
Subcellular Distribution of Thyroid Hormone Receptor Beta in Ovarian Cancer


 , , ,
, , ,
Abstract
1. Introduction
2. Results
2.1. Study Cohort
2.2. TR Beta/TR Beta-1 in Ovarian Cancer Tissue
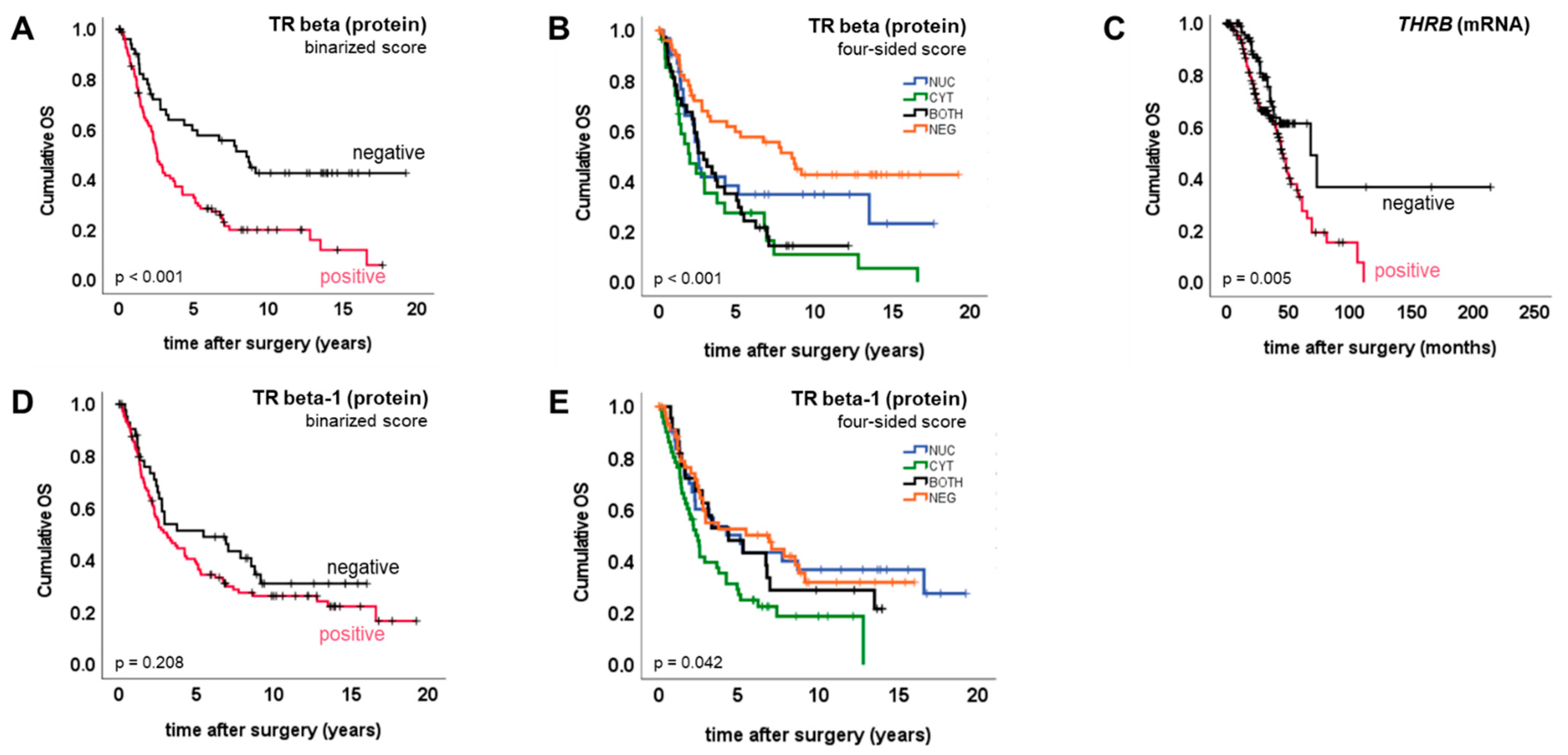
2.3. TR Beta/Beta-1 and Ovarian Cancer Prognosis
3. Discussion
4. Materials and Methods
4.1. Tissue Samples
4.2. Immunohistochemistry
4.3. Statistical Analysis Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Perri, A.; Catalano, S.; Bonofiglio, D.; Vizza, D.; Rovito, D.; Qi, H.; Aquila, S.; Panza, S.; Rizza, P.; Lanzino, M.; et al. T3 enhances thyroid cancer cell proliferation through TRbeta1/Oct-1-mediated cyclin D1 activation. Mol. Cell. Endocrinol. 2014, 382, 205–217. [Google Scholar] [CrossRef]
- Munoz, A.; Bernal, J. Biological activities of thyroid hormone receptors. Eur. J. Endocrinol. Eur. Fed. Endocr. Soc. 1997, 137, 433–445. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lazar, M.A.; Berrodin, T.J.; Harding, H.P. Differential DNA binding by monomeric, homodimeric, and potentially heteromeric forms of the thyroid hormone receptor. Mol. Cell. Biol. 1991, 11, 5005–5015. [Google Scholar] [CrossRef]
- Weinberger, C.; Thompson, C.C.; Ong, E.S.; Lebo, R.; Gruol, D.J.; Evans, R.M. The c-erb-A gene encodes a thyroid hormone receptor. Nature 1986, 324, 641–646. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.G.; Cheng, S.-Y. Thyroid hormone receptors and cancer. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2013, 1830, 3928–3936. [Google Scholar] [CrossRef]
- Mishkin, S.Y.; Pollack, R.; Yalovsky, M.A.; Morris, H.P. Inhibition of local and metastatic hepatoma growth and prolongation of survival after induction of hypothyroidism. Cancer Res. 1981, 41, 3040–3045. [Google Scholar]
- Mousa, S.A.; Yalcin, M.; Bharali, D.J.; Meng, R.; Tang, H.-Y.; Lin, H.-Y.; Davis, F.B.; Davis, P.J. Tetraiodothyroacetic acid and its nanoformulation inhibit thyroid hormone stimulation of non-small cell lung cancer cells in vitro and its growth in xenografts. Lung Cancer 2011, 76, 39–45. [Google Scholar] [CrossRef]
- Ness, R.B.; Grisso, J.A.; Cottreau, C.; Klapper, J.; Vergona, R.; Wheeler, J.E.; Morgan, M.; Schlesselman, J.J. Factors Related to Inflammation of the Ovarian Epithelium and Risk of Ovarian Cancer. Epidemiology 2000, 11, 111–117. [Google Scholar] [CrossRef]
- Aghajanova, L.; Lindeberg, M.; Carlsson, I.B.; Stavreus-Evers, A.; Zhang, P.; Scott, J.E.; Hovatta, O.; Skjöldebrand-Sparre, L. Receptors for thyroid-stimulating hormone and thyroid hormones in human ovarian tissue. Reprod. Biomed. Online 2009, 18, 337–347. [Google Scholar] [CrossRef]
