Abstract
Plant miRNAs are powerful regulators of gene expression at the post-transcriptional level, which was repeatedly proved in several model plant species. miRNAs are considered to be key regulators of many developmental, homeostatic, and immune processes in plants. However, our understanding of plant miRNAs is still limited, despite the fact that an increasing number of studies have appeared. This systematic review aims to summarize our current knowledge about miRNAs in spring barley (Hordeum vulgare), which is an important agronomical crop worldwide and serves as a common monocot model for studying abiotic stress responses as well. This can help us to understand the connection between plant miRNAs and (not only) abiotic stresses in general. In the end, some future perspectives and open questions are summarized.
1. Introduction
MicroRNAs (miRNAs) are important players in post-transcriptional gene expression regulation in multicellular species. miRNAs can modify/decrease the expression of fully or partially complementary mRNA molecules [,]. Original reports about RNA silencing, where miRNAs belong, date back to the 1990s [] when the first attempt to introduce a chimeric chalcone synthase using Agrobacterium tumefaciens vector led to the decrease or complete loss of anthocyanin pigmentation in petals (flowers) of Petunia hybrida []. A similar observation was in 1992 documented also for the Neurospora crassa where the transformation using plasmids containing artificial constructs led to the albino phenotype []. Another experiment in 1993 carried out by Victor Ambros, Rosalind Lee and Rhonda Feinbaum resulted in the final revelation that the lin-4 gene (involved in the regulation of Caenorhabditis elegans developmental events) codes not for a protein, but for small RNA with regulatory function [], and such RNAs were later called miRNAs and their nomenclature was established []. Today, miRNAs are considered to be master regulators of many cell differentiation, developmental, and homeostatic processes in animals [] and plants [,], and are also widely accepted as an important component of the cellular immune system, which is documented even for plant species [,,,].
Despite the fact that the number of studies dealing with plant miRNAs during the past few years has steeply increased (from 11 articles in 2002 to the current 928 in 2021), our complex understanding of their role in environmental stress responses remains limited, see, e.g., current review dealing with miRNA regulation and stress adaptation in plants []. Our specific focus on barley stems from the importance of this agronomical crop, which was one of the first cultivated grains as early as 10,000 years ago []. Barley is currently ranked 4th in worldwide production after wheat, maize, and rice, and is extensively used in food production, for feeding cattle, or brewery worldwide [,]. Additionally, barley plants serve as an important commodity with potential health benefits—from hail to barley grass food supplements. Effects on the gut microbiota and suppression of already developed chronic diseases including obesity, diabetes, circulatory disorders, and cancer were documented or at least hypothesized [,]. An additional and equally important application of barley seedlings is its use as a monocot model species (often considered as a model plant for the whole Triticeae tribe). Barley has a short cultivation period from seed planting to sampling the material (14 days) and does not require complex growth conditions which makes it a suitable species for a wide range of experiments [,,]. Lastly, barley has a sequenced reference genome (cultivar Morex, NCBI ID: GCF_904849725.1) of a length of 4.27 Gbps consisting of 7 chromosomes, and circular chloroplastic DNA []. More than one-half of all 58,438 predicted genes (53%; total of 31,449) are protein-coding genes, while approximately one-tenth are pseudogenes (9.9%; total of 5778) []. Most importantly, almost one-third of the barley genes are coding for small RNAs (30.3%; total of 17,729) [] which further highlights their functional relevance in gene expression regulation and justifies the need to properly understand their involvement in physiological and stress-related processes. The rest of the genes are uncharacterized (6%; a total of 3481) []. For the above-mentioned reasons, we decided to perform a review focused specifically on spring barley (Hordeum vulgare) miRNAs. In the following chapters, we will briefly discuss plant miRNAs biogenesis and regulatory potential in general, then we will move to specific roles of miRNAs in barley physiology and stress responses, conserved barley miRNAs and their high-confidence mRNA targets, and finally give some possible future directions of research in this field, with focus on barley.
2. Plant miRNAs—Biogenesis and Regulatory Potential
Mature plant miRNAs are 19–25-nucleotide-long ribonucleic acids that can have either intergenic (miRNA gene is localized between two protein-coding sequences of the DNA) or intragenic origin [], where miRNAs are cleaved from the mRNA sequences during the splicing (also called intron-derived miRNAs []). Specifically, in barley, more than 75% of miRNAs are transcribed from intergenic loci []. The biogenesis of miRNAs is ensured by the DNA-dependent RNA polymerase II which is responsible for the biosynthesis itself []. In some cases, multiple plant miRNAs are synthesized all at once (multiple miRNAs localized in one long transcript) [] and often form a miRNA family, which is a group of miRNAs derived from a common ancestor []. Emerging miRNAs can be modified co-transcriptionally, or post-transcriptionally. Similar to other transcripts, a 7-methylguanosine (m7G) cap is attached to the 5′ end of the miRNA, and the 3′ end is polyadenylated (or can be spliced) []. Later, the transcript encoding miRNA (or multiple miRNAs) is folded to the stem-loop structure which is called pri-miRNA [] (meaning primary miRNA transcript). Such pri-miRNAs are further cleaved by the dicing bodies. Dicing bodies consists of several proteins including DICER-LIKE 1 (DCL1), DAWDLE (DDL), HYL1, TGH, and SE [,], resulting in miRNA duplex formation which can be later 2′-O-methylated by the HEN1 methylase [] and incorporated into the RNA-induced silencing complex (RISC) [,]. The complex issue of further proteins involved in plant miRNA biogenesis is reviewed in Li et al., 2021 []. miRNAs of both origins (intragenic as well as intergenic) lead to the formation of a mature RISC with incorporated mature miRNA. In most cases, only the sense/guide miRNA strand is incorporated into the RISC, while the antisense/passenger miRNA (miRNA*) strand is disrupted, but recently also the regulation potential of the passenger miRNA became the center of interest [,,]. For a clear summary of miRNA biogenesis see Figure 1 below.
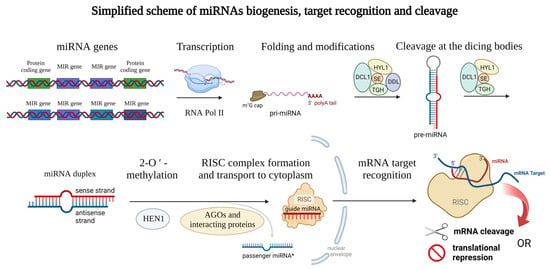
Figure 1.
Schematic representation of the miRNAs biogenesis. Genes encoding miRNAs are transcribed by RNA polymerase II and modified on their ends (m7G cap and polyA tail) and thus the primary microRNA (pri-miRNA) arise. Then, the typical stem-loop structure is formed by complementary base pairing and cleaved at the dicing bodies (consisting of several proteins including DCL1, HYL1, SE, TIGH, and DDL) resulting in miRNA duplex formation which can be later 2′-O-methylated (ensured by the HEN1 protein). Guide miRNA is incorporated into the RISC consisting of several proteins, and transported into the cytoplasm, where mRNA target recognition and cleavage can take place while the passenger miRNA is released away. Proteins from the Argonaute family (AGOs) can modify the stability of the miRNAs and also affect the interaction with target mRNAs. This figure was created using BioRender (https://biorender.com/; accessed on 20 June 2022).
miRNAs interact with their target mRNAs mostly at their 3′ UTRs, but interactions occurring in the 5′ UTRs or coding regions were documented as well [,]. RISC is directed to the complementary mRNA transcript, whereby the Watson–Crick base-pairing aligns guide miRNA and target mRNA transcript, and depending on the central miRNA region complementarity, mRNA is cleaved (usually when there is perfect base-pair complementarity), or translation repression occurs (central miRNA region is not completely complementary to mRNA) []. Moreover, in the case the target mRNA is cleaved, so-called phased secondary small interfering RNAs (phasiRNAs) can arise []. phasiRNAs are 21 or 24-nucleotide-long siRNAs having important roles in plant stress responses [], development [], and reproduction [].
Similar to the other genes, miRNA transcription is precisely fine-tuned. This is assured mainly by transcription factors binding [] and methylation status of DNA [], both heavily influenced by endogenous and exogenous stimuli. In 2018, protein WHIRLY1 was found to be involved in increased levels of nuclear miRNAs in high-light conditions in barley. It was therefore proposed that WHIRLY1 can bind to RNA and it might be a general factor influencing the biogenesis and/or stability of various miRNAs [].
An additional level of miRNA complexity is their dynamic stability [,]. It was documented that the processes such as 3′-end modifications and interaction with Argonaute proteins (AGOs) can both reduce and increase the stability of miRNAs depending on the actual needs of the plant. For example, AGO1 from Arabidopsis thaliana was proposed to stabilize miRNAs, and miRNA–mRNA target interaction [].
Besides post-transcriptional gene silencing (PTGS), miRNAs can regulate plant genes via RNA-induced methylation of DNA [,]. Such a process was in detail described in the Arabidopsis thaliana, where miRNAs (miR165, miR166) regulate the methylation status of PHB and PHV genes [], and are responsible for the determination of the abaxial and adaxial leaf side. Similarly, the miRNA-induced gene methylation was described even in the Oryza sativa where the miR1873 ensures the methylation of its own gene []. To make our understanding of miRNAs-based regulation of gene expression more challenging, the stimulative effect of miRNAs on gene expression was observed and documented as well [].
