
The Family Keeps on Growing: Four Novel Fungal OYEs Characterized

 ,
,  , , and
, , and
Abstract
1. Introduction
2. Results and Discussion
2.1. Identification of the New Putative ERs and Sequence Analysis
2.2. Production and Purification of the Recombinant Proteins
2.3. Oligomeric State Determination
2.4. Substrate Spectra
2.5. Steady State Kinetic Parameters
2.6. pH Optimum and Thermal Stability
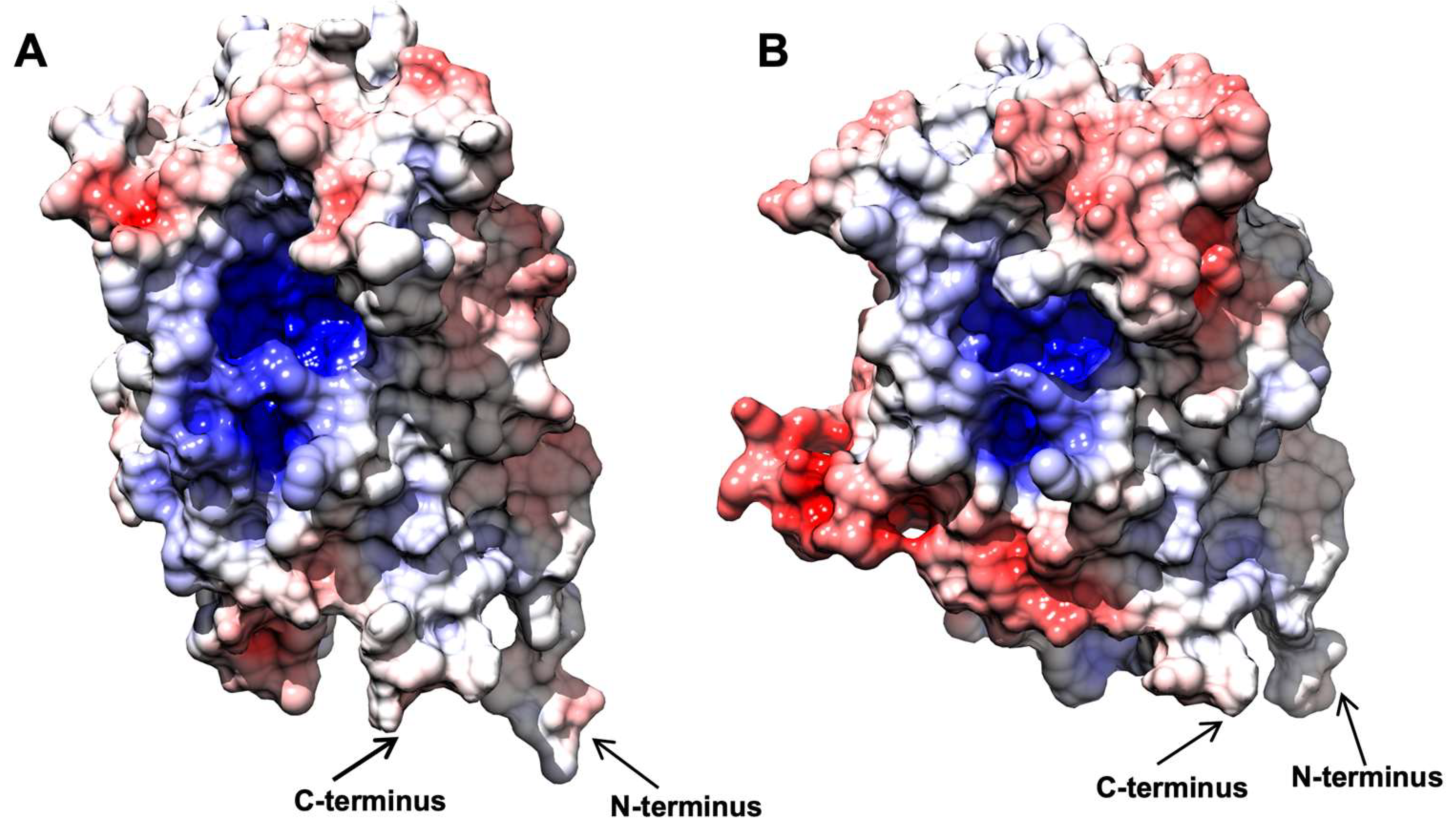
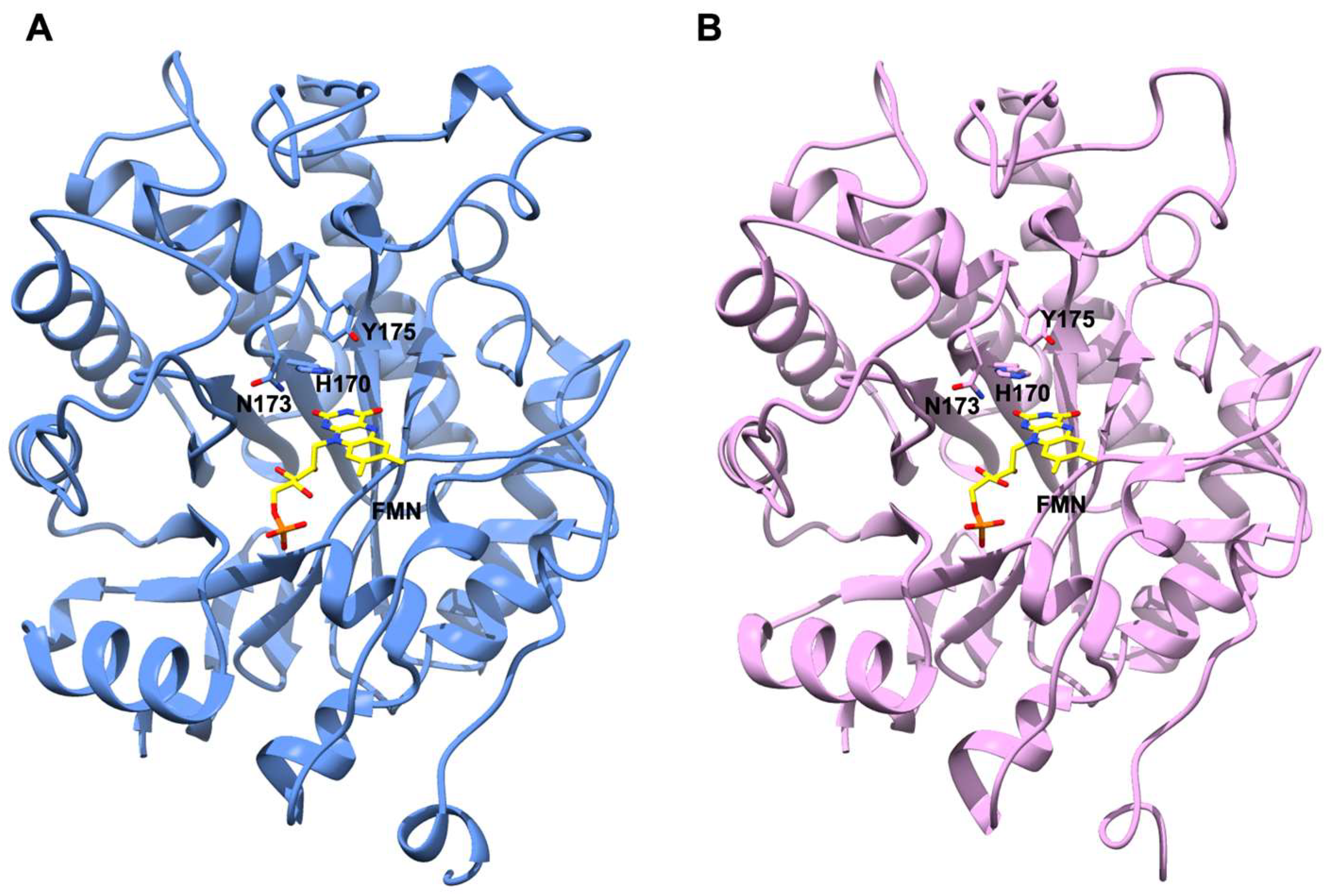
2.7. AnOYE8 X-ray Crystal Structure
2.8. AnOYE2 and BfOYE1 Model Structures
3. Materials and Methods
3.1. Organisms and Culture Conditions
3.2. Sequence Analysis and Cloning
3.3. Production, Analysis, and Purification of Recombinant Proteins
3.4. Oligomeric State Determination by Analytical Gel Filtration and BN-PAGE
3.5. Activity Assay and Kinetics
3.6. Determination of pH Optimum
3.7. Thermofluor Measurements
3.8. Crystallization and Data Collection
3.9. Model Building and Refinement
3.10. Bioinformatic Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Toogood, H.S.; Scrutton, N.S. Discovery, Characterization, Engineering, and Applications of Ene-Reductases for Industrial Biocatalysis. ACS Catal. 2018, 8, 3532–3549. [Google Scholar] [CrossRef] [PubMed]
- Nizam, S.; Verma, S.; Borah, N.N.; Gazara, R.K.; Verma, P.K. Comprehensive Genome-Wide Analysis Reveals Different Classes of Enigmatic Old Yellow Enzyme in Fungi. Sci. Rep. 2014, 4, 4013–4024. [Google Scholar] [CrossRef] [PubMed]
- Romagnolo, A.; Spina, F.; Brenna, E.; Crotti, M.; Parmeggiani, F.; Varese, G.C. Identification of Fungal Ene-Reductase Activity by Means of a Functional Screening. Fungal Biol. 2015, 119, 487–493. [Google Scholar] [CrossRef] [PubMed]
- Carballeira, J.D.; Valmaseda, M.; Alvarez, E.; Gago, J.V.S. Gongronella butleri, Schizosaccharomyces octosporus and Diplogelasinospora grovesii: Novel Microorganisms Useful for the Stereoselective Reduction of Ketones. Enzyme Microb. Technol. 2004, 34, 611–623. [Google Scholar] [CrossRef]