- Rae, M.T.; Gubbay, O.; Kostogiannou, A.; Price, D.; Critchley, H.O.D.; Hillier, S.G. Thyroid Hormone Signaling in Human Ovarian Surface Epithelial Cells. J. Clin. Endocrinol. Metab. 2007, 92, 322–327. [Google Scholar] [CrossRef]
- Ditsch, N.; Heublein, S.; Jeschke, U.; Sattler, C.; Kuhn, C.; Hester, A.; Czogalla, B.; Trillsch, F.; Mahner, S.; Engel, J.; et al. Cytoplasmic versus nuclear THR alpha expression determines survival of ovarian cancer patients. J. Cancer Res. Clin. Oncol. 2020, 146, 1923–1932. [Google Scholar] [CrossRef]
- Cao, X.; Kambe, F.; Moeller, L.; Refetoff, S.; Seo, H. Thyroid hormone induces rapid activation of Akt/protein kinase B-mammalian target of rapamycin-p70S6K cascade through phosphatidylinositol 3-kinase in human fibroblasts. Mol. Endocrinol. 2005, 19, 102–112. [Google Scholar] [CrossRef] [PubMed]
- Tothill, R.W.; Tinker, A.V.; George, J.; Brown, R.; Fox, S.B.; Lade, S.; Johnson, D.S.; Trivett, M.K.; Etemadmoghadam, D.; Locandro, B.; et al. Novel molecular subtypes of serous and endometrioid ovarian cancer linked to clinical outcome. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2008, 14, 5198–5208. [Google Scholar] [CrossRef]
- Roggero, V.R.; Zhang, J.; Parente, L.E.; Doshi, Y.; Dziedzic, R.C.; McGregor, E.L.; Varjabedian, A.D.; Schad, S.E.; Bondzi, C.; Allison, L.A. Nuclear import of the thyroid hormone receptor α1 is mediated by importin 7, importin β1, and adaptor importin α1. Mol. Cell. Endocrinol. 2016, 419, 185–197. [Google Scholar] [CrossRef]
- Subramanian, K.S.; Dziedzic, R.C.; Nelson, H.N.; Stern, M.E.; Roggero, V.R.; Bondzi, C.; Allison, L.A. Multiple exportins influence thyroid hormone receptor localization. Mol. Cell. Endocrinol. 2015, 411, 86–96. [Google Scholar] [CrossRef]
- Heublein, S.; Mayr, R.; Meindl, A.; Angele, M.; Gallwas, J.; Jeschke, U.; Ditsch, N. Thyroid Hormone Receptors Predict Prognosis in BRCA1 Associated Breast Cancer in Opposing Ways. PLoS ONE 2015, 10, e0127072. [Google Scholar] [CrossRef]
- Bonamy, G.M.; Allison, L.A. Oncogenic conversion of the thyroid hormone receptor by altered nuclear transport. Nucl. Recept. Signal. 2006, 4, e008. [Google Scholar] [CrossRef]
- Martin, N.P.; Fernandez de Velasco, E.M.; Mizuno, F.; Scappini, E.L.; Gloss, B.; Erxleben, C.; Williams, J.G.; Stapleton, H.M.; Gentile, S.; Armstrong, D.L. A Rapid Cytoplasmic Mechanism for PI3 Kinase Regulation by the Nuclear Thyroid Hormone Receptor, TRβ, and Genetic Evidence for Its Role in the Maturation of Mouse Hippocampal Synapses In Vivo. Endocrinology 2014, 155, 3713–3724. [Google Scholar] [CrossRef]