Last but not least, in 2015 it was proposed that plant pri-miRNAs are capable of encoding small functional peptides [,] described as miPEPs. The best-characterized miPEPs (miPEP171d, miPEP172c, and miPEP858a) were found in plant species including Arabidopsis thaliana (miPEP165a [], miPEP858 []), Medicago truncatula (miPEP171b []), Glycine max (miPEP172c []), and Vitis vinifera (miPEP171d1 []). The mechanism of miPEPs molecular function is still largely unclear, but generally, miPEPs positively affect the accumulation of their associated miRNAs []. It is also likely that many of miPEPs will be species-specific [].
3. miRNAs in Barley Physiology and Stress Responses
miRNAs in plants are important regulators of various physiological processes including shoot apical meristem development [], leaf growth [], flower formation [], seed production [], and root expansion []. It was found that miRNA171 in barley is responsible for the regulation of shoot meristem development through three independent pathways, i.e., firstly through the down-regulation of SCARECROW-LIKE (SCL) transcription factors, secondly via up-regulation of miRNA156 and repressing vegetative phase transitions (a possibly monocotyledon-specific mechanism), and thirdly by repressing expression of TRD and HvPLA1 genes []. Additionally, flower development in grasses including barley is tightly regulated by miRNAs. It was found that miRNA159, miRNA171, miRNA172, and miRNA396 regulate the expression of floral organ identity genes in barley, rice, and maize []. In barley, cleistogamous flowering (i.e., shedding its pollen before opening) arises from the suppression of the AP2 transcription factor via miR172, originally thought to be a result of target mRNA cleavage [], but later it was proved that miR172-mediated AP2 regulation occurs at the translational level []. It is also known that the expression of barley miR393 is active in the developmental period, and its misexpression affects seedling growth and stomatal density []. In 2018, it was found that miR160 in barley simultaneously targets class II ARF members which are functionally involved in developmental stages by regulating the auxin-mediated genes []. Figure 2 illustratively depicts some of the most known barley miRNAs (and their targets) that play important roles in developmental processes.
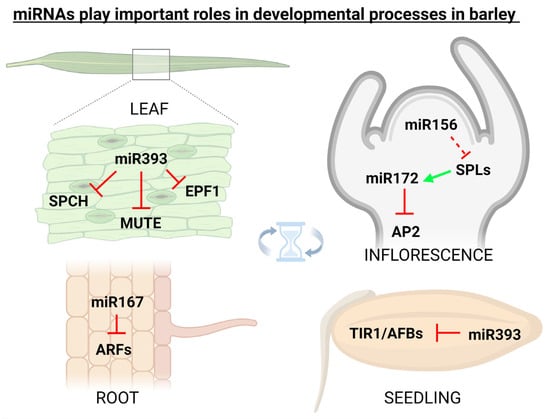
Figure 2.
miRNAs play important roles also in the developmental processes. In spring barley (Hordeum vulgare), specific miRNAs were linked with the targets involved in the regulation of flowering, root development, seed germination, and also with stomata development. Inhibition is indicated by the red ┴ mark, while positive effect by the green arrow. This figure was created using BioRender (https://biorender.com/; accessed on 20 June 2022).
Besides the non-stress conditions, miRNAs play key roles in gene expression regulation in response to a variety of abiotic stimuli, including several stress responses. In plants, their involvement in many abiotic stress responses including heat stress responses, low-temperature responses, drought exposure responses, carbon dioxide responses, light stress responses, or gamma radiation responses was reported [,,,,]. Specifically in barley, miRNAs responsive to salinity stress [,,,], drought [,,,,], nitrogen [], boron [], phosphorus [,], aluminum [,,], cadmium [], cold deacclimation [], heat stress [], and possibly to light [] were identified till date. A chronological summary of the most impactful miRNA studies in barley (starting from 2010) can be found below in Table 1.

Table 1.
Chronological summary of studies dealing with miRNAs in barley species and most important results obtained.
From the above-mentioned studies, it is evident that barley miRNAs play a complex role in responses to various abiotic and biotic stresses or stimuli, which is schematically depicted in Figure 3.
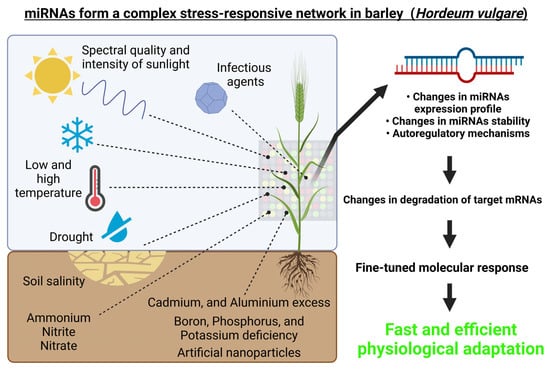
Figure 3.
miRNAs form a complex regulatory network in barley (Hordeum vulgare). Environmental cues, both abiotic (i.e., spectral quality and intensity of the incident light, growth temperature, drought, high salinity, heavy metals exposure, etc.) and biotic (for example pathogens) can affect the expression of miRNAs and thus also their target genes. This figure was created using BioRender (https://biorender.com/; accessed on 20 June 2022).
4. Target Transcripts of Barley miRNAs
Several web-based tools, resources, and databases related to small RNAs comprising miRNAs in plants exist []. To browse miRNAs identified in barley to date, mainly PNRD [], PmiREN [], miRBase [], Plant small RNA genes [], or the integrative miRNEST database [] can be used. As these databases use slightly different methods of required miRNAs evidence or data sources, overall counts of miRNAs deposited here differ. Total counts of barley miRNAs in each of the databases together with direct links and other useful information are listed in Table 2.
Table 2.
User-friendly online databases comprising barley miRNAs.
According to TarDB: A miRNA Target Database in Plants [] (http://www.biosequencing.cn/TarDB/browse.html, accessed 20 June 2022), there are currently 20 conserved miRNAs in barley (Table 3). It is worth mentioning that TarDB uses relatively strict parameters to identify high-confidence plant miRNAs and their targets based on cross-species conservation filter, degradome, and sRNA-seq data, so Table 3 below is rather illustrative than exhaustive. It is supposed that the overall number of functional miRNAs in barley is much higher, according to a study published in November 2021 [], a total of 156 miRNAs including 35 known and 121 novel miRNAs experimentally identified in Tibetan hull-less barleys, targeting over 1200 genes (nonetheless it was done by computational prediction, only selected targets were also in vitro verified using RLM-5′ RACE method) [].
Table 3.
High-confidence miRNA targets in Hordeum vulgare together with their experimentally verified or supposed biological functions in higher plants.
Many barley miRNAs are targeting mRNA transcripts encoding transcription factors. This is maybe not too surprising, as it was previously known that most of the plant miRNA targets are transcription factors that regulate plant growth and development []. In Figure 4, known barley miRNA targets from TarDB [] are grouped according to their gene ontologies (GOs), separately for ‘all conserved miRNA targets’ (dataset containing 92 mRNAs) and ‘degradome-supported miRNA targets’ (dataset containing 37 mRNAs, 15 of them are common with the first subset of ‘all conserved miRNA targets’). GO terms for these datasets were acquired using PLAZA Workbench [,]. It can be seen that most miRNA targets participate in diverse biological processes, comprising metabolic, developmental, regulatory, and reproductive processes. This fact may be in good aggreement with general observations and knowledge from miRNA studies not only in barley plants. From the point of molecular functions view, barley miRNA targets are employed mainly in binding processes (e.g., organic cyclic compound binding, protein binding, nucleic acid binding, etc.) and some targets possess catalytic activity (Figure 4). It is worth noting that the above-mentioned GOs are more general than specific, and they deserve more detailed analysis in the future. Moreover, it may be interesting that many miRNA targets (once translated into proteins) can bind DNA and theoretically act as transcriptional activators or repressors influencing the expression of their superior miRNA genes, thus forming another regulatory layer (or feedback loop) []. This issue could certainly serve as a potential theme for further research in the field of plant development and stress responses.
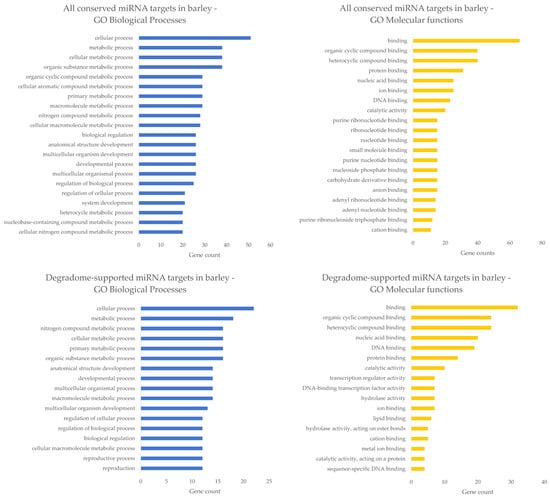
Figure 4.
Biological processes and Molecular functions of miRNA targets in barley. In the upper half of the image, the most abundant GOs of all conserved miRNA targets in barley are shown. In the lower half of the image, the most abundant GOs of degradome-supported miRNA targets are depicted. Blue bar plots stand for biological processes, whereas the orange ones correspond to molecular functions.
It is essential to bear in mind that a particular miRNA can interact with many different mRNA molecules [], and that particular miRNA targets can be relatively quickly acquired through plant evolution []. A good example is miR168a from sweet orange (Citrus sinensis L. Osbeck), where besides its original target (AGO1 mRNA) it gained a novel target, CUC2 mRNA []. Another specific case was observed in rice (Oryza sativa japonica cv. Nipponbare), where miR159 triggers MAP kinase 8 mRNA, in addition to its original target (MYB mRNA) []. Interestingly, miRNA activity can be regulated by bait in the form of long non-coding RNAs (lncRNAs), where such lncRNAs mimic the target mRNAs and sequester specific miRNAs (preventing them from interacting with their mRNA targets)—this phenomenon is usually described as (mi)RNA decoy [] or Target Mimics []. In barley, there is a study from 2020 where authors identified about 8000 lncRNAs and found a total of 32 endogenous target mimics that may potentially decoy 18 different miRNAs [].
