
Cell Therapy with Human Reprogrammed CD8+ T-Cells Has Antimetastatic Effects on Lewis Lung Carcinoma in C57BL/6 Mice

 , , , ,
, , , ,  ,
,
Abstract
1. Introduction
2. Results
2.1. Morphological and Histological Analysis of the Lungs of Mice with Lewis Lung Carcinoma after Cell Therapy with Human Reprogrammed CD8+ T-Cells
2.1.1. Histological Examination
2.1.2. Tumor Growth and Metastases
2.2. Effect of Human Reprogrammed CD8+ T-Cells on Lung Cancer Cells and CSCs Isolated from Mice with Lewis Lung Carcinoma
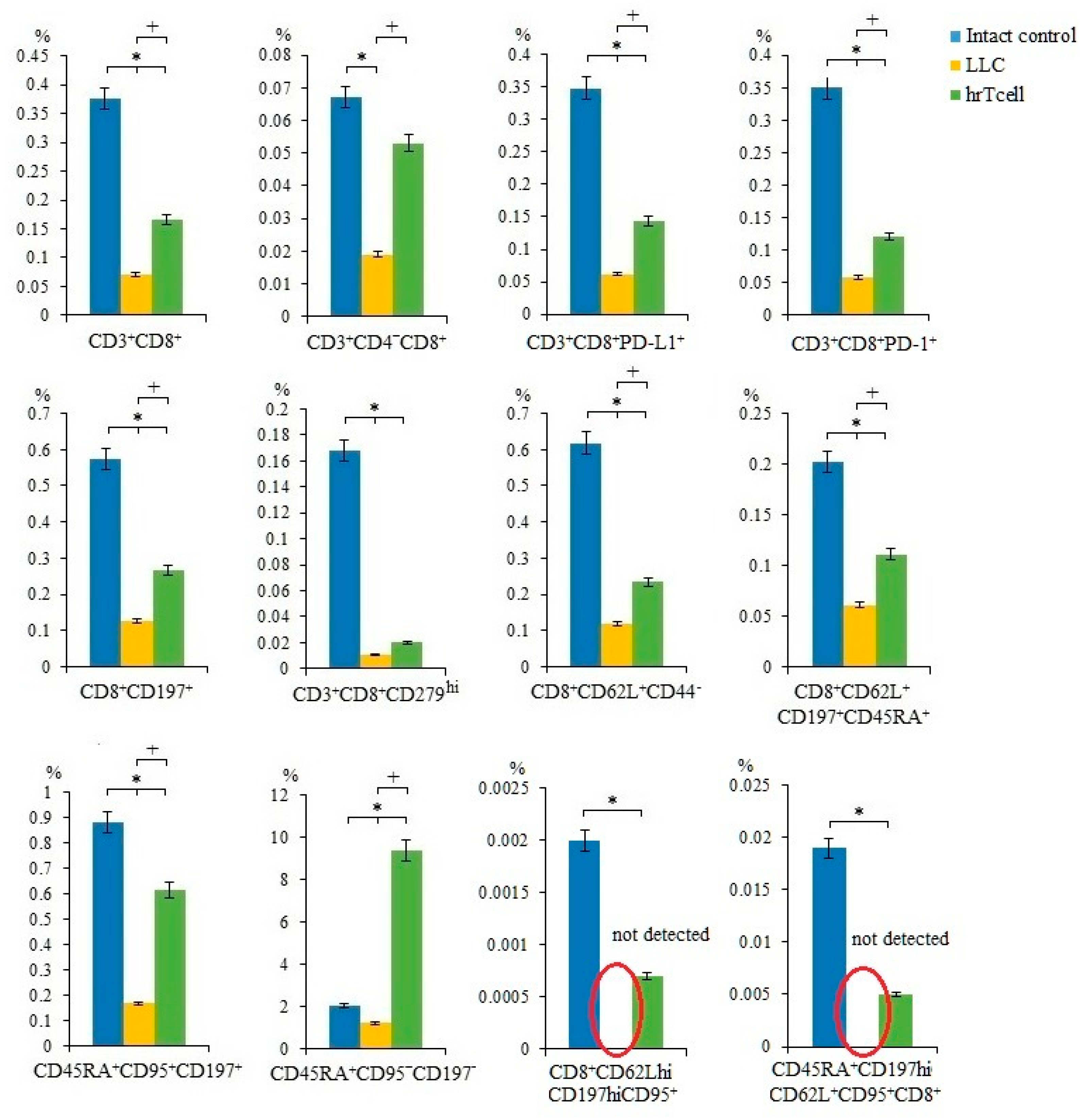
2.3. Effect of Human Reprogrammed CD8+ T-Cells on Blood T-Cell Populations in Mice with Lewis Lung Carcinoma
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Lewis Lung Carcinoma Cell Line
4.3. Lewis Lung Carcinoma Cell Culture
4.4. Modeling Immunosuppression in Mice
4.5. Metastatic Model of Lewis Lung Carcinoma
4.6. Characteristics of a Healthy Volunteer
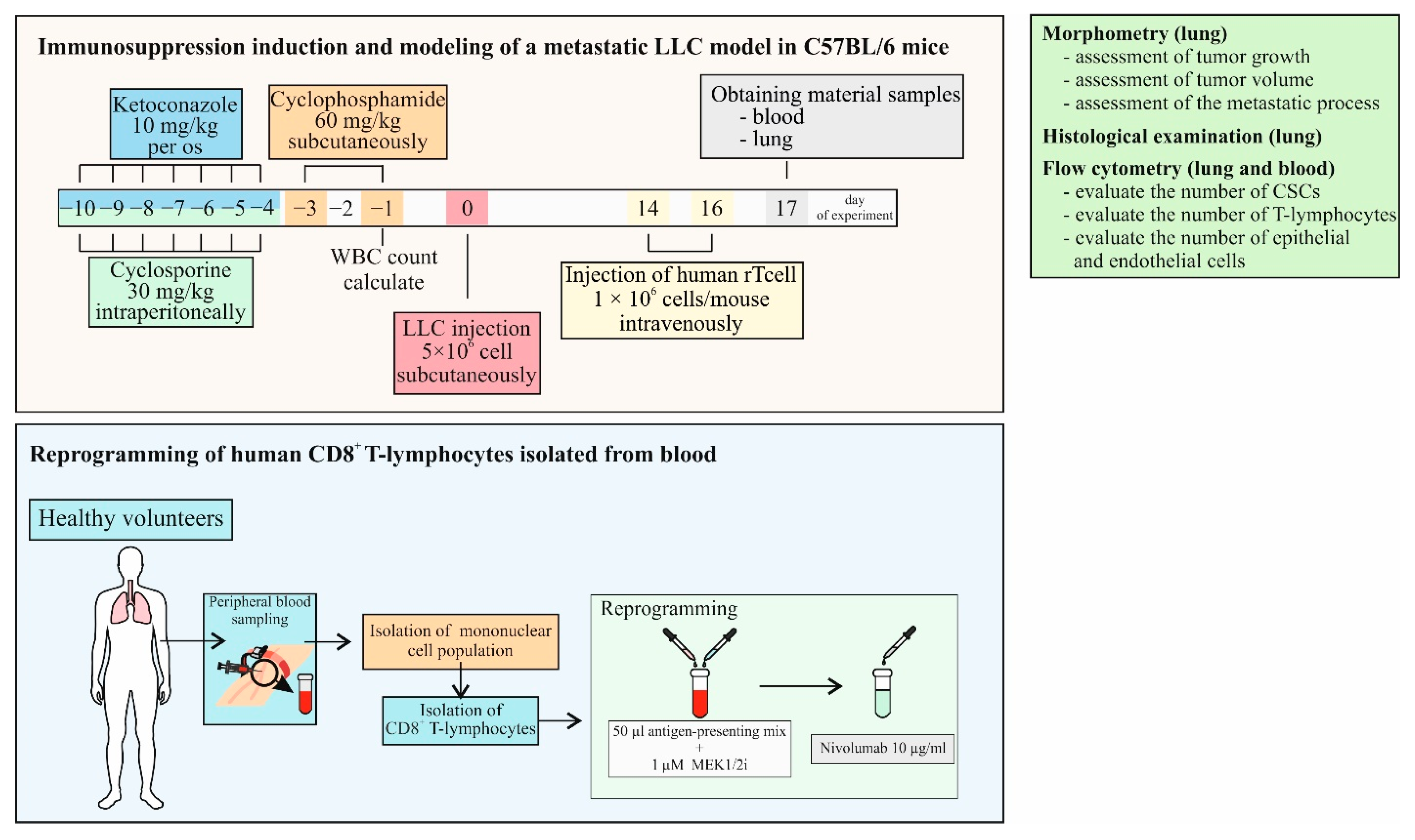
4.7. Study Design
4.8. Isolation of Human Blood Mononuclear Cells
4.9. Cryopreservation of Mononuclear Cells Obtained from Human Blood
4.10. Magnetic Separation of Human CD8+ T-Cells
4.11. Reprogramming of Human CD8+ T-Cells
4.12. Detection of the CCR7 Expression and Cytotoxicity in CD8+ T-Cells In Vitro
4.13. Injection of Reprogrammed CD8+ T-Cells
4.14. Isolation of Mouse Mononuclear Cells
4.15. Detection of the Cancer Cells and CSCs Receptor Expression on Mouse Blood and Lung Mononuclear Cells
4.16. Histological Examination of the Lungs
4.17. Assessment of Tumor Growth
4.18. Assessment of Tumor Volume
4.19. Evaluation of the Severity of the Metastatic Process [49]
- -