- Birolli, W.G.; Lima, R.N.; Porto, A.L.M. Applications of Marine-Derived Microorganisms and Their Enzymes in Biocatalysis and Biotransformation, the Underexplored Potentials. Front. Microbiol. 2019, 10, 1453. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, I.M.; Rocha, L.C.; Yoshioka, S.A.; Nitschke, M.; Jeller, A.H.; Pizzutti, L.; Seleghim, M.H.R.; Porto, A.L.M. Chemoselective Reduction of Chalcones by Whole Hyphae of Marine Fungus Penicillium citrinum CBMAI 1186, Free and Immobilized on Biopolymers. Biocatal. Agric. Biotechnol. 2014, 3, 358–364. [Google Scholar] [CrossRef]
- Ferreira, I.M.; Meira, E.B.; Rosset, I.G.; Porto, A.L.M. Chemoselective Biohydrogenation of α,β- and α,β,γ,δ-Unsaturated Ketones by the Marine-Derived Fungus Penicillium citrinum CBMAI 1186 in a Biphasic System. J. Mol. Catal. B Enzym. 2015, 115, 59–65. [Google Scholar] [CrossRef]
- Ferreira, I.M.; de Vasconcellos, S.P.; da Cruz, J.B.; Comassetto, J.V.; Porto, A.L.M.; Rocha, L.C. Hydrogenation of Bis-α,β-Unsaturated Enones Mediated by Filamentous Fungi. Biocatal. Agric. Biotechnol. 2015, 4, 144–149. [Google Scholar] [CrossRef]
- Ferreira, I.M.; Fiamingo, A.; Campana-Filho, S.P.; Porto, A.L.M. Biotransformation of (E)-2-Methyl-3-Phenylacrylaldehyde Using Mycelia of Penicillium citrinum CBMAI 1186, Both Free and Immobilized on Chitosan. Mar. Biotechnol. 2020, 22, 348–356. [Google Scholar] [CrossRef]
- Jimenez, D.E.Q.; Barreiro, J.C.; Santos, F.M.; Vasconcellos, S.P.; Porto, A.L.M.; Batista, J.M. Enantioselective Ene-Reduction of (E)-2-Cyano-3-(Furan-2-yl) Acrylamide by Marine and Terrestrial Fungi and Absolute Configuration of (R)-2-Cyano-3-(Furan-2-yl) Propanamide Determined by Calculations of Electronic Circular Dichroism (ECD) Spectra. Chirality 2019, 31, 534–542. [Google Scholar] [CrossRef]
- Matos, I.L.; Nitschke, M.; Porto, A.L.M. Hydrogenation of Halogenated 2′-Hydroxychalcones by Mycelia of Marine-Derived Fungus Penicillium raistrickii. Mar. Biotechnol. 2019, 21, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Janicki, I.; Kielbasinski, P.; Szelag, J.; Glebski, A.; Szczesna-Antczak, M. Preparative Scale Application of Mucor circinelloides Ene–Reductase and Alcohol Dehydrogenase Activity for the Asymmetric Bioreduction of α,β-Unsaturated γ-Ketophosphonates. Bioorg. Chem. 2020, 98, 103548. [Google Scholar] [CrossRef] [PubMed]
