
Phenotypic and Genotypic Virulence Characterisation of Staphylococcus pettenkoferi Strains Isolated from Human Bloodstream and Diabetic Foot Infections

 , , , , , , and
, , , , , , and 
Abstract
1. Introduction
2. Results
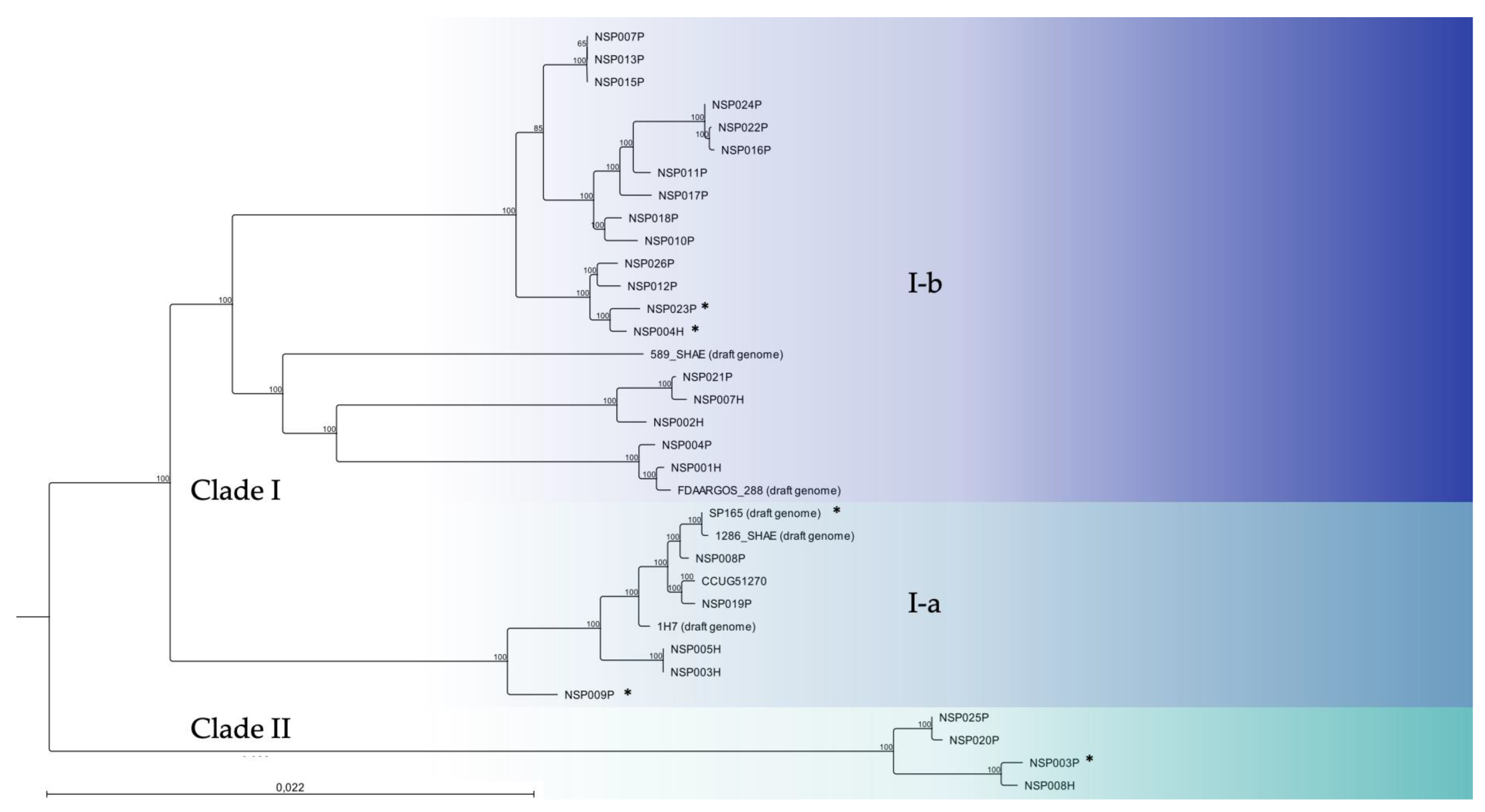
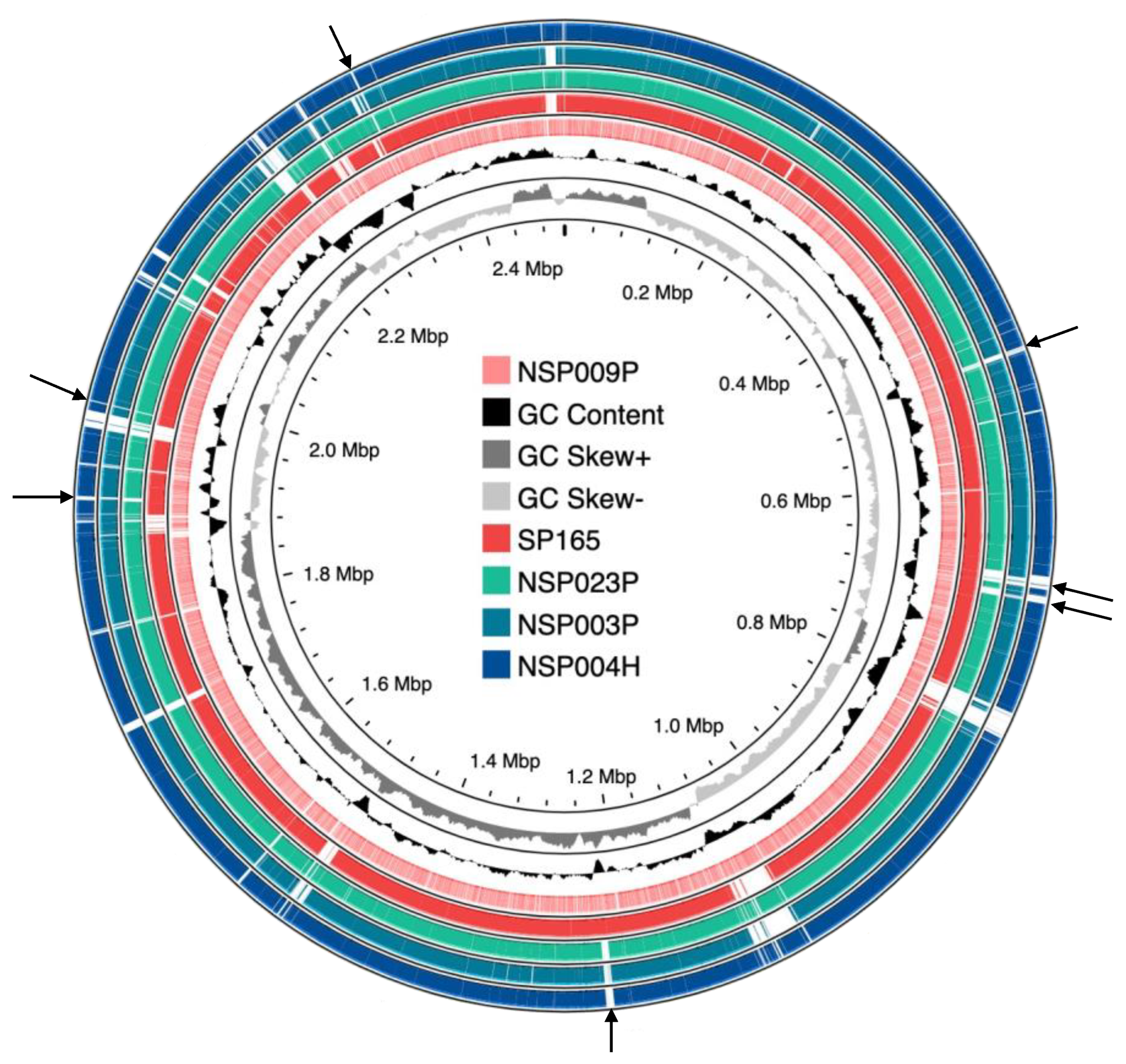
2.1. Genetic Diversity of Clinical S. pettenkoferi Strains
2.2. Resistome of S. pettenkoferi Strains
