
Putrescine: A Key Metabolite Involved in Plant Development, Tolerance and Resistance Responses to Stress

 , , ,
, , ,
Abstract
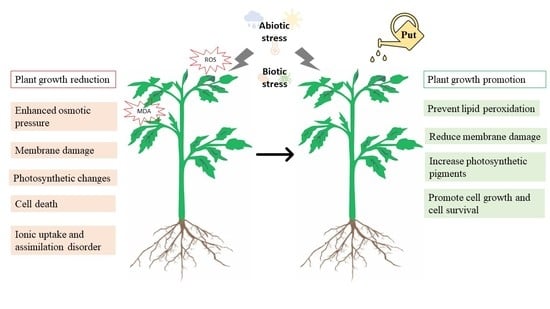
1. Introduction
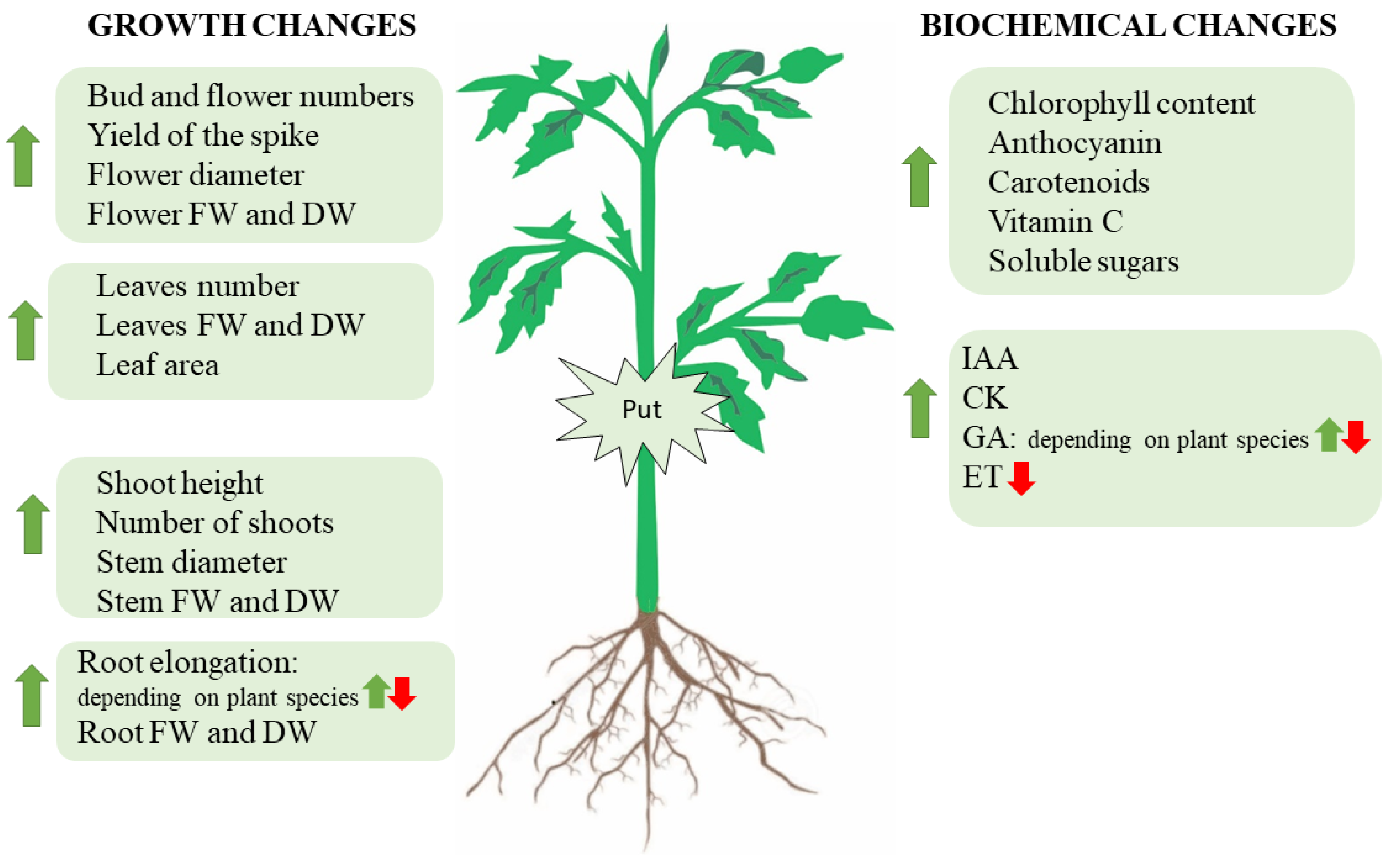
2. Plant Growth Responses
3. Tolerance Responses to Abiotic Stresses
4. Resistance Responses to Biotic Stresses
5. Conclusions and Perspectives
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alcázar, R.; Marco, F.; Cuevas, J.C.; Patron, M.; Ferrando, A.; Carrasco, P.; Tiburcio, A.F.; Altabella, T. Involvement of Polyamines in Plant Response to Abiotic Stress. Biotechnol. Lett. 2006, 28, 1867–1876. [Google Scholar] [CrossRef] [PubMed]
- Groppa, M.D.; Benavides, M.P. Polyamines and Abiotic Stress: Recent Advances. Amino Acids 2008, 34, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Kaur-Sawhney, R.; Tiburcio, A.F.; Altabella, T.; Galston, A.W. Polyamines in Plants: An Overview. J. Cell Mol. Biol. 2003, 2, 1–12. [Google Scholar]
- Chen, D.; Shao, Q.; Yin, L.; Younis, A.; Zheng, B. Polyamine Function in Plants: Metabolism, Regulation on Development, and Roles in Abiotic Stress Responses. Front. Plant Sci. 2019, 9, 1945. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.W.; Watanabe, K.; Murayama, C.; Izawa, S.; Niitsu, M.; Michael, A.J.; Berberich, T.; Kusano, T. Polyamine Oxidase5 Regulates Arabidopsis Growth through Thermospermine Oxidase Activity. Plant Physiol. 2014, 165, 1575–1590. [Google Scholar] [CrossRef]
- Sobieszczuk-Nowicka, E. Polyamine Catabolism Adds Fuel to Leaf Senescence. Amino Acids 2017, 49, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Regla-Márquez, C.F.; Canto-Flick, A.; Avilés-Viñas, S.A.; Valle-Gough, R.E.; Pérez-Pastrana, J.; García-Villalobos, F.J.; Santana-Buzzy, N. Cadaverine: A Common Polyamine in Zygotic Embryos and Somatic Embryos of the Species Capsicum chinense Jacq. Plant Cell Tissue Organ Cult. (PCTOC) 2016, 124, 253–264. [Google Scholar] [CrossRef]
- Nahar, K.; Hasanuzzaman, M.; Rahman, A.; Alam, M.M.; Mahmud, J.-A.; Suzuki, T.; Fujita, M. Polyamines Confer Salt Tolerance in Mung Bean (Vigna radiata L.) by Reducing Sodium Uptake, Improving Nutrient Homeostasis, Antioxidant Defense, and Methylglyoxal Detoxification Systems. Front. Plant Sci. 2016, 7, 1104. [Google Scholar] [CrossRef]
- Walters, D. Resistance to Plant Pathogens: Possible Roles for Free Polyamines and Polyamine Catabolism. New Phytol. 2003, 159, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Docimo, T.; Reichelt, M.; Schneider, B.; Kai, M.; Kunert, G.; Gershenzon, J.; D’Auria, J.C. The First Step in the Biosynthesis of Cocaine in Erythroxylum Coca: The Characterization of Arginine and Ornithine Decarboxylases. Plant Mol. Biol. 2012, 78, 599–615. [Google Scholar] [CrossRef]
- Sekula, B.; Ruszkowski, M.; Malinska, M.; Dauter, Z. Structural Investigations of N-Carbamoylputrescine Amidohydrolase from Medicago Truncatula: Insights into the Ultimate Step of Putrescine Biosynthesis in Plants. Front. Plant Sci. 2016, 7, 350. [Google Scholar] [CrossRef] [PubMed]
- Hanfrey, C.; Sommer, S.; Mayer, M.J.; Burtin, D.; Michael, A.J. Arabidopsis Polyamine Biosynthesis: Absence of Ornithine Decarboxylase and the Mechanism of Arginine Decarboxylase Activity. Plant J. 2001, 27, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Soyka, S.; Heyer, A.G. Arabidopsis Knockout Mutation of ADC2 Gene Reveals Inducibility by Osmotic Stress. FEBS Lett. 1999, 458, 219–223. [Google Scholar] [CrossRef]
- Kim, S.-H.; Kim, S.-H.; Yoo, S.-J.; Min, K.-H.; Nam, S.-H.; Cho, B.H.; Yang, K.-Y. Putrescine Regulating by Stress-Responsive MAPK Cascade Contributes to Bacterial Pathogen Defense in Arabidopsis. Biochem. Biophys. Res. Commun. 2013, 437, 502–508. [Google Scholar] [CrossRef] [PubMed]
- Vuosku, J.; Karppinen, K.; Muilu-Mäkelä, R.; Kusano, T.; Sagor, G.H.M.; Avia, K.; Alakärppä, E.; Kestilä, J.; Suokas, M.; Nickolov, K.; et al. Scots Pine Aminopropyltransferases Shed New Light on Evolution of the Polyamine Biosynthesis Pathway in Seed Plants. Ann. Bot. 2018, 121, 1243–1256. [Google Scholar] [CrossRef] [PubMed]
- Knott, J.M.; Römer, P.; Sumper, M. Putative Spermine Synthases from Thalassiosira Pseudonana and Arabidopsis Thaliana Synthesize Thermospermine Rather than Spermine. FEBS Lett. 2007, 581, 3081–3086. [Google Scholar] [CrossRef] [PubMed]
- Handa, A.K.; Fatima, T.; Mattoo, A.K. Polyamines: Bio-Molecules with Diverse Functions in Plant and Human Health and Disease. Front. Chem. 2018, 6, 10. [Google Scholar] [CrossRef] [PubMed]
- Moschou, P.N.; Paschalidis, K.A.; Delis, I.D.; Andriopoulou, A.H.; Lagiotis, G.D.; Yakoumakis, D.I.; Roubelakis-Angelakis, K.A. Spermidine Exodus and Oxidation in the Apoplast Induced by Abiotic Stress Is Responsible for H2O2 Signatures That Direct Tolerance Responses in Tobacco. Plant Cell 2008, 20, 1708–1724. [Google Scholar] [CrossRef] [PubMed]
- Farvardin, A.; González-Hernández, A.I.; Llorens, E.; García-Agustín, P.; Scalschi, L.; Vicedo, B. The Apoplast: A Key Player in Plant Survival. Antioxidants 2020, 9, 604. [Google Scholar] [CrossRef] [PubMed]
- Wojtasik, W.; Kulma, A.; Namysł, K.; Preisner, M.; Szopa, J. Polyamine Metabolism in Flax in Response to Treatment with Pathogenic and Non–Pathogenic Fusarium Strains. Front. Plant Sci. 2015, 6, 291. [Google Scholar] [CrossRef] [PubMed]
- Grossi, M.; Phanstiel, O.; Rippe, C.; Swärd, K.; Alajbegovic, A.; Albinsson, S.; Forte, A.; Persson, L.; Hellstrand, P.; Nilsson, B.-O. Inhibition of Polyamine Uptake Potentiates the Anti-Proliferative Effect of Polyamine Synthesis Inhibition and Preserves the Contractile Phenotype of Vascular Smooth Muscle Cells. J. Cell. Physiol. 2016, 231, 1334–1342. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, A.; Shim, I.-S.; Fujihara, S. Inhibition of Putrescine Biosynthesis Enhanced Salt Stress Sensitivity and Decreased Spermidine Content in Rice Seedlings. Biol. Plant. 2017, 61, 385–388. [Google Scholar] [CrossRef]
- Hashem, A.M.; Moore, S.; Chen, S.; Hu, C.; Zhao, Q.; Elesawi, I.E.; Feng, Y.; Topping, J.F.; Liu, J.; Lindsey, K.; et al. Putrescine Depletion Affects Arabidopsis Root Meristem Size by Modulating Auxin and Cytokinin Signaling and ROS Accumulation. Int. J. Mol. Sci. 2021, 22, 4094. [Google Scholar] [CrossRef]
- He, L.; Nada, K.; Kasukabe, Y.; Tachibana, S. Enhanced Susceptibility of Photosynthesis to Low-Temperature Photoinhibition Due to Interruption of Chill-Induced Increase of S-Adenosylmethionine Decarboxylase Activity in Leaves of Spinach (Spinacia oleracea L.). Plant Cell Physiol. 2002, 43, 196–206. [Google Scholar] [CrossRef] [PubMed]
- Navakoudis, E.; Lütz, C.; Langebartels, C.; Lütz-Meindl, U.; Kotzabasis, K. Ozone Impact on the Photosynthetic Apparatus and the Protective Role of Polyamines. Biochim. Biophys. Acta (BBA) Gen. Subj. 2003, 1621, 160–169. [Google Scholar] [CrossRef]
- Fariduddin, Q.; Varshney, P.; Yusuf, M.; Ahmad, A. Polyamines: Potent Modulators of Plant Responses to Stress. J. Plant Interact. 2013, 8, 1–16. [Google Scholar] [CrossRef]