- Wallwey, C.; Matuschek, M.; Xie, X.-L.; Li, S.-M. Ergot Alkaloid Biosynthesis in Aspergillus fumigatus: Conversion of Chanoclavine-I Aldehyde to Festuclavine by the Festuclavine Synthase FgaFS in the Presence of the Old Yellow Enzyme FgaOx3. Org. Biomol. Chem. 2010, 8, 3500–3508. [Google Scholar] [CrossRef]
- Cheng, J.Z.; Coyle, C.M.; Panaccione, D.G.; O’Connor, S.E. A Role for Old Yellow Enzyme in Ergot Alkaloid Biosynthesis. J. Am. Chem. Soc. 2010, 132, 1776–1777. [Google Scholar] [CrossRef]
- Matuschek, M.; Wallwey, C.; Wollinsky, B.; Xie, X.; Li, S.-M. In Vitro Conversion of Chanoclavine-I Aldehyde to the Stereoisomers Festuclavine and Pyroclavine Controlled by the Second Reduction Step. RSC Adv. 2012, 2, 3662–3669. [Google Scholar] [CrossRef]
- Gerhards, N.; Li, S.-M. A Bifunctional Old Yellow Enzyme from Penicillium roqueforti Is Involved in Ergot Alkaloid Biosynthesis. Org. Biomol. Chem. 2017, 15, 8059. [Google Scholar] [CrossRef]
- Li, Z.; Wang, Z.; Meng, G.; Lu, H.; Huang, Z.; Chen, F. Identification of an Ene-Reductase from Yeast Kluyveromyces marxianus and Application in the Asymmetric Synthesis of (R)-profen Esters. Asian J. Org. Chem. 2018, 7, 763–769. [Google Scholar] [CrossRef]
- Janicki, I.; Kielbasinski, P.; Turrini, N.G.; Faber, K.; Hall, M. Asymmetric Bioreduction of β-activated Vinylphosphonate Derivatives Using Ene-reductases. Adv. Synth. Catal. 2017, 359, 4190–4196. [Google Scholar] [CrossRef]
- Robescu, M.S.; Cendron, L.; Bacchin, A.; Wagner, K.; Reiter, T.C.; Janicki, I.; Merusic, K.; Illek, M.; Bergantino, E.; Hall, M. Asymmetric Proton Transfer Catalysis by Stereocomplementary Old Yellow Enzymes for C=C-Bond Isomerization Reaction. 2022; under revision. [Google Scholar]
- Böhmer, S.; Marx, C.; Gomez-Baraibar, A.; Nowaczyk, M.M.; Tischler, D.; Hemschemeier, A.; Happe, T. Evolutionary Diverse Chlamydomonas reinhardtii Old Yellow Enzymes Reveal Distinctive Catalytic Properties and Potential for Whole-Cell Biotransformations. Algal Res. 2020, 50, 101970. [Google Scholar] [CrossRef]
- Nizam, S.; Gazara, R.K.; Verma, S.; Singh, K.; Verma, P.K. Comparative Structural Modeling of Six Old Yellow Enzymes (OYEs) from the Necrotrophic Fungus Ascochyta rabiei: Insight into Novel OYE Classes with Differences in Cofactor Binding, Organization of Active Site Residues and Stereopreferences. PLoS ONE 2014, 9, e95989. [Google Scholar] [CrossRef]