- The frequency of tumor metastasis is the percentage of animals with metastases in relation to the total number of animals in the group;
- -
- The degree of damage to the lungs by LLC metastases, where
- -
- Average number of metastases per animal in each group;
- -
- Is the average weight of the lungs affected by LLC metastases;
- -
- Metastasis inhibition index (MII)—MII was calculated by the formula:
4.20. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Friedlaender, A.; Addeo, A.; Russo, A.; Gregorc, V.; Cortinovis, D.; Rolfo, C.D. Targeted Therapies in Early Stage NSCLC: Hype or Hope? Int. J. Mol. Sci. 2020, 21, 6329. [Google Scholar] [CrossRef] [PubMed]
- Sarode, P.; Mansouri, S.; Karger, A.; Schaefer, M.B.; Grimminger, F.; Seeger, W.; Savai, R. Epithelial cell plasticity defines heterogeneity in lung cancer. Cell Signal. 2020, 65, 109463. [Google Scholar] [CrossRef] [PubMed]
- Min, H.Y.; Lee, H.Y. Mechanisms of resistance to chemotherapy in non-small cell lung cancer. Arch. Pharm. Res. 2021, 44, 146–164. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.; Xin, W.; Tong, Y.; Sun, J.; Xu, G.; Ye, Z.; Rao, Y. Cost effectiveness of immune checkpoint inhibitors for treatment of non-small cell lung cancer: A systematic review. PLoS ONE 2020, 15, e0238536. [Google Scholar] [CrossRef]
- Luo, W.; Wang, Z.; Zhang, T.; Yang, L.; Xian, J.; Li, Y.; Li, W. Immunotherapy in non-small cell lung cancer: Rationale, recent advances and future perspectives. Precis. Clin. Med. 2021, 4, 258–270. [Google Scholar] [CrossRef]