- Couée, I.; Hummel, I.; Sulmon, C.; Gouesbet, G.; El Amrani, A. Involvement of Polyamines in Root Development. Plant Cell Tissue Organ Cult. (PCTOC) 2004, 76, 1–10. [Google Scholar] [CrossRef]
- Liu, J.-H.; Wang, W.; Wu, H.; Gong, X.; Moriguchi, T. Polyamines Function in Stress Tolerance: From Synthesis to Regulation. Front. Plant Sci. 2015, 6, 827. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.S.; Ali, M.; Ahmad, M.; Siddique, K.H.M. Polyamines: Natural and Engineered Abiotic and Biotic Stress Tolerance in Plants. Biotechnol. Adv. 2011, 29, 300–311. [Google Scholar] [CrossRef] [PubMed]
- Mustafavi, S.H.; Naghdi Badi, H.; Sękara, A.; Mehrafarin, A.; Janda, T.; Ghorbanpour, M.; Rafiee, H. Polyamines and Their Possible Mechanisms Involved in Plant Physiological Processes and Elicitation of Secondary Metabolites. Acta Physiol. Plant. 2018, 40, 102. [Google Scholar] [CrossRef]
- Anwar, R.; Mattoo, A.K.; Handa, A.K. Polyamine Interactions with Plant Hormones: Crosstalk at Several Levels. In Polyamines; Springer: Tokyo, Japan, 2015; pp. 267–302. [Google Scholar]
- Alcázar, R.; Altabella, T.; Marco, F.; Bortolotti, C.; Reymond, M.; Koncz, C.; Carrasco, P.; Tiburcio, A.F. Polyamines: Molecules with Regulatory Functions in Plant Abiotic Stress Tolerance. Planta 2010, 231, 1237–1249. [Google Scholar] [CrossRef]
- Wimalasekera, R.; Villar, C.; Begum, T.; Scherer, G.F.E. COPPER AMINE OXIDASE1 (CuAO1) of Arabidopsis Thaliana Contributes to Abscisic Acid-and Polyamine-Induced Nitric Oxide Biosynthesis and Abscisic Acid Signal Transduction. Mol. Plant 2011, 4, 663–678. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, S.; Wani, K.I.; Naeem, M.; Khan, M.M.A.; Aftab, T. Cellular Responses, Osmotic Adjustments, and Role of Osmolytes in Providing Salt Stress Resilience in Higher Plants: Polyamines and Nitric Oxide Crosstalk. J. Plant Growth Regul. 2022, 7, 347. [Google Scholar] [CrossRef]
- Khanna, K.; Sharma, N.; Kour, S.; Ali, M.; Ohri, P.; Bhardwaj, R. Hydrogen Sulfide: A Robust Combatant against Abiotic Stresses in Plants. Hydrogen 2021, 2, 319–342. [Google Scholar] [CrossRef]
- Deng, G.; Zhou, L.; Wang, Y.; Zhang, G.; Chen, X. Hydrogen Sulfide Acts Downstream of Jasmonic Acid to Inhibit Stomatal Development in Arabidopsis. Planta 2020, 251, 42. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Huo, J.; Liao, W. Hydrogen Sulfide: Roles in Plant Abiotic Stress Response and Crosstalk with Other Signals. Plant Sci. 2021, 302, 110733. [Google Scholar] [CrossRef] [PubMed]
- Minocha, R.; Majumdar, R.; Minocha, S.C. Polyamines and Abiotic Stress in Plants: A Complex Relationship1. Front. Plant Sci. 2014, 5, 175. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Xing, S.T.; Sun, X. Effects of Polyamines on Hormones Contents and the Relationship with the Flower Bud Differentiation in Chrysanthemum. Plant Physiol. J. 2014, 50, 1195–1202. [Google Scholar]
- Cai, Q.; Zhang, J.; Guo, C.; Al, E. Reviews of the Physiological Roles and Molecular Biology of Polyamines in Higher Plants. J. Fujian Educ. Coll. 2006, 7, 118–124. [Google Scholar]
- Zhao, W.; Sun, G.; Li, S. Polyamines and Plant Stress Resistance. J. South. Agric. 2004, 35, 443–447. [Google Scholar]
- Imai, A.; Akiyama, T.; Kato, T.; Sato, S.; Tabata, S.; Yamamoto, K.T.; Takahashi, T. Spermine Is Not Essential for Survival of Arabidopsis. FEBS Lett. 2004, 556, 148–152. [Google Scholar] [CrossRef]
- Kusano, T.; Berberich, T.; Tateda, C.; Takahashi, Y. Polyamines: Essential Factors for Growth and Survival. Planta 2008, 228, 367–381. [Google Scholar] [CrossRef]
- Pál, M.; Szalai, G.; Janda, T. Speculation: Polyamines Are Important in Abiotic Stress Signaling. Plant Sci. 2015, 237, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Palavan-Ünsal, N. Polyamine Metabolism in the Roots of Phaseolus vulgaris Interaction of the Inhibitors of Polyamine Biosynthesis with Putrescine in Growth and Polyamine Biosynthesis. Plant Cell Physiol. 1987, 28, 565–572. [Google Scholar]
- Lee, T.-M. Polyamine Regulation of Growth and Chilling Tolerance of Rice (Oryza sativa L.) Roots Cultured in Vitro. Plant Sci. 1997, 122, 111–117. [Google Scholar] [CrossRef]
- Tarenghi, E.; Carré, M.; Martin-Tanguy, J. Effects of Inhibitors of Polyamine Biosynthesis and of Polyamines on Strawberry Microcutting Growth and Development. Plant Cell Tissue Organ Cult. (PCTOC) 1995, 42, 47–55. [Google Scholar] [CrossRef]
- Wu, Q.S.; Zou, Y.N.; He, X.H. Exogenous Putrescine, Not Spermine or Spermidine, Enhances Root Mycorrhizal Development and Plant Growth of Trifoliate Orange (Poncirus trifoliata) Seedlings. Int. J. Agric. Biol. 2010, 12, 576–580. [Google Scholar]
- Ben-Hayyim, G.; Martin-Tanguy, J.; Tepfer, D. Changing Root and Shoot Architecture with the RolA Gene from Agrobacterium Rhizogenes: Interactions with Gibberellic Acid and Polyamine Metabolism. Physiol. Plant. 1996, 96, 237–243. [Google Scholar] [CrossRef]
- Tang, W.; Newton, R.J. Polyamines Promote Root Elongation and Growth by Increasing Root Cell Division in Regenerated Virginia Pine (Pinus virginiana Mill.) Plantlets. Plant Cell Rep. 2005, 24, 581–589. [Google Scholar] [CrossRef]
- Sánchez-Rangel, D.; Chávez-Martínez, A.I.; Rodríguez-Hernández, A.A.; Maruri-López, I.; Urano, K.; Shinozaki, K.; Jiménez-Bremont, J.F. Simultaneous Silencing of Two Arginine Decarboxylase Genes Alters Development in Arabidopsis. Front. Plant Sci. 2016, 7, 300. [Google Scholar] [CrossRef] [PubMed]
- Matam, P.; Parvatam, G. Putrescine and Polyamine Inhibitors in Culture Medium Alter in Vitro Rooting Response of Decalepis Hamiltonii Wight & Arn. Plant Cell Tissue Organ Cult. (PCTOC) 2017, 128, 273–282. [Google Scholar]
- Badawy, E.S.M.; Kandil, M.M.; Habib, A.M.; El-Sayed, I.M. Influence of Diatomite, Putrescine and Alpha-Tocopherol on Some Vegetative Growth and Flowering of Antirrhinum majus L. Plants. J. Hortic. Sci. Ornam. Plants 2015, 7, 7–18. [Google Scholar]
- Danaee, E.; Abdossi, V. Phytochemical And Morphophysiological Responses In Basil (Ocimum basilicum L.) Plant To Application Of Polyamines. J. Med. Plants 2019, 18, 125–133. [Google Scholar]
- Houdusse, F.; Zamarreño, A.M.; Garnica, M.; García-Mina, J. The Importance of Nitrate in Ameliorating the Effects of Ammonium and Urea Nutrition on Plant Development: The Relationships with Free Polyamines and Plant Proline Contents. Funct. Plant Biol. 2005, 32, 1057. [Google Scholar] [CrossRef] [PubMed]
- Awasthi, V.; Gautam, I.K.; Sengar, R.S.; Garg, S.K. Influence of Putrescine on Enzymes of Ammonium Assimilation in Maize Seedling. Am. J. Plant Sci. 2013, 04, 297–301. [Google Scholar] [CrossRef]
- González-Hernández, A.I.; Scalschi, L.; Troncho, P.; García-Agustín, P.; Camañes, G. Putrescine Biosynthetic Pathways Modulate Root Growth Differently in Tomato Seedlings Grown under Different N Sources. J. Plant Physiol. 2022, 268, 153560. [Google Scholar] [CrossRef]