As an illustrative example of miRNAs targets diversity, we depicted all computationally predicted mRNA targets of single barley miRNA, particularly miR5049c (Figure 5). According to TarDB [], this miRNA has the potential to target 17 different mRNAs originating from various genes across the whole barley genome (Chromosomes 1 to 7). The molecular and biological functions of proteins encoded by these mRNAs are also very diverse, and some of them participate in response to external stimuli, e.g., HSP20-like chaperones superfamily protein (by homology) []. The fact that one miRNA can bind multiple mRNA targets is relatively well-known for many years [,]. Obviously, at the same time, a single miRNA molecule can bind only a single mRNA target, therefore one may imagine that the relative accessibility of particular mRNA to a particular miRNA determines the proportion of specific mRNA-miRNA interactions. The cell usually produces only a fraction of all possible mRNAs, and therefore such a mechanism of regulation would seem efficient.
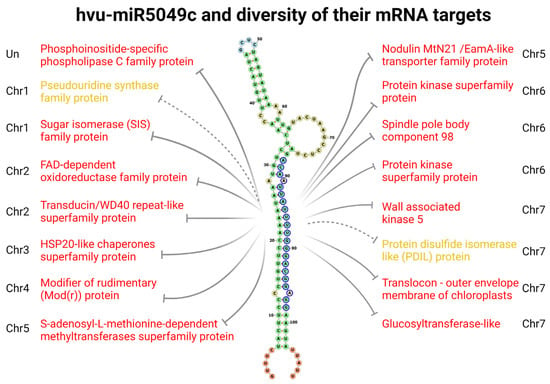
Figure 5.
Barley pri-miR5049c structure together with miR5049 putative mRNA targets. pri-miRNA structure was computed via RNAfold web server [] and visualized in the form of a Forna diagram []. Nucleotides in blue circles correspond to the mature 21nt-long miRNA region. Grey lines depict inhibition of specific mRNA targets (if the line is full and the description is in red, mRNA cleavage was predicted, according to TarDB: “Cleavage is predicted if miRNA 5′ positions 9–11 have the perfect match”). Chromosome numbers correspond to the location of genes encoding particular mRNAs, Un stands for Unplaced locus.
5. Conclusions and Future Directions
This review gives a basic overview of a rapidly growing amount of miRNA studies in barley (Hordeum vulgare). From what we know, it is clear that miRNAs play an important role in many developmental processes as well as in a variety of stress-induced molecular and biological responses. It is likely that more and more putative miRNAs will be discovered in barley, and many of them will be linked to abiotic or biotic stresses, including drought, cold, high temperatures, high salinity, micronutrient excess or deficiency in the soil, spectral quality of incident light, or infectious agents. Identification of plant miRNA targets on a large scale has traditionally been made mainly by bioinformatic approaches [,,,,,]. On the other hand, experimental validation is needed to verify predicted mRNA targets—historically, this has been done using laboratory-intensive in vitro methods like the 5′ RACE assay [], but nowadays, the high-throughput degradome sequencing technique can be employed to validate (at least partially) predicted miRNA targets. Nonetheless, four criteria (according to a nice review by Giulia Riolo et al. []) should ideally be fulfilled when validating novel miRNAs:
- (a)
- Showing co-expression of miRNA and target mRNA in vivo;
- (b)
- Proving interaction between miRNA and a specific site within target mRNA;
- (c)
- Demonstrating miRNA-mediated effects on target protein expression;
- (d)
- Demonstrating miRNA effects on biological function.
What is quite difficult for our complex understanding of miRNA mechanisms is that even different genotypes/cultivars of barley tend to express unique miRNA patterns. This may point to rapid miRNA evolution allowing gene expression fine-tuning in a dynamically changing environment and agriculture. In addition, different plant tissues may express a different ‘miRNome’ in response to various stress signals [].
Several studies have discussed the possibility of miRNA-based technology to improve plant resistance to abiotic factors [,]. In 2017, Jannatul Ferdous et al. published a study where the drought-inducible expression of miR827 enhanced drought tolerance in transgenic barley []. In maize, the knock-down of miR166 using short tandem target mimics technology resulted in enhanced abiotic stress resistance, abscisic acid level elevation, and indole acetic acid level reduction []. As miR166 is conserved also in barley, it would be interesting to identify whether its knock-down would have similar effects. Another promising possibility offers CRISPR/Cas technology already utilized for miRNA gene editing in rice [,] and Arabidopsis thaliana [], further reviewed in []. Finally, there is an increasing effort to use exogenous/artificially made miRNAs (or siRNAs) in modern plant protection and improvement, and such RNA interference technology is usually considered GMO-free []. Among the various options, chitosan nanoparticles bearing miRNAs seem to be particularly attractive [].
Barley miRNAs could also be efficiently used as molecular markers. In 2020, researchers proposed selected miRNAs as a tool to monitor the barley response to soil compaction [].
Below, several outstanding questions are summarized:
- (1)
- Are some of the barley miRNAs tissue/developmental, or stage-specific? Are we able to catalog it in some integrative and user-friendly way? For this purpose, it would be beneficial to have something like a barley miRNA atlas (similar to PmiRExAt, where wheat, rice, maize, and Arabidopsis miRNAs in multiple tissues and developmental stages can be found) [].
- (2)
- Which barley miRNAs have the potential to become a useful stress biomarker? In other words, do some stress-specific miRNAs exist?
- (3)
- Is barley miRNome rather complete, or not? Compared to rice, wheat, and Arabidopsis, the total number of known barley miRNAs is still lack behind, and bona fide many discoveries waiting for us!
To better depict the above-mentioned perspectives in barley miRNAs research, we have created a diagram where particular future aims are divided into two categories, i.e., work to be done either using dry-lab or wet-lab methods, together with possible future applications (Figure 6).
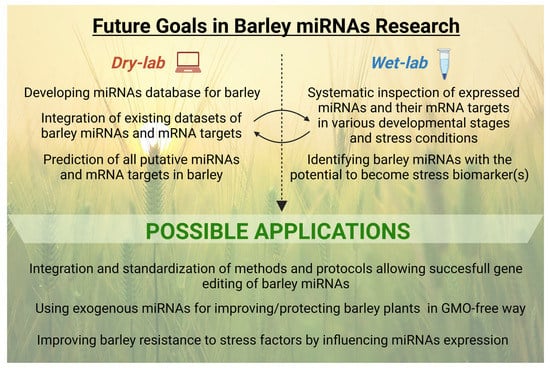
Figure 6.
Future goals in miRNAs research in barley, divided into dry-lab and wet-lab categories, and possible future applications.
All in all, even though a lot is known about miRNAs in barley, much remains to be resolved. Aristotle said “the more you know, the more you realize you don’t know”, and complex miRNAs problematics in barley (and generally in plants) could definitely fit this quote.
Author Contributions
Conceptualization, A.V. and M.B.; resources, M.B. and A.V.; writing—original draft preparation, A.V. and M.B.; writing—review and editing, V.Š., P.P. and J.Č.; visualization, A.V. and M.B.; supervision, J.Č.; project administration, J.Č.; funding acquisition, J.Č. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Czech Science Foundation (GACR 21-18532S to V.Š. and A.V.), the University of Ostrava (SGS11/PřF/2022 to A.V.; SGS10/PřF/2022 to P.P.). Participation of V.Š. was also supported by the Ministry of Education, Youth and Sports of the Czech Republic, project “SustES—Adaptation strategies for sustainable ecosystem services and food security under adverse environmental conditions” (CZ.02.1.01/0.0/0.0/16_019/0000797). J.Č., P.P. and M.B. were supported by the National Agency for Agricultural Research (NAZV) of the Czech Republic grant no. QK1810391 “Utilization of genomic and transcriptomic approaches to create genetic resources and breeding materials of poppy with specific traits”.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study.