- D’Incecco, A.; Andreozzi, M.; Ludovini, V.; Rossi, E.; Capodanno, A.; Landi, L.; Tibaldi, C.; Minuti, G.; Salvini, J.; Coppi, E.; et al. PD-1 and PD-L1 expression in molecularly selected non-small-cell lung cancer patients. Br. J. Cancer. 2015, 112, 95–102. [Google Scholar] [CrossRef]
- Qu, J.; Mei, Q.; Chen, L.; Zhou, J. Chimeric antigen receptor (CAR)-T-cell therapy in non-small-cell lung cancer (NSCLC): Current status and future perspectives. Cancer Immunol. Immunother. 2021, 70, 619–631. [Google Scholar] [CrossRef]
- Kandra, P.; Nandigama, R.; Eul, B.; Huber, M.; Kobold, S.; Seeger, W.; Grimminger, F.; Savai, R. Utility and Drawbacks of Chimeric Antigen Receptor T Cell (CAR-T) Therapy in Lung Cancer. Front. Immunol. 2022, 13, 903562. [Google Scholar] [CrossRef]
- Suresh, R.; Ali, S.; Ahmad, A.; Philip, P.A.; Sarkar, F.H. The Role of Cancer Stem Cells in Recurrent and Drug-Resistant Lung Cancer. Adv. Exp. Med. Biol. 2016, 890, 57–74. [Google Scholar] [CrossRef]
- Herreros-Pomares, A.; de-Maya-Girones, J.D.; Calabuig-Fariñas, S.; Lucas, R.; Martínez, A.; Pardo-Sánchez, J.M.; Alonso, S.; Blasco, A.; Guijarro, R.; Martorell, M.; et al. Lung tumorspheres reveal cancer stem cell-like properties and a score with prognostic impact in resected non-small-cell lung cancer. Cell Death. Dis. 2019, 10, 660. [Google Scholar] [CrossRef]
- Maiuthed, A.; Chantarawong, W.; Chanvorachote, P. Lung Cancer Stem Cells and Cancer Stem Cell-targeting Natural Compounds. Anticancer. Res. 2018, 38, 3797–3809. [Google Scholar] [CrossRef] [PubMed]