2.3. Biofilm Capacity of S. pettenkoferi Strains
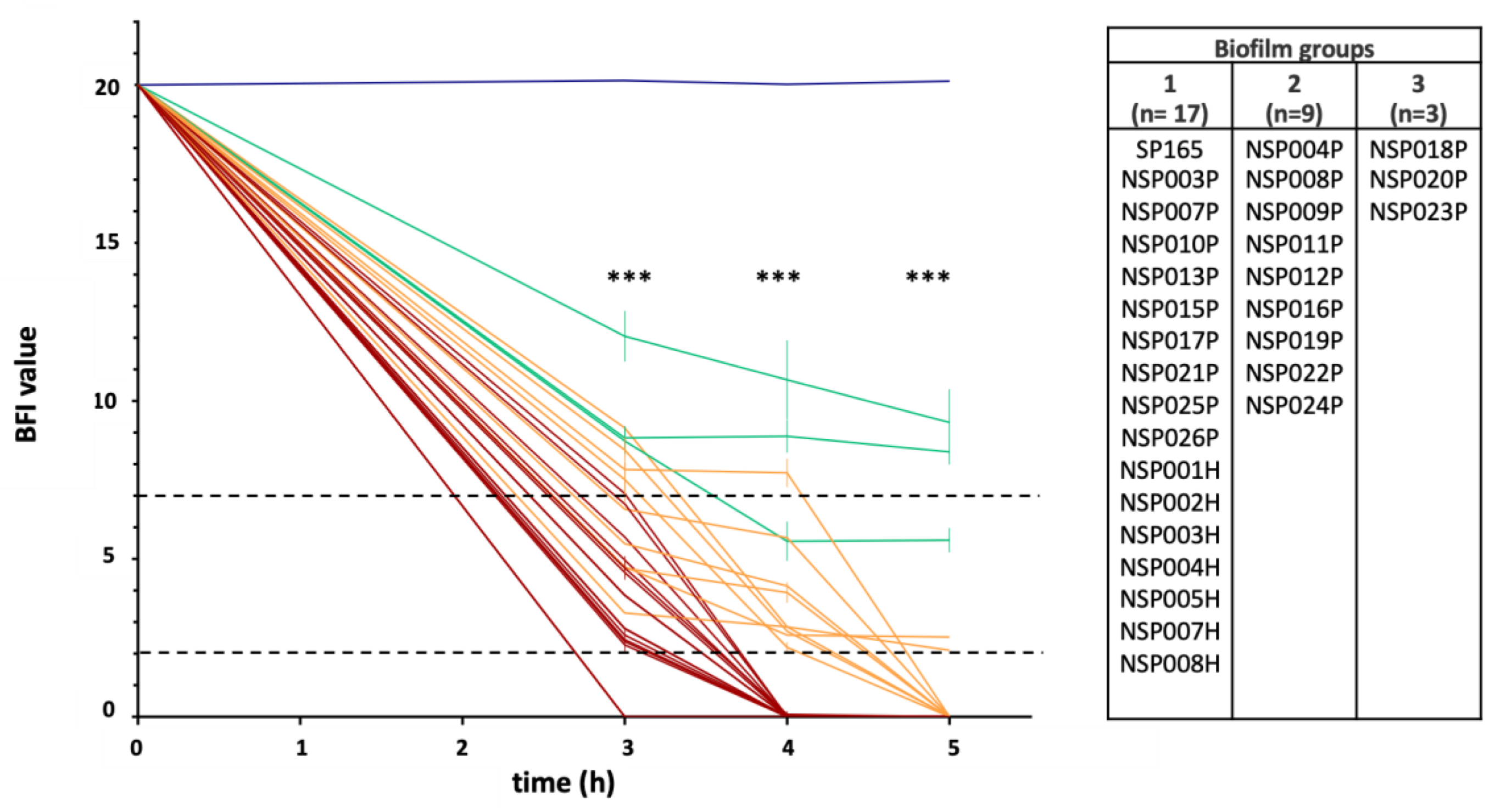
2.3.1. Ability to Form Early Biofilm
2.3.2. Biofilm-Related Genes within S. pettenkoferi Genomes
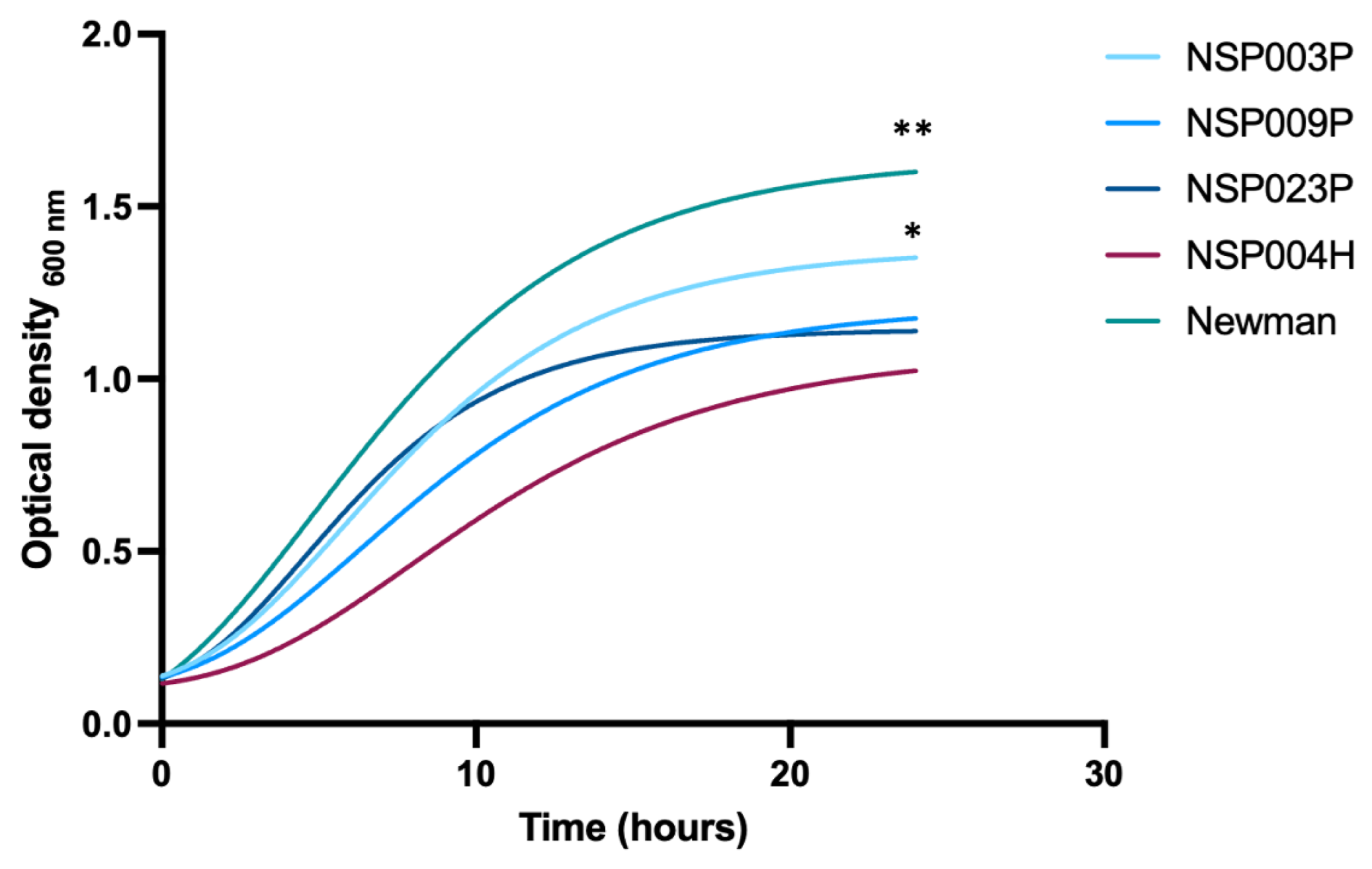
2.4. Fitness of S. pettenkoferi Strains
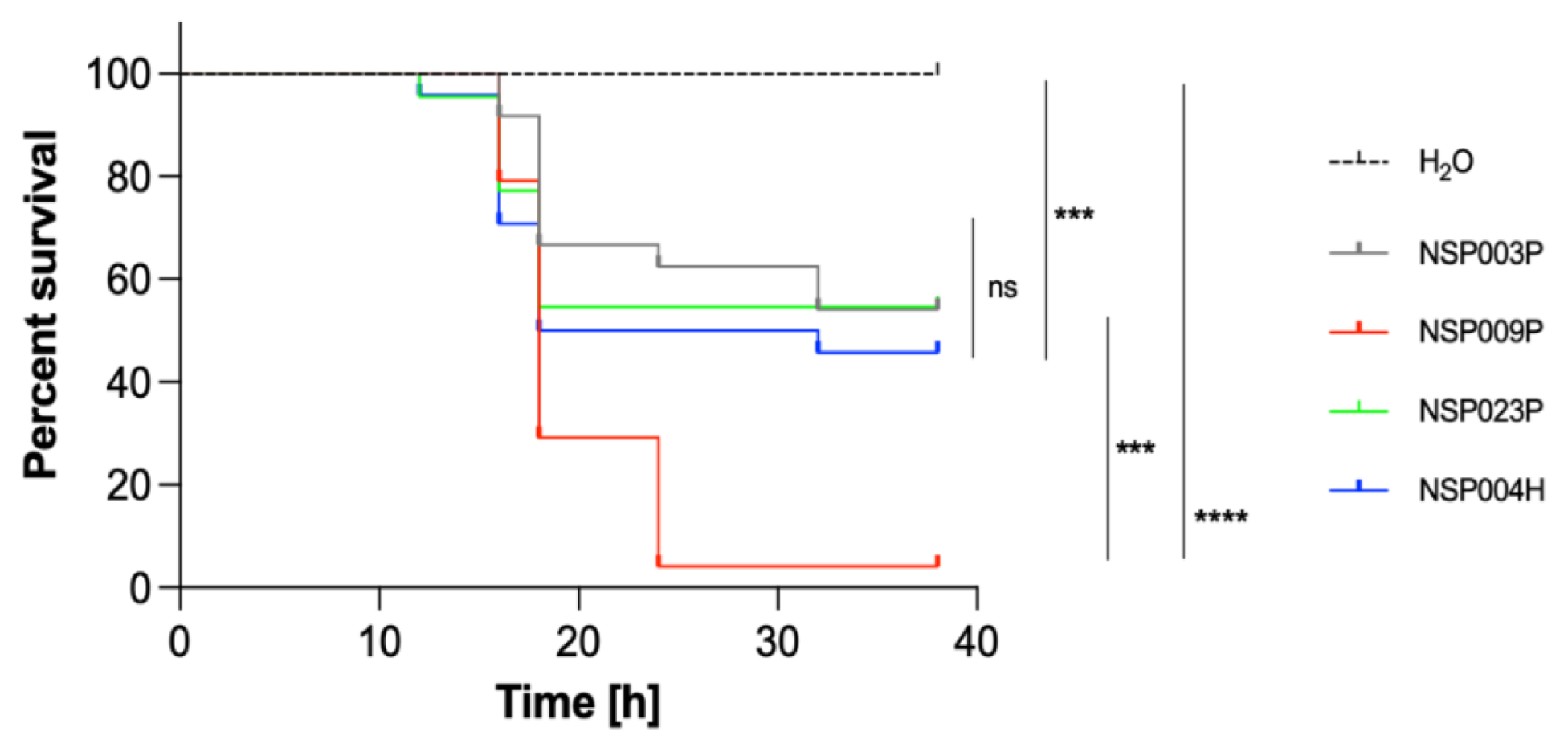
2.5. Virulence Study of S. pettenkoferi Strains Using a Zebrafish Embryos Model
2.6. Genomic Analysis of Putative Virulence Factors of the S. pettenkoferi Strains
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains
4.2. Whole-Genome Sequencing and Bioinformatics Analysis
4.3. Kinetics of Early Biofilm Formation
4.4. Growth Curves Evaluation
4.5. Infection of Danio Rerio Embryos
4.6. Ethical Statement
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IDF Diabetes Atlas—10th Edition. Available online: http://www.diabetesatlas.org (accessed on 1 September 2022).