- Jerzak, K.J.; Cockburn, J.G.; Dhesy-Thind, S.K.; Pond, G.R.; Pritchard, K.I.; Nofech-Mozes, S.; Sun, P.; Narod, S.A.; Bane, A. Thyroid hormone receptor beta-1 expression in early breast cancer: A validation study. Breast Cancer Res. Treat. 2018, 171, 709–717. [Google Scholar] [CrossRef]
- Shao, W.; Kuhn, C.; Mayr, D.; Ditsch, N.; Kailuweit, M.; Wolf, V.; Harbeck, N.; Mahner, S.; Jeschke, U.; Cavaillès, V.; et al. Cytoplasmic and Nuclear Forms of Thyroid Hormone Receptor β1 Are Inversely Associated with Survival in Primary Breast Cancer. Int. J. Mol. Sci. 2020, 21, 330. [Google Scholar] [CrossRef]
- Piqué, D.G.; Greally, J.M.; Mar, J.C. Identification of a novel subgroup of endometrial cancer patients with loss of thyroid hormone receptor beta expression and improved survival. BMC Cancer 2020, 20, 857. [Google Scholar] [CrossRef] [PubMed]
- Park, J.W.; Zhao, L.; Willingham, M.C.; Cheng, S.-Y. Loss of tyrosine phosphorylation at Y406 abrogates the tumor suppressor functions of the thyroid hormone receptor β. Mol. Carcinog. 2016, 56, 489–498. [Google Scholar] [CrossRef]
- Krashin, E.; Piekiełko-Witkowska, A.; Ellis, M.; Ashur-Fabian, O. Thyroid Hormones and Cancer: A Comprehensive Review of Preclinical and Clinical Studies. Front. Endocrinol. 2019, 10, 59. [Google Scholar] [CrossRef]
- Kenessey, A.; Ojamaa, K. Thyroid hormone stimulates protein synthesis in the cardiomyocyte by activating the Akt-mTOR and p70S6K pathways. J. Biol. Chem. 2006, 281, 20666–20672. [Google Scholar] [CrossRef]
- Davidson, C.D.; Gillis, N.E.; Carr, F.E. Thyroid Hormone Receptor Beta as Tumor Suppressor: Untapped Potential in Treatment and Diagnostics in Solid Tumors. Cancers 2021, 13, 4254. [Google Scholar] [CrossRef]
- Ditsch, N.; Toth, B.; Himsl, I.; Lenhard, M.; Ochsenkühn, R.; Friese, K.; Mayr, D.; Jeschke, U. Thyroid hormone receptor (TR)alpha and TRbeta expression in breast cancer. Histol. Histopathol. 2013, 28, 227–237. [Google Scholar]
- Ditsch, N.; Mayr, R.; Lenhard, M.; Strauss, C.; Vodermaier, A.; Gallwas, J.; Stoeckl, D.; Graeser, M.; Weissenbacher, T.; Friese, K.; et al. Correlation of thyroid hormone, retinoid X, peroxisome proliferator-activated, vitamin D and oestrogen/progesterone receptors in breast carcinoma. Oncol. Lett. 2012, 4, 665–671. [Google Scholar] [CrossRef]
- Ditsch, N.; Toth, B.; Mayr, D.; Lenhard, M.; Gallwas, J.; Weissenbacher, T.; Dannecker, C.; Friese, K.; Jeschke, U. The Association between Vitamin D Receptor Expression and Prolonged Overall Survival in Breast Cancer. J. Histochem. Cytochem. 2011, 60, 121–129. [Google Scholar] [CrossRef]