- Scholtissek, A.; Tischler, D.; Westphal, A.H.; van Berkel, W.J.H.; Paul, C.E. Old Yellow Enzyme-Catalysed Asymmetric Hydrogenation: Linking Family Roots with Improved Catalysis. Catalysts 2017, 7, 130. [Google Scholar] [CrossRef]
- Joseph, R.E.; Andreotti, A.H. Bacterial Expression and Purification of Interleukin-2 Tyrosine Kinase: Single Step Separation of the Chaperonin Impurity. Protein Expr. Purif. 2008, 60, 194–197. [Google Scholar] [CrossRef] [PubMed]
- Adalbjornsson, B.V.; Toogood, H.S.; Fryszkowska, A.; Pudney, C.R.; Jowitt, T.A.; Leys, D.; Scrutton, N.S. Biocatalysis with Thermostable Enzymes: Structure and Properties of a Thermophilic Ene-Reductase Related to Old Yellow Enzyme. ChemBioChem 2010, 11, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Opperman, D.J.; Piater, L.A.; van Heerden, E. A Novel Chromate Reductase from Thermus scotoductus SA-01 Related to Old Yellow Enzyme. J. Bacteriol. 2008, 190, 3076–3082. [Google Scholar] [CrossRef]
- Litthauer, S.; Gargiulo, S.; van Heerden, E.; Hollmann, F.; Opperman, D.J. Heterologous Expression and Characterization of the Ene-Reductases from Deinococcus radiodurans and Ralstonia metallidurans. J. Mol. Catal. B Enzym. 2014, 99, 89–95. [Google Scholar] [CrossRef]
- Robescu, M.S.; Niero, M.; Loprete, G.; Cendron, L.; Bergantino, E. A New Thermophilic Ene-Reductase from the Filamentous Anoxygenic Phototrophic Bacterium Chloroflexus aggregans. Microorganisms 2021, 9, 953. [Google Scholar] [CrossRef]
- Riedel, A.; Mehnert, M.; Paul, C.E.; Westphal, A.H.; van Berkel, W.J.H.; Tischler, D. Functional Characterization and Stability Improvement of a Thermophilic-like Ene-Reductase from Rhodococcus opacus 1CP. Front. Microbiol. 2015, 6, 1073. [Google Scholar] [CrossRef]
- Catucci, G.; Romagnolo, A.; Spina, F.; Varese, G.C.; Gilardi, G.; Di Nardo, G. Enzyme-Substrate Matching in Biocatalysis: In Silico Studies to Predict Substrate Preference of Ten Putative Ene-Reductases from Mucor circinelloides MUT44. J. Mol. Catal. B Enzym. 2016, 131, 94–100. [Google Scholar] [CrossRef]
