; Kori, R.; John, B.; Tan, T.-H. Caspase-Mediated Cleavage of Actin-Binding and SH3-Domain-Containing Proteins Cortactin, HS1, and HIP-55 during Apoptosis. Biochem. Biophys. Res. Commun. 2001, 288, 981–989. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.-Y.; Husain, M. Caspase-Mediated Cleavage of Human Cortactin during Influenza A Virus Infection Occurs in Its Actin-Binding Domains and Is Associated with Released Virus Titres. Viruses 2020, 12, 87. [Google Scholar] [CrossRef]
- Hashimoto, S.; Hirose, M.; Hashimoto, A.; Morishige, M.; Yamada, A.; Hosaka, H.; Akagi, K.I.; Ogawa, E.; Oneyama, C.; Agatsuma, T.; et al. Targeting AMAP1 and cortactin binding bearing an atypical src homology 3/proline interface for prevention of breast cancer invasion and metastasis. Proc. Natl. Acad. Sci. USA 2006, 103, 7036–7041. [Google Scholar] [CrossRef]
- Oneyama, C.; Nakano, H.; Sharma, S.V. UCS15A, a novel small molecule, SH3 domain-mediated protein-protein interaction blocking drug. Oncogene 2002, 21, 2037–2050. [Google Scholar] [CrossRef]
- Kenney, S.P.; Meng, X.-J. An SH3 binding motif within the nucleocapsid protein of porcine reproductive and respiratory syndrome virus interacts with the host cellular signaling proteins STAMI, TXK, Fyn, Hck, and cortactin. Virus Res. 2015, 204, 31–39. [Google Scholar] [CrossRef]
- Snyder, B.N.; Cho, Y.; Qian, Y.; Coad, J.E.; Flynn, D.C.; Cunnick, J.M. AFAP1L1 is a novel adaptor protein of the AFAP family that interacts with cortactin and localizes to invadosomes. Eur. J. Cell Biol. 2011, 90, 376–389. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Chen, L.; McClafferty, H.; Sailer, C.A.; Ruth, P.; Knaus, H.; Shipston, M. A noncanonical SH3 domain binding motif links BK channels to the actin cytoskeleton via the SH3 adapter cortactin. FASEB J. 2006, 20, 2588–2590. [Google Scholar] [CrossRef] [PubMed]
- Lynch, D.K.; Winata, S.C.; Lyons, R.J.; Hughes, W.; Lehrbach, G.M.; Wasinger, V.; Corthals, G.; Cordwell, S.; Daly, R. A Cortactin-CD2-associated Protein (CD2AP) Complex Provides a Novel Link between Epidermal Growth Factor Receptor Endocytosis and the Actin Cytoskeleton. J. Biol. Chem. 2003, 278, 21805–21813. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Bruck, S.; Cemerski, S.; Zhang, L.; Butler, B.; Dani, A.; Cooper, J.; Shaw, A.S. CD2AP Links Cortactin and Capping Protein at the Cell Periphery To Facilitate Formation of Lamellipodia. Mol. Cell. Biol. 2013, 33, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Adams, G.; Zhou, J.; Wang, W.; Wu, H.; Quan, J.; Liu, Y.; Xia, P.; Wang, Z.; Zhou, S.; Jiang, J.; et al. The Microtubule Plus End Tracking Protein TIP150 Interacts with Cortactin to Steer Directional Cell Migration. J. Biol. Chem. 2016, 291, 20692–20706. [Google Scholar] [CrossRef]
- Ohoka, Y.; Takai, Y. Isolation and characterization of cortactin isoforms and a novel cortactin-binding protein, CBP90. Genes Cells 1998, 3, 603–612. [Google Scholar] [CrossRef]
- Kim, L.; Wong, T.W. Growth Factor-dependent Phosphorylation of the Actin-binding Protein Cortactin Is Mediated by the Cytoplasmic Tyrosine Kinase FER. J. Biol. Chem. 1998, 273, 23542–23548. [Google Scholar] [CrossRef]
- Kelley, L.C.; Weed, S.A. Cortactin Is a Substrate of Activated Cdc42-Associated Kinase 1 (ACK1) during Ligand-induced Epidermal Growth Factor Receptor Downregulation. PLoS ONE 2012, 7, e44363. [Google Scholar] [CrossRef]
- Wang, W.; Liu, Y.; Liao, K. Tyrosine phosphorylation of cortactin by the FAK-Src complex at focal adhesions regulates cell motility. BMC Cell Biol. 2011, 12, 49. [Google Scholar] [CrossRef]
- Katsube, T.; Takahisa, M.; Ueda, R.; Hashimoto, N.; Kobayashi, M.; Togashi, S. Cortactin Associates with the Cell-Cell Junction Protein ZO-1 in both Drosophila and Mouse. J. Biol. Chem. 1998, 273, 29672–29677. [Google Scholar] [CrossRef]
- von Holleben, M.; Gohla, A.; Janssen, K.-P.; Iritani, B.M.; Beer-Hammer, S. Immunoinhibitory Adapter Protein Src Homology Domain 3 Lymphocyte Protein 2 (SLy2) Regulates Actin Dynamics and B Cell Spreading. J. Biol. Chem. 2011, 286, 13489–13501. [Google Scholar] [CrossRef] [PubMed]
- Kinley, A.W.; Weed, S.A.; Weaver, A.M.; Karginov, A.V.; Bissonette, E.; Cooper, J.A.; Parsons, J.T. Cortactin interacts with WIP in regulating Arp2/3 activation and membrane protrusion. Curr. Biol. 2003, 13, 384–393. [Google Scholar] [CrossRef]
- Kim, K.; Hou, P.; Gorski, J.L.; Cooper, J.A. Effect of Fgd1 on cortactin in Arp2/3 complex-mediated actin assembly. Biochemistry 2004, 43, 2422–2427. [Google Scholar] [CrossRef]
- Ren, X.; Qiao, Y.; Li, J.; Li, X.; Zhang, D.; Zhang, X.; Zhu, X.; Zhou, W.; Shi, J.; Wang, W.; et al. Cortactin recruits FMNL2 to promote actin polymerization and endosome motility in invadopodia formation. Cancer Lett. 2018, 419, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Lua, B.L.; Low, B.C. BPGAP1 Interacts with Cortactin and Facilitates Its Translocation to Cell Periphery for Enhanced Cell Migration. Mol. Biol. Cell 2004, 15, 2873–2883. [Google Scholar] [CrossRef] [PubMed]