- Nahed, G.A.A.; Lobna, S.T.; Soad, M.I. Some Studies on The Effect Of Putrescine, Ascorbic Acid And Thiamine On Growth, Flowering And Some Chemical Constituents Of Gladiolus Plants At Nubaria. Ozean J. Appl. Sci. 2009, 2, 169–179. [Google Scholar]
- Youssef, A.A.; Mahgoub, M.H.; Talaat, I.M. Physiological and Biochemical Aspects of Matthiola incana L. Plants under the Effect of Putrescine and Kinetin Treatments. Egypt. J. Appl. Sci. 2004, 19, 492–510. [Google Scholar]
- Talaat, I.M.; Bekheta, M.A.; Mahgoubi, M.H. Physiological Response of Periwinkle Plants (Catharanthus roseus L.) to Tryptophan and Putrescine. Int. J. Agric. Biol. 2005, 7, 210–213. [Google Scholar]
- Mahgoub, M.; el Aziz, N.G.A.; Mazhar, A.M.A. Response of Dahlia pinnata L. Plant to Foliar Spray with Putrescine and Thiamine on Growth, Flowering and Photosynthetic Pigments. Am. Eurasian J. Agric. Environ. Sci. 2011, 10, 769–775. [Google Scholar]
- Dastyaran, M.; Hosseini Farahi, M. The Effect of Humic Acid and Putrescin on Vegetative Properties and Vase Life of Rose in a Soilless System. J. Soil Plant Interact. 2014, 5, 241–250. [Google Scholar]
- Yadav, V.; Singh, N.B.; Singh, H.; Singh, A.; Hussain, I. Putrescine Affects Tomato Growth and Response of Antioxidant Defense System Due to Exposure to Cinnamic Acid. Int. J. Veg. Sci. 2019, 25, 259–277. [Google Scholar] [CrossRef]
- Mora, V.; Bacaicoa, E.; Zamarreño, A.-M.; Aguirre, E.; Garnica, M.; Fuentes, M.; García-Mina, J.-M. Action of Humic Acid on Promotion of Cucumber Shoot Growth Involves Nitrate-Related Changes Associated with the Root-to-Shoot Distribution of Cytokinins, Polyamines and Mineral Nutrients. J. Plant Physiol. 2010, 167, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Krizek, D.T.; Kramer, G.F.; Mirecki, R.M. Influence of UV-B Radiation and Putrescine on Shoot and Root Growth of Cucumber Seedlings Grown in Nutrient Solution. J. Plant Nutr. 1997, 20, 613–623. [Google Scholar] [CrossRef]
- Farooq, M.; Basra, S.M.A.; Rehman, H.; Hussain, M. Seed Priming with Polyamines Improves the Germination and Early Seedling Growth in Fine Rice. J. New Seeds 2008, 9, 145–155. [Google Scholar] [CrossRef]
- Fazilati, M.; Hossein Forghani, A.; Fazilati, M. The Role of Polyamine to Increasing Growth of Plant: As a Key Factor in Health Crisis. Int. J. Health Syst. Disaster Manag. 2015, 3, 89–94. [Google Scholar]
- Bais, H.P.; Sudha, G.S.; Ravishankar, G.A. Putrescine and Silver Nitrate Influences Shoot Multiplication, In Vitro Flowering and Endogenous Titers of Polyamines in Cichorium intybus L. Cv. Lucknow Local. J. Plant Growth Regul. 2000, 19, 238–248. [Google Scholar] [CrossRef]
- Badawy, E.M.; Kandil, M.M.; Mahgoub, M.; Shanan, N.; Hegazi, N. Chemical Constituents of Celosia argentea var. Cristata L. Plants as Affected by Foliar Application of Putrescine and Alpha-Tocopherol. Int. J. ChemTech Res. 2015, 8, 464–470. [Google Scholar]
- Yousefi, F.; Jabbarzadeh, Z.; Amiri, J.; Rasouli-Sadaghiani, M.H. Response of Roses (Rosa hybrida L. ‘Herbert Stevens’) to Foliar Application of Polyamines on Root Development, Flowering, Photosynthetic Pigments, Antioxidant Enzymes Activity and NPK. Sci. Rep. 2019, 9, 16025. [Google Scholar] [CrossRef]
- Kandil, M.M.; Ibrahaim, S.M.M.; El-Hanafy, S.H.; El-Sabwah, M.M. Effect of Putrescine and Uniconazole on Some Flowering Characteristics, and Some Chemical Constituents of Salvia splendens F. Plant. Int. J. ChemTech Res. 2015, 8, 174–186. [Google Scholar]
- Amin, A.A.; Gharib, F.A.; Abouziena, H.F.; Dawood, M.G. Role of Indole-3-Butyric Acid or/and Putrescine in Improving Productivity of Chickpea (Cicer arientinum L.) Plants. Pak. J. Biol. Sci. 2013, 16, 1894–1903. [Google Scholar] [CrossRef] [PubMed]
- El-Bassiouny, H.M.; Mostafa, H.A.; El-Khawas, S.A.; Hassanein, R.A.; Khalil, S.I.; Abd El-Monem, A.A. Physiological Responses of Wheat Plant to Foliar Treatments with Arginine or Putrescine. Aust. J. Basic Appl. Sci. 2008, 2, 1390–1403. [Google Scholar]
- El-Bassiouny, H.M.S.; Bekheta, M.A. Role of Putrescine on Growth, Regulation of Stomatal Aperture, Ionic Contents and Yield by Two Wheat Cultivars under Salinity Stress. Egypt. J. Physiol. Sci. 2001, 23, 235–258. [Google Scholar]
- Ioannidis, N.E.; Sfichi-Duke, L.; Kotzabasis, K. Polyamines Stimulate Non-Photochemical Quenching of Chlorophyll a Fluorescence in Scenedesmus Obliquus. Photosynth. Res. 2011, 107, 169–175. [Google Scholar] [CrossRef]
- Singh, T.; Bala, M. Effect of Putrescine and Benzyl Adenine on Growth, Flowering and Post-Harvest Keeping Quality Parameters in Chrysanthemum (Chrysanthemum morifolium Ramat.). J. Hortic. Sci. 2020, 15, 191–196. [Google Scholar] [CrossRef]
- Li, C.; Pei, Z.X.; Gan, L.Y. Effects of Photoperiod on Flowering and Polyamine Contents of Nobile-Type Dendrobium. Zhiwu Shengli Xuebao/Plant Physiol. J. 2014, 50, 1167–1170. [Google Scholar]
- Mahgoub, M.H.; El-Ghorab, A.H.; Bekheta, M. Effect of Some Bioregulators on the Endogenous Phytohormones, Chemical Composition, Essential Oil and Its Antioxidant Activity of Carnation (Dianthus caryophyllus L.). J. Agric. Sci. Mansoura Univ. 2006, 31, 4229–4245. [Google Scholar]
- Iman, M.E.S.; Kandil, M.M.; Badawy, M.E.S.; Abdalla, M.A.E.F.; Mahgoub, H.M.; Habib, M.A. Effect of Diatomite, Putrescine and Alpha-Tocopherol on Flower Characters and Anatomical Flower Bud Structure of Antirhinum majus L. Plant. Middle East J. Agric. Res. 2018, 7, 1747–1755. [Google Scholar]
- Kazemi, M. Influence of Foliar Application of 5-Sulfosalicylic Acid, Malic Acid, Putrescine and Potassium Nitrate on Vegetative Growth and Reproductive Characteristics of Strawberry Cv. ‘Selva.’ J. Biol. Environ. Sci. 2013, 7, 93–101. [Google Scholar]
- Bani Assadi, F.; Safari, V.R.; Maghsoudi Mod, A.A. Effect of Putrescine and Salinity on the Morphological, Biochemical, and Pigments of English Marigold Plant (Calendula officinalis L.). J. Sci. Technol. Greenh. Cult. 2015, 6, 125–133. [Google Scholar]
- Killiny, N.; Nehela, Y. Citrus Polyamines: Structure, Biosynthesis, and Physiological Functions. Plants 2020, 9, 426. [Google Scholar] [CrossRef] [PubMed]
- Bekheta, M.A.; El-Bassiouny, H.M.S. Responses of Two Wheat Cultivars Grown under Salinity Stress to Putrescine Treatment. J. Agric. Sci. 2005, 30, 4505–4521. [Google Scholar]
- Alcázar, R.; García-Martínez, J.L.; Cuevas, J.C.; Tiburcio, A.F.; Altabella, T. Overexpression of ADC2 in Arabidopsis Induces Dwarfism and Late-Flowering through GA Deficiency. Plant J. 2005, 43, 425–436. [Google Scholar] [CrossRef] [PubMed]
- Sobieszczuk-Nowicka, E.; Rorat, T.; Legocka, J. Polyamine Metabolism and S-Adenosylmethionine Decarboxylase Gene Expression during the Cytokinin-Stimulated Greening Process. Acta Physiol. Plant. 2007, 29, 495–502. [Google Scholar] [CrossRef]
- Legocka, J.; Żarnowska, A. Role of Polyamines in the Cytokinin-Dependent Physiological Processes II. Modulation of Polyamine Levels during Cytokinin-Stimulated Expansion of Cucumber Cotyledons. Acta Physiol. Plant. 2000, 22, 395–401. [Google Scholar] [CrossRef]