- Ying, J.; Shi, C.; Li, C.S.; Hu, L.P.; Zhang, W.D. Expression and significance of SOX2 in non-small cell lung carcinoma. Oncol. Lett. 2016, 12, 3195–3198. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Huang, Y.; Huang, Y.; Chen, J.; Wang, S.; Zhou, J. The prognostic value of SOX2 expression in non-small cell lung cancer: A meta-analysis. PLoS ONE 2013, 8, e71140. [Google Scholar] [CrossRef] [PubMed]
- Mathew, M.; Enzler, T.; Shu, C.A.; Rizvi, N.A. Combining chemotherapy with PD-1 blockade in NSCLC. Pharmacol. Ther. 2018, 186, 130–137. [Google Scholar] [CrossRef]
- Skurikhin, E.G.; Pershina, O.; Ermakova, N.; Pakhomova, A.; Widera, D.; Zhukova, M.; Pan, E.; Sandrikina, L.; Kogai, L.; Kushlinskii, N.; et al. Reprogrammed CD8+ T-Lymphocytes Isolated from Bone Marrow Have Anticancer Potential in Lung Cancer. Biomedicines. 2022, 10, 1450. [Google Scholar] [CrossRef]
- Polson, A.G.; Fuji, R.N. The successes and limitations of preclinical studies in predicting the pharmacodynamics and safety of cell-surface-targeted biological agents in patients. Br. J. Pharmacol. 2012, 166, 1600–1602. [Google Scholar] [CrossRef]
- Robinson, N.B.; Krieger, K.; Khan, F.M.; Huffman, W.; Chang, M.; Naik, A.; Yongle, R.; Hameed, I.; Krieger, K.; Girardi, L.N.; et al. The current state of animal models in research: A review. Int. J. Surg. 2019, 72, 9–13. [Google Scholar] [CrossRef]
- Liu, Q.; Zhao, C.; Jiang, P.; Liu, D. Circulating tumor cells counts are associated with CD8+ T-cell levels in programmed death-ligand 1-negative non-small cell lung cancer patients after radiotherapy: A retrospective study. Medicine 2021, 100, e26674. [Google Scholar] [CrossRef]
- American Cancer Society. Available online: https://www.cancer.org/cancer/lung-cancer/treating-non-small-cell.html (accessed on 1 November 2022).