- Lipsky, B.A.; Senneville, E.; Abbas, Z.G.; Aragón-Sánchez, J.; Diggle, M.; Embil, J.M.; Kono, S.; Lavery, L.A.; Malone, M.; van Asten, S.A.; et al. Guidelines on the Diagnosis and Treatment of Foot Infection in Persons with Diabetes (IWGDF 2019 Update). Diabetes Metab. Res. Rev. 2020, 36, e3280. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, D.G.; Boulton, A.J.M.; Bus, S.A. Diabetic foot ulcers and their recurrence. N. Engl. J. Med. 2017, 376, 2367–2375. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, D.G.; Kanda, V.A.; Lavery, L.A.; Marston, W.; Mills, J.L., Sr.; Boulton, A.J. Mind the gap: Disparity between research funding and costs of care for diabetic foot ulcers. Diabetes Care 2013, 36, 1815–1817. [Google Scholar] [CrossRef] [PubMed]
- Cascini, S.; Agabiti, N.; Davoli, M.; Uccioli, L.; Meloni, M.; Giurato, L.; Marino, C.; Bargagli, A.M. Survival and factors predicting mortality after major and minor lower-extremity amputations among patients with diabetes: A population-based study using Health information systems. BMJ Open Diabetes Res. Care 2020, 8, e001355. [Google Scholar] [CrossRef] [PubMed]
- Barshes, N.R.; Sigireddi, M.; Wrobel, J.S.; Mahankali, A.; Robbins, J.M.; Kougias, P.; Armstrong, D.G. The system of care for the diabetic foot: Objectives, outcomes, and opportunities. Diabet. Foot Ankle 2013, 4, 21847. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Ponsero, A.J.; Armstrong, D.G.; Lipsky, B.A.; Hurwitz, B.L. The dynamic wound microbiome. BMC Med. 2020, 18, 358. [Google Scholar] [CrossRef]
- Radzieta, M.; Sadeghpour-Heravi, F.; Peters, T.J.; Hu, H.; Vickery, K.; Jeffries, T.; Dickson, H.G.; Schwarzer, S.; Jensen, S.O.; Malone, M. A multiomics approach to identify host-microbe alterations associated with infection severity in diabetic foot infections: A pilot study. NPJ Biofilms Microbiomes 2021, 7, 29. [Google Scholar] [CrossRef]
- Jneid, J.; Cassir, N.; Schuldiner, S.; Jourdan, N.; Sotto, A.; Lavigne, J.-P.; La Scola, B. Exploring the microbiota of diabetic foot infections with culturomics. Front. Cell. Infect. Microbiol. 2018, 8, 282. [Google Scholar] [CrossRef]
- Dunyach-Remy, C.; Ngba Essebe, C.; Sotto, A.; Lavigne, J.-P. Staphylococcus aureus toxins and diabetic foot ulcers: Role in pathogenesis and interest in diagnosis. Toxins 2016, 8, 209. [Google Scholar] [CrossRef]
- Désidéri-Vaillant, C.; Nédelec, Y.; Guichon, J.-M.; Le Louarn, S.; Noyer, V.; Sapin-Lory, J.; Le Guen, P.; Nicolas, X. Staphylococcus simulans osteitis in a diabetic patient. Diabetes Metab. 2011, 37, 560–562. [Google Scholar] [CrossRef]
- Nguyen, M.T.T.; Ahern, N.R.; Train, M.K. Staphylococcus schleiferi diabetic foot osteomyelitis and bacteraemia in an immunocompromised host. BMJ Case Rep. 2020, 13, e238302. [Google Scholar] [CrossRef] [PubMed]
- Loïez, C.; Wallet, F.; Pischedda, P.; Renaux, E.; Senneville, E.; Mehdi, N.; Courcol, R.J. First case of osteomyelitis caused by “Staphylococcus pettenkoferi”. J. Clin. Microbiol. 2007, 45, 1069–1071. [Google Scholar] [CrossRef]
- Argemi, X.; Hansmann, Y.; Prola, K.; Prévost, G. Coagulase-negative staphylococci pathogenomics. Int. J. Mol. Sci. 2019, 20, 1215. [Google Scholar] [CrossRef]
- Trülzsch, K.; Rinder, H.; Trček, J.; Bader, L.; Wilhelm, U.; Heesemann, J. “Staphylococcus pettenkoferi”, a novel staphylococcal species isolated from clinical specimens. Diagn. Microbiol. Infect. Dis. 2002, 43, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Vecchia, M.; Lombardi, A.; Zuccaro, V.; Mariani, B.; Morea, A.; Asperges, E.; Grecchi, C.; Sacchi, P.; Bruno, R. A clinical case of sepsis due to Staphylococcus pettenkoferi. Infez. Med. 2018, 26, 369–373. [Google Scholar] [PubMed]
- d’Azevedo, P.A.; Comin, G.; Cantarelli, V. Characterization of a new coagulase-negative Staphylococcus species (Staphylococcus pettenkoferi) isolated from blood cultures from a hospitalized patient in Porto Alegre, Brazil. Rev. Soc. Bras. Med. Trop. 2010, 43, 331–332. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Park, S.; Chung, H.-S.; Lee, M. Clinical and microbiological characteristics of six Staphylococcus pettenkoferi isolates from blood samples. Ann. Lab. Med. 2015, 35, 250–253. [Google Scholar] [CrossRef] [PubMed]
- Song, S.H.; Park, J.S.; Kwon, H.R.; Kim, S.H.; Kim, H.B.; Chang, H.E.; Park, K.U.; Song, J.; Kim, E.C. Human bloodstream infection caused by Staphylococcus pettenkoferi. J. Med. Microbiol. 2009, 58, 270–272. [Google Scholar] [CrossRef]
- Kang, H.; Ryoo, N. Identification of Staphylococcus pettenkoferi isolated from blood culture. Ann. Clin. Microbiol. 2019, 22, 77. [Google Scholar] [CrossRef]
- Hadano, Y.; Hijikata, T.; Miura, A.; Fujii, S.; Awaya, Y. Staphylococcus pettenkoferi bacteremia in a tertiary care hospital in Japan: Report of three cases. J. Infect. Chemother. 2022, 28, 1424–1426. [Google Scholar] [CrossRef]
- Strong, C.; Cosiano, M.; Cabezas, M.; Barwatt, J.W.; Tillekeratne, L.G. Staphylococcus pettenkoferi bacteremia in an American intensive care unit. Case Rep. Infect. Dis. 2021, 2021, 5235691. [Google Scholar] [CrossRef] [PubMed]
- Hashi, A.A.; Delport, J.A.; Elsayed, S.; Silverman, M.S. Staphylococcus pettenkoferi bacteremia: A case report and review of the literature. Can. J. Infect. Dis. Med. Microbiol. 2015, 26, 319–322. [Google Scholar] [CrossRef] [PubMed]
- Eke, U.A.; Fairfax, M.R.; Mitchell, R.; Taylor, M.; Salimnia, H. Staphylococcus pettenkoferi-positive blood cultures in hospitalized patients in a multi-site tertiary center. Diagn. Microbiol. Infect. Dis. 2021, 99, 115284. [Google Scholar] [CrossRef]
- Ugaban, K.B.; She, R.C. The clinical significance of Staphylococcus pettenkoferi: A retrospective review at a tertiary care medical center. Diagn. Microbiol. Infect. Dis. 2022, 102, 115592. [Google Scholar] [CrossRef]
- Kalan, L.; Meisel, J.S.; Loesche, M.A.; Horwinski, J.; Soaita, I.; Chen, X.; Gardner, S.E.; Grice, E.A. Strain and species level variation in the microbiome of diabetic wounds is associated with clinical outcomes and therapeutic efficacy. Cell Host Microbe 2019, 25, 641–655. [Google Scholar] [CrossRef] [PubMed]
- Ahmad-Mansour, N.; Plumet, L.; Huc-Brandt, S.; Magnan, C.; Yahiaoui-Martinez, A.; Kissa, K.; Pantel, A.; Lavigne, J.-P.; Molle, V. Investigating pathogenicity and virulence of Staphylococcus pettenkoferi: An emerging pathogen. Int. J. Mol. Sci. 2021, 22, 13614. [Google Scholar] [CrossRef]