- Romagnolo, A.; Spina, F.; Poli, A.; Risso, S.; Serito, B.; Crotti, M.; Monti, D.; Brenna, E.; Lanfranco, L.; Varese, G.C. Old Yellow Enzyme Homologues in Mucor circinelloides: Expression Profile and Biotransformation. Sci. Rep. 2017, 7, 12093. [Google Scholar] [CrossRef]
- Krissinel, E.; Henrick, K. Inference of Macromolecular Assemblies from Crystalline State. J. Mol. Biol. 2007, 372, 774–797. [Google Scholar] [CrossRef]
- Oberdorfer, G.; Steinkellner, G.; Stueckler, C.; Faber, K.; Gruber, K. Stereopreferences of Old Yellow Enzymes: Structure Correlations and Sequence Patterns in Enoate Reductases. ChemCatChem 2011, 3, 1562–1566. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A New Generation of Protein Database Search Programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; LI, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Soding, J.; et al. Fast, Scalable Generation of High-quality Protein Multiple Sequence Alignments Using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef] [PubMed]
- Allen, G.; Flores-Vergara, M.; Krasnyanski, S.; Kumar, S.; Thompson, W. A Modified Protocol for Rapid DNA Isolation from Plant Tissues Using Cetyltrimethylammonium Bromide. Nat. Protoc. 2006, 1, 2320–2325. [Google Scholar] [CrossRef]
- Robescu, M.S.; Niero, M.; Hall, M.; Cendron, L.; Bergantino, E. Two New Ene-Reductases from Photosynthetic Extremophiles Enlarge the Panel of Old Yellow Enzymes: CtOYE and GsOYE. Appl. Microbiol. Biotechnol. 2020, 104, 2051–2066. [Google Scholar] [CrossRef]
- Fu, Y.; Castiglione, K.; Weuster-Botz, D. Comparative Characterization of Novel Ene-Reductases from Cyanobacteria. Biotechnol. Bioeng. 2013, 110, 1293–1301. [Google Scholar] [CrossRef]
- Kabsch, W. XDS. Acta Crystallogr. D Biol. Crystallogr. 2010, D66, 125–132. [Google Scholar] [CrossRef]
- Winn, M.D.; Ballard, C.C.; Cowtan, K.D.; Dodson, E.J.; Emsley, P.; Evans, P.R.; Keegan, R.M.; Leslie, A.G.W.; McCoy, A.; McNicholas, S.J.; et al. Overview of the CCP4 Suite and Current Developments. Acta Crystallogr. D Biol. Crystallogr. 2011, D67, 235–242. [Google Scholar] [CrossRef]