- Pinheiro, F.D.; Teixeira, A.F.; de Brito, B.B.; da Silva, F.A.F.; Santos, M.L.C.; de Melo, F.F. Immunotherapy—New perspective in lung cancer. World J. Clin. Oncol. 2020, 11, 250–259. [Google Scholar] [CrossRef]
- Steven, A.; Fisher, S.A.; Robinson, B.W. Immunotherapy for lung cancer. Respirology 2016, 21, 821–833. [Google Scholar] [CrossRef]
- Kinoshita, T.; Terai, H.; Yaguchi, T. Clinical Efficacy and Future Prospects of Immunotherapy in Lung Cancer. Life 2021, 11, 1029. [Google Scholar] [CrossRef] [PubMed]
- Raskov, H.; Orhan, A.; Christensen, J.P.; Gögenur, I. Cytotoxic CD8+ T-cells in cancer and cancer immunotherapy. Br. J. Cancer. 2021, 124, 359–367. [Google Scholar] [CrossRef]
- Reiser, J.; Banerjee, A. Effector, Memory, and Dysfunctional CD8(+) T Cell Fates in the Antitumor Immune Response. J. Immunol. Res. 2016, 2016, 8941260. [Google Scholar] [CrossRef] [PubMed]
- Janker, F.; Weder, W.; Jang, J.H.; Jungraithmayr, W. Preclinical, non-genetic models of lung adenocarcinoma: A comparative survey. Oncotarget 2018, 9, 30527–30538. [Google Scholar] [CrossRef] [PubMed]
- Codony-Servat, J.; Verlicchi, A.; Rosell, R. Cancer stem cells in small cell lung cancer. Transl. Lung Cancer Res. 2016, 5, 16–25. [Google Scholar] [CrossRef]
- Skurikhin, E.; Pershina, O.; Zhukova, M.; Widera, D.; Ermakova, N.; Pan, E.; Pakhomova, A.; Morozov, S.; Kubatiev, A.; Dygai, A. Potential of Stem Cells and CART as a Potential Polytherapy for Small Cell Lung Cancer. Front. Cell Dev. Biol. 2021, 9, 778020. [Google Scholar] [CrossRef]
- Von Itzstein, M.S.; Drapkin, B.J.; Minna, J.D. How lung cancer cells change identity. Elife 2021, 10, e71610. [Google Scholar] [CrossRef]
- Li, L.; Lu, G.; Liu, Y.; Gong, L.; Zheng, X.; Zheng, H.; Gu, W.; Yang, L. Low Infiltration of CD8+ PD-L1+ T Cells and M2 Macrophages Predicts Improved Clinical Outcomes After Immune Checkpoint Inhibitor Therapy in Non-Small Cell Lung Carcinoma. Front. Oncol. 2021, 11, 658690. [Google Scholar] [CrossRef]
- Sathaliyawala, T.; Kubota, M.; Yudanin, N.; Turner, D.; Camp, P.; Thome, J.J.; Bickham, K.L.; Lerner, H.; Goldstein, M.; Sykes, M.; et al. Distribution and compartmentalization of human circulating and tissue-resident memory T-cell subsets. Immunity 2013, 38, 187–197. [Google Scholar] [CrossRef]
- Verma, V.; Jafarzadeh, N.; Boi, S.; Kundu, S.; Jiang, Z.; Fan, Y.; Lopez, J.; Nandre, R.; Zeng, P.; Alolaqi, F.; et al. MEK inhibition reprograms CD8+ T lymphocytes into memory stem cells with potent antitumor effects. Nat. Immunol. 2021, 22, 53–66. [Google Scholar] [CrossRef]
- Gattinoni, L.; Speiser, D.E.; Lichterfeld, M.; Bonini, C. T memory stem cells in health and disease. Nat. Med. 2017, 23, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Berard, M.; Tough, D.F. Qualitative differences between naïve and memory T-cells. Immunology 2002, 106, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Gattinoni, L.; Klebanoff, C.A.; Restifo, N.P. Paths to stemness: Building the ultimate antitumour T-cell. Nat. Rev. Cancer. 2012, 12, 671–684. [Google Scholar] [CrossRef] [PubMed]
- Gascón, M.; Isla, D.; Cruellas, M.; Gálvez, E.M.; Lastra, R.; Ocáriz, M.; Paño, J.R.; Ramírez, A.; Sesma, A.; Torres-Ramón, I.; et al. Intratumoral Versus Circulating Lymphoid Cells as Predictive Biomarkers in Lung Cancer Patients Treated with Immune Checkpoint Inhibitors: Is the Easiest Path the Best One? Cells 2020, 9, 1525. [Google Scholar] [CrossRef] [PubMed]