- King, N.P.; Beatson, S.A.; Totsika, M.; Ulett, G.C.; Alm, R.A.; Manning, P.A.; Schembri, M.A. UafB is a serine-rich repeat adhesin of Staphylococcus saprophyticus that mediates binding to fibronectin, fibrinogen and human uroepithelial cells. Microbiology 2011, 157, 1161–1175. [Google Scholar] [CrossRef]
- Wang, Y.; Armato, U.; Wu, J. Targeting tunable physical properties of materials for chronic wound care. Front. Bioeng. Biotechnol. 2020, 8, 584. [Google Scholar] [CrossRef]
- Bakker, K.; Apelqvist, J.; Lipsky, B.A.; Van Netten, J.J.; on behalf of the International Working Group on the Diabetic Foot (IWGDF). The 2015 IWGDF guidance documents on prevention and management of foot problems in diabetes: Development of an evidence-based global consensus. Diabetes Metab. Res. Rev. 2016, 32, 2–6. [Google Scholar] [CrossRef]
- Ravaioli, S.; Selan, L.; Visai, L.; Pirini, V.; Campoccia, D.; Maso, A.; Speziale, P.; Montanaro, L.; Arciola, C.R. Staphylococcus lugdunensis, an aggressive coagulase-negative pathogen not to be underestimated. Int. J. Artif. Organs 2012, 35, 742–753. [Google Scholar] [CrossRef]
- Pouget, C.; Dunyach-Remy, C.; Pantel, A.; Schuldiner, S.; Sotto, A.; Lavigne, J.-P. Biofilms in diabetic foot ulcers: Significance and clinical relevance. Microorganisms 2020, 8, 1580. [Google Scholar] [CrossRef] [PubMed]
- França, A.; Gaio, V.; Lopes, N.; Melo, L.D.R. Virulence factors in coagulase-negative Staphylococci. Pathogens 2021, 10, 170. [Google Scholar] [CrossRef] [PubMed]
- Dutta, T.K.; Chakraborty, S.; Das, M.; Mandakini, R.; Vanrahmlimphuii; Roychoudhury, P.; Ghorai, S.; Behera, S.K. Multidrug-resistant Staphylococcus pettenkoferi isolated from cat in India. Vet. World 2018, 11, 1380–1384. [Google Scholar] [CrossRef]
- Månsson, E.; Hellmark, B.; Stegger, M.; Skytt Andersen, P.; Sundqvist, M.; Söderquist, B. Genomic relatedness of Staphylococcus pettenkoferi isolates of different origins. J. Med. Microbiol. 2017, 66, 601–608. [Google Scholar] [CrossRef]
- Pedroso, S.H.S.P.; Sandes, S.H.C.; Luiz, K.C.M.; Dias, R.S.; Filho, R.A.T.; Serufo, J.C.; Farias, L.M.; Carvalho, M.A.R.; Bomfim, M.R.Q.; Santos, S.G. Biofilm and toxin profile: A phenotypic and genotypic characterization of coagulase-negative staphylococci isolated from human bloodstream infections. Microb. Pathog. 2016, 100, 312–318. [Google Scholar] [CrossRef] [PubMed]
- Szczuka, E.; Krzymińska, S.; Kaznowski, A. Clonality, virulence and the occurrence of genes encoding antibiotic resistance among Staphylococcus warneri isolates from bloodstream infections. J. Med. Microbiol. 2016, 65, 828–836. [Google Scholar] [CrossRef]
- Silva, P.V.; Cruz, R.S.; Keim, L.S.; Paula, G.R.; Carvalho, B.T.; Coelho, L.R.; Carvalho, M.C.; Rosa, J.M.; Figueiredo, A.M.; Teixeira, L.A. The antimicrobial susceptibility, biofilm formation and genotypic profiles of Staphylococcus haemolyticus from bloodstream infections. Mem. Inst. Oswaldo Cruz 2013, 108, 812–813. [Google Scholar] [CrossRef]
- Heilmann, C.; Hussain, M.; Peters, G.; Götz, F. Evidence for autolysin-mediated primary attachment of Staphylococcus epidermidis to a polystyrene surface. Mol. Microbiol. 1997, 24, 1013–1024. [Google Scholar] [CrossRef]
- Qin, Z.; Ou, Y.; Yang, L.; Zhu, Y.; Tolker-Nielsen, T.; Molin, S.; Qu, D. Role of autolysin-mediated DNA release in biofilm formation of Staphylococcus epidermidis. Microbiology 2007, 153, 2083–2092. [Google Scholar] [CrossRef] [PubMed]
- Bowden, M.G.; Chen, W.; Singvall, J.; Xu, Y.; Peacock, S.J.; Valtulina, V.; Speziale, P.; Höök, M. Identification and preliminary characterization of cell-wall-anchored proteins of Staphylococcus epidermidis. Microbiology 2005, 151, 1453–1464. [Google Scholar] [CrossRef]
- Cucarella, C.; Solano, C.; Valle, J.; Amorena, B.; Lasa, Í.; Penadés, J.R. Bap, a Staphylococcus aureus surface protein involved in biofilm formation. J. Bacteriol. 2001, 183, 2888–2896. [Google Scholar] [CrossRef] [PubMed]
- Potter, A.; Ceotto, H.; Giambiagi-deMarval, M.; dos Santos, K.R.N.; Nes, I.F.; Bastos, M.D.C.D.F. The gene bap, involved in biofilm production, is present in Staphylococcus spp. strains from nosocomial infections. J. Microbiol. 2009, 47, 319–326. [Google Scholar] [CrossRef]
- Frank, K.L.; Patel, R. Poly-N-acetylglucosamine is not a major component of the extracellular matrix in biofilms formed by icaADBC-positive Staphylococcus lugdunensis isolates. Infect. Immun. 2007, 75, 4728–4742. [Google Scholar] [CrossRef] [PubMed]
- Schilcher, K.; Horswill, A.R. Staphylococcal biofilm development: Structure, regulation, and treatment strategies. Microbiol. Mol. Biol. Rev. 2020, 84, e00026-19. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Khan, B.A.; Cheung, G.Y.; Bach, T.H.; Jameson-Lee, M.; Kong, K.-F.; Queck, S.Y.; Otto, M. Staphylococcus epidermidis surfactant peptides promote biofilm maturation and dissemination of biofilm-associated infection in mice. J. Clin. Investig. 2011, 121, 238–248. [Google Scholar] [CrossRef]
- Towle, K.M.; Lohans, C.T.; Miskolzie, M.; Acedo, J.Z.; van Belkum, M.J.; Vederas, J.C. Solution structures of phenol-soluble modulins a1, a3, and b2, virulence factors from Staphylococcus aureus. Biochemistry 2016, 55, 4798–4806. [Google Scholar] [CrossRef]
- Cheung, G.Y.C.; Joo, H.S.; Chatterjee, S.S.; Otto, M. Phenol-soluble modulins—Critical determinants of staphylococcal virulence. FEMS Microbiol. Rev. 2014, 38, 698–719. [Google Scholar] [CrossRef]
- Wang, C.; Li, M.; Dong, D.; Wang, J.; Ren, J.; Otto, M.; Gao, Q. Role of ClpP in biofilm formation and virulence of Staphylococcus epidermidis. Microbes Infect. 2007, 9, 1376–1383. [Google Scholar] [CrossRef]
- Leggett, H.C.; Cornwallis, C.K.; Buckling, A.; West, S.A. Growth rate, transmission mode and virulence in human pathogens. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2017, 372, 20160094. [Google Scholar] [CrossRef]
- Gomes, M.C.; Mostowy, S. The case for modeling human infection in zebrafish. Trends Microbiol. 2020, 28, 10–18. [Google Scholar] [CrossRef]
- Torraca, V.; Mostowy, S. Zebrafish infection: From pathogenesis to cell biology. Trends Cell Biol. 2018, 28, 143–156. [Google Scholar] [CrossRef] [PubMed]
- Keightley, M.C.; Wang, C.H.; Pazhakh, V.; Lieschke, G.J. Delineating the roles of neutrophils and macrophages in zebrafish regeneration models. Int. J. Biochem. Cell Biol. 2014, 56, 92–106. [Google Scholar] [CrossRef] [PubMed]
- EUCAST: Clinical Breakpoints and Dosing of Antibiotics. Available online: https://eucast.org/clinical_breakpoints/ (accessed on 1 June 2022).