- Biasini, M.; Bienert, S.; Waterhouse, A.; Arnold, K.; Studer, G.; Schmidt, T.; Kiefer, F.; Cassarino, T.G.; Bertoni, M.; Bordoli, L.; et al. SWISS-MODEL: Modelling Protein Tertiary and Quaternary Structure Using Evolutionary Information. Nucleic Acids Res. 2014, 42, W252–W258. [Google Scholar] [CrossRef]
- Murshudov, G.N.; Skubák, P.; Lebedev, A.A.; Pannu, N.S.; Steiner, R.A.; Nicholls, R.A.; Winn, M.D.; Long, F.; Vagin, A.A. REFMAC 5 for the Refinement of Macromolecular Crystal Structures. Acta Crystallogr. D Biol. Crystallogr. 2011, 67, 355–367. [Google Scholar] [CrossRef]
- Adams, P.D.; Afonine, P.V.; Bunkóczi, G.; Chen, V.B.; Davis, I.W.; Echols, N.; Headd, J.J.; Hung, L.-W.; Kapral, G.J.; Grosse-Kunstleve, R.W.; et al. PHENIX: A Comprehensive Python-Based System for Macromolecular Structure Solution. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 213–221. [Google Scholar] [CrossRef]
- Benkert, P.; Biasini, M.; Schwede, T. Toward the Estimation of the Absolute Quality of Individual Protein Structure Models. Bioinformatics 2011, 27, 343–350. [Google Scholar] [CrossRef]
- Chen, V.B.; Bryan, W.; Iii, A.; Headd, J.J.; Keedy, D.A.; Kapral, G.J.; Murray, L.W.; Richardson, J.S.; Richardson, D.C. MolProbity: All-Atom Structure Validation for Macromolecular Crystallography. Acta Crystallogr. D Biol. Crystallogr. 2010, D66, 12–21. [Google Scholar] [CrossRef]
- Dolinsky, T.J.; Nielsen, J.E.; McCammon, J.A.; Baker, N.A. PDB2PQR: An Automated Pipeline for the Setup of Poisson-Boltzmann Electrostatics Calculations. Nucleic Acids Res. 2004, 32, W665–W667. [Google Scholar] [CrossRef]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Pall, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High Performance Molecular Simulations through Multi-Level Parallelism from Laptops to Supercomputers. SoftwareX 2015, 1–2, 19–25. [Google Scholar] [CrossRef]
- Huang, J.; Rauscher, S.; Nawrocki, G.; Ran, T.; Feig, M.; de Groot, B.L.; Grubmuller, H.; MacKerell, A.D., Jr. CHARMM36m: An Improved Force Field for Folded and Intrinsically Disordered Proteins. Nat. Methods 2017, 14, 71–73. [Google Scholar] [CrossRef]
- Zhang, C.; Freddolino, P.L.; Zhang, Y. COFACTOR: Improved Protein Function Prediction by Combining Structure, Sequence and Protein–Protein Interaction Information. Nucleic Acids Res. 2017, 45, W291–W299. [Google Scholar] [CrossRef]