- Ito, A.; Shimazu, T.; Maeda, S.; Shah, A.A.; Tsunoda, T.; Iemura, S.-I.; Natsume, T.; Suzuki, T.; Motohashi, H.; Yamamoto, M.; et al. The subcellular localization and activity of cortactin is regulated by acetylation and interaction with Keap1. Sci. Signal. 2015, 8, ra120. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, J.; Zhan, X. Tyrosine Phosphorylation of Cortactin Is Required for H2O2-mediated Injury of Human Endothelial Cells. J. Biol. Chem. 2000, 275, 37187–37193. [Google Scholar] [CrossRef]
- Ren, Y.; He, Y.; Brown, S.; Zbornik, E.; Mlodzianoski, M.; Ma, D.; Huang, F.; Mattoo, S.; Suter, D.M. A single tyrosine phosphorylation site in cortactin is important for filopodia formation in neuronal growth cones. Mol. Biol. Cell 2019, 30, 1817–1833. [Google Scholar] [CrossRef]
- Ran, J.; Yang, Y.; Li, D.; Liu, M.; Zhou, J. Deacetylation of α-tubulin and cortactin is required for HDAC6 to trigger ciliary disassembly. Sci. Rep. 2015, 5, 12917. [Google Scholar] [CrossRef]
- Walsh, G.; Jefferis, R. Post-translational modifications in the context of therapeutic proteins. Nat. Biotechnol. 2006, 24, 1241–1252. [Google Scholar] [CrossRef]
- Parsons, S.J.; Parsons, J.T. Src family kinases, key regulators of signal transduction. Oncogene 2004, 23, 7906–7909. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Wei, J.; Mialki, R.; Zou, C.; Mallampalli, R.K.; Zhao, Y. Extracellular Signal-regulated Kinase (ERK) Regulates Cortactin Ubiquitination and Degradation in Lung Epithelial Cells. J. Biol. Chem. 2012, 287, 19105–19114. [Google Scholar] [CrossRef] [PubMed]
- Tomar, A.; Lawson, C.; Ghassemian, M.; Schlaepfer, D.D. Cortactin as a Target for FAK in the Regulation of Focal Adhesion Dynamics. PLoS ONE 2012, 7, e44041. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Quiles, N.; Ho, H.-Y.H.; Kirschner, M.W.; Ramesh, N.; Geha, R.S. Erk/Src Phosphorylation of Cortactin Acts as a Switch On-Switch Off Mechanism That Controls Its Ability To Activate N-WASP. Mol. Cell. Biol. 2004, 24, 5269–5280. [Google Scholar] [CrossRef]
- Lua, B.L.; Low, B.C. Cortactin phosphorylation as a switch for actin cytoskeletal network and cell dynamics control. FEBS Lett. 2005, 579, 577–585. [Google Scholar] [CrossRef] [PubMed]
- Hoye, A.M.; Couchman, J.R.; Wewer, U.M.; Yoneda, A. The Phosphorylation and Distribution of Cortactin Downstream of Integrin alpha9beta1 Affects Cancer Cell Behaviour. Sci. Rep. 2016, 6, 28529. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Shrikhande, U.; Alicie, B.M.; Zhou, Q.; Geahlen, R.L. Role of the Protein Tyrosine Kinase Syk in Regulating Cell-Cell Adhesion and Motility in Breast Cancer Cells. Mol. Cancer Res. 2009, 7, 634–644. [Google Scholar] [CrossRef]
- Bandela, M.; Letsiou, E.; Natarajan, V.; Ware, L.B.; Garcia, J.G.N.; Singla, S.; Dudek, S.M. Cortactin Modulates Lung Endothelial Apoptosis Induced by Cigarette Smoke. Cells 2021, 10, 2869. [Google Scholar] [CrossRef]
- Zhao, Y.; Lei, Y.; He, S.-W.; Li, Y.-Q.; Wang, Y.-Q.; Hong, X.-H.; Liang, Y.-L.; Li, J.-Y.; Chen, Y.; Luo, W.-J.; et al. Hypermethylation of UCHL1 Promotes Metastasis of Nasopharyngeal Carcinoma by Suppressing Degradation of Cortactin (CTTN). Cells 2020, 9, 559. [Google Scholar] [CrossRef]
- Cen, G.; Ding, H.-H.; Liu, B.; Wu, W.-D. FBXL5 targets cortactin for ubiquitination-mediated destruction to regulate gastric cancer cell migration. Tumor Biol. 2014, 35, 8633–8638. [Google Scholar] [CrossRef]
- Wei, C.Y.; Zhu, M.X.; Yang, Y.W.; Zhang, P.F.; Yang, X.; Peng, R.; Gao, C.; Lu, J.C.; Wang, L.; Deng, X.Y.; et al. Downregulation of RNF128 activates Wnt/beta-catenin signaling to induce cellular EMT and stemness via CD44 and CTTN ubiquitination in melanoma. J. Hematol. Oncol. 2019, 12, 21. [Google Scholar] [CrossRef] [PubMed]
- Noh, S.J.; Baek, H.A.; Park, H.S.; Jang, K.Y.; Moon, W.S.; Kang, M.J.; Lee, D.G.; Kim, M.H.; Lee, J.H.; Chung, M.J. Expression of SIRT1 and cortactin is associated with progression of non-small cell lung cancer. Pathol. Res. Pr. 2013, 209, 365–370. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yuan, Z.; Zhang, Y.; Yong, S.; Salas-Burgos, A.; Koomen, J.; Olashaw, N.; Parsons, J.T.; Yang, X.-J.; Dent, S.R.; et al. HDAC6 Modulates Cell Motility by Altering the Acetylation Level of Cortactin. Mol. Cell 2007, 27, 197–213. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, M.; Dong, H.; Yong, S.; Li, X.; Olashaw, N.; Kruk, P.A.; Cheng, J.Q.; Bai, W.; Chen, J.; et al. Deacetylation of cortactin by SIRT1 promotes cell migration. Oncogene 2008, 28, 445–460. [Google Scholar] [CrossRef]