References
- Lam, J.K.W.; Chow, M.Y.T.; Zhang, Y.; Leung, S.W.S. SiRNA versus MiRNA as Therapeutics for Gene Silencing. Mol. Ther. Nucleic Acids 2015, 4, e252. [Google Scholar] [CrossRef]
- Wang, J.; Mei, J.; Ren, G. Plant MicroRNAs: Biogenesis, Homeostasis, and Degradation. Front. Plant Sci. 2019, 10, 360. [Google Scholar] [CrossRef] [PubMed]
- Sen, G.L.; Blau, H.M. A Brief History of RNAi: The Silence of the Genes. FASEB J. 2006, 20, 1293–1299. [Google Scholar] [CrossRef]
- Napoli, C.; Lemieux, C.; Jorgensen, R. Introduction of a Chimeric Chalcone Synthase Gene into Petunia Results in Reversible Co-Suppression of Homologous Genes in Trans. Plant Cell 1990, 2, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Romano, N.; Macino, G. Quelling: Transient Inactivation of Gene Expression in Neurospora Crassa by Transformation with Homologous Sequences. Mol. Microbiol. 1992, 6, 3343–3353. [Google Scholar] [CrossRef]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans Heterochronic Gene Lin-4 Encodes Small RNAs with Antisense Complementarity to Lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef] [PubMed]
- Ambros, V.; Bartel, B.; Bartel, D.P.; Burge, C.B.; Carrington, J.C.; Chen, X.; Dreyfuss, G.; Eddy, S.R.; Griffiths-Jones, S.; Marshall, M.; et al. A Uniform System for MicroRNA Annotation. RNA 2003, 9, 277–279. [Google Scholar] [CrossRef] [PubMed]
- Gebert, L.F.R.; MacRae, I.J. Regulation of MicroRNA Function in Animals. Nat. Rev. Mol. Cell Biol. 2019, 20, 21–37. [Google Scholar] [CrossRef] [PubMed]
- Garcia, D. A MiRacle in Plant Development: Role of MicroRNAs in Cell Differentiation and Patterning. Semin. Cell Dev. Biol. 2008, 19, 586–595. [Google Scholar] [CrossRef]
- Millar, A.A. The Function of MiRNAs in Plants. Plants 2020, 9, 198. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, T.; Zhang, F.; Zhang, Y.; Liang, Y. RNA Interference: A Natural Immune System of Plants to Counteract Biotic Stressors. Cells 2019, 8, 38. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Galili, G. Tuning the Orchestra: MiRNAs in Plant Immunity. Trends Plant Sci. 2019, 24, 189–191. [Google Scholar] [CrossRef] [PubMed]
- Cui, C.; Wang, J.-J.; Zhao, J.-H.; Fang, Y.-Y.; He, X.-F.; Guo, H.-S.; Duan, C.-G. A Brassica MiRNA Regulates Plant Growth and Immunity through Distinct Modes of Action. Mol. Plant 2020, 13, 231–245. [Google Scholar] [CrossRef]
- Mengistu, A.A.; Tenkegna, T.A. The Role of MiRNA in Plant–Virus Interaction: A Review. Mol. Biol. Rep. 2021, 48, 2853–2861. [Google Scholar] [CrossRef]
- Pagano, L.; Rossi, R.; Paesano, L.; Marmiroli, N.; Marmiroli, M. MiRNA Regulation and Stress Adaptation in Plants. Environ. Exp. Bot. 2021, 184, 104369. [Google Scholar] [CrossRef]
- Nevo, E. Evolution of Wild Barley and Barley Improvement. In Advance in Barley Sciences; Springer: Dordrecht, The Netherlands, 2013; pp. 1–23. [Google Scholar]
- Pourkheirandish, M.; Komatsuda, T. The Importance of Barley Genetics and Domestication in a Global Perspective. Ann. Bot. 2007, 100, 999–1008. [Google Scholar] [CrossRef]
- Ullrich, S.E. Barley: Production, Improvement, and Uses; John Wiley & Sons: New York, NY, USA, 2010. [Google Scholar]
- Tosh, S.M.; Bordenave, N. Emerging Science on Benefits of Whole Grain Oat and Barley and Their Soluble Dietary Fibers for Heart Health, Glycemic Response, and Gut Microbiota. Nutr. Rev. 2020, 78, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Lahouar, L.; El-Bok, S.; Achour, L. Therapeutic Potential of Young Green Barley Leaves in Prevention and Treatment of Chronic Diseases: An Overview. Am. J. Chin. Med. 2015, 43, 1311–1329. [Google Scholar] [CrossRef]
- Pech, R.; Volná, A.; Hunt, L.; Bartas, M.; Červeň, J.; Pečinka, P.; Špunda, V.; Nezval, J. Regulation of Phenolic Compound Production by Light Varying in Spectral Quality and Total Irradiance. Int. J. Mol. Sci. 2022, 23, 6533. [Google Scholar] [CrossRef]
- Harwood, W.A. An Introduction to Barley: The Crop and the Model. In Barley; Humana Press: New York, NY, UAS, 2019; pp. 1–5. [Google Scholar]
- Sato, K. History and Future Perspectives of Barley Genomics. DNA Res. 2020, 27, dsaa023. [Google Scholar] [CrossRef] [PubMed]
- Saski, C.; Lee, S.-B.; Fjellheim, S.; Guda, C.; Jansen, R.K.; Luo, H.; Tomkins, J.; Rognli, O.A.; Daniell, H.; Clarke, J.L. Complete Chloroplast Genome Sequences of Hordeum vulgare, Sorghum bicolor and Agrostis stolonifera, and Comparative Analyses with Other Grass Genomes. Theor. Appl. Genet. 2007, 115, 571–590. [Google Scholar] [CrossRef]
- Schoch, C.L.; Ciufo, S.; Domrachev, M.; Hotton, C.L.; Kannan, S.; Khovanskaya, R.; Leipe, D.; Mcveigh, R.; O’Neill, K.; Robbertse, B. NCBI Taxonomy: A Comprehensive Update on Curation, Resources and Tools. Database 2020, 2020, baaa062. [Google Scholar] [CrossRef] [PubMed]
- Sato, F.; Tsuchiya, S.; Meltzer, S.J.; Shimizu, K. MicroRNAs and Epigenetics. FEBS J. 2011, 278, 1598–1609. [Google Scholar] [CrossRef] [PubMed]
- Shapulatov, U.; van Hoogdalem, M.; Schreuder, M.; Bouwmeester, H.; Abdurakhmonov, I.Y.; van der Krol, A.R. Functional Intron-Derived MiRNAs and Host-Gene Expression in Plants. Plant Methods 2018, 14, 83. [Google Scholar] [CrossRef]
- Wu, X.; Hornyik, C.; Bayer, M.; Marshall, D.; Waugh, R.; Zhang, R. In Silico Identification and Characterization of Conserved Plant MicroRNAs in Barley. Open Life Sci. 2014, 9, 841–852. [Google Scholar] [CrossRef][Green Version]
- Baldrich, P.; Hsing, Y.-I.C.; San Segundo, B. Genome-Wide Analysis of Polycistronic MicroRNAs in Cultivated and Wild Rice. Genome. Biol. Evol. 2016, 8, 1104–1114. [Google Scholar] [CrossRef] [PubMed]
- Zou, Q.; Mao, Y.; Hu, L.; Wu, Y.; Ji, Z. MiRClassify: An Advanced Web Server for MiRNA Family Classification and Annotation. Comput. Biol. Med. 2014, 45, 157–160. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Dou, Y.; Li, S.; Ren, G.; Chevalier, D.; Zhang, C.; Yu, B. DAWDLE Interacts with DICER-LIKE Proteins to Mediate Small RNA Biogenesis. Plant Physiol. 2018, 177, 1142–1151. [Google Scholar] [CrossRef]
- Zhang, L.; Xiang, Y.; Chen, S.; Shi, M.; Jiang, X.; He, Z.; Gao, S. Mechanisms of MicroRNA Biogenesis and Stability Control in Plants. Front. Plant Sci. 2022, 13, 844149. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Yang, Z.; Li, J.; Minakhina, S.; Yang, M.; Padgett, R.W.; Steward, R.; Chen, X. Methylation as a Crucial Step in Plant MicroRNA Biogenesis. Science 2005, 307, 932–935. [Google Scholar] [CrossRef]
- Djami-Tchatchou, A.T.; Sanan-Mishra, N.; Ntushelo, K.; Dubery, I.A. Functional Roles of MicroRNAs in Agronomically Important Plants—Potential as Targets for Crop Improvement and Protection. Front. Plant Sci. 2017, 8, 378. [Google Scholar] [CrossRef]
- Li, M.; Yu, B. Recent Advances in the Regulation of Plant MiRNA Biogenesis. RNA Biol. 2021, 18, 2087–2096. [Google Scholar] [CrossRef] [PubMed]
- Medley, J.C.; Panzade, G.; Zinovyeva, A.Y. MicroRNA Strand Selection: Unwinding the Rules. Wiley Interdiscip. Rev. RNA 2021, 12, e1627. [Google Scholar] [CrossRef] [PubMed]
- Meijer, H.A.; Smith, E.M.; Bushell, M. Regulation of MiRNA Strand Selection: Follow the Leader? Biochem. Soc. Trans. 2014, 42, 1135–1140. [Google Scholar] [CrossRef]