- Yu, Y.; Jin, C.; Sun, C.; Wang, J.; Ye, Y.; Zhou, W.; Lu, L.; Lin, X. Inhibition of Ethylene Production by Putrescine Alleviates Aluminium-Induced Root Inhibition in Wheat Plants. Sci. Rep. 2016, 6, 18888. [Google Scholar] [CrossRef]
- Takahashi, Y.; Cong, R.; Sagor, G.H.M.; Niitsu, M.; Berberich, T.; Kusano, T. Characterization of Five Polyamine Oxidase Isoforms in Arabidopsis thaliana. Plant Cell Rep. 2010, 29, 955–965. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Polyamines and Abiotic Stress Tolerance in Plants. Plant Signal. Behav. 2010, 5, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.-H.; Kitashiba, H.; Wang, J.; Ban, Y.; Moriguchi, T. Polyamines and Their Ability to Provide Environmental Stress Tolerance to Plants. Plant Biotechnol. 2007, 24, 117–126. [Google Scholar] [CrossRef]
- Hummel, I.; Bourdais, G.; Gouesbet, G.; Couée, I.; Malmberg, R.L.; el Amrani, A. Differential Gene Expression of ARGININE DECARBOXYLASE ADC1 and ADC2 in Arabidopsis thaliana: Characterization of Transcriptional Regulation during Seed Germination and Seedling Development. New Phytol. 2004, 163, 519–531. [Google Scholar] [CrossRef] [PubMed]
- Perez-Amador, M.A.; Leon, J.; Green, P.J.; Carbonell, J. Induction of the Arginine Decarboxylase ADC2 Gene Provides Evidence for the Involvement of Polyamines in the Wound Response in Arabidopsis. Plant Physiol. 2002, 130, 1454–1463. [Google Scholar] [CrossRef] [PubMed]
- Urano, K.; Yoshiba, Y.; Nanjo, T.; Igarashi, Y.; Seki, M.; Sekiguchi, F.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Characterization of Arabidopsis Genes Involved in Biosynthesis of Polyamines in Abiotic Stress Responses and Developmental Stages. Plant Cell Environ. 2003, 26, 1917–1926. [Google Scholar] [CrossRef]
- Armengaud, P.; Breitling, R.; Amtmann, A. The Potassium-Dependent Transcriptome of Arabidopsis Reveals a Prominent Role of Jasmonic Acid in Nutrient Signaling. Plant Physiol. 2004, 136, 2556–2576. [Google Scholar] [CrossRef]
- Alcázar, R.; Cuevas, J.C.; Patron, M.; Altabella, T.; Tiburcio, A.F. Abscisic Acid Modulates Polyamine Metabolism under Water Stress in Arabidopsis Thaliana. Physiol. Plant. 2006, 128, 448–455. [Google Scholar] [CrossRef]
- Parihar, P.; Singh, S.; Singh, R.; Singh, V.P.; Prasad, S.M. Effect of Salinity Stress on Plants and Its Tolerance Strategies: A Review. Environ. Sci. Pollut. Res. 2015, 22, 4056–4075. [Google Scholar] [CrossRef] [PubMed]
- Tanveer, M.; Shabala, S. Targeting Redox Regulatory Mechanisms for Salinity Stress Tolerance in Crops. In Salinity Responses and Tolerance in Plants, Volume 1; Springer International Publishing: Cham, Switzerland, 2018; pp. 213–234. [Google Scholar]
- Zapata, P.J.; Serrano, M.; Pretel, M.T.; Amorós, A.; Botella, M.Á. Polyamines and Ethylene Changes during Germination of Different Plant Species under Salinity. Plant Sci. 2004, 167, 781–788. [Google Scholar] [CrossRef]
- Antoniou, C.; Zarza, X.; Gohari, G.; Panahirad, S.; Filippou, P.; Tiburcio, A.F.; Fotopoulos, V. Involvement of Polyamine Metabolism in the Response of Medicago truncatula Genotypes to Salt Stress. Plants 2021, 10, 269. [Google Scholar] [CrossRef] [PubMed]
- Krishnamurthy, R.; Bhagwat, K.A. Polyamines as Modulators of Salt Tolerance in Rice Cultivars. Plant Physiol. 1989, 91, 500–504. [Google Scholar] [CrossRef] [PubMed]
- Santa-Cruz, A.; Acosta, M.; Rus, A.; Bolarin, M.C. Short-Term Salt Tolerance Mechanisms in Differentially Salt Tolerant Tomato Species. Plant Physiol. Biochem. 1999, 37, 65–71. [Google Scholar] [CrossRef]
- Ali, R.M. Role of Putrescine in Salt Tolerance of Atropa belladonna Plant. Plant Sci. 2000, 152, 173–179. [Google Scholar] [CrossRef]
- Legocka, J.; Kluk, A. Effect of Salt and Osmotic Stress on Changes in Polyamine Content and Arginine Decarboxylase Activity in Lupinus Luteus Seedlings. J. Plant Physiol. 2005, 162, 662–668. [Google Scholar] [CrossRef]
- Quinet, M.; Ndayiragije, A.; Lefevre, I.; Lambillotte, B.; Dupont-Gillain, C.C.; Lutts, S. Putrescine Differently Influences the Effect of Salt Stress on Polyamine Metabolism and Ethylene Synthesis in Rice Cultivars Differing in Salt Resistance. J. Exp. Bot. 2010, 61, 2719–2733. [Google Scholar] [CrossRef] [PubMed]
- Urano, K.; Yoshiba, Y.; Nanjo, T.; Ito, T.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Arabidopsis Stress-Inducible Gene for Arginine Decarboxylase AtADC2 Is Required for Accumulation of Putrescine in Salt Tolerance. Biochem. Biophys. Res. Commun. 2004, 313, 369–375. [Google Scholar] [CrossRef]
- Xiong, F.; Liao, J.; Ma, Y.; Wang, Y.; Fang, W.; Zhu, X. The Protective Effect of Exogenous Putrescine in the Response of Tea Plants (Camellia sinensis) to Salt Stress. HortScience 2018, 53, 1640–1646. [Google Scholar] [CrossRef]
- Ekinci, M.; Yildirim, E.; Dursun, A.; Mohamedsrajaden, N. Putresin, Spermin ve Spermidin Uygulamalarının Biber (Capsicum annum L.) Fidesinde Tuz Stresi Zararını Hafifletici Etkisi. Yüzüncü Yıl Üniversitesi Tarım Bilimleri Derg. 2019, 29, 290–299. [Google Scholar] [CrossRef]
- Esfandiari Ghalati, R.; Shamili, M.; Homaei, A. Effect of Putrescine on Biochemical and Physiological Characteristics of Guava (Psidium guajava L.) Seedlings under Salt Stress. Sci. Hortic. 2020, 261, 108961. [Google Scholar] [CrossRef]
- Shu, S.; Yuan, Y.; Chen, J.; Sun, J.; Zhang, W.; Tang, Y.; Zhong, M.; Guo, S. The Role of Putrescine in the Regulation of Proteins and Fatty Acids of Thylakoid Membranes under Salt Stress. Sci. Rep. 2015, 5, 14390. [Google Scholar] [CrossRef] [PubMed]
- Zhong, M.; Yuan, Y.; Shu, S.; Sun, J.; Guo, S.; Yuan, R.; Tang, Y. Effects of Exogenous Putrescine on Glycolysis and Krebs Cycle Metabolism in Cucumber Leaves Subjected to Salt Stress. Plant Growth Regul. 2016, 79, 319–330. [Google Scholar] [CrossRef]
- Shen, J.; Wang, Y.; Shu, S.; Jahan, M.S.; Zhong, M.; Wu, J.; Sun, J.; Guo, S. Exogenous Putrescine Regulates Leaf Starch Overaccumulation in Cucumber under Salt Stress. Sci. Hortic. 2019, 253, 99–110. [Google Scholar] [CrossRef]
- Espasandin, F.D.; Calzadilla, P.I.; Maiale, S.J.; Ruiz, O.A.; Sansberro, P.A. Overexpression of the Arginine Decarboxylase Gene Improves Tolerance to Salt Stress in Lotus Tenuis Plants. J. Plant Growth Regul. 2018, 37, 156–165. [Google Scholar] [CrossRef]
- Shi, K.; Huang, Y.Y.; Xia, X.J.; Zhang, Y.L.; Zhou, Y.H.; Yu, J.Q. Protective Role of Putrescine Against Salt Stress Is Partially Related to the Improvement of Water Relation and Nutritional Imbalance in Cucumber. J. Plant Nutr. 2008, 31, 1820–1831. [Google Scholar] [CrossRef]
- Reginato, M.A.; Abdala, G.I.; Miersch, O.; Ruíz, O.A.; Moschetti, E.; Luna, V. Changes in the Levels of Jasmonates and Free Polyamines Induced by Na2SO4 and NaCl in Roots and Leaves of the Halophyte Prosopis strombulifera. Biologia 2012, 67, 689–697. [Google Scholar] [CrossRef]