- Terman, J.R.; Kashina, A. Post-translational modification and regulation of actin. Curr. Opin. Cell Biol. 2012, 25, 30–38. [Google Scholar] [CrossRef]
- Tokmakov, A.A.; Kurotani, A.; Takagi, T.; Toyama, M.; Shirouzu, M.; Fukami, Y.; Yokoyama, S. Multiple Post-translational Modifications Affect Heterologous Protein Synthesis. J. Biol. Chem. 2012, 287, 27106–27116. [Google Scholar] [CrossRef]
- Oliveira-Ferrer, L.; Legler, K.; Milde-Langosch, K. Role of protein glycosylation in cancer metastasis. Semin. Cancer Biol. 2017, 44, 141–152. [Google Scholar] [CrossRef]
- Niethamer, T.K.; Stabler, C.T.; Leach, J.P.; Zepp, J.A.; Morley, M.P.; Babu, A.; Zhou, S.; Morrisey, E.E. Defining the role of pulmonary endothelial cell heterogeneity in the response to acute lung injury. eLife 2020, 9, e53072. [Google Scholar] [CrossRef]
- Ponce, A.G.; Madrid, A.F.C.; Robles, H.V.; Paredes, S.C.; Nava, P.; Betanzos, A.; Zarbock, A.; Rottner, K.; Vestweber, D.; Schnoor, M. Loss of cortactin causes endothelial barrier dysfunction via disturbed adrenomedullin secretion and actomyosin contractility. Sci. Rep. 2016, 6, 29003. [Google Scholar] [CrossRef]
- Belvitch, P.; Htwe, Y.M.; Brown, M.E.; Dudek, S. Cortical Actin Dynamics in Endothelial Permeability. Curr. Top. Membr. 2018, 82, 141–195. [Google Scholar] [CrossRef]
- Dudek, S.M.; Garcia, J.G.N. Cytoskeletal regulation of pulmonary vascular permeability. J. Appl. Physiol. 2001, 91, 1487–1500. [Google Scholar] [CrossRef] [PubMed]
- Fu, P.; Ebenezer, D.L.; Berdyshev, E.V.; Bronova, I.A.; Shaaya, M.; Harijith, A.; Natarajan, V. Role of Sphingosine Kinase 1 and S1P Transporter Spns2 in HGF-mediated Lamellipodia Formation in Lung Endothelium. J. Biol. Chem. 2016, 291, 27187–27203. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Gawlak, G.; Shah, A.S.; Higginbotham, K.; Tian, X.; Kawasaki, Y.; Akiyama, T.; Sacks, D.B.; Birukova, A.A. Hepatocyte Growth Factor-induced Asef-IQGAP1 Complex Controls Cytoskeletal Remodeling and Endothelial Barrier. J. Biol. Chem. 2015, 290, 4097–4109. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, J.R.; Dudek, S.M.; Singleton, P.A.; Kolosova, I.A.; Verin, A.D.; Garcia, J.G.N. Endothelial cell barrier enhancement by ATP is mediated by the small GTPase Rac and cortactin. Am. J. Physiol. Cell. Mol. Physiol. 2006, 291, L289–L295. [Google Scholar] [CrossRef][Green Version]
- Jacobson, J.R.; Dudek, S.M.; Birukov, K.G.; Ye, S.Q.; Grigoryev, D.N.; Girgis, R.E.; Garcia, J.G.N. Cytoskeletal Activation and Altered Gene Expression in Endothelial Barrier Regulation by Simvastatin. Am. J. Respir. Cell Mol. Biol. 2004, 30, 662–670. [Google Scholar] [CrossRef]
- Dudek, S.M.; Chiang, E.T.; Camp, S.M.; Guo, Y.; Zhao, J.; Brown, M.E.; Singleton, P.A.; Wang, L.; Desai, A.; Arce, F.T.; et al. Abl Tyrosine Kinase Phosphorylates Nonmuscle Myosin Light Chain Kinase to Regulate Endothelial Barrier Function. Mol. Biol. Cell 2010, 21, 4042–4056. [Google Scholar] [CrossRef]
- Belvitch, P.; Adyshev, D.; Elangovan, V.R.; Brown, M.E.; Naureckas, C.; Rizzo, A.N.; Siegler, J.H.; Garcia, J.G.; Dudek, S.M. Proline-rich region of non-muscle myosin light chain kinase modulates kinase activity and endothelial cytoskeletal dynamics. Microvasc. Res. 2014, 95, 94–102. [Google Scholar] [CrossRef]
- Usatyuk, P.V.; Singleton, P.A.; Pendyala, S.; Kalari, S.K.; He, D.; Gorshkova, I.A.; Camp, S.M.; Moitra, J.; Dudek, S.M.; Garcia, J.G.; et al. Novel Role for Non-muscle Myosin Light Chain Kinase (MLCK) in Hyperoxia-induced Recruitment of Cytoskeletal Proteins, NADPH Oxidase Activation, and Reactive Oxygen Species Generation in Lung Endothelium. J. Biol. Chem. 2012, 287, 9360–9375. [Google Scholar] [CrossRef]
- Arce, F.T.; Whitlock, J.L.; Birukova, A.A.; Birukov, K.G.; Arnsdorf, M.F.; Lal, R.; Garcia, J.G.; Dudek, S.M. Regulation of the Micromechanical Properties of Pulmonary Endothelium by S1P and Thrombin: Role of Cortactin. Biophys. J. 2008, 95, 886–894. [Google Scholar] [CrossRef]
- Brown, M.; Adyshev, D.; Bindokas, V.; Moitra, J.; Garcia, J.G.; Dudek, S.M. Quantitative distribution and colocalization of non-muscle myosin light chain kinase isoforms and cortactin in human lung endothelium. Microvasc. Res. 2010, 80, 75–88. [Google Scholar] [CrossRef]
- Wang, X.; Bleher, R.; Wang, L.; Garcia, J.G.N.; Dudek, S.M.; Shekhawat, G.S.; Dravid, V.P. Imatinib Alters Agonists-mediated Cytoskeletal Biomechanics in Lung Endothelium. Sci. Rep. 2017, 7, 14152. [Google Scholar] [CrossRef]
- Itoh, T.; Hasegawa, J.; Tsujita, K.; Kanaho, Y.; Takenawa, T. The Tyrosine Kinase Fer Is a Downstream Target of the PLD-PA Pathway that Regulates Cell Migration. Sci. Signal. 2009, 2, ra52. [Google Scholar] [CrossRef] [PubMed]