- McKindles, K.M.; McKay, R.M.; Bullerjahn, G.S. Genomic comparison of Planktothrix agardhii isolates from a Lake Erie embayment. PLoS ONE 2022, 17, e0273454. [Google Scholar] [CrossRef] [PubMed]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for predictions of phenotypes from genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef]
- Zankari, E.; Allesøe, R.; Joensen, K.G.; Cavaco, L.M.; Lund, O.; Aarestrup, F.M. PointFinder: A novel web tool for WGS-based detection of antimicrobial resistance associated with chromosomal point mutations in bacterial pathogens. J. Antimicrob. Chemother. 2017, 72, 2764–2768. [Google Scholar] [CrossRef] [PubMed]
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.Y.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.L.V.; Cheng, A.A.; Liu, S.; et al. CARD 2020: Antibiotic resistome surveillance with the comprehensive antibiotic resistance database. Nucleic Acids Res. 2020, 48, D517–D525. [Google Scholar] [CrossRef]
- Jukes, T.H.; Cantor, C.R. Evolution of protein molecules. In Mammalian Protein Metabolism; Munro, H.N., Ed.; Academic Press Inc.: New York, NY, USA, 1969; Volume 3, pp. 21–132. [Google Scholar]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Grant, J.R.; Stothard, P. The CGView server: A comparative genomics tool for circular genomes. Nucleic Acids Res. 2008, 36, W181–W184. [Google Scholar] [CrossRef]
- Chavant, P.; Gaillard-Martinie, B.; Talon, R.; Hébraud, M.; Bernardi, T. A new device for rapid evaluation of biofilm formation potential by bacteria. J. Microbiol. Methods 2007, 68, 605–612. [Google Scholar] [CrossRef]
- Tjørve, K.M.C.; Tjørve, E. The use of Gompertz models in growth analyses, and new Gompertz-model approach: An addition to the Unified-Richards family. PLoS ONE 2017, 12, e0178691. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate ID | Year | Type of Sampling | Hospital Department | Patients | Resistance Profile | |
|---|---|---|---|---|---|---|
| Age | Sex | |||||
| SP165 | 2018 | Tissue biopsy | Diabetic foot clinic | 60 | M | PEN, OXA, ERY, CMN, L, OFX, RIF, FOS |
| NSP003P | 2020 | Tissue biopsy | Diabetic foot clinic | 58 | M | / |
| NSP004P | 2020 | Tissue biopsy | Diabetic foot clinic | 89 | M | OFX, FA, FOS |
| NSP007P | 2020 | Bone biopsy | Diabetic foot clinic | 69 | M | PEN, FA |
| NSP008P | 2020 | Bone biopsy | Vascular surgery | 90 | M | FOS |
| NSP009P | 2020 | Bone biopsy | Diabetic foot clinic | 62 | M | PEN, ERY, CMN, L, PT, SYN, TET, OFX, FA, FOS |
| NSP010P | 2020 | Bone biopsy | Orthopaedic surgery | 61 | F | FOS |
| NSP011P | 2020 | Tissue biopsy | Diabetic foot clinic | 89 | M | / |
| NSP012P | 2021 | Bone biopsy | Orthopaedic surgery | 67 | M | PEN, OFX, FA |
| NSP013P | 2021 | Tissue biopsy | Diabetic foot clinic | 94 | M | PEN, OFX |
| NSP015P | 2021 | Bone biopsy | Orthopaedic surgery | 64 | M | FOS |
| NSP016P | 2021 | Bone biopsy | Diabetic foot clinic | 56 | M | FA |
| NSP017P | 2021 | Tissue biopsy | Diabetic foot clinic | 84 | M | / |
| NSP018P | 2021 | Bone biopsy | Vascular surgery | 76 | M | / |
| NSP019P | 2021 | Bone biopsy | Vascular surgery | 71 | M | FOS |
| NSP020P | 2021 | Tissue biopsy | Orthopaedic surgery | 63 | M | / |
| NSP021P | 2021 | Bone biopsy | Diabetic foot clinic | 60 | M | PEN, FOS |
| NSP022P | 2021 | Tissue biopsy | Diabetic foot clinic | 66 | F | FA |
| NSP023P | 2021 | Bone biopsy | Orthopaedic surgery | 94 | F | PEN |
| NSP024P | 2021 | Tissue biopsy | Diabetic foot clinic | 78 | F | PEN |
| NSP025P | 2022 | Tissue biopsy | Diabetic foot clinic | 93 | M | / |
| NSP026P | 2022 | Tissue biopsy | Diabetic foot clinic | 63 | M | PEN |
| NSP001H | 2020 | Blood culture | Geriatric | 83 | M | PEN, OFX, FA, FOS |
| NSP002H | 2020 | Blood culture | Emergency | 59 | F | ERY, FOS |
| NSP003H | 2020 | Blood culture | Emergency | 65 | M | FA, FOS |
| NSP004H | 2021 | Blood culture | Polyvalent medicine | 44 | F | PEN |
| NSP005H | 2021 | Blood culture | Intensive care unit | 58 | M | FOS |
| NSP007H | 2022 | Blood culture | Neurology | 72 | M | PEN, FOS |
| NSP008H | 2022 | Blood culture | Intensive care unit | 62 | M | FOS |
| Resistance Genes | Blood Culture Strains (n = 7) | DFU Strains (n = 22) | Total (n = 29) | p-Value |
|---|---|---|---|---|
| Blood vs. DFU | ||||
| blaZ | 3 (42.9%) | 9 (40.9%) | 12 (41.4%) | 1 |
| mecA | 0 (0%) | 1 (4.5%) | 1 (3.4%) | 1 |
| ermA | 0 (0%) | 3 (13.6%) | 3 (10.3%) | 0.56 |
| vgaA | 3 (42.9%) | 3 (13.6%) | 6 (20.7%) | 0.13 |
| gyrA | 1 (14.3%) | 5 (22.7%) | 6 (20.7%) | 1 |
| grlA | 6 (87.5%) | 17 (77.2%) | 23 (79.3%) | 1 |
| grlB | 1 (14.3%) | 13 (59%) | 14 (48.3%) | 0.08 |
| tetk | 0 (0%) | 1 (4.5%) | 1 (3.4%) | 1 |
| rpoB | 0 (0%) | 1 (4.5%) | 1 (3.4%) | 1 |
| fusB | 1 (14.3%) | 7 (31.8%) | 8 (27.6%) | 0.63 |
| ant(9)-Ia | 0 (0%) | 3 (13.6%) | 3 (10.3%) | 0.56 |
| fosB | 4 (57.1%) | 6 (27.3%) | 11 (37.9%) | 0.19 |