- Reinoso, H.; Sosa, L.; Reginato, M.; Luna, V. Histological Alterations Induced by Sodium Sulfate in the Vegetative Anatomy of Prosopis strombulifera (Lam.) Benth. World J. Agric. Sci. 2005, 1, 109–119. [Google Scholar]
- Altamura, M.M.; Torrigiani, P.; Capitani, F.; Scaramagli, S.; Bagni, N. De Novo Root Formation in Tobacco Thin Layers Is Affected by Inhibition of Polyamine Biosynthesis. J. Exp. Bot. 1991, 42, 1575–1582. [Google Scholar] [CrossRef]
- Gong, B.; Li, X.; Bloszies, S.; Wen, D.; Sun, S.; Wei, M.; Li, Y.; Yang, F.; Shi, Q.; Wang, X. Sodic Alkaline Stress Mitigation by Interaction of Nitric Oxide and Polyamines Involves Antioxidants and Physiological Strategies in Solanum lycopersicum. Free Radic. Biol. Med. 2014, 71, 36–48. [Google Scholar] [CrossRef] [PubMed]
- Recalde, L.; Gómez Mansur, N.M.; Cabrera, A.v.; Matayoshi, C.L.; Gallego, S.M.; Groppa, M.D.; Benavides, M.P. Unravelling Ties in the Nitrogen Network: Polyamines and Nitric Oxide Emerging as Essential Players in Signalling Roadway. Ann. Appl. Biol. 2021, 178, 192–208. [Google Scholar] [CrossRef]
- García-Jiménez, P.; Just, P.M.; Delgado, A.M.; Robaina, R.R. Transglutaminase Activity Decrease during Acclimation to Hyposaline Conditions in Marine Seaweed Grateloupia Doryphora (Rhodophyta, Halymeniaceae). J. Plant Physiol. 2007, 164, 367–370. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Donaire, M.d.P.; Giráldez, J.V.; Vanwalleghem, T. Impact of Climate Change on Agricultural Droughts in Spain. Water 2020, 12, 3214. [Google Scholar] [CrossRef]
- Hussain, H.A.; Hussain, S.; Khaliq, A.; Ashraf, U.; Anjum, S.A.; Men, S.; Wang, L. Chilling and Drought Stresses in Crop Plants: Implications, Cross Talk, and Potential Management Opportunities. Front. Plant Sci. 2018, 9, 393. [Google Scholar] [CrossRef]
- Gupta, S.; Agarwal, V.P.; Gupta, N.K. Efficacy of Putrescine and Benzyladenine on Photosynthesis and Productivity in Relation to Drought Tolerance in Wheat (Triticum aestivum L.). Physiol. Mol. Biol. Plants 2012, 18, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Wang, L.; Yang, R.; Han, Y.; Hao, J.; Liu, C.; Fan, S. Effects of Exogenous Putrescine on the Ultrastructure of and Calcium Ion Flow Rate in Lettuce Leaf Epidermal Cells under Drought Stress. Hortic. Environ. Biotechnol. 2019, 60, 479–490. [Google Scholar] [CrossRef]
- Shallan, M.A.; Hassan, H.M.M.; Namich, A.A.M.; Ibrahim, A.A. Effect of Sodium Niroprusside, Putrescine and Glycine Betaine on Alleviation of Drought Stress in Cotton Plant. J. Agric. Environ. Sci. 2012, 12, 1252–1265. [Google Scholar]
- Zeid, I.M.; Shedeed, Z.A. Response of Alfalfa to Putrescine Treatment under Drought Stress. Biol. Plant. 2006, 50, 635–640. [Google Scholar] [CrossRef]
- Abd Elbar, O.H.; Farag, R.E.; Shehata, S.A. Effect of Putrescine Application on Some Growth, Biochemical and Anatomical Characteristics of Thymus vulgaris L. under Drought Stress. Ann. Agric. Sci. 2019, 64, 129–137. [Google Scholar] [CrossRef]
- Zhao, J.; Wang, X.; Pan, X.; Jiang, Q.; Xi, Z. Exogenous Putrescine Alleviates Drought Stress by Altering Reactive Oxygen Species Scavenging and Biosynthesis of Polyamines in the Seedlings of Cabernet Sauvignon. Front. Plant Sci. 2021, 12, 767992. [Google Scholar] [CrossRef] [PubMed]
- Khosrowshahi, Z.T.; Slehi-Lisar, Y.; Ghassemi-Golezani, K.; Motafakkerazad, R. Physiological Responses of Safolew to Exogenous Putwescine Undew Watew Defcit. J. Stress Physiol. Biochem. 2018, 14, 38–48. [Google Scholar]
- Islam, M.J.; Uddin, M.J.; Hossain, M.A.; Henry, R.; Begum, M.K.; Sohel, M.A.T.; Mou, M.A.; Ahn, J.; Cheong, E.J.; Lim, Y.-S. Exogenous Putrescine Attenuates the Negative Impact of Drought Stress by Modulating Physio-Biochemical Traits and Gene Expression in Sugar Beet (Beta vulgaris L.). PLoS ONE 2022, 17, e0262099. [Google Scholar] [CrossRef] [PubMed]
- Montilla-Bascón, G.; Rubiales, D.; Hebelstrup, K.H.; Mandon, J.; Harren, F.J.M.; Cristescu, S.M.; Mur, L.A.J.; Prats, E. Reduced Nitric Oxide Levels during Drought Stress Promote Drought Tolerance in Barley and Is Associated with Elevated Polyamine Biosynthesis. Sci. Rep. 2017, 7, 13311. [Google Scholar] [CrossRef] [PubMed]
- An, Z.F.; Li, C.Y.; Zhang, L.X.; Alva, A.K. Role of Polyamines and Phospholipase D in Maize (Zea mays L.) Response to Drought Stress. S. Afr. J. Bot. 2012, 83, 145–150. [Google Scholar] [CrossRef]
- Karimi, Z. Putrescine Foliar Application Effect on Physiologic and Morphologic Characteristics of Wheat (Triticum aestivum var Sw-82-9) under Water Deficit Stress. Biol. Forum Int. J. 2016, 8, 532–539. [Google Scholar]
- Alcázar, R.; Planas, J.; Saxena, T.; Zarza, X.; Bortolotti, C.; Cuevas, J.; Bitrián, M.; Tiburcio, A.F.; Altabella, T. Putrescine Accumulation Confers Drought Tolerance in Transgenic Arabidopsis Plants Over-Expressing the Homologous Arginine Decarboxylase 2 Gene. Plant Physiol. Biochem. 2010, 48, 547–552. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi-Cheraghabadi, M.; Modarres-Sanavy, S.A.M.; Sefidkon, F.; Rashidi-Monfared, S.; Mokhtassi-Bidgoli, A. Improving Water Deficit Tolerance of Salvia officinalis L. Using Putrescine. Sci. Rep. 2021, 11, 21997. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.; Farooq, M.; Wahid, M.A.; Wahid, A. Seed Priming with Putrescine Improves the Drought Resistance of Maize Hybrids. Int. J. Agric. Biol 2014, 15, 1349–1353. [Google Scholar]
- Do, P.T.; Degenkolbe, T.; Erban, A.; Heyer, A.G.; Kopka, J.; Köhl, K.I.; Hincha, D.K.; Zuther, E. Dissecting Rice Polyamine Metabolism under Controlled Long-Term Drought Stress. PLoS ONE 2013, 8, e60325. [Google Scholar]
- Ahangir, A.; Ghotbi-Ravandi, A.A.; Rezadoost, H.; Bernard, F. Drought Tolerant Maize Cultivar Accumulates Putrescine in Roots. Rhizosphere 2020, 16, 100260. [Google Scholar] [CrossRef]
- Chen, J.; Shang, Y.-T.; Wang, W.-H.; Chen, X.-Y.; He, E.-M.; Zheng, H.-L.; Shangguan, Z. Hydrogen Sulfide-Mediated Polyamines and Sugar Changes Are Involved in Hydrogen Sulfide-Induced Drought Tolerance in Spinacia oleracea Seedlings. Front. Plant Sci. 2016, 7, 1173. [Google Scholar] [CrossRef] [PubMed]
- Pucciariello, C.; Voesenek, L.A.C.J.; Perata, P.; Sasidharan, R. Plant Responses to Flooding. Front. Plant Sci. 2014, 5, 226. [Google Scholar] [CrossRef] [PubMed]
- Herrera, A. Responses to Flooding of Plant Water Relations and Leaf Gas Exchange in Tropical Tolerant Trees of a Black-Water Wetland. Front. Plant Sci. 2013, 4, 106. [Google Scholar] [CrossRef]
- Bertani, A.; Brambilla, I.; Mapelli, S.; Reggiani, R. Elongation Growth in the Absence of Oxygen: The Rice Coleoptile. Russ. J. Plant Physiol. 1997, 44, 543–547. [Google Scholar]
- Mahdavian, M.; Sarikhani, H.; Hadadinejad, M.; Dehestani, A. Putrescine Effect on Physiological, Morphological, and Biochemical Traits of Carrizo Citrange and Volkameriana Rootstocks under Flooding Stress. Int. J. Fruit Sci. 2020, 20, 164–177. [Google Scholar] [CrossRef]
- Yiu, J.-C.; Juang, L.-D.; Fang, D.Y.-T.; Liu, C.-W.; Wu, S.-J. Exogenous Putrescine Reduces Flooding-Induced Oxidative Damage by Increasing the Antioxidant Properties of Welsh Onion. Sci. Hortic. 2009, 120, 306–314. [Google Scholar] [CrossRef]