- Climent, M.; Viggiani, G.; Chen, Y.-W.; Coulis, G.; Castaldi, A. MicroRNA and ROS Crosstalk in Cardiac and Pulmonary Diseases. Int. J. Mol. Sci. 2020, 21, 4370. [Google Scholar] [CrossRef] [PubMed]
- Usatyuk, P.V.; Romer, L.H.; He, D.; Parinandi, N.L.; Kleinberg, M.E.; Zhan, S.; Jacobson, J.R.; Dudek, S.M.; Pendyala, S.; Garcia, J.G.; et al. Regulation of Hyperoxia-induced NADPH Oxidase Activation in Human Lung Endothelial Cells by the Actin Cytoskeleton and Cortactin. J. Biol. Chem. 2007, 282, 23284–23295. [Google Scholar] [CrossRef] [PubMed]
- Hogan, B.L.; Barkauskas, C.E.; Chapman, H.A.; Epstein, J.A.; Jain, R.; Hsia, C.C.; Niklason, L.; Calle, E.; Le, A.; Randell, S.H.; et al. Repair and Regeneration of the Respiratory System: Complexity, Plasticity, and Mechanisms of Lung Stem Cell Function. Cell Stem Cell 2014, 15, 123–138. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; He, D.; Berdyshev, E.; Zhong, M.; Salgia, R.; Morris, A.J.; Smyth, S.S.; Natarajan, V.; Zhao, Y. Autotaxin induces lung epithelial cell migration through lysoPLD activity-dependent and -independent pathways. Biochem. J. 2011, 439, 45–55. [Google Scholar] [CrossRef]
- Wu, Q.; Nadesalingam, J.; Moodley, S.; Bai, X.; Liu, M. XB130 translocation to microfilamentous structures mediates NNK-induced migration of human bronchial epithelial cells. Oncotarget 2015, 6, 18050–18065. [Google Scholar] [CrossRef][Green Version]
- Chen, D.-Y.; Husain, M. Caspase-mediated degradation of host cortactin that promotes influenza A virus infection in epithelial cells. Virology 2016, 497, 146–156. [Google Scholar] [CrossRef]
- Johnson, E.R.; Matthay, M.A. Acute Lung Injury: Epidemiology, Pathogenesis, and Treatment. J. Aerosol Med. Pulm. Drug Deliv. 2010, 23, 243–252. [Google Scholar] [CrossRef]
- Thompson, B.T.; Chambers, R.C.; Liu, K.D. Acute Respiratory Distress Syndrome. N. Engl. J. Med. 2017, 377, 562–572. [Google Scholar] [CrossRef]
- Natarajan, V.; Dudek, S.M.; Jacobson, J.R.; Moreno-Vinasco, L.; Huang, L.S.; Abassi, T.; Mathew, B.; Zhao, Y.; Wang, L.; Bittman, R.; et al. Sphingosine-1-phosphate, FTY720, and sphingosine-1-phosphate receptors in the pathobiology of acute lung injury. Am. J. Respir. Cell Mol. Biol. 2013, 49, 6–17. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, A.N.; Belvitch, P.; Demeritte, R.; Garcia, J.G.; Letsiou, E.; Dudek, S.M. Arg mediates LPS-induced disruption of the pulmonary endothelial barrier. Vasc. Pharmacol. 2020, 128–129, 106677. [Google Scholar] [CrossRef] [PubMed]
- Htwe, Y.; Wang, H.; Belvitch, P.; Meliton, L.; Bandela, M.; Letsiou, E.; Dudek, S. Group V Phospholipase A2 Mediates Endothelial Dysfunction and Acute Lung Injury Caused by Methicillin-Resistant Staphylococcus aureus. Cells 2021, 10, 1731. [Google Scholar] [CrossRef] [PubMed]
- Letsiou, E.; Sammani, S.; Wang, H.; Belvitch, P.; Dudek, S.M. Parkin regulates lipopolysaccharide-induced proinflammatory responses in acute lung injury. Transl. Res. 2017, 181, 71–82. [Google Scholar] [CrossRef]
- Ohmura, T.; Tian, Y.; Sarich, N.; Ke, Y.; Meliton, A.; Shah, A.S.; Andreasson, K.; Birukov, K.G.; Birukova, A.A. Regulation of lung endothelial permeability and inflammatory responses by prostaglandin A2: Role of EP4 receptor. Mol. Biol. Cell 2017, 28, 1622–1635. [Google Scholar] [CrossRef]
- Tian, X.; Tian, Y.; Gawlak, G.; Meng, F.; Kawasaki, Y.; Akiyama, T.; Birukova, A.A. Asef controls vascular endothelial permeability and barrier recovery in the lung. Mol. Biol. Cell 2015, 26, 636–650. [Google Scholar] [CrossRef]
- Chiang, E.T.; Camp, S.M.; Dudek, S.M.; Brown, M.E.; Usatyuk, P.V.; Zaborina, O.; Alverdy, J.C.; Garcia, J.G. Protective effects of high-molecular weight Polyethylene Glycol (PEG) in human lung endothelial cell barrier regulation: Role of actin cytoskeletal rearrangement. Microvasc. Res. 2009, 77, 174–186. [Google Scholar] [CrossRef]
- Wang, L.; Bittman, R.; Garcia, J.G.; Dudek, S.M. Junctional complex and focal adhesion rearrangement mediates pulmonary endothelial barrier enhancement by FTY720 S-phosphonate. Microvasc. Res. 2015, 99, 102–109. [Google Scholar] [CrossRef]
- Wang, L.; Letsiou, E.; Wang, H.; Belvitch, P.; Meliton, L.; Brown, M.E.; Bandela, M.; Chen, J.; Garcia, J.G.; Dudek, S.M. MRSA-Induced Endothelial Permeability and Acute Lung Injury are Attenuated by FTY720 S-Phosphonate. Am. J. Physiol. Cell. Mol. Physiol. 2022, 322, L149–L161. [Google Scholar] [CrossRef]
- Dudek, S.; Camp, S.; Chiang, E.; Singleton, P.; Usatyuk, P.; Zhao, Y.; Natarajan, V.; Garcia, J. Pulmonary endothelial cell barrier enhancement by FTY720 does not require the S1P1 receptor. Cell. Signal. 2007, 19, 1754–1764. [Google Scholar] [CrossRef]