- Vimalraj, S.; Selvamurugan, N. MicroRNAs: Synthesis, Gene Regulation and Osteoblast Differentiation. Curr. Issues Mol. Biol. 2013, 15, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Forman, J.J.; Coller, H.A. The Code within the Code: MicroRNAs Target Coding Regions. Cell Cycle 2010, 9, 1533–1541. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Wang, H. Translational Inhibition by MicroRNAs in Plants. Prog. Mol. Subcell. Biol. 2010, 50, 41–57. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Teng, C.; Xia, R.; Meyers, B.C. PhasiRNAs in Plants: Their Biogenesis, Genic Sources, and Roles in Stress Responses, Development, and Reproduction. Plant Cell 2020, 32, 3059–3080. [Google Scholar] [CrossRef] [PubMed]
- Araki, S.; Le, N.T.; Koizumi, K.; Villar-Briones, A.; Nonomura, K.-I.; Endo, M.; Inoue, H.; Saze, H.; Komiya, R. MiR2118-Dependent U-Rich PhasiRNA Production in Rice Anther Wall Development. Nat. Commun. 2020, 11, 3115. [Google Scholar] [CrossRef]
- Xia, R.; Chen, C.; Pokhrel, S.; Ma, W.; Huang, K.; Patel, P.; Wang, F.; Xu, J.; Liu, Z.; Li, J.; et al. 24-Nt Reproductive PhasiRNAs Are Broadly Present in Angiosperms. Nat. Commun. 2019, 10, 627. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Khanna, K.; Ruan, S. Expression of MicroRNAs and Its Regulation in Plants. Semin. Cell Dev. Biol. 2010, 21, 790–797. [Google Scholar] [CrossRef] [PubMed]
- Chhabra, R. MiRNA and Methylation: A Multifaceted Liaison. ChemBioChem 2015, 16, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Świda-Barteczka, A.; Krieger-Liszkay, A.; Bilger, W.; Voigt, U.; Hensel, G.; Szweykowska-Kulinska, Z.; Krupinska, K. The Plastid-Nucleus Located DNA/RNA Binding Protein WHIRLY1 Regulates MicroRNA-Levels during Stress in Barley (Hordeum vulgare L.). RNA Biol. 2018, 15, 886–891. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Mo, B.; Chen, X. Mechanisms That Impact MicroRNA Stability in Plants. RNA Biol. 2012, 9, 1218–1223. [Google Scholar] [CrossRef]
- Gallego-Bartolomé, J. DNA Methylation in Plants: Mechanisms and Tools for Targeted Manipulation. New Phytol. 2020, 227, 38–44. [Google Scholar] [CrossRef]
- Wu, L.; Zhou, H.; Zhang, Q.; Zhang, J.; Ni, F.; Liu, C.; Qi, Y. DNA Methylation Mediated by a MicroRNA Pathway. Mol. Cell 2010, 38, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Bao, N.; Lye, K.-W.; Barton, M.K. MicroRNA Binding Sites in Arabidopsis Class III HD-ZIP MRNAs Are Required for Methylation of the Template Chromosome. Dev. Cell 2004, 7, 653–662. [Google Scholar] [CrossRef] [PubMed]
- Vasudevan, S. Posttranscriptional Upregulation by MicroRNAs. WIREs RNA 2012, 3, 311–330. [Google Scholar] [CrossRef] [PubMed]
- Lauressergues, D.; Couzigou, J.-M.; Clemente, H.S.; Martinez, Y.; Dunand, C.; Bécard, G.; Combier, J.-P. Primary Transcripts of MicroRNAs Encode Regulatory Peptides. Nature 2015, 520, 90–93. [Google Scholar] [CrossRef] [PubMed]
- Prasad, A.; Sharma, N.; Prasad, M. Noncoding but Coding: Pri-MiRNA into the Action. Trends Plant Sci. 2021, 26, 204–206. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Badola, P.K.; Bhatia, C.; Sharma, D.; Trivedi, P.K. Primary Transcript of MiR858 Encodes Regulatory Peptide and Controls Flavonoid Biosynthesis and Development in Arabidopsis. Nat. Plants 2020, 6, 1262–1274. [Google Scholar] [CrossRef]
- Couzigou, J.-M.; André, O.; Guillotin, B.; Alexandre, M.; Combier, J.-P. Use of MicroRNA-Encoded Peptide MiPEP172c to Stimulate Nodulation in Soybean. New Phytol. 2016, 211, 379–381. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Deng, B.; Gao, J.; Zhao, Z.; Chen, Z.; Song, S.; Wang, L.; Zhao, L.; Xu, W.; Zhang, C.; et al. A MiRNA-Encoded Small Peptide, Vvi-MiPEP171d1, Regulates Adventitious Root Formation. Plant Physiol. 2020, 183, 656–670. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.E.; Zhao, Y.-T.; Wang, X.-J.; Croft, L.; Wang, Z.-H.; Haerizadeh, F.; Mattick, J.S.; Singh, M.B.; Carroll, B.J.; Bhalla, P.L. MicroRNAs in the Shoot Apical Meristem of Soybean. J. Exp. Bot. 2011, 62, 2495–2506. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, A.; Kumar, A.; Kaur, H.; Kaur, N. MiRNA: The Taskmaster of Plant World. Biologia 2021, 76, 1551–1567. [Google Scholar] [CrossRef]
- Waheed, S.; Zeng, L. The Critical Role of MiRNAs in Regulation of Flowering Time and Flower Development. Genes 2020, 11, 319. [Google Scholar] [CrossRef] [PubMed]
- Curaba, J.; Spriggs, A.; Taylor, J.; Li, Z.; Helliwell, C. MiRNA Regulation in the Early Development of Barley Seed. BMC Plant Biol. 2012, 12, 120. [Google Scholar] [CrossRef]
- Curaba, J.; Talbot, M.; Li, Z.; Helliwell, C. Over-Expression of MicroRNA171 Affects Phase Transitions and Floral Meristem Determinancy in Barley. BMC Plant Biol. 2013, 13, 6. [Google Scholar] [CrossRef]
- Smoczynska, A.; Szweykowska-Kulinska, Z. MicroRNA-Mediated Regulation of Flower Development in Grasses. Acta Biochim. Pol. 2016, 63, 687–692. [Google Scholar] [CrossRef] [PubMed]
- Nair, S.K.; Wang, N.; Turuspekov, Y.; Pourkheirandish, M.; Sinsuwongwat, S.; Chen, G.; Sameri, M.; Tagiri, A.; Honda, I.; Watanabe, Y.; et al. Cleistogamous Flowering in Barley Arises from the Suppression of MicroRNA-Guided HvAP2 MRNA Cleavage. Proc. Natl. Acad. Sci. USA 2010, 107, 490–495. [Google Scholar] [CrossRef] [PubMed]
- Anwar, N.; Ohta, M.; Yazawa, T.; Sato, Y.; Li, C.; Tagiri, A.; Sakuma, M.; Nussbaumer, T.; Bregitzer, P.; Pourkheirandish, M.; et al. MiR172 Downregulates the Translation of Cleistogamy 1 in Barley. Ann. Bot. 2018, 122, 251–265. [Google Scholar] [CrossRef]
- Yuan, W.; Suo, J.; Shi, B.; Zhou, C.; Bai, B.; Bian, H.; Zhu, M.; Han, N. The Barley MiR393 Has Multiple Roles in Regulation of Seedling Growth, Stomatal Density, and Drought Stress Tolerance. Plant Physiol. Biochem. 2019, 142, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Tombuloglu, H. Genome-Wide Analysis of the Auxin Response Factors (ARF) Gene Family in Barley (Hordeum vulgare L.). J. Plant Biochem. Biotechnol. 2019, 28, 14–24. [Google Scholar] [CrossRef]
- Shriram, V.; Kumar, V.; Devarumath, R.M.; Khare, T.S.; Wani, S.H. MicroRNAs as Potential Targets for Abiotic Stress Tolerance in Plants. Front. Plant Sci. 2016, 7, 817. [Google Scholar] [CrossRef]
- Barczak-Brzyżek, A.; Brzyżek, G.; Koter, M.; Siedlecka, E.; Gawroński, P.; Filipecki, M. Plastid Retrograde Regulation of MiRNA Expression in Response to Light Stress. BMC Plant Biol. 2022, 22, 150. [Google Scholar] [CrossRef]
- Subburaj, S.; Ha, H.-J.; Jin, Y.-T.; Jeon, Y.; Tu, L.; Kim, J.-B.; Kang, S.-Y.; Lee, G.-J. Identification of γ-Radiation-Responsive MicroRNAs and Their Target Genes in Tradescantia (BNL Clone 4430). J. Plant Biol. 2017, 60, 116–128. [Google Scholar] [CrossRef]
- Visentin, I.; Pagliarani, C.; Deva, E.; Caracci, A.; Turečková, V.; Novák, O.; Lovisolo, C.; Schubert, A.; Cardinale, F. A Novel Strigolactone-MiR156 Module Controls Stomatal Behaviour during Drought Recovery. Plant Cell Environ. 2020, 43, 1613–1624. [Google Scholar] [CrossRef] [PubMed]
- Saminathan, T.; Alvarado, A.; Lopez, C.; Shinde, S.; Gajanayake, B.; Abburi, V.L.; Vajja, V.G.; Jagadeeswaran, G.; Raja Reddy, K.; Nimmakayala, P.; et al. Elevated Carbon Dioxide and Drought Modulate Physiology and Storage-Root Development in Sweet Potato by Regulating MicroRNAs. Funct. Integr. Genom. 2019, 19, 171–190. [Google Scholar] [CrossRef] [PubMed]
- Deng, P.; Wang, L.; Cui, L.; Feng, K.; Liu, F.; Du, X.; Tong, W.; Nie, X.; Ji, W.; Weining, S. Global Identification of MicroRNAs and Their Targets in Barley under Salinity Stress. PLoS ONE 2015, 10, e0137990. [Google Scholar] [CrossRef] [PubMed]
- Kuang, L.; Yu, J.; Shen, Q.; Fu, L.; Wu, L. Identification of MicroRNAs Responding to Aluminium, Cadmium and Salt Stresses in Barley Roots. Plants 2021, 10, 2754. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Sun, G. Micro RNA s Contribute to Enhanced Salt Adaptation of the Autopolyploid Hordeum bulbosum Compared with Its Diploid Ancestor. Plant J. 2017, 91, 57–69. [Google Scholar] [CrossRef]