- Bita, C.E.; Gerats, T. Plant Tolerance to High Temperature in a Changing Environment: Scientific Fundamentals and Production of Heat Stress-Tolerant Crops. Front. Plant Sci. 2013, 4, 273. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Roychowdhury, R.; Fujita, M. Physiological, Biochemical, and Molecular Mechanisms of Heat Stress Tolerance in Plants. Int. J. Mol. Sci. 2013, 14, 9643–9684. [Google Scholar] [CrossRef] [PubMed]
- Mitra, R.; Bhatia, C.R. Bioenergetic Cost of Heat Tolerance in Wheat Crop. Curr. Sci. 2008, 94, 10491053. [Google Scholar]
- Kusano, T.; Yamaguchi, K.; Berberich, T.; Takahashi, Y. Advances in Polyamine Research in 2007. J. Plant Res. 2007, 120, 345–350. [Google Scholar] [CrossRef]
- Hassanein, R.A.; El-Khawas, S.A.; Ibrahim, S.K.; El-Bassiouny, H.M.; Mostafa, H.A.; Abd El-Monem, A.A. Improving the thermo tolerance of wheat plant by foliar application of arginine or putrescine. Pak. J. Bot. 2013, 45, 111–118. [Google Scholar]
- Mostafa, H.A.M.; Hassanein, R.A.; Khalil, S.I.; El-Khawas, S.A.; El-Bassiouny, H.M.S.; El-Monem, A.A.A. Effect of Arginine or Putrescine on Growth, Yield and Yield Components of Late Sowing Wheat. J. Appl. Sci. Res. 2010, 54, 177–183. [Google Scholar]
- Collado-González, J.; Piñero, M.C.; Otálora, G.; López-Marín, J.; Amor, F.M. del The Effect of Foliar Putrescine Application, Ammonium Exposure, and Heat Stress on Antioxidant Compounds in Cauliflower Waste. Antioxidants 2021, 10, 707. [Google Scholar] [CrossRef] [PubMed]
- Amooaghaie, R.; Moghym, S. Effect of Polyamines on Thermotolerance and Membrane Stability of Soybean Seedling. Afr. J. Biotechnol. 2011, 10, 9673–9676. [Google Scholar]
- Khalil, S.I.; El-Monem, H.M.S.; Hassanein, R.A.; Mostafa, H.A.; El-Khawas, S.A.; Abd El-Monem, A.A. Antioxidant Defense System in Heat Shocked Wheat Plants Previously Treated with Arginine or Putrescine. Aust. J. Basic Appl. Sci. 2009, 3, 1517–1526. [Google Scholar]
- Königshofer, H.; Lechner, S. Are Polyamines Involved in the Synthesis of Heat-Shock Proteins in Cell Suspension Cultures of Tobacco and Alfalfa in Response to High-Temperature Stress? Plant Physiol. Biochem. 2002, 40, 51–59. [Google Scholar] [CrossRef]
- Jahan, M.S.; Shu, S.; Wang, Y.; Chen, Z.; He, M.; Tao, M.; Sun, J.; Guo, S. Melatonin Alleviates Heat-Induced Damage of Tomato Seedlings by Balancing Redox Homeostasis and Modulating Polyamine and Nitric Oxide Biosynthesis. BMC Plant Biol. 2019, 19, 414. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.R.; Singh, G.P.; Sharma, S.K.; Singh, K.; Goswami, S.; Rai, R.D. Molecular Cloning of HSP17 Gene (SHSP) and Their Differential Expression under Exogenous Putrescine and Heat Shock in Wheat (Triticum aestivum). Afr. J. Biotechnol. 2012, 11, 16800–16808. [Google Scholar]
- Marco, F.; Alcázar, R.; Tiburcio, A.F.; Carrasco, P. Interactions between Polyamines and Abiotic Stress Pathway Responses Unraveled by Transcriptome Analysis of Polyamine Overproducers. OMICS J. Integr. Biol. 2011, 15, 775–781. [Google Scholar] [CrossRef] [PubMed]
- Theocharis, A.; Clément, C.; Barka, E.A. Physiological and Molecular Changes in Plants Grown at Low Temperatures. Planta 2012, 235, 1091–1105. [Google Scholar] [CrossRef]
- Cuevas, J.C.; López-Cobollo, R.; Alcázar, R.; Zarza, X.; Koncz, C.; Altabella, T.; Salinas, J.; Tiburcio, A.F.; Ferrando, A. Putrescine Is Involved in Arabidopsis Freezing Tolerance and Cold Acclimation by Regulating Abscisic Acid Levels in Response to Low Temperature. Plant Physiol. 2008, 148, 1094–1105. [Google Scholar] [CrossRef]
- Song, Y.; Diao, Q.; Qi, H. Putrescine Enhances Chilling Tolerance of Tomato (Lycopersicon esculentum Mill.) through Modulating Antioxidant Systems. Acta Physiol. Plant. 2014, 36, 3013–3027. [Google Scholar] [CrossRef]
- Diao, Q.; Song, Y.; Shi, D.; Qi, H. Interaction of Polyamines, Abscisic Acid, Nitric Oxide, and Hydrogen Peroxide under Chilling Stress in Tomato (Lycopersicon esculentum Mill.) Seedlings. Front. Plant Sci. 2017, 8, 203. [Google Scholar] [CrossRef]
- Hummel, I.; Couée, I.; el Amrani, A.; Martin-Tanguy, J.; Hennion, F. Involvement of Polyamines in Root Development at Low Temperature in the Subantarctic Cruciferous Species Pringlea antiscorbutica. J. Exp. Bot. 2002, 53, 1463–1473. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yin, X.; Yang, Y.; Lv, Y.; Li, Y.; Yang, D.; Yue, Y.; Yang, Y. BrrICE1.1 Is Associated with Putrescine Synthesis through Regulation of the Arginine Decarboxylase Gene in Freezing Tolerance of Turnip (Brassica rapa Var. Rapa). BMC Plant Biol. 2020, 20, 504. [Google Scholar] [CrossRef] [PubMed]
- Mustafavi, S.H.; Shekari, F.; Abbasi, A. Putrescine Improve Low Temperature Tolerance of Fennel (Foeniculum vulgare Mill.) Seeds. Cercet. Agron. Mold. 2015, 48, 69–76. [Google Scholar] [CrossRef]
- Abbasi, N.; Ali, I.; Hafiz, I.; Alenazi, M.; Shafiq, M. Effects of Putrescine Application on Peach Fruit during Storage. Sustainability 2019, 11, 2013. [Google Scholar] [CrossRef]
- Spormann, S.; Soares, C.; Teixeira, J.; Fidalgo, F. Polyamines as Key Regulatory Players in Plants under Metal Stress—A Way for an Enhanced Tolerance. Ann. Appl. Biol. 2021, 178, 209–226. [Google Scholar] [CrossRef]
- Sharma, S.S.; Dietz, K.-J. The Significance of Amino Acids and Amino Acid-Derived Molecules in Plant Responses and Adaptation to Heavy Metal Stress. J. Exp. Bot. 2006, 57, 711–726. [Google Scholar] [CrossRef]
- Weinstein, L.H.; Kaur-Sawhney, R.; Rajam, M.V.; Wettlaufer, S.H.; Galston, A.W. Cadmium-Induced Accumulation of Putrescine in Oat and Bean Leaves. Plant Physiol. 1986, 82, 641–645. [Google Scholar] [CrossRef]
- Zacchini, M.; de Agazio, M. Spread of Oxidative Damage and Antioxidative Response through Cell Layers of Tobacco Callus after UV-C Treatment. Plant Physiol. Biochem. 2004, 42, 445–450. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Wang, Z.; Zhao, Y.; Zhang, X.; Zhang, S.; Bo, L.; Wang, Y.; Ding, Y.; An, L. Putrescine Protects Hulless Barley from Damage Due to UV-B Stress via H2S- and H2O2-Mediated Signaling Pathways. Plant Cell Rep. 2016, 35, 1155–1168. [Google Scholar] [CrossRef] [PubMed]
- Roy, M.; Wu, R. Arginine Decarboxylase Transgene Expression and Analysis of Environmental Stress Tolerance in Transgenic Rice. Plant Sci. 2001, 160, 869–875. [Google Scholar] [CrossRef]
- Capell, T.; Bassie, L.; Christou, P. Modulation of the Polyamine Biosynthetic Pathway in Transgenic Rice Confers Tolerance to Drought Stress. Proc. Natl. Acad. Sci. USA 2004, 101, 9909–9914. [Google Scholar] [CrossRef] [PubMed]
- Prabhavathi, V.R.; Rajam, M.V. Polyamine Accumulation in Transgenic Eggplant Enhances Tolerance to Multiple Abiotic Stresses and Fungal Resistance. Plant Biotechnol. 2007, 24, 273–282. [Google Scholar] [CrossRef]