- Rogers, A.J. Genome-Wide Association Study in Acute Respiratory Distress Syndrome. Finding the Needle in the Haystack to Advance Our Understanding of Acute Respiratory Distress Syndrome. Am. J. Respir. Crit. Care Med. 2018, 197, 1373–1374. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.; Camp, S.M.; Dan, A.; Garcia, J.G.N.; Dudek, S.M.; Leckband, D.E. A genetic variant of cortactin linked to acute lung injury impairs lamellipodia dynamics and endothelial wound healing. Am. J. Physiol. Cell. Mol. Physiol. 2015, 309, L983–L994. [Google Scholar] [CrossRef] [PubMed]
- Belvitch, P.; Casanova, N.; Sun, X.; Camp, S.M.; Sammani, S.; Brown, M.E.; Mascarhenas, J.; Lynn, H.; Adyshev, D.; Siegler, J.; et al. A cortactin CTTN coding SNP contributes to lung vascular permeability and inflammatory disease severity in African descent subjects. Transl. Res. 2022, in press. [Google Scholar] [CrossRef] [PubMed]
- Hikichi, M.; Mizumura, K.; Maruoka, S.; Gon, Y. Pathogenesis of chronic obstructive pulmonary disease (COPD) induced by cigarette smoke. J. Thorac. Dis. 2019, 11, S2129–S2140. [Google Scholar] [CrossRef] [PubMed]
- Eisner, M.D.; Anthonisen, N.; Coultas, D.; Künzli, N.; Perez-Padilla, R.; Postma, D.; Romieu, I.; Silverman, E.K.; Balmes, J.R. An Official American Thoracic Society Public Policy Statement: Novel Risk Factors and the Global Burden of Chronic Obstructive Pulmonary Disease. Am. J. Respir. Crit. Care Med. 2010, 182, 693–718. [Google Scholar] [CrossRef]
- Lee, H.-M.; Kim, C.-W.; Hwang, K.-A.; Choi, D.-W.; Choi, K.-C. Three components of cigarette smoke altered the growth and apoptosis of metastatic colon cancer cells via inducing the synthesis of reactive oxygen species and endoplasmic reticulum stress. Environ. Toxicol. Pharmacol. 2016, 45, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Zeidler, R.; Albermann, K.; Lang, S. Nicotine and apoptosis. Apoptosis 2007, 12, 1927–1943. [Google Scholar] [CrossRef]
- Vij, N.; Chandramani-Shivalingappa, P.; Van Westphal, C.; Hole, R.; Bodas, M. Cigarette smoke-induced autophagy impairment accelerates lung aging, COPD-emphysema exacerbations and pathogenesis. Am. J. Physiol. Physiol. 2018, 314, C73–C87. [Google Scholar] [CrossRef]
- Carnevali, S.; Petruzzelli, S.; Longoni, B.; Vanacore, R.; Barale, R.; Cipollini, M.; Scatena, F.; Paggiaro, P.; Celi, A.; Giuntini, C. Cigarette smoke extract induces oxidative stress and apoptosis in human lung fibroblasts. Am. J. Physiol. Cell. Mol. Physiol. 2003, 284, L955–L963. [Google Scholar] [CrossRef]
- Bandela, M.; Suryadevara, V.; Fu, P.; Reddy, S.P.; Bikkavilli, K.; Huang, L.S.; Dhavamani, S.; Subbaiah, P.V.; Singla, S.; Dudek, S.M.; et al. Role of Lysocardiolipin Acyltransferase in Cigarette Smoke-Induced Lung Epithelial Cell Mitochondrial ROS, Mitochondrial Dynamics, and Apoptosis. Cell Biophys. 2021, 80, 203–216. [Google Scholar] [CrossRef]
- Luanpitpong, S.; Talbott, S.J.; Rojanasakul, Y.; Nimmannit, U.; Pongrakhananon, V.; Wang, L.; Chanvorachote, P. Regulation of Lung Cancer Cell Migration and Invasion by Reactive Oxygen Species and Caveolin-1. J. Biol. Chem. 2010, 285, 38832–38840. [Google Scholar] [CrossRef] [PubMed]
- Lin, B.-H.; Tsai, M.-H.; Lii, C.-K.; Wang, T.-S. IP3 and calcium signaling involved in the reorganization of the actin cytoskeleton and cell rounding induced by cigarette smoke extract in human endothelial cells. Environ. Toxicol. 2015, 31, 1293–1306. [Google Scholar] [CrossRef] [PubMed]
- Silva, D.; Cáceres, M.; Arancibia, R.; Martinez, C.; Smith, P.C. Effects of cigarette smoke and nicotine on cell viability, migration and myofibroblastic differentiation. J. Periodontal Res. 2012, 47, 599–607. [Google Scholar] [CrossRef] [PubMed]
- D’Anna, C.; Cigna, D.; Di Sano, C.; Di Vincenzo, S.; Dino, P.; Ferraro, M.; Bini, L.; Bianchi, L.; Di Gaudio, F.; Gjomarkaj, M.; et al. Exposure to cigarette smoke extract and lipopolysaccharide modifies cytoskeleton organization in bronchial epithelial cells. Exp. Lung Res. 2017, 43, 347–358. [Google Scholar] [CrossRef] [PubMed]
- Unachukwu, U.; Trischler, J.; Goldklang, M.; Xiao, R.; D’Armiento, J. Maternal smoke exposure decreases mesenchymal proliferation and modulates Rho-GTPase-dependent actin cytoskeletal signaling in fetal lungs. FASEB J. 2017, 31, 2340–2351. [Google Scholar] [CrossRef]
- Leung, E.L.-H.; Fiscus, R.R.; Tung, J.W.; Tin, V.P.-C.; Cheng, L.C.; Sihoe, A.D.-L.; Fink, L.M.; Ma, Y.; Wong, M.P. Non-Small Cell Lung Cancer Cells Expressing CD44 Are Enriched for Stem Cell-Like Properties. PLoS ONE 2010, 5, e14062. [Google Scholar] [CrossRef]
- Ouhtit, A.; Thouta, R.; Zayed, H.; Gaur, R.L.; Fernando, A.; Rahman, M.; Welsh, D.A. CD44 mediates stem cell mobilization to damaged lung via its novel transcriptional targets, Cortactin and Survivin. Int. J. Med Sci. 2020, 17, 103–111. [Google Scholar] [CrossRef]
- Moore, L.D.; Le, T.; Fan, G. DNA Methylation and Its Basic Function. Neuropsychopharmacology 2013, 38, 23–38. [Google Scholar] [CrossRef]