- Kuang, L.; Shen, Q.; Wu, L.; Yu, J.; Fu, L.; Wu, D.; Zhang, G. Identification of MicroRNAs Responding to Salt Stress in Barley by High-Throughput Sequencing and Degradome Analysis. Environ. Exp. Bot. 2019, 160, 59–70. [Google Scholar] [CrossRef]
- Smoczynska, A.; Pacak, A.M.; Nuc, P.; Swida-Barteczka, A.; Kruszka, K.; Karlowski, W.M.; Jarmolowski, A.; Szweykowska-Kulinska, Z. A Functional Network of Novel Barley MicroRNAs and Their Targets in Response to Drought. Genes 2020, 11, 488. [Google Scholar] [CrossRef] [PubMed]
- Ferdous, J.; Sanchez-Ferrero, J.C.; Langridge, P.; Milne, L.; Chowdhury, J.; Brien, C.; Tricker, P.J. Differential Expression of MicroRNAs and Potential Targets under Drought Stress in Barley. Plant Cell Environ. 2017, 40, 11–24. [Google Scholar] [CrossRef]
- Kantar, M.; Unver, T.; Budak, H. Regulation of Barley MiRNAs upon Dehydration Stress Correlated with Target Gene Expression. Funct. Integr. Genom. 2010, 10, 493–507. [Google Scholar] [CrossRef]
- Hackenberg, M.; Gustafson, P.; Langridge, P.; Shi, B.-J. Differential Expression of MicroRNAs and Other Small RNAs in Barley between Water and Drought Conditions. Plant Biotechnol. J. 2015, 13, 2–13. [Google Scholar] [CrossRef] [PubMed]
- Qiu, C.-W.; Liu, L.; Feng, X.; Hao, P.-F.; He, X.; Cao, F.; Wu, F. Genome-Wide Identification and Characterization of Drought Stress Responsive MicroRNAs in Tibetan Wild Barley. Int. J. Mol. Sci. 2020, 21, 2795. [Google Scholar] [CrossRef]
- Grabowska, A.; Smoczynska, A.; Bielewicz, D.; Pacak, A.; Jarmolowski, A.; Szweykowska-Kulinska, Z. Barley MicroRNAs as Metabolic Sensors for Soil Nitrogen Availability. Plant Sci. 2020, 299, 110608. [Google Scholar] [CrossRef] [PubMed]
- Ozhuner, E.; Eldem, V.; Ipek, A.; Okay, S.; Sakcali, S.; Zhang, B.; Boke, H.; Unver, T. Boron Stress Responsive MicroRNAs and Their Targets in Barley. PLoS ONE 2013, 8, e59543. [Google Scholar] [CrossRef]
- Hackenberg, M.; Huang, P.-J.; Huang, C.-Y.; Shi, B.-J.; Gustafson, P.; Langridge, P. A Comprehensive Expression Profile of MicroRNAs and Other Classes of Non-Coding Small RNAs in Barley Under Phosphorous-Deficient and -Sufficient Conditions. DNA Res. 2013, 20, 109–125. [Google Scholar] [CrossRef]
- Sega, P.; Kruszka, K.; Bielewicz, D.; Karlowski, W.; Nuc, P.; Szweykowska-Kulinska, Z.; Pacak, A. Pi-Starvation Induced Transcriptional Changes in Barley Revealed by a Comprehensive RNA-Seq and Degradome Analyses. BMC Genom. 2021, 22, 165. [Google Scholar] [CrossRef]
- Bai, B.; Bian, H.; Zeng, Z.; Hou, N.; Shi, B.; Wang, J.; Zhu, M.; Han, N. MiR393-Mediated Auxin Signaling Regulation Is Involved in Root Elongation Inhibition in Response to Toxic Aluminum Stress in Barley. Plant Cell Physiol. 2017, 58, 426–439. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Yu, J.; Shen, Q.; Huang, L.; Wu, D.; Zhang, G. Identification of MicroRNAs in Response to Aluminum Stress in the Roots of Tibetan Wild Barley and Cultivated Barley. BMC Genom. 2018, 19, 560. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Wu, L.; Fu, L.; Shen, Q.; Kuang, L.; Wu, D.; Zhang, G. Genotypic Difference of Cadmium Tolerance and the Associated MicroRNAs in Wild and Cultivated Barley. Plant Growth Regul. 2019, 87, 389–401. [Google Scholar] [CrossRef]
- Chen, F.; He, J.; Jin, G.; Chen, Z.-H.; Dai, F. Identification of Novel MicroRNAs for Cold Deacclimation in Barley. Plant Growth Regul. 2020, 92, 389–400. [Google Scholar] [CrossRef]
- Kruszka, K.; Pacak, A.; Swida-Barteczka, A.; Nuc, P.; Alaba, S.; Wroblewska, Z.; Karlowski, W.; Jarmolowski, A.; Szweykowska-Kulinska, Z. Transcriptionally and Post-Transcriptionally Regulated MicroRNAs in Heat Stress Response in Barley. J. Exp. Bot. 2014, 65, 6123–6135. [Google Scholar] [CrossRef]
- Schreiber, A.W.; Shi, B.-J.; Huang, C.-Y.; Langridge, P.; Baumann, U. Discovery of Barley MiRNAs through Deep Sequencing of Short Reads. BMC Genom. 2011, 12, 129. [Google Scholar] [CrossRef]
- Lv, S.; Nie, X.; Wang, L.; Du, X.; Biradar, S.S.; Jia, X.; Weining, S. Identification and Characterization of MicroRNAs from Barley (Hordeum vulgare L.) by High-Throughput Sequencing. Int. J. Mol. Sci. 2012, 13, 2973–2984. [Google Scholar] [CrossRef]
- Kruszka, K.; Pacak, A.; Swida-Barteczka, A.; Stefaniak, A.K.; Kaja, E.; Sierocka, I.; Karlowski, W.; Jarmolowski, A.; Szweykowska-Kulinska, Z. Developmentally Regulated Expression and Complex Processing of Barley Pri-MicroRNAs. BMC Genom. 2013, 14, 34. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Cheng, X.; Liu, D.; Xu, W.; Wise, R.; Shen, Q.-H. The MiR9863 Family Regulates Distinct Mla Alleles in Barley to Attenuate NLR Receptor-Triggered Disease Resistance and Cell-Death Signaling. PLoS Genet. 2014, 10, e1004755. [Google Scholar] [CrossRef] [PubMed]
- Kis, A.; Tholt, G.; Ivanics, M.; Várallyay, É.; Jenes, B.; Havelda, Z. Polycistronic Artificial MiRNA-Mediated Resistance to Wheat Dwarf Virus in Barley Is Highly Efficient at Low Temperature. Mol. Plant Pathol. 2016, 17, 427–437. [Google Scholar] [CrossRef]
- Deng, P.; Bian, J.; Yue, H.; Feng, K.; Wang, M.; Du, X.; Weining, S.; Nie, X. Characterization of MicroRNAs and Their Targets in Wild Barley (Hordeum vulgare Subsp. Spontaneum) Using Deep Sequencing. Genome 2016, 59, 339–348. [Google Scholar] [CrossRef]
- Pacak, A.M.; Kruszka, K.; Świda-Barteczka, A.; Nuc, P.; Karlowski, W.; Jarmołowski, A.; Szweykowska-Kulińska, Z. Developmental Changes in Barley MicroRNA Expression Profiles Coupled with MiRNA Targets Analysis. Acta Biochim. Pol. 2016, 63, 799–809. [Google Scholar] [CrossRef]
- Bai, B.; Shi, B.; Hou, N.; Cao, Y.; Meng, Y.; Bian, H.; Zhu, M.; Han, N. MicroRNAs Participate in Gene Expression Regulation and Phytohormone Cross-Talk in Barley Embryo during Seed Development and Germination. BMC Plant Biol. 2017, 17, 150. [Google Scholar] [CrossRef] [PubMed]
- Smith, O.; Palmer, S.A.; Clapham, A.J.; Rose, P.; Liu, Y.; Wang, J.; Allaby, R.G. Small RNA Activity in Archeological Barley Shows Novel Germination Inhibition in Response to Environment. Mol. Biol. Evol. 2017, 34, 2555–2562. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, R.K.; Bregitzer, P.; Singh, J. Genome-Wide Analysis of the SPL/MiR156 Module and Its Interaction with the AP2/MiR172 Unit in Barley. Sci. Rep. 2018, 8, 7085. [Google Scholar] [CrossRef] [PubMed]
- Plaksenkova, I.; Kokina, I.; Petrova, A.; Jermaļonoka, M.; Gerbreders, V.; Krasovska, M. The Impact of Zinc Oxide Nanoparticles on Cytotoxicity, Genotoxicity, and MiRNA Expression in Barley (Hordeum vulgare L.) Seedlings. Sci. World J. 2020, 2020, 6649746. [Google Scholar] [CrossRef]
- Ye, Z.; Zeng, J.; Long, L.; Ye, L.; Zhang, G. Identification of MicroRNAs in Response to Low Potassium Stress in the Shoots of Tibetan Wild Barley and Cultivated. Curr. Plant Biol. 2021, 25, 100193. [Google Scholar] [CrossRef]
- Puchta, M.; Groszyk, J.; Małecka, M.; Koter, M.D.; Niedzielski, M.; Rakoczy-Trojanowska, M.; Boczkowska, M. Barley Seeds MiRNome Stability during Long-Term Storage and Aging. Int. J. Mol. Sci. 2021, 22, 4315. [Google Scholar] [CrossRef]
- Yao, X.; Wang, Y.; Yao, Y.; Bai, Y.; Wu, K.; Qiao, Y. Identification MicroRNAs and Target Genes in Tibetan Hulless Barley to BLS Infection. Agron. J. 2021, 113, 2273–2292. [Google Scholar] [CrossRef]