| Biofilm Steps | Putative Genetic Determinants of Virulence | Blood Culture (n = 7) | DFU (n = 22) | p-Value B vs. D | Biofilm Group 1 (n = 17) | Biofilm Group 2 (n = 9) | Biofilm Group 3 (n = 3) | p-Value G1 vs. G2 vs. G3 | Total (n = 29) | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| INITIAL ADHESION | To abiotic surfaces | bhp | Biofilm associated protein | 0 (0%) | 2 (9%) | 1 | 0 (0%) | 1 (11%) | 1 (33%) | 0.07 | 2 (6.9%) |
| atlE | Autolysin E | 7 (100%) | 22 (100%) | - | 17 (100%) | 9 (100%) | 3 (100%) | - | 29 (100%) | ||
| sesC, sasC, sasG | Surface proteins | 0 (0%) | 0 (0%) | - | 0 (0%) | 0 (0%) | 0 (0%) | - | 0 (0%) | ||
| embp | Extracellular matrix binding protein Embp | 0 (0%) | 0 (0%) | - | 0 (0%) | 0 (0%) | 0 (0%) | - | 0 (0%) | ||
| To biotic surfaces | fnbpA, fnbpB | Fibronectin binding protein A and B | 0 (0%) | 0 (0%) | - | 0 (0%) | 0 (0%) | 0 (0%) | - | 0 (0%) | |
| fbl, fib | Fibrinogen binding protein | 0 (0%) | 0 (0%) | - | 0 (0%) | 0 (0%) | 0 (0%) | - | 0 (0%) | ||
| clfA, clfB | Clumping factors A and B | 0 (0%) | 0 (0%) | - | 0 (0%) | 0 (0%) | 0 (0%) | - | 0 (0%) | ||
| sdrCDEFH | Serine-aspartate repeat-containing protein C,D,E,F,H | 0 (0%) | 0 (0%) | - | 0 (0%) | 0 (0%) | 0 (0%) | - | 0 (0%) | ||
| sdrD B-like domain | B-like domain from the SdrD protein | 4 (57.1%) | 5 (22.7%) | 0.15 | 7 (41.2%) | 1 (11%) | 1 (33%) | 0.24 | 9 (31%) | ||
| MATURATION | Intercellular aggregation | icaA, icaB, icaC | Polysaccharide Intercellular Adhesion (PIA) | 7 (100%) | 22 (100%) | - | 17 (100%) | 9 (100%) | 3 (100%) | - | 29 (100%) |
| icaD | Polysaccharide Intercellular Adhesion (PIA) | 0 (0%) | 0 (0%) | - | 0 (0%) | 0 (0%) | 0 (0%) | - | 0 (0%) | ||
| isdA, isdB | Iron-regulated surface determinant protein A,B | 0 (0%) | 0 (0%) | - | 0 (0%) | 0 (0%) | 0 (0%) | - | 0 (0%) | ||
| aap | Accumulation associated protein | 0 (0%) | 0 (0%) | - | 0 (0%) | 0 (0%) | 0 (0%) | - | 0 (0%) | ||
| sbp | S. epidermidis biofilm accumulation | 0 (0%) | 0 (0%) | - | 0 (0%) | 0 (0%) | 0 (0%) | - | 0 (0%) | ||
| Biofilm structuration | splABCDE | Serine proteases SplA, SplB, SplC, SplD, SplE | 0 (0%) | 0 (0%) | - | 0 (0%) | 0 (0%) | 0 (0%) | - | 0 (0%) | |
| splF | Serine protease SplF | 7 (100%) | 22 (100%) | - | 17 (100%) | 9 (100%) | 3 (100%) | - | 29 (100%) | ||
| clpP | ClpP protease | 7 (100%) | 22 (100%) | - | 17 (100%) | 9 (100%) | 3 (100%) | - | 29 (100%) | ||
| SSP1763 | Serine protease HtrA-like | 7 (100%) | 22 (100%) | - | 17 (100%) | 9 (100%) | 3 (100%) | - | 29 (100%) | ||
| HMPREF2802_03390 | Serine protease | 7 (100%) | 22 (100%) | - | 17 (100%) | 9 (100%) | 3 (100%) | - | 29 (100%) | ||
| REGULATION | icaR | Biofilm operon icaADBC regulator | 0 (0%) | 0 (0%) | - | 0 (0%) | 0 (0%) | 0 (0%) | - | 0 (0%) | |
| agrA, agrC | Accessory genes regulators A and C | 7 (100%) | 22 (100%) | - | 17 (100%) | 9 (100%) | 3 (100%) | - | 29 (100%) | ||
| agrB, agrD | Accessory gene sregulators B and D | 0 (0%) | 0 (0%) | - | 0 (0%) | 0 (0%) | 0 (0%) | - | 0 (0%) | ||
| sigB | Sigma B | 7 (100%) | 22 (100%) | - | 17 (100%) | 9 (100%) | 3 (100%) | - | 29 (100%) | ||
| rsbUVW | / | 7 (100%) | 22 (100%) | - | 17 (100%) | 9 (100%) | 3 (100%) | - | 29 (100%) | ||
| lytSR | LytSR two component regulatory system | 7 (100%) | 22 (100%) | - | 17 (100%) | 9 (100%) | 3 (100%) | - | 29 (100%) | ||
| arlRS | ArlRS two-component system | 7 (100%) | 22 (100%) | - | 17 (100%) | 9 (100%) | 3 (100%) | - | 29 (100%) | ||
| mgrA | HTH-type transcription regulator MgrA | 7 (100%) | 22 (100%) | - | 17 (100%) | 9 (100%) | 3 (100%) | - | 29 (100%) | ||
| cidA | Holin-like protein CidA | 6 (85.7%) | 17 (77.3%) | 1 | 12 (70.6%) | 9 (100%) | 2 (67%) | 0.22 | 23 (79.3%) | ||
| lrgA | Antiholin-like protein LrgA | 7 (100%) | 22 (100%) | - | 17 (100%) | 9 (100%) | 3 (100%) | - | 29 (100%) | ||
| sarA | Transcriptional regulator SarA | 7 (100%) | 22 (100%) | - | 17 (100%) | 9 (100%) | 3 (100%) | - | 29 (100%) | ||
| luxS | S-ribosylhomocysteine lyase | 7 (100%) | 22 (100%) | - | 17 (100%) | 9 (100%) | 3 (100%) | - | 29 (100%) | ||
| DISPERSION | BUY34_08050 | Hemolysin H3C: Phenol soluble modulin beta protein | 7 (100%) | 22 (100%) | - | 17 (100%) | 9 (100%) | 3 (100%) | - | 29 (100%) | |
| Putative Genetic Determinants of Virulence | Blood Culture (n = 7) | DFU (n = 22) | p-Value B vs. D | Biofilm Group 1 (n = 17) | Biofilm Group 2 (n = 9) | Biofilm Group 3 (n = 3) | p-Value G1 vs. G2 vs. G3 | Total (n = 29) | ||
|---|---|---|---|---|---|---|---|---|---|---|
| Toxins | yqfA | Hemolysin III | 7 (100%) | 22 (100%) | - | 17 (100%) | 9 (100%) | 3 (100%) | - | 29 (100%) |
| hld | Delta hemolysin | 5 (71.4%) | 18 (81.8%) | 0.61 | 14 (82.4%) | 6 (66.7%) | 3 (100%) | 0.53 | 23 (79.3%) | |
| ERS010499_02103 | Hemolysin H1U | 6 (85.7%) | 9 (40.9%) | 0.0801 | 11 (65.7%) | 4 (44.4%) | 0 (0%) | 0.1218 | 15 (51.7%) | |
| hly (hla), hlb, hlg | α- β- γ-hemolysins | 0 (0%) | 0 (0%) | - | 0 (0%) | 0 (0%) | 0 (0%) | - | 0 (0%) | |
| sea, sel, seg, sed | Enterotoxins | 0 (0%) | 0 (0%) | - | 0 (0%) | 0 (0%) | 0 (0%) | - | 0 (0%) | |
| tst-1 | Toxic shock syndrome toxin | 0 (0%) | 0 (0%) | - | 0 (0%) | 0 (0%) | 0 (0%) | - | 0 (0%) | |