- Joehanes, R.; Just, A.C.; Marioni, R.E.; Pilling, L.C.; Reynolds, L.M.; Mandaviya, P.R.; Guan, W.; Xu, T.; Elks, C.E.; Aslibekyan, S.; et al. Epigenetic Signatures of Cigarette Smoking. Circ. Cardiovasc. Genet. 2016, 9, 436–447. [Google Scholar] [CrossRef]
- Dharmage, S.; Perret, J.L.; Custovic, A. Epidemiology of Asthma in Children and Adults. Front. Pediatr. 2019, 7, 246. [Google Scholar] [CrossRef]
- Yayan, J.; Rasche, K. Asthma and COPD: Similarities and Differences in the Pathophysiology, Diagnosis and Therapy. 2015, 910, 31–38. Adv. Exp. Med. Biol. 2015, 910, 31–38. [Google Scholar] [CrossRef]
- Wan, Y.I.; Soler-Artigas, M.; Shrine, N.R.; Wain, L.V.; Tobin, M.D.; Moffatt, M.F.; Bush, A.; Cookson, W.O.; Strachan, D.; Heaney, L.; et al. Genome-Wide Association Study To Identify Genetic Determinants Of Severe Asthma (AUGOSA). Am. J. Respir. Crit. Care Med. 2011, 183, A6170. [Google Scholar] [CrossRef]
- Ma, S.-F.; Flores, C.; Wade, M.S.; Dudek, S.M.; Nicolae, D.L.; Ober, C.; Garcia, J.G. A common cortactin gene variation confers differential susceptibility to severe asthma. Genet. Epidemiol. 2008, 32, 757–766. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.Q.; Shirakawa, T.; Kawai, M.; Enomoto, T.; Sasaki, S.; Dake, Y.; Kitano, H.; Hagihara, A.; Hopkin, J.M.; Morimoto, K. Association between asthma and an intragenic variant of CC16 on chromosome 11q13. Clin. Genet. 1998, 53, 54–56. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Cleary, R.A.; Wang, T.; Li, J.; Tang, D.D. The Association of Cortactin with Profilin-1 Is Critical for Smooth Muscle Contraction. J. Biol. Chem. 2014, 289, 14157–14169. [Google Scholar] [CrossRef]
- Wang, Y.; Rezey, A.C.; Wang, R.; Tang, D.D. Role and regulation of Abelson tyrosine kinase in Crk-associated substrate/profilin-1 interaction and airway smooth muscle contraction. Respir. Res. 2018, 19, 4. [Google Scholar] [CrossRef]
- Cleary, R.A.; Wang, R.; Waqar, O.; Singer, H.A.; Tang, D.D. Role of c-Abl tyrosine kinase in smooth muscle cell migration. Am. J. Physiol. Physiol. 2014, 306, C753-61. [Google Scholar] [CrossRef][Green Version]
- Rizzo, A.N.; Aman, J.; Amerongen, G.P.V.N.; Dudek, S.M. Targeting Abl Kinases to Regulate Vascular Leak During Sepsis and Acute Respiratory Distress Syndrome. Arter. Thromb. Vasc. Biol. 2015, 35, 1071–1079. [Google Scholar] [CrossRef]
- Brescia, G.; Parrino, D.; Nicolè, L.; Zanotti, C.; Lanza, C.; Barion, U.; Marino, F.; Marioni, G. Cortactin expression in nasal polyps of Aspirin-Exacerbated Respiratory Disease (AERD) patients. Am. J. Otolaryngol. 2018, 39, 293–298. [Google Scholar] [CrossRef]
- Li, J.; Chen, S.; Cleary, R.A.; Wang, R.; Gannon, O.J.; Seto, E.; Tang, D.D. Histone deacetylase 8 regulates cortactin deacetylation and contraction in smooth muscle tissues. Am. J. Physiol. Physiol. 2014, 307, C288–C295. [Google Scholar] [CrossRef]
- Liu, C.; Li, Q.; Zhou, X.; Kolosov, V.P.; Perelman, J.M. Cortactin mediates elevated shear stress-induced mucin hypersecretion via actin polymerization in human airway epithelial cells. Int. J. Biochem. Cell Biol. 2013, 45, 2756–2763. [Google Scholar] [CrossRef] [PubMed]
- Thekkeveedu, R.K.; Guaman, M.C.; Shivanna, B. Bronchopulmonary dysplasia: A review of pathogenesis and pathophysiology. Respir. Med. 2017, 132, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.; Heyob, K.; Tipple, T.; Pryhuber, G.S.; Rogers, L.K. Alterations in VASP phosphorylation and profilin1 and cofilin1 expression in hyperoxic lung injury and BPD. Respir. Res. 2018, 19, 229. [Google Scholar] [CrossRef]
- Xing, Y.; Fu, J.; Yang, H.; Yao, L.; Qiao, L.; Du, Y.; Xue, X. MicroRNA expression profiles and target prediction in neonatal Wistar rat lungs during the development of bronchopulmonary dysplasia. Int. J. Mol. Med. 2015, 36, 1253–1263. [Google Scholar] [CrossRef][Green Version]
- Yin, M.; Ma, W.; An, L. Cortactin in cancer cell migration and invasion. Oncotarget 2017, 8, 88232–88243. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, H.; Gong, H.; Yuan, Y.; Li, Y.; Wang, C.; Li, W.; Zhang, Z.; Liu, M.; Liu, H.; et al. miR-182 suppresses invadopodia formation and metastasis in non-small cell lung cancer by targeting cortactin gene. J. Exp. Clin. Cancer Res. 2018, 37, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Shenoy, A.K.; Padia, R.; Fang, D.; Jing, Q.; Yang, P.; Su, S.-B.; Huang, S. Suppression of lung cancer progression by isoliquiritigenin through its metabolite 2, 4, 2’, 4’-Tetrahydroxychalcone. J. Exp. Clin. Cancer Res. 2018, 37, 243. [Google Scholar] [CrossRef]
- Fan, M.; Wu, J.; Li, X.; Jiang, Y.; Wang, X.; Bie, M.; Weng, Y.; Chen, S.; Chen, B.; An, L.; et al. CX 3 CL1 promotes tumour cell by inducing tyrosine phosphorylation of cortactin in lung cancer. J. Cell. Mol. Med. 2020, 25, 132–146. [Google Scholar] [CrossRef]