- Wang, N.-H.; Zhou, X.-Y.; Shi, S.-H.; Zhang, S.; Chen, Z.-H.; Ali, M.A.; Ahmed, I.M.; Wang, Y.; Wu, F. An MiR156-Regulated Nucleobase-Ascorbate Transporter 2 Confers Cadmium Tolerance via Enhanced Anti-Oxidative Capacity in Barley. J. Adv. Res. 2022, in press. [Google Scholar] [CrossRef]
- Liao, P.; Li, S.; Cui, X.; Zheng, Y. A Comprehensive Review of Web-Based Resources of Non-Coding RNAs for Plant Science Research. Int. J. Biol. Sci. 2018, 14, 819–832. [Google Scholar] [CrossRef]
- Yi, X.; Zhang, Z.; Ling, Y.; Xu, W.; Su, Z. PNRD: A Plant Non-Coding RNA Database. Nucleic Acids Res. 2015, 43, D982–D989. [Google Scholar] [CrossRef]
- Guo, Z.; Kuang, Z.; Zhao, Y.; Deng, Y.; He, H.; Wan, M.; Tao, Y.; Wang, D.; Wei, J.; Li, L. PmiREN2.0: From Data Annotation to Functional Exploration of Plant MicroRNAs. Nucleic Acids Res. 2022, 50, D1475–D1482. [Google Scholar] [CrossRef] [PubMed]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. MiRBase: From MicroRNA Sequences to Function. Nucleic Acids Res. 2019, 47, D155–D162. [Google Scholar] [CrossRef] [PubMed]
- Lunardon, A.; Johnson, N.R.; Hagerott, E.; Phifer, T.; Polydore, S.; Coruh, C.; Axtell, M.J. Integrated Annotations and Analyses of Small RNA–Producing Loci from 47 Diverse Plants. Genome Res. 2020, 30, 497–513. [Google Scholar] [CrossRef] [PubMed]
- Szcześniak, M.W.; Makalowska, I. MiRNEST 2.0: A Database of Plant and Animal MicroRNAs. Nucleic Acids Res. 2014, 42, D74–D77. [Google Scholar] [CrossRef]
- Liu, J.; Liu, X.; Zhang, S.; Liang, S.; Luan, W.; Ma, X. TarDB: An Online Database for Plant MiRNA Targets and MiRNA-Triggered Phased SiRNAs. BMC Genom. 2021, 22, 348. [Google Scholar] [CrossRef] [PubMed]
- Dou, X.; Zhou, Z.; Zhao, L. Identification and Expression Analysis of MiRNAs in Germination and Seedling Growth of Tibetan Hulless Barley. Genomics 2021, 113, 3735–3749. [Google Scholar] [CrossRef]
- Xing, S.; Salinas, M.; Höhmann, S.; Berndtgen, R.; Huijser, P. MiR156-Targeted and Nontargeted SBP-Box Transcription Factors Act in Concert to Secure Male Fertility in Arabidopsis. Plant Cell 2010, 22, 3935–3950. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Zheng, F.; Wang, J.; Zhang, C.; Xiao, F.; Ye, J.; Li, C.; Ye, Z.; Zhang, J. MiR156a-Targeted SBP-Box Transcription Factor SlSPL13 Regulates Inflorescence Morphogenesis by Directly Activating SFT in Tomato. Plant Biotechnol. J. 2020, 18, 1670–1682. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Cheng, X.; Liu, P.; Sun, J. MiR156-Targeted SBP-Box Transcription Factors Interact with DWARF53 to Regulate TEOSINTE BRANCHED1 and BARREN STALK1 Expression in Bread Wheat. Plant Physiol. 2017, 174, 1931–1948. [Google Scholar] [CrossRef] [PubMed]
- Millar, A.A.; Lohe, A.; Wong, G. Biology and Function of MiR159 in Plants. Plants 2019, 8, 255. [Google Scholar] [CrossRef] [PubMed]
- Csukasi, F.; Donaire, L.; Casañal, A.; Martínez-Priego, L.; Botella, M.A.; Medina-Escobar, N.; Llave, C.; Valpuesta, V. Two Strawberry MiR159 Family Members Display Developmental-Specific Expression Patterns in the Fruit Receptacle and Cooperatively Regulate Fa-GAMYB. New Phytol. 2012, 195, 47–57. [Google Scholar] [CrossRef] [PubMed]
- da Silva, E.M.; Silva, G.F.F.E.; Bidoia, D.B.; da Silva Azevedo, M.; de Jesus, F.A.; Pino, L.E.; Peres, L.E.P.; Carrera, E.; López-Díaz, I.; Nogueira, F.T.S. Micro RNA 159-Targeted Sl GAMYB Transcription Factors Are Required for Fruit Set in Tomato. Plant J. 2017, 92, 95–109. [Google Scholar] [CrossRef] [PubMed]
- Yadav, A.; Kumar, S.; Verma, R.; Lata, C.; Sanyal, I.; Rai, S.P. MicroRNA 166: An Evolutionarily Conserved Stress Biomarker in Land Plants Targeting HD-ZIP Family. Physiol. Mol. Biol. Plants 2021, 27, 2471–2485. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Fang, R.; Deng, R.; Li, J. The OsmiRNA166b-OsHox32 Pair Regulates Mechanical Strength of Rice Plants by Modulating Cell Wall Biosynthesis. Plant Biotechnol. J. 2021, 19, 1468–1480. [Google Scholar] [CrossRef] [PubMed]
- Javed, M.; Solanki, M.; Sinha, A.; Shukla, L.I. Position Based Nucleotide Analysis of MiR168 Family in Higher Plants and Its Targets in Mammalian Transcripts. MicroRNA 2017, 6, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Um, T.; Choi, J.; Park, T.; Chung, P.J.; Jung, S.E.; Shim, J.S.; Kim, Y.S.; Choi, I.-Y.; Park, S.C.; Oh, S.-J. Rice MicroRNA171f/SCL6 Module Enhances Drought Tolerance by Regulation of Flavonoid Biosynthesis Genes. Plant Direct 2022, 6, e374. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Zhou, J.; Gao, L.; Tang, Y. Plant MiR397 and Its Functions. Funct. Plant Biol. 2020, 48, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.-H.; Pei, H. Over-Expression of MiR397 Improves Plant Tolerance to Cold Stress in Arabidopsis Thaliana. J. Plant Biol. 2014, 57, 209–217. [Google Scholar] [CrossRef]
- Pegler, J.L.; Oultram, J.M.; Grof, C.P.; Eamens, A.L. Molecular Manipulation of the MiR399/PHO2 Expression Module Alters the Salt Stress Response of Arabidopsis Thaliana. Plants 2020, 10, 73. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Ahn, H.J.; Chiou, T.-J.; Ahn, J.H. The Role of the MiR399-PHO2 Module in the Regulation of Flowering Time in Response to Different Ambient Temperatures in Arabidopsis Thaliana. Mol. Cells 2011, 32, 83–88. [Google Scholar] [CrossRef]
- Yan, Y.; Wang, H.; Hamera, S.; Chen, X.; Fang, R. MiR444a Has Multiple Functions in the Rice Nitrate-Signaling Pathway. Plant J. 2014, 78, 44–55. [Google Scholar] [CrossRef]
- Wang, H.; Jiao, X.; Kong, X.; Hamera, S.; Wu, Y.; Chen, X.; Fang, R.; Yan, Y. A Signaling Cascade from MiR444 to RDR1 in Rice Antiviral RNA Silencing Pathway. Plant Physiol. 2016, 170, 2365–2377. [Google Scholar] [CrossRef]
- Jiao, X.; Wang, H.; Yan, J.; Kong, X.; Liu, Y.; Chu, J.; Chen, X.; Fang, R.; Yan, Y. Promotion of BR Biosynthesis by MiR444 Is Required for Ammonium-Triggered Inhibition of Root Growth. Plant Physiol. 2020, 182, 1454–1466. [Google Scholar] [CrossRef]
- Sun, L.; Sun, G.; Shi, C.; Sun, D. Transcriptome Analysis Reveals New MicroRNAs-Mediated Pathway Involved in Anther Development in Male Sterile Wheat. BMC Genom. 2018, 19, 333. [Google Scholar] [CrossRef] [PubMed]
- Bano, N.; Fakhrah, S.; Nayak, S.P.; Bag, S.K.; Mohanty, C.S. Identification of MiRNA and Their Target Genes in Cestrum nocturnum L. and Cestrum diurnum L. in Stress Responses. Physiol. Mol. Biol. Plants 2022, 28, 31–49. [Google Scholar] [CrossRef]
- Li, T.; Ma, L.; Geng, Y.; Hao, C.; Chen, X.; Zhang, X. Small RNA and Degradome Sequencing Reveal Complex Roles of MiRNAs and Their Targets in Developing Wheat Grains. PLoS ONE 2015, 10, e0139658. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, X.; Yuan, L.; Liu, Y.; Shen, T.; Zhang, Y. Comparative Small RNA Profiling and Functional Exploration on Wheat with High-and Low-Cadmium Accumulation. Front. Genet. 2021, 12, 635599. [Google Scholar] [CrossRef]
- Samad, A.F.A.; Sajad, M.; Nazaruddin, N.; Fauzi, I.A.; Murad, A.M.A.; Zainal, Z.; Ismail, I. MicroRNA and Transcription Factor: Key Players in Plant Regulatory Network. Front. Plant Sci. 2017, 8, 565. [Google Scholar] [CrossRef] [PubMed]
- Van Bel, M.; Diels, T.; Vancaester, E.; Kreft, L.; Botzki, A.; Van de Peer, Y.; Coppens, F.; Vandepoele, K. PLAZA 4.0: An Integrative Resource for Functional, Evolutionary and Comparative Plant Genomics. Nucleic Acids Res. 2018, 46, D1190–D1196. [Google Scholar] [CrossRef]