- Alet, A.I.; Sanchez, D.H.; Cuevas, J.C.; del Valle, S.; Altabella, T.; Tiburcio, A.F.; Marco, F.; Ferrando, A.; Espasandín, F.D.; González, M.E.; et al. Putrescine Accumulation in Arabidopsis thaliana Transgenic Lines Enhances Tolerance to Dehydration and Freezing Stress. Plant Signal. Behav. 2011, 6, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.-Q.; Zhang, Q.-F.; Liu, J.-H.; Li, G.-H. Overexpression of PtADC Confers Enhanced Dehydration and Drought Tolerance in Transgenic Tobacco and Tomato: Effect on ROS Elimination. Biochem. Biophys. Res. Commun. 2011, 413, 10–16. [Google Scholar] [CrossRef]
- Wang, J.; Sun, P.-P.; Chen, C.-L.; Wang, Y.; Fu, X.-Z.; Liu, J.-H. An Arginine Decarboxylase Gene PtADC from Poncirus trifoliata Confers Abiotic Stress Tolerance and Promotes Primary Root Growth in Arabidopsis. J. Exp. Bot. 2011, 62, 2899–2914. [Google Scholar] [CrossRef]
- Kumria, R.; Rajam, M.V. Ornithine Decarboxylase Transgene in Tobacco Affects Polyamines, In Vitro-Morphogenesis and Response to Salt Stress. J. Plant Physiol. 2002, 159, 983–990. [Google Scholar] [CrossRef]
- Jiménez-Bremont, J.F.; Marina, M.; Guerrero-González, M.d.l.L.; Rossi, F.R.; Sánchez-Rangel, D.; Rodrí guez-Kessler, M.; Ruiz, O.A.; Gárriz, A. Physiological and Molecular Implications of Plant Polyamine Metabolism during Biotic Interactions. Front. Plant Sci. 2014, 5, 95. [Google Scholar] [PubMed]
- Angelini, R.; Cona, A.; Federico, R.; Fincato, P.; Tavladoraki, P.; Tisi, A. Plant Amine Oxidases “on the Move”: An Update. Plant Physiol. Biochem. 2010, 48, 560–564. [Google Scholar] [CrossRef] [PubMed]
- Planas-Portell, J.; Gallart, M.; Tiburcio, A.F.; Altabella, T. Copper-Containing Amine Oxidases Contribute to Terminal Polyamine Oxidation in Peroxisomes and Apoplast of Arabidopsis thaliana. BMC Plant Biol. 2013, 13, 109. [Google Scholar] [CrossRef] [PubMed]
- Tavladoraki, P.; Cona, A.; Angelini, R. Copper-Containing Amine Oxidases and FAD-Dependent Polyamine Oxidases Are Key Players in Plant Tissue Differentiation and Organ Development. Front. Plant Sci. 2016, 7, 824. [Google Scholar] [CrossRef] [PubMed]
- Cona, A.; Rea, G.; Angelini, R.; Federico, R.; Tavladoraki, P. Functions of Amine Oxidases in Plant Development and Defence. Trends Plant Sci. 2006, 11, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Alcázar, R. A New Insight into the Contribution of Putrescine to Defense in Arabidopsis thaliana. Plant Signal. Behav. 2021, 16, 1885187. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Atanasov, K.E.; Arafaty, N.; Murillo, E.; Tiburcio, A.F.; Zeier, J.; Alcázar, R. Putrescine Elicits ROS-dependent Activation of the Salicylic Acid Pathway in Arabidopsis thaliana. Plant Cell Environ. 2020, 43, 2755–2768. [Google Scholar] [CrossRef] [PubMed]
- Marini, F.; Betti, L.; Scaramagli, S.; Biondi, S.; Torrigiani, P. Polyamine Metabolism Is Upregulated in Response to Tobacco Mosaic Virus in Hypersensitive, but Not in Susceptible, Tobacco. New Phytol. 2001, 149, 301–309. [Google Scholar] [CrossRef]
- Rodríguez-Kessler, M.; Ruiz, O.A.; Maiale, S.; Ruiz-Herrera, J.; Jiménez-Bremont, J.F. Polyamine Metabolism in Maize Tumors Induced by Ustilago maydis. Plant Physiol. Biochem. 2008, 46, 805–814. [Google Scholar] [CrossRef]
- Foster, S.A.; Walters, D.R. Polyamine Concentrations and Activities of Ornithine and Arginine Decarboxylase in Wheat Infected with the Stem Rust Fungus. J. Plant Physiol. 1992, 140, 134–136. [Google Scholar] [CrossRef]
- Marina, M.; Maiale, S.J.; Rossi, F.R.; Romero, M.F.; Rivas, E.I.; Gárriz, A.; Ruiz, O.A.; Pieckenstain, F.L. Apoplastic Polyamine Oxidation Plays Different Roles in Local Responses of Tobacco to Infection by the Necrotrophic Fungus Sclerotinia sclerotiorum and the Biotrophic Bacterium Pseudomonas viridiflava. Plant Physiol. 2008, 147, 2164–2178. [Google Scholar] [CrossRef]
- Brauc, S.; de Vooght, E.; Claeys, M.; Geuns, J.M.C.; Höfte, M.; Angenon, G. Overexpression of Arginase in Arabidopsis Thaliana Influences Defence Responses against Botrytis cinerea. Plant Biol. 2012, 14, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Mitsuya, Y.; Takahashi, Y.; Berberich, T.; Miyazaki, A.; Matsumura, H.; Takahashi, H.; Terauchi, R.; Kusano, T. Spermine Signaling Plays a Significant Role in the Defense Response of Arabidopsis thaliana to Cucumber Mosaic Virus. J. Plant Physiol. 2009, 166, 626–643. [Google Scholar] [CrossRef]
- Liu, C.; Atanasov, K.E.; Tiburcio, A.F.; Alcázar, R. The Polyamine Putrescine Contributes to H2O2 and RbohD/F-Dependent Positive Feedback Loop in Arabidopsis PAMP-Triggered Immunity. Front. Plant Sci. 2019, 10, 894. [Google Scholar] [CrossRef] [PubMed]
- Rossi, F.R.; Gárriz, A.; Marina, M.; Pieckenstain, F.L. Modulation of Polyamine Metabolism in Arabidopsis thaliana by Salicylic Acid. Physiol. Plant. 2021, 173, 843–855. [Google Scholar] [CrossRef]
- Kim, N.H.; Kim, B.S.; Hwang, B.K. Pepper Arginine Decarboxylase Is Required for Polyamine and γ-Aminobutyric Acid Signaling in Cell Death and Defense Response. Plant Physiol. 2013, 162, 2067–2083. [Google Scholar] [CrossRef]
- Kim, N.H.; Choi, H.W.; Hwang, B.K. Xanthomonas campestris Pv. Vesicatoria Effector AvrBsT Induces Cell Death in Pepper, but Suppresses Defense Responses in Tomato. Mol. Plant-Microbe Interact. 2010, 23, 1069–1082. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; von Roepenack-Lahaye, E.; Buntru, M.; de Lange, O.; Schandry, N.; Pérez-Quintero, A.L.; Weinberg, Z.; Lowe-Power, T.M.; Szurek, B.; Michael, A.J.; et al. A Plant Pathogen Type III Effector Protein Subverts Translational Regulation to Boost Host Polyamine Levels. Cell Host Microbe 2019, 26, 638–649. [Google Scholar] [CrossRef]
- Chávez-Martínez, A.I.; Ortega-Amaro, M.A.; Torres, M.; Serrano, M.; Jiménez-Bremont, J.F. Arabidopsis Adc-Silenced Line Exhibits Differential Defense Responses to Botrytis cinerea and Pseudomonas syringae Infection. Plant Physiol. Biochem. 2020, 156, 494–503. [Google Scholar] [CrossRef] [PubMed]
- Hewezi, T.; Howe, P.J.; Maier, T.R.; Hussey, R.S.; Mitchum, M.G.; Davis, E.L.; Baum, T.J. Arabidopsis Spermidine Synthase Is Targeted by an Effector Protein of the Cyst Nematode Heterodera schachtii. Plant Physiol. 2010, 152, 968–984. [Google Scholar] [CrossRef] [PubMed]
- Khajuria, A.; Ohri, P. Exogenously Applied Putrescine Improves the Physiological Responses of Tomato Plant during Nematode Pathogenesis. Sci. Hortic. 2018, 230, 35–42. [Google Scholar] [CrossRef]
- Vilas, J.M.; Romero, F.M.; Rossi, F.R.; Marina, M.; Maiale, S.J.; Calzadilla, P.I.; Pieckenstain, F.L.; Ruiz, O.A.; Gárriz, A. Modulation of Plant and Bacterial Polyamine Metabolism during the Compatible Interaction between Tomato and Pseudomonas syringae. J. Plant Physiol. 2018, 231, 281–290. [Google Scholar] [CrossRef]
- Stes, E.; Vandeputte, O.M.; el Jaziri, M.; Holsters, M.; Vereecke, D. A Successful Bacterial Coup d’État: How Rhodococcus fascians Redirects Plant Development. Annu. Rev. Phytopathol. 2011, 49, 69–86. [Google Scholar] [CrossRef]