- Lenhard, M.; Tsvilina, A.; Schumacher, L.; Kupka, M.; Ditsch, N.; Mayr, D.; Friese, K.; Jeschke, U. Human chorionic gonadotropin and its relation to grade, stage and patient survival in ovarian cancer. BMC Cancer 2012, 12, 2. [Google Scholar] [CrossRef]
- Lenhard, M.; Lennerová, T.; Ditsch, N.; Kahlert, S.; Friese, K.; Mayr, D.; Jeschke, U. Opposed roles of follicle-stimulating hormone and luteinizing hormone receptors in ovarian cancer survival. Histopathology 2011, 58, 990–994. [Google Scholar] [CrossRef]
- Gyorffy, B.; Lánczky, A.; Szallasi, Z. Implementing an online tool for genome-wide validation of survival-associated biomarkers in ovarian-cancer using microarray data from 1287 patients. Endocr. Relat. Cancer 2012, 19, 197–208. [Google Scholar] [CrossRef] [PubMed]
- Sattler, C. Expressionsanalyse nukleärer Hormonrezeptoren im Ovarialkarzinom. Ph.D. Thesis, LMU München, Munich, Germany, 2015. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| n (Total) | n | % | ||
|---|---|---|---|---|
| Histology | 153 | |||
| HGSOC | 82 | 54 | ||
| LGSOC | 26 | 17 | ||
| clear cell | 12 | 8 | ||
| endometrioid | 21 | 14 | ||
| mucinous | 12 | 8 | ||
| FIGO stage | 152 | |||
| I, II | 43 | 28 | ||
| III, IV | 109 | 72 | ||
| pN | 93 | |||
| N0 | 41 | 45 | ||
| N+ | 51 | 55 | ||
| Patient age | 152 | |||
| ≤55 years | 62 | 41 | ||
| >55 years | 90 | 59 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heublein, S.; Jeschke, U.; Sattler, C.; Kuhn, C.; Hester, A.; Czogalla, B.; Trillsch, F.; Mahner, S.; Mayr, D.; Schmoeckel, E.; et al. Subcellular Distribution of Thyroid Hormone Receptor Beta in Ovarian Cancer. Int. J. Mol. Sci. 2022, 23, 2698. https://doi.org/10.3390/ijms23052698
Heublein S, Jeschke U, Sattler C, Kuhn C, Hester A, Czogalla B, Trillsch F, Mahner S, Mayr D, Schmoeckel E, et al. Subcellular Distribution of Thyroid Hormone Receptor Beta in Ovarian Cancer. International Journal of Molecular Sciences. 2022; 23(5):2698. https://doi.org/10.3390/ijms23052698
Chicago/Turabian StyleHeublein, Sabine, Udo Jeschke, Cornelia Sattler, Christina Kuhn, Anna Hester, Bastian Czogalla, Fabian Trillsch, Sven Mahner, Doris Mayr, Elisa Schmoeckel, and et al. 2022. "Subcellular Distribution of Thyroid Hormone Receptor Beta in Ovarian Cancer" International Journal of Molecular Sciences 23, no. 5: 2698. https://doi.org/10.3390/ijms23052698
APA StyleHeublein, S., Jeschke, U., Sattler, C., Kuhn, C., Hester, A., Czogalla, B., Trillsch, F., Mahner, S., Mayr, D., Schmoeckel, E., & Ditsch, N. (2022). Subcellular Distribution of Thyroid Hormone Receptor Beta in Ovarian Cancer. International Journal of Molecular Sciences, 23(5), 2698. https://doi.org/10.3390/ijms23052698