- Tian, W.; Chen, C.; Lei, X.; Zhao, J.; Liang, J. CASTp 3.0: Computed Atlas of Surface Topography of Proteins. Nucleic Acid Res. 2018, 46, W363–W367. [Google Scholar] [CrossRef]
- Jurrus, E.; Engel, D.; Star, K.; Monson, K.; Brandi, J.; Felberg, L.E.; Brookes, D.H.; Wilson, L.; Chen, J.; Liles, K.; et al. Improvements to the APBS Biomolecular Solvation Software Suite. Protein Sci. 2018, 27, 112–128. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A Visualization System for Exploratory Research and Analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef]
- Schutz, C.N.; Warshel, A. What Are the Dielectric Constants of Proteins and How to Validate Electrostatic Models? Proteins 2001, 44, 400–417. [Google Scholar] [CrossRef] [PubMed]
- Gorham, R.D., Jr.; Kieslich, C.A.; Morikis, D. Electrostatic Clustering and Free Energy Calculations Provide a Foundation for Protein Design and Optimization. Ann. Biomed. Eng. 2010, 39, 1252–1263. [Google Scholar] [CrossRef] [PubMed]
- Guo, T.; Gong, L.-C.; Sui, S.-F. An Electrostatically Preferred Lateral Orientation of SNARE Complex Suggests Novel Mechanisms for Driving Membrane Fusion. PLoS ONE 2010, 5, e8900. [Google Scholar] [CrossRef]
- Dobrijevic, D.; Benhamou, L.; Aliev, A.E.; Mendez-Sanchez, D.; Dawson, N.; Baud, D.; Tappertzhofen, N.; Moody, T.S.; Orengo, C.A.; Hailes, H.C.; et al. Metagenomic ene-reductases for the bioreduction of sterically challenging enones. RSC Adv. 2019, 9, 36608–36614. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Specific Activity (U/mg) | ||||
|---|---|---|---|---|
| Substrate | AnOYE2 | AnOYE8 | BfOYE1 | BfOYE4 |
| O2 | 0.37 ± 0.07 | 0.28 ± 0.07 | 0.32 ± 0.00 | 0.17 ± 0.00 |
 1 | N.D. | 1.1 ± 0.08 | 3.8 ± 0.22 | 2.2 ± 0.19 |
 2 | N.D. | 0.7 ± 0.08 | 10.3 ± 0.00 | 3.3 ± 0.00 |
 3 | N.D. | 0.8 ± 0.08 | 5.7 ± 0.37 | 1.4 ± 0.31 |
 4 | N.D. | N.D. | 10.3 ± 0.30 | N.D. |
 5 | 16.6 ± 0.53 | N.D. | 11.3 ± 0.30 | 22.5 ± 0.25 |
 6 | 1.5 ± 0.07 | 1.0 ± 0.00 | 7.5 ± 0.60 | 0.9 ± 0.06 |
 7 | N.D. | 1.0 ± 0.08 | 9.4 ± 0.50 | 0.8 ± 0.13 |
 8 | N.D. | 1.2 ± 0.00 | 4.7 ± 0.15 | 23.4 ± 1.87 |