- Fernández-Crespo, E.; Scalschi, L.; Llorens, E.; García-Agustín, P.; Camañes, G. NH4+ Protects Tomato Plants against Pseudomonas syringae by Activation of Systemic Acquired Acclimation. J. Exp. Bot. 2015, 66, 6777–6790. [Google Scholar] [CrossRef]
- González-Hernández, A.I.; Fernández-Crespo, E.; Scalschi, L.; Hajirezaei, M.-R.; von Wirén, N.; García-Agustín, P.; Camañes, G. Ammonium Mediated Changes in Carbon and Nitrogen Metabolisms Induce Resistance against Pseudomonas syringae in Tomato Plants. J. Plant Physiol. 2019, 239, 28–37. [Google Scholar] [CrossRef]
- Walters, D.R. Polyamines and Plant Disease. Phytochemistry 2003, 64, 97–107. [Google Scholar] [CrossRef]
- Zeiss, D.R.; Piater, L.A.; Dubery, I.A. Hydroxycinnamate Amides: Intriguing Conjugates of Plant Protective Metabolites. Trends Plant Sci. 2021, 26, 184–195. [Google Scholar] [CrossRef]
- Edreva, A.M.; Velikova, V.B.; Tsonev, T.D. Phenylamides in Plants. Russ. J. Plant Physiol. 2007, 54, 287–301. [Google Scholar] [CrossRef]
- Alamgir, K.M.; Hojo, Y.; Christeller, J.T.; Fukumoto, K.; Isshiki, R.; Shinya, T.; Baldwin, I.T.; Galis, I. Systematic Analysis of Rice (Oryza sativa) Metabolic Responses to Herbivory. Plant Cell Environ. 2016, 39, 453–466. [Google Scholar] [CrossRef]
- Kaur, H.; Heinzel, N.; Schöttner, M.; Baldwin, I.T.; GÁlis, I. R2R3-NaMYB8 Regulates the Accumulation of Phenylpropanoid-Polyamine Conjugates, Which Are Essential for Local and Systemic Defense against Insect Herbivores in Nicotiana Attenuata. Plant Physiol. 2010, 152, 1731–1747. [Google Scholar] [CrossRef]
- Tanabe, K.; Hojo, Y.; Shinya, T.; Galis, I. Molecular Evidence for Biochemical Diversification of Phenolamide Biosynthesis in Rice Plants. J. Integr. Plant Biol. 2016, 58, 903–913. [Google Scholar] [CrossRef]
- Muroi, A.; Ishihara, A.; Tanaka, C.; Ishizuka, A.; Takabayashi, J.; Miyoshi, H.; Nishioka, T. Accumulation of Hydroxycinnamic Acid Amides Induced by Pathogen Infection and Identification of Agmatine Coumaroyltransferase in Arabidopsis thaliana. Planta 2009, 230, 517–527. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Shen, L.; Li, F.; Meng, D.; Sheng, J. Methyl Salicylate-Induced Arginine Catabolism Is Associated with Up-Regulation of Polyamine and Nitric Oxide Levels and Improves Chilling Tolerance in Cherry Tomato Fruit. J. Agric. Food Chem. 2011, 59, 9351–9357. [Google Scholar] [CrossRef] [PubMed]
- Jang, E.-K.; Min, K.-H.; Kim, S.-H.; Nam, S.-H.; Zhang, S.; Kim, Y.C.; Cho, B.H.; Yang, K.-Y. Mitogen-Activated Protein Kinase Cascade in the Signaling for Polyamine Biosynthesis in Tobacco. Plant Cell Physiol. 2009, 50, 658–664. [Google Scholar] [CrossRef]
- Németh, M.; Janda, T.; Horváth, E.; Páldi, E.; Szalai, G. Exogenous Salicylic Acid Increases Polyamine Content but May Decrease Drought Tolerance in Maize. Plant Sci. 2002, 162, 569–574. [Google Scholar] [CrossRef]
- Walters, D.; Cowley, T.; Mitchell, A. Methyl Jasmonate Alters Polyamine Metabolism and Induces Systemic Protection against Powdery Mildew Infection in Barley Seedlings. J. Exp. Bot. 2002, 53, 747–756. [Google Scholar] [CrossRef]
- Haggag, W.M.; Abd-El-Kareem, F. Methyl Jasmonate Stimulates Polyamines Biosynthesisand Resistance against Leaf Rust in Wheat Plants. Arch. Phytopathol. Plant Prot. 2009, 42, 16–31. [Google Scholar] [CrossRef]
- Xu, B.; Sheehan, M.J.; Timko, M.P. Differential Induction of Ornithine Decarboxylase (ODC) Gene Family Members in Transgenic Tobacco (Nicotiana tabacum L. Cv. Bright Yellow 2) Cell Suspensions by Methyl-Jasmonate Treatment. Plant Growth Regul. 2004, 44, 101–116. [Google Scholar] [CrossRef]
- Peremarti, A.; Bassie, L.; Yuan, D.; Pelacho, A.; Christou, P.; Capell, T. Transcriptional Regulation of the Rice Arginine Decarboxylase (Adc1) and S-Adenosylmethionine Decarboxylase (Samdc) Genes by Methyl Jasmonate. Plant Physiol. Biochem. 2010, 48, 553–559. [Google Scholar] [CrossRef] [PubMed]
- An, Z.; Jing, W.; Liu, Y.; Zhang, W. Hydrogen Peroxide Generated by Copper Amine Oxidase Is Involved in Abscisic Acid-Induced Stomatal Closure in Vicia faba. J. Exp. Bot. 2008, 59, 815–825. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Gene | Source | Plant | Enhanced Tolerance to | Reference |
|---|---|---|---|---|
| ADC | Avena sativa | Oryza sativa | Salinity | [170] |
| Datura stramonium | Oryza sativa | Drought (PEG8000) | [171] | |
| Avena sativa | Solanum melongena | Salinity, drought, high temperature and heavy metal | [172] | |
| Avena sativa | Arabidopsis thaliana | Dehydration and low temperature | [173] | |
| Poncirus trifoliate | Arabidopsis thaliana | Dehydration and drought | [174] | |
| Poncirus trifoliate | Arabidopsis thaliana | Osmotic stress, dehydration, drought and low temperature | [175] | |
| Avena sativa | Lotus tenuis | Drought | [112] | |
| ADC1 | Arabidopsis thaliana | Arabidopsis thaliana | Low temperatures | [158] |
| ADC2 | Arabidopsis thaliana | Arabidopsis thaliana | Drought | [133] |
| ODC | Mus musculus | Tobacco | Salinity | [176] |
| Gene | Plant | Biotic Stress | Reference |
|---|---|---|---|
| ADC, ODC | Nicotiana tabacum | Tobacco Mosaic virus (TMV) | [184] |
| ADC, ODC | Hordeum vulgare | Blumeria graminis f. sp. Hordei fungus | [202] |
| ODC | Triticum aestivum | Puccinia graminis f. sp. tritici | [186] |
| ADC | Solanum melongena | Fusarium oxysporium | [172] |
| ADC | Capsicum annuum | avirulent Xanthomonas campestris pv versicatoria | [192] |
| ADC | Solanum lycopersicum | Ralstonia solanacearum | [174] |
| ADC1, ADC2 | Arabidopsis thaliana | Heteodera schachtii | [196] |
| ADC1, ADC2 | Arabidopsis thaliana | Avirulent Cucumber Mosaic virus | [189] |
| ADC2 | Arabidopsis thaliana | Pseudomonas syringae | [14] |
| Arginase 2 | Arabidopsis thaliana | Botrytis cinerea | [188] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-Hernández, A.I.; Scalschi, L.; Vicedo, B.; Marcos-Barbero, E.L.; Morcuende, R.; Camañes, G. Putrescine: A Key Metabolite Involved in Plant Development, Tolerance and Resistance Responses to Stress. Int. J. Mol. Sci. 2022, 23, 2971. https://doi.org/10.3390/ijms23062971
González-Hernández AI, Scalschi L, Vicedo B, Marcos-Barbero EL, Morcuende R, Camañes G. Putrescine: A Key Metabolite Involved in Plant Development, Tolerance and Resistance Responses to Stress. International Journal of Molecular Sciences. 2022; 23(6):2971. https://doi.org/10.3390/ijms23062971
Chicago/Turabian StyleGonzález-Hernández, Ana Isabel, Loredana Scalschi, Begonya Vicedo, Emilio Luis Marcos-Barbero, Rosa Morcuende, and Gemma Camañes. 2022. "Putrescine: A Key Metabolite Involved in Plant Development, Tolerance and Resistance Responses to Stress" International Journal of Molecular Sciences 23, no. 6: 2971. https://doi.org/10.3390/ijms23062971
APA StyleGonzález-Hernández, A. I., Scalschi, L., Vicedo, B., Marcos-Barbero, E. L., Morcuende, R., & Camañes, G. (2022). Putrescine: A Key Metabolite Involved in Plant Development, Tolerance and Resistance Responses to Stress. International Journal of Molecular Sciences, 23(6), 2971. https://doi.org/10.3390/ijms23062971