- Van Bel, M.; Silvestri, F.; Weitz, E.M.; Kreft, L.; Botzki, A.; Coppens, F.; Vandepoele, K. PLAZA 5.0: Extending the Scope and Power of Comparative and Functional Genomics in Plants. Nucleic Acids Res. 2022, 50, D1468–D1474. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.-M.; Kuang, S.; Xiong, X.; Gao, T.; Liu, C.; Guo, A.-Y. Transcription Factor and MicroRNA Co-Regulatory Loops: Important Regulatory Motifs in Biological Processes and Diseases. Brief. Bioinform. 2015, 16, 45–58. [Google Scholar] [CrossRef] [PubMed]
- Selbach, M.; Schwanhäusser, B.; Thierfelder, N.; Fang, Z.; Khanin, R.; Rajewsky, N. Widespread Changes in Protein Synthesis Induced by MicroRNAs. Nature 2008, 455, 58–63. [Google Scholar] [CrossRef]
- Baldrich, P.; Beric, A.; Meyers, B.C. Despacito: The Slow Evolutionary Changes in Plant MicroRNAs. Curr. Opin. Plant Biol. 2018, 42, 16–22. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, L.; Chen, D.; Wu, X.; Huang, D.; Chen, L.; Li, L.; Deng, X.; Xu, Q. Genome-Wide Comparison of MicroRNAs and Their Targeted Transcripts among Leaf, Flower and Fruit of Sweet Orange. BMC Genom. 2014, 15, 695. [Google Scholar] [CrossRef] [PubMed]
- Baldrich, P.; Campo, S.; Wu, M.-T.; Liu, T.-T.; Hsing, Y.-I.C.; Segundo, B.S. MicroRNA-Mediated Regulation of Gene Expression in the Response of Rice Plants to Fungal Elicitors. RNA Biol. 2015, 12, 847–863. [Google Scholar] [CrossRef] [PubMed]
- Banks, I.R.; Zhang, Y.; Wiggins, B.E.; Heck, G.R.; Ivashuta, S. RNA Decoys. Plant Signal. Behav. 2012, 7, 1188–1193. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Liu, C.; Gu, L.; Mo, B.; Cao, X.; Chen, X. TarHunter, a Tool for Predicting Conserved MicroRNA Targets and Target Mimics in Plants. Bioinformatics 2018, 34, 1574–1576. [Google Scholar] [CrossRef]
- Unver, T.; Tombuloglu, H. Barley Long Non-Coding RNAs (LncRNA) Responsive to Excess Boron. Genomics 2020, 112, 1947–1955. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Cheng, Y.; Feng, K.; Ruan, M.; Ye, Q.; Wang, R.; Li, Z.; Zhou, G.; Yao, Z.; Yang, Y.; et al. Genome-Wide Identification and Expression Profiling of Tomato Hsp20 Gene Family in Response to Biotic and Abiotic Stresses. Front. Plant Sci. 2016, 7, 1215. [Google Scholar] [CrossRef] [PubMed]
- Lewis, B.P.; Burge, C.B.; Bartel, D.P. Conserved Seed Pairing, Often Flanked by Adenosines, Indicates That Thousands of Human Genes Are MicroRNA Targets. Cell 2005, 120, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Ni, W.-J.; Leng, X.-M. Dynamic MiRNA–MRNA Paradigms: New Faces of MiRNAs. Biochem. Biophys. Rep. 2015, 4, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Gruber, A.R.; Lorenz, R.; Bernhart, S.H.; Neuböck, R.; Hofacker, I.L. The Vienna RNA Websuite. Nucleic Acids Res. 2008, 36, W70–W74. [Google Scholar] [CrossRef] [PubMed]
- Kerpedjiev, P.; Hammer, S.; Hofacker, I.L. Forna (Force-Directed RNA): Simple and Effective Online RNA Secondary Structure Diagrams. Bioinformatics 2015, 31, 3377–3379. [Google Scholar] [CrossRef]
- Jones-Rhoades, M.W.; Bartel, D.P. Computational Identification of Plant MicroRNAs and Their Targets, Including a Stress-Induced MiRNA. Mol. Cell 2004, 14, 787–799. [Google Scholar] [CrossRef]
- Sun, Y.-H.; Lu, S.; Shi, R.; Chiang, V.L. Computational Prediction of Plant MiRNA Targets. In RNAi and Plant Gene Function Analysis; Humana Press: Totowa, NJ, USA, 2011; pp. 175–186. [Google Scholar]
- Dai, X.; Zhuang, Z.; Zhao, P.X. Computational Analysis of MiRNA Targets in Plants: Current Status and Challenges. Brief. Bioinform. 2011, 12, 115–121. [Google Scholar] [CrossRef]
- Fahlgren, N.; Carrington, J.C. MiRNA Target Prediction in Plants. In Plant MicroRNAs; Humana Press: Totowa, NJ, USA, 2010; pp. 51–57. [Google Scholar]
- Pandey, P.; Srivastava, P.K.; Pandey, S.P. Prediction of Plant MiRNA Targets. In Plant MicroRNAs; Humana Press: Totowa, NJ, USA, 2019; pp. 99–107. [Google Scholar]
- Rhoades, M.W.; Reinhart, B.J.; Lim, L.P.; Burge, C.B.; Bartel, B.; Bartel, D.P. Prediction of Plant MicroRNA Targets. Cell 2002, 110, 513–520. [Google Scholar] [CrossRef]
- Llave, C.; Xie, Z.; Kasschau, K.D.; Carrington, J.C. Cleavage of Scarecrow-like MRNA Targets Directed by a Class of Arabidopsis MiRNA. Science 2002, 297, 2053–2056. [Google Scholar] [CrossRef] [PubMed]
- Riolo, G.; Cantara, S.; Marzocchi, C.; Ricci, C. MiRNA Targets: From Prediction Tools to Experimental Validation. Methods Protoc. 2020, 4, 1. [Google Scholar] [CrossRef] [PubMed]
- Patel, P.; Yadav, K.; Ganapathi, T.R.; Penna, S. Plant MiRNAome: Cross Talk in Abiotic Stressful Times. In Genetic Enhancement of Crops for Tolerance to Abiotic Stress: Mechanisms and Approaches, Vol. I; Rajpal, V.R., Sehgal, D., Kumar, A., Raina, S.N., Eds.; Sustainable Development and Biodiversity; Springer International Publishing: Cham, Switzesland, 2019; pp. 25–52. ISBN 978-3-319-91956-0. [Google Scholar]
- Zhang, B.; Wang, Q. MicroRNA-Based Biotechnology for Plant Improvement. J. Cell. Physiol. 2015, 230, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B. MicroRNA: A New Target for Improving Plant Tolerance to Abiotic Stress. J. Exp. Bot. 2015, 66, 1749–1761. [Google Scholar] [CrossRef] [PubMed]
- Ferdous, J.; Whitford, R.; Nguyen, M.; Brien, C.; Langridge, P.; Tricker, P.J. Drought-Inducible Expression of Hv-MiR827 Enhances Drought Tolerance in Transgenic Barley. Funct. Integr. Genom. 2017, 17, 279–292. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Yang, T.; Guo, Z.; Wang, Q.; Chai, M.; Wu, M.; Li, X.; Li, W.; Li, G.; Tang, J.; et al. Maize MicroRNA166 Inactivation Confers Plant Development and Abiotic Stress Resistance. Int. J. Mol. Sci. 2020, 21, 9506. [Google Scholar] [CrossRef] [PubMed]
- Chung, P.J.; Chung, H.; Oh, N.; Choi, J.; Bang, S.W.; Jung, S.E.; Jung, H.; Shim, J.S.; Kim, J.-K. Efficiency of Recombinant CRISPR/RCas9-Mediated MiRNA Gene Editing in Rice. Int. J. Mol. Sci. 2020, 21, 9606. [Google Scholar] [CrossRef]
- Hua, K.; Tao, X.; Zhu, J.-K. Expanding the Base Editing Scope in Rice by Using Cas9 Variants. Plant Biotechnol. J. 2019, 17, 499–504. [Google Scholar] [CrossRef]
- Ó’Maoiléidigh, D.S.; van Driel, A.D.; van Singh, A.; Sang, Q.; LeBec, N.; Vincent, C.; de Olalla, E.B.G.; Vayssières, A.; Branchat, M.R.; Severing, E.; et al. Systematic Analyses of the MIR172 Family Members of Arabidopsis Define Their Distinct Roles in Regulation of APETALA2 during Floral Transition. PLoS Biol. 2021, 19, e3001043. [Google Scholar] [CrossRef] [PubMed]
- Deng, F.; Zeng, F.; Shen, Q.; Abbas, A.; Cheng, J.; Jiang, W.; Chen, G.; Shah, A.N.; Holford, P.; Tanveer, M.; et al. Molecular Evolution and Functional Modification of Plant MiRNAs with CRISPR. Trends Plant Sci. 2022, 27, 890–907. [Google Scholar] [CrossRef] [PubMed]
- Dalakouras, A.; Wassenegger, M.; Dadami, E.; Ganopoulos, I.; Pappas, M.L.; Papadopoulou, K. Genetically Modified Organism-Free RNA Interference: Exogenous Application of RNA Molecules in Plants. Plant Physiol. 2020, 182, 38–50. [Google Scholar] [CrossRef] [PubMed]
- Mujtaba, M.; Wang, D.; Carvalho, L.B.; Oliveira, J.L.; Espirito Santo Pereira, A.D.; Sharif, R.; Jogaiah, S.; Paidi, M.K.; Wang, L.; Ali, Q.; et al. Nanocarrier-Mediated Delivery of MiRNA, RNAi, and CRISPR-Cas for Plant Protection: Current Trends and Future Directions. ACS Agric. Sci. Technol. 2021, 1, 417–435. [Google Scholar] [CrossRef]
- Ražná, K.; Rataj, V.; Macák, M.; Galambošová, J. MicroRNA-Based Markers as a Tool to Monitor the Barley (Hordeum vulgare L.) Response to Soil Compaction. Acta Fytotech. Zootech. 2020, 23, 139–146. [Google Scholar] [CrossRef]
- Gurjar, A.K.S.; Panwar, A.S.; Gupta, R.; Mantri, S.S. PmiRExAt: Plant MiRNA Expression Atlas Database and Web Applications. Database 2016, 2016, baw060. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).