| Substrate | AnOYE2 | BfOYE1 | BfOYE4 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| KM (mM) | kcat (s−1) | kcat/KM (mM−1 s−1) | KM (mM) | kcat (s−1) | kcat/KM (mM−1 s−1) | KM (mM) | kcat (s−1) | kcat/KM (mM−1 s−1) | |
| NADPH 2 | 0.037 ± 0.01 | 8.00 ± 0.40 | 216.2 | 0.025 ± 0.00 | 5.83 ± 0.35 | 233.2 | 0.13 ± 0.02 | 89.22 ± 3.37 | 686.3 |
| 1 | N.M. | N.M. | N.M. | 2.82 ± 0.36 | 5.98 ± 0.21 | 2.1 | 1.07 ± 0.12 | 1.06 ± 0.03 | 1.0 |
| 2 | N.M. | N.M. | N.M. | 0.44 ± 0.06 | 5.00 ± 0.16 | 11.4 | 0.65 ± 0.09 | 2.57 ± 0.07 | 3.9 |
| 3 | N.M. | N.M. | N.M. | 0.44 ± 0.06 | 4.13 ± 0.12 | 9.4 | 1.83 ± 0.18 | 0.56 ± 0.013 | 0.3 |
| 4 | N.M. | N.M. | N.M. | 2.14 ± 0.28 | 6.32 ± 0.28 | 2.9 | N.M. | N.M. | N.M. |
| 5 | 0.01 ± 0.00 | 7.38 ± 0.66 | 738.0 | 0.01 ± 0.00 | 9.91 ± 0.29 | 991.0 | 0.07 ± 0.01 | 36.3 ± 1.12 | 518.6 |
| 6 | N.M. | N.M. | N.M. | 0.07 ± 0.01 | 7.84 ± 0.01 | 112.0 | N.M. | N.M. | N.M. |
| 7 | N.M. | N.M. | N.M. | 0.45 ± 0.06 | 10.6 ± 0.47 | 23.6 | N.M. | N.M. | N.M. |
| 8 | N.M. | N.M. | N.M. | N.D. | N.D. | N.D. | 2.72 ± 0.50 | 22.6 ± 1.14 | 8.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Robescu, M.S.; Loprete, G.; Gasparotto, M.; Vascon, F.; Filippini, F.; Cendron, L.; Bergantino, E. The Family Keeps on Growing: Four Novel Fungal OYEs Characterized. Int. J. Mol. Sci. 2022, 23, 3050. https://doi.org/10.3390/ijms23063050
Robescu MS, Loprete G, Gasparotto M, Vascon F, Filippini F, Cendron L, Bergantino E. The Family Keeps on Growing: Four Novel Fungal OYEs Characterized. International Journal of Molecular Sciences. 2022; 23(6):3050. https://doi.org/10.3390/ijms23063050
Chicago/Turabian StyleRobescu, Marina Simona, Giovanni Loprete, Matteo Gasparotto, Filippo Vascon, Francesco Filippini, Laura Cendron, and Elisabetta Bergantino. 2022. "The Family Keeps on Growing: Four Novel Fungal OYEs Characterized" International Journal of Molecular Sciences 23, no. 6: 3050. https://doi.org/10.3390/ijms23063050
APA StyleRobescu, M. S., Loprete, G., Gasparotto, M., Vascon, F., Filippini, F., Cendron, L., & Bergantino, E. (2022). The Family Keeps on Growing: Four Novel Fungal OYEs Characterized. International Journal of Molecular Sciences, 23(6), 3050. https://doi.org/10.3390/ijms23063050