- Yamada, H.; Takeda, T.; Michiue, H.; Abe, T.; Takei, K. Actin bundling by dynamin 2 and cortactin is implicated in cell migration by stabilizing filopodia in human non-small cell lung carcinoma cells. Int. J. Oncol. 2016, 49, 877–886. [Google Scholar] [CrossRef]
- McNiven, M.A.; Kim, L.; Krueger, E.W.; Orth, J.; Cao, H.; Wong, T.W. Regulated Interactions between Dynamin and the Actin-Binding Protein Cortactin Modulate Cell Shape. J. Cell Biol. 2000, 151, 187–198. [Google Scholar] [CrossRef]
- Shi, X.; Duan, F.; Lin, L.; Xu, Q.; Xu, T.; Zhang, R. WIP-1 and DBN-1 promote scission of endocytic vesicles by bridging actin and Dynamin-1 in the C. elegans intestine. J. Cell Sci. 2019, 132, jcs228023. [Google Scholar] [CrossRef] [PubMed]
- Leong, H.; Robertson, A.E.; Stoletov, K.; Leith, S.J.; Chin, C.A.; Chien, A.E.; Hague, M.N.; Ablack, A.; Carmine-Simmen, K.; McPherson, V.A.; et al. Invadopodia Are Required for Cancer Cell Extravasation and Are a Therapeutic Target for Metastasis. Cell Rep. 2014, 8, 1558–1570. [Google Scholar] [CrossRef] [PubMed]
- Armacki, M.; Polaschek, S.; Waldenmaier, M.; Morawe, M.; Ruhland, C.; Schmid, R.; Lechel, A.; Tharehalli, U.; Steup, C.; Bektas, Y.; et al. Protein Kinase D1, Reduced in Human Pancreatic Tumors, Increases Secretion of Small Extracellular Vesicles From Cancer Cells That Promote Metastasis to Lung in Mice. Gastroenterology 2020, 159, 1019–1035.e22. [Google Scholar] [CrossRef] [PubMed]
- Su, C.-M.; Su, Y.-H.; Chiu, C.-F.; Chang, Y.-W.; Hong, C.-C.; Yu, Y.-H.; Ho, Y.-S.; Wu, C.-H.; Yen, C.-S.; Su, J.-L. Vascular Endothelial Growth Factor-C Upregulates Cortactin and Promotes Metastasis of Esophageal Squamous Cell Carcinoma. Ann. Surg. Oncol. 2014, 21, 767–775. [Google Scholar] [CrossRef]
- Debreova, M.; Csaderova, L.; Burikova, M.; Lukacikova, L.; Kajanova, I.; Sedlakova, O.; Kery, M.; Kopacek, J.; Zatovicova, M.; Bizik, J.; et al. CAIX Regulates Invadopodia Formation through Both a pH-Dependent Mechanism and Interplay with Actin Regulatory Proteins. Int. J. Mol. Sci. 2019, 20, 2745. [Google Scholar] [CrossRef]
- Damiano, L.; Le Dévédec, S.E.; Di Stefano, P.; Repetto, D.; Lalai, R.; Truong, H.; Xiong, J.L.; Danen, E.H.; Yan, K.; Verbeek, F.J.; et al. p140Cap suppresses the invasive properties of highly metastatic MTLn3-EGFR cells via impaired cortactin phosphorylation. Oncogene 2011, 31, 624–633. [Google Scholar] [CrossRef]
- Majumder, S.; Sowden, M.P.; Gerber, S.A.; Thomas, T.; Christie, C.K.; Mohan, A.; Yin, G.; Lord, E.M.; Berk, B.C.; Pang, J. G-Protein–Coupled Receptor-2–Interacting Protein-1 Is Required for Endothelial Cell Directional Migration and Tumor Angiogenesis via Cortactin-Dependent Lamellipodia Formation. Arter. Thromb. Vasc. Biol. 2014, 34, 419–426. [Google Scholar] [CrossRef]
- Kaluza, D.; Kroll, J.; Gesierich, S.; Yao, T.-P.; Boon, R.; Hergenreider, E.; Tjwa, M.; Rössig, L.; Seto, E.; Augustin, H.; et al. Class IIb HDAC6 regulates endothelial cell migration and angiogenesis by deacetylation of cortactin. EMBO J. 2011, 30, 4142–4156. [Google Scholar] [CrossRef]
- Wen, Y.; Gamazon, E.R.; Bleibel, W.K.; Wing, C.; Mi, S.; McIlwee, B.E.; Delaney, S.M.; Duan, S.; Im, H.K.; Dolan, M.E. An eQTL-based method identifies CTTN and ZMAT3 as pemetrexed susceptibility markers. Hum. Mol. Genet. 2012, 21, 1470–1480. [Google Scholar] [CrossRef][Green Version]
- Faoro, L.; Singleton, P.A.; Cervantes, G.M.; Lennon, F.E.; Choong, N.W.; Kanteti, R.; Ferguson, B.; Husain, A.N.; Tretiakova, M.S.; Ramnath, N.; et al. EphA2 Mutation in Lung Squamous Cell Carcinoma Promotes Increased Cell Survival, Cell Invasion, Focal Adhesions, and Mammalian Target of Rapamycin Activation. J. Biol. Chem. 2010, 285, 18575–18585. [Google Scholar] [CrossRef]
- Surjit, M.; Liu, B.; Jameel, S.; Chow, V.; Lal, S.K. The SARS coronavirus nucleocapsid protein induces actin reorganization and apoptosis in COS-1 cells in the absence of growth factors. Biochem. J. 2004, 383, 13–18. [Google Scholar] [CrossRef]
- Lv, X.; Li, Z.; Guan, J.; Hu, S.; Zhang, J.; Lan, Y.; Zhao, K.; Lu, H.; Song, D.; He, H. Porcine Hemagglutinating Encephalomyelitis Virus Activation of the Integrin alpha5beta1-FAK-Cofilin Pathway Causes Cytoskeletal Rearrangement To Promote Its Invasion of N2a Cells. J. Virol. 2019, 93, e01736-18. [Google Scholar] [CrossRef]
- Zhou, P.; Yang, X.-L.; Wang, X.-G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.-R.; Zhu, Y.; Li, B.; Huang, C.-L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [PubMed]
- Eisenhut, M.; Shin, J.I. Pathways in the Pathophysiology of Coronavirus 19 Lung Disease Accessible to Prevention and Treatment. Front. Physiol. 2020, 11, 872. [Google Scholar] [CrossRef] [PubMed]