| etA, etB, etD | Exfoliatin | 0 (0%) | 0 (0%) | - | 0 (0%) | 0 (0%) | 0 (0%) | - | 0 (0%) | |
| lukDE | Leukotoxin LukD and LukE | 0 (0%) | 0 (0%) | - | 0 (0%) | 0 (0%) | 0 (0%) | - | 0 (0%) | |
| Avoid Host Immune Response and Invasiveness | capA | Capsule biosynthesis protein CapA | 7 (100%) | 22 (100%) | - | 17 (100%) | 9 (100%) | 3 (100%) | - | 29 (100%) |
| eno | Enolase | 7 (100%) | 22 (100%) | - | 17 (100%) | 9 (100%) | 3 (100%) | - | 29 (100%) | |
| katA | Catalase | 7 (100%) | 22 (100%) | - | 17 (100%) | 9 (100%) | 3 (100%) | - | 29 (100%) | |
| / | esXA: Type VII secretion system (T7SS) substrate | 2 (28.6%) | 4 (18.2%) | 0.61 | 4 (23.5%) | 1 (11.1%) | 1 (33%) | 0.67 | 6 (20.7%) | |
| T7SSB apparatus encoding genes | T7SSb apparatus: EsaA, EsaB, EssA, EssB and EssC | 1 (14.3%) | 1 (4.5%) | 0.43 | 1 (5.9%) | 1 (11.1%) | 0 (0%) | 1 | 2 (6.9%) | |
| tuf | Elongation factor tu | 7 (100%) | 22 (100%) | - | 17 (100%) | 9 (100%) | 3 (100%) | - | 29 (100%) | |
| lspA | Lipoprotein signal peptidase | 7 (100%) | 22 (100%) | - | 17 (100%) | 9 (100%) | 3 (100%) | - | 29 (100%) | |
| lgt | Lipoprotein diacylglycerol transferase | 7 (100%) | 22 (100%) | - | 17 (100%) | 9 (100%) | 3 (100%) | - | 29 (100%) | |
| nucH | Thermonuclease H | 7 (100%) | 22 (100%) | - | 17 (100%) | 9 (100%) | 3 (100%) | - | 29 (100%) | |
| coA | Coagulase | 0 (0%) | 0 (0%) | - | 0 (0%) | 0 (0%) | 0 (0%) | - | 0 (0%) | |
| spA | Staphylococcus protein A | 0 (0%) | 0 (0%) | - | 0 (0%) | 0 (0%) | 0 (0%) | - | 0 (0%) | |
| vwb | Von Willebrand factor binding protein Vwb | 0 (0%) | 0 (0%) | - | 0 (0%) | 0 (0%) | 0 (0%) | - | 0 (0%) | |
| saK | Staphylokinase | 0 (0%) | 0 (0%) | - | 0 (0%) | 0 (0%) | 0 (0%) | - | 0 (0%) | |
| Protease activity or implicated in proteolysis | clpC, clpX, clpXP | Clp protases | 7 (100%) | 22 (100%) | - | 17 (100%) | 9 (100%) | 3 (100%) | - | 29 (100%) |
| mecA | Adapter protein MecA | 7 (100%) | 22 (100%) | - | 17 (100%) | 9 (100%) | 3 (100%) | - | 29 (100%) | |
| aur | Aureolysin | 0 (0%) | 0 (0%) | - | 0 (0%) | 0 (0%) | 0 (0%) | - | 0 (0%) | |
| Regulation | nsaS | Sensor histidine kinase | 7 (100%) | 22 (100%) | - | 17 (100%) | 9 (100%) | 3 (100%) | - | 29 (100%) |
| nsaR | DNA-binding response regulator | 0 (0%) | 0 (0%) | - | 0 (0%) | 0 (0%) | 0 (0%) | - | 0 (0%) | |
| srrB | Sensor protein SrrB | 7 (100%) | 22 (100%) | - | 17 (100%) | 9 (100%) | 3 (100%) | - | 29 (100%) | |
| srrA | Transcriptional regulatory protein SrrA | 0 (0%) | 0 (0%) | - | 0 (0%) | 0 (0%) | 0 (0%) | - | 0 (0%) | |
| vraSR | Sensor Pt VraS/ Response regulator pt VraR | 7 (100%) | 22 (100%) | - | 17 (100%) | 9 (100%) | 3 (100%) | - | 29 (100%) | |
| saeSR | Histidine protein kinase SaeS/ Response regulator SaeR | 7 (100%) | 22 (100%) | - | 17 (100%) | 9 (100%) | 3 (100%) | - | 29 (100%) | |
| Others | virE | Virulence-associated protein E | 0 (0%) | 2 (9.1%) | 1 | 0 (0%) | 1 (11%) | 1 (33%) | 0.07 | 2 (6.9%) |
| cvfB, cvfC | Conserved virulence factor B and C | 7 (100%) | 22 (100%) | - | 17 (100%) | 9 (100%) | 3 (100%) | - | 29 (100%) | |
| CJ235_09000 | α- β- hydrolase | 7 (100%) | 22 (100%) | - | 17 (100%) | 9 (100%) | 3 (100%) | - | 29 (100%) | |
| isdF | Iron-regulated surface determinant protein F | 7 (100%) | 22 (100%) | - | 17 (100%) | 9 (100%) | 3 (100%) | - | 29 (100%) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Magnan, C.; Ahmad-Mansour, N.; Pouget, C.; Morsli, M.; Huc-Brandt, S.; Pantel, A.; Dunyach-Remy, C.; Sotto, A.; Molle, V.; Lavigne, J.-P. Phenotypic and Genotypic Virulence Characterisation of Staphylococcus pettenkoferi Strains Isolated from Human Bloodstream and Diabetic Foot Infections. Int. J. Mol. Sci. 2022, 23, 15476. https://doi.org/10.3390/ijms232415476
Magnan C, Ahmad-Mansour N, Pouget C, Morsli M, Huc-Brandt S, Pantel A, Dunyach-Remy C, Sotto A, Molle V, Lavigne J-P. Phenotypic and Genotypic Virulence Characterisation of Staphylococcus pettenkoferi Strains Isolated from Human Bloodstream and Diabetic Foot Infections. International Journal of Molecular Sciences. 2022; 23(24):15476. https://doi.org/10.3390/ijms232415476
Chicago/Turabian StyleMagnan, Chloé, Nour Ahmad-Mansour, Cassandra Pouget, Madjid Morsli, Sylvaine Huc-Brandt, Alix Pantel, Catherine Dunyach-Remy, Albert Sotto, Virginie Molle, and Jean-Philippe Lavigne. 2022. "Phenotypic and Genotypic Virulence Characterisation of Staphylococcus pettenkoferi Strains Isolated from Human Bloodstream and Diabetic Foot Infections" International Journal of Molecular Sciences 23, no. 24: 15476. https://doi.org/10.3390/ijms232415476
APA StyleMagnan, C., Ahmad-Mansour, N., Pouget, C., Morsli, M., Huc-Brandt, S., Pantel, A., Dunyach-Remy, C., Sotto, A., Molle, V., & Lavigne, J.-P. (2022). Phenotypic and Genotypic Virulence Characterisation of Staphylococcus pettenkoferi Strains Isolated from Human Bloodstream and Diabetic Foot Infections. International Journal of Molecular Sciences, 23(24), 15476. https://doi.org/10.3390/ijms232415476