- Ling, M.; Yang, X. Correlation between cancer stem cells (CSCs) and tumor-infiltrating lymphocytes (TILs): Do TILs interact with CSCs in non-small cell lung cancer? Ann. Transl. Med. 2020, 8, 914. [Google Scholar] [CrossRef] [PubMed]
- Badoual, C.; Hans, S.; Merillon, N.; Van Ryswick, C.; Ravel, P.; Benhamouda, N.; Levionnois, E.; Nizard, M.; Si-Mohamed, A.; Besnier, N.; et al. PD-1-expressing tumor-infiltrating T-cells are a favorable prognostic biomarker in HPV-associated head and neck cancer. Cancer Res. 2013, 73, 128–138. [Google Scholar] [CrossRef]
- Chawla, A.S.; Khalsa, J.K.; Dhar, A.; Gupta, S.; Umar, D.; Arimbasseri, G.A.; Bal, V.; George, A.; Rath, S. A role for cell-autocrine interleukin-2 in regulatory T-cell homeostasis. Immunology 2020, 160, 295–309. [Google Scholar] [CrossRef]
- Crofts, K.F.; Holbrook, B.C.; Soto-Pantoja, D.R.; Ornelles, D.A.; Alexander-Miller, M.A. TCR Dependent Metabolic Programming Regulates Autocrine IL-4 Production Resulting in Self-Tuning of the CD8+ T Cell Activation Set point. Front. Immunol. 2020, 11, 540. [Google Scholar] [CrossRef]
- Jivrajani, M.; Shaikh, M.V.; Shrivastava, N.; Nivsarkar, M. An improved and versatile immunosuppression protocol for the development of tumor xenograft in mice. Anticancer. Res. 2014, 34, 7177–7183. [Google Scholar]
- Stankevicius, V.; Kuodyte, K.; Schveigert, D.; Bulotiene, D.; Paulauskas, T.; Daniunaite, K.; Suziedelis, K. Gene and miRNA expression profiles of mouse Lewis lung carcinoma LLC1 cells following single or fractionated dose irradiation. Oncol. Lett. 2017, 13, 4190–4200. [Google Scholar] [CrossRef]
- Meehan, B.; Garnier, D.; Dombrovsky, A.; Lau, K.; D’Asti, E.; Magnus, N.; Rak, J. Ageing-related responses to antiangiogenic effects of sunitinib in atherosclerosis-prone mice. Mech. Ageing Dev. 2014, 140, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Skurikhin, E.G.; Ermakova, N.; Zhukova, M.; Pershina, O.; Pan, E.; Pakhomova, A.; Kogai, L.; Goldberg, V.; Simolina, E.; Skurikhina, V.; et al. Analysis of Circulating Tumor and Cancer Stem Cells Provides New Opportunities in Diagnosis and Treatment of Small Cell Lung Cancer. Int. J. Mol. Sci. 2022, 23, 10853. [Google Scholar] [CrossRef] [PubMed]
- Skurikhin, E.G.; Ermakova, N.N.; Pershina, O.V.; Krupin, V.A.; Pakhomova, A.V.; Dygai, A.M. Response of Hematopoietic Stem and Progenitor Cells to Reserpine in C57Bl/6 Mice. Bull. Exp. Biol. Med. 2016, 160, 439–443. [Google Scholar] [CrossRef]
- Skurikhin, E.; Pershina, O.; Zhukova, M.; Widera, D.; Pan, E.; Pakhomova, A.; Krupin, V.; Ermakova, N.; Skurikhina, V.; Sandrikina, L.; et al. Spiperone Stimulates Regeneration in Pulmonary Endothelium Damaged by Cigarette Smoke and Lipopolysaccharide. Int. J. Chron. Obstruct. Pulmon. Dis. 2021, 16, 3575–3591. [Google Scholar] [CrossRef] [PubMed]
- Cardiff, R.D.; Miller, C.H.; Munn, R.J. Manual hematoxylin and eosin staining of mouse tissue sections. Cold Spring Harb. Protoc. 2014, 2014, 655–658. [Google Scholar] [CrossRef]
- Hather, G.; Liu, R.; Bandi, S.; Mettetal, J.; Manfredi, M.; Shyu, W.C.; Donelan, J.; Chakravarty, A. Growth rate analysis and efficient experimental design for tumor xenograft studies. Cancer Inform. 2014, 13, 65–72. [Google Scholar] [CrossRef]