- Oldstone, M.B.A.; Sinha, Y.N.; Blount, P.; Tishon, A.; Rodriguez, M.; von Wedel, R.; Lampert, P.W. Virus-Induced Alterations in Homeostasis: Alteration in Differentiated Functions of Infected Cells in Vivo. Science 1982, 218, 1125–1127. [Google Scholar] [CrossRef] [PubMed]
- Taylor, M.P.; Koyuncu, O.O.; Enquist, L.W. Subversion of the actin cytoskeleton during viral infection. Nat. Rev. Genet. 2011, 9, 427–439. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Zhu, L.; Yang, X.; Lin, J.; Yang, Q. The epidermal growth factor receptor regulates cofilin activity and promotes transmissible gastroenteritis virus entry into intestinal epithelial cells. Oncotarget 2016, 7, 12206–12221. [Google Scholar] [CrossRef]
- Swaine, T.; Dittmar, M.T. CDC42 Use in Viral Cell Entry Processes by RNA Viruses. Viruses 2015, 7, 6526–6536. [Google Scholar] [CrossRef]
- Barrantes, F.J. While We Wait for a Vaccine Against SARS-CoV-2, Why Not Think About Available Drugs? Front. Physiol. 2020, 11, 820. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Protein | Site of Interaction with CTTN | References | |
|---|---|---|---|
| 1 | SHANK2 and SHANK3 | SH3 domain | [15] |
| 2 | FGD1 | SH3 domain | [16] |
| 3 | ABL2 | Homology | [17] |
| 4 | KCNA2 | C terminus | [18] |
| 5 | SAMSN1 | SH3 domain | [19] |
| 6 | ASAP1 | SH3 domain/Pro-rich region | [20] |
| 7 | DNM2 | SH3 domain/homology | [21] |
| 8 | ACTN1 | Homology | [22] |
| 9 | N-WASP | SH3 domain | [23] |
| 10 | Arp2/3 complex | NTA/SH3 domain | [24] |
| 11 | Kv1.3/Kv1.2 | CTTN | [18,25] |
| 12 | Caspase cleavage sites | Actin binding domains/SH3 domain | [26,27] |
| 13 | UCS15A/AMAP1 | SH3 domain/pro-rich region | [28,29] |
| 14 | STAM1, TXK, Fyn, Hck | SH3 domain | [30] |
| 15 | AFAP1L1 | SH3 domain | [31] |
| 16 | Calcium activated potassium channels | SH3 domain | [32] |
| 17 | CD2AP | SH3 domain | [33,34] |
| 18 | TIP150 | C terminal tail | [35] |
| 19 | CBP90 | SH3 domain | [36] |
| 20 | Cytoplasmic tyrosine kinase FER | CTTN | [37] |
| 21 | MLCK | SH3 domain | [14] |
| 22 | Cdc42-associated kinase 1(ACK1) | SH3 domain | [38] |
| 23 | FAK | SH3 domain | [39] |
| 24 | Protein ZO-1 | SH3 domain | [40] |
| 25 | Lymphocyte protein 2(Sly2) | SH3 domain | [41] |
| 26 | WIP | SH3 domain | [42] |
| 27 | Fgd1, Cdc42, GEF | SH3 domain | [43,44] |
| 28 | BPGAP1 | SH3 domain | [45] |
| 29 | Keap1 | CTTN | [46] |
| Protein | Function | References | |
|---|---|---|---|
| 1 | miR-182 | ↓ invadopodia formation and metastasis Inhibits invasion and proliferation | [136,142,143] |
| 2 | Isoliquiritigenin,2,4,2′,4, Tetrahydroxychalcone metabolite | ↓ lung cancer invasion | [137] |
| 3 | SIRT1 (sirtuin1) | Involved in tumor progression | [62] |
| 4 | Dynamin1 | Cell migration, stabilizes filopodia formation | [140,141] |
| 5 | Actinin-1, Ect2 | Localization of invadopodia, matrix degradation and migration | [22] |
| 6 | MT1-MMP, Tks4, Tks5 | Invadopodia during cancer extravasation and metastasis | [142] |
| 7 | Protein Kinase D1 | ↑ extracellular vesicles, promotes metastasis | [143] |
| 8 | Vascular endothelial growth factor- C | Promotes metastasis | [144] |
| 9 | CAIX-Tumor associated carbonic anhydrase IX | Promotes Invadopodia formation and matrix degradation | [145] |
| 10 | Dynamin2 | Suppresses lamellipodia formation and invasion | [139] |
| 11 | P140Cap | Suppresses invasion of MTLn3-EGFR | [146] |
| 12 | G-protein-coupled receptor-2-interacting protein-1 | Directional migration and tumor angiogenesis, lamellipodia formation | [147] |
| 13 | XB130 | Mediates NNK-induced migration | [87] |
| 14 | Class II HDAC6 | Endothelial cell migration and angiogenesis | [148] |
| 15 | Abl Kinases | Cancer progression | [128] |
| 16 | ZMAT3 | Knockdown of CTTN/ZMAT3 ↓ cell survival | [149] |
| 17 | EphA2 mutation | Promotes cell survival, cell invasion and mammalian target of rapamycin activation | [150] |
| 18 | CD44 | HGF mediated vascular integrity | [116,151,152] |
| 19 | CX (3) CL1 | Cell invasion and migration | [138] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bandela, M.; Belvitch, P.; Garcia, J.G.N.; Dudek, S.M. Cortactin in Lung Cell Function and Disease. Int. J. Mol. Sci. 2022, 23, 4606. https://doi.org/10.3390/ijms23094606
Bandela M, Belvitch P, Garcia JGN, Dudek SM. Cortactin in Lung Cell Function and Disease. International Journal of Molecular Sciences. 2022; 23(9):4606. https://doi.org/10.3390/ijms23094606
Chicago/Turabian StyleBandela, Mounica, Patrick Belvitch, Joe G. N. Garcia, and Steven M. Dudek. 2022. "Cortactin in Lung Cell Function and Disease" International Journal of Molecular Sciences 23, no. 9: 4606. https://doi.org/10.3390/ijms23094606
APA StyleBandela, M., Belvitch, P., Garcia, J. G. N., & Dudek, S. M. (2022). Cortactin in Lung Cell Function and Disease. International Journal of Molecular Sciences, 23(9), 4606. https://doi.org/10.3390/ijms23094606