- Tomayko, M.M.; Reynolds, C.P. Determination of subcutaneous tumor size in athymic (nude) mice. Cancer Chemother. Pharmacol. 1989, 24, 148–154. [Google Scholar] [CrossRef]
- Mikuleak, N.I.; Minnigaleeva, S.D.; Magdeyev, R.R.; Kinzirsky, A.S.; Mikulyak, A.L.; Solomanina, O.O. Valuation of the impact of separate and combined use of carubicinum with probucol, mexidol and Ė-tocopherolon the growth of primary tumor and metastasis oflew is lung carcinoma. Fundam. Research. 2014, 7, 753–758. [Google Scholar]
- Buggert, M.; Nguyen, S.; Salgado-Montes de Oca, G.; Bengsch, B.; Darko, S.; Ransier, A.; Roberts, E.R.; Del Alcazar, D.; Brody, I.B.; Vella, L.A.; et al. Identification and characterization of HIV-specific resident memory CD8+ T-cells in human lymphoid tissue. Sci. Immunol. 2018, 3, eaar4526. [Google Scholar] [CrossRef]
- Czystowska, M.; Gooding, W.; Szczepanski, M.J.; Lopez-Abaitero, A.; Ferris, R.L.; Johnson, J.T.; Whiteside, T.L. The immune signature of CD8(+)CCR7(+) T-cells in the peripheral circulation associates with disease recurrence in patients with HNSCC. Clin. Cancer Res. 2013, 19, 889–899. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Tumor Weight, mg | Tumor Weight Coefficient, mg/g |
|---|---|---|
| Intact | 0 | 0 |
| Mice with LLC | 4.84 ± 0.57 | 175.62 ± 18.95 |
| Mice with LLC + hrT-cells | 5.09 ± 0.42 | 181.08 ± 13.54 |
| Group | X, mm | Y, mm | Z, mm | V, mm3 |
|---|---|---|---|---|
| Mice with LLC | 27.33 ± 1.44 | 17.33 ± 1.33 | 12.78 ± 1.0 8 | 2899.60 ± 513.88 |
| Mice with LLC + hrT-cells | 27.44 ± 1.55 | 16.00 ± 0.67 | 14.33 ± 1.38 | 3055.4 ± 525.95 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skurikhin, E.G.; Pershina, O.; Ermakova, N.; Pakhomova, A.; Zhukova, M.; Pan, E.; Sandrikina, L.; Widera, D.; Kogai, L.; Kushlinskii, N.; et al. Cell Therapy with Human Reprogrammed CD8+ T-Cells Has Antimetastatic Effects on Lewis Lung Carcinoma in C57BL/6 Mice. Int. J. Mol. Sci. 2022, 23, 15780. https://doi.org/10.3390/ijms232415780
Skurikhin EG, Pershina O, Ermakova N, Pakhomova A, Zhukova M, Pan E, Sandrikina L, Widera D, Kogai L, Kushlinskii N, et al. Cell Therapy with Human Reprogrammed CD8+ T-Cells Has Antimetastatic Effects on Lewis Lung Carcinoma in C57BL/6 Mice. International Journal of Molecular Sciences. 2022; 23(24):15780. https://doi.org/10.3390/ijms232415780
Chicago/Turabian StyleSkurikhin, Evgenii G., Olga Pershina, Natalia Ermakova, Angelina Pakhomova, Mariia Zhukova, Edgar Pan, Lubov Sandrikina, Darius Widera, Lena Kogai, Nikolai Kushlinskii, and et al. 2022. "Cell Therapy with Human Reprogrammed CD8+ T-Cells Has Antimetastatic Effects on Lewis Lung Carcinoma in C57BL/6 Mice" International Journal of Molecular Sciences 23, no. 24: 15780. https://doi.org/10.3390/ijms232415780
APA StyleSkurikhin, E. G., Pershina, O., Ermakova, N., Pakhomova, A., Zhukova, M., Pan, E., Sandrikina, L., Widera, D., Kogai, L., Kushlinskii, N., Kubatiev, A., Morozov, S. G., & Dygai, A. (2022). Cell Therapy with Human Reprogrammed CD8+ T-Cells Has Antimetastatic Effects on Lewis Lung Carcinoma in C57BL/6 Mice. International Journal of Molecular Sciences, 23(24), 15780. https://doi.org/10.3390/ijms232415780