
Tocotrienol-Rich Fractions Offer Potential to Suppress Pulmonary Fibrosis Progression

 ,
,  ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Results and Discussion
2.1. Effects on Weight (g), Feed Intake (g) and Lung Index of TRF
2.2. Effects on HYP and MMP-7 Level of TRF
2.3. Effects on Inflammatory Markers of TRF
2.4. Effects on Antioxidant Enzymes of TRF
2.5. Effects on Histological Evaluation of TRF
2.6. IHC Determination on Collagen I and Collagen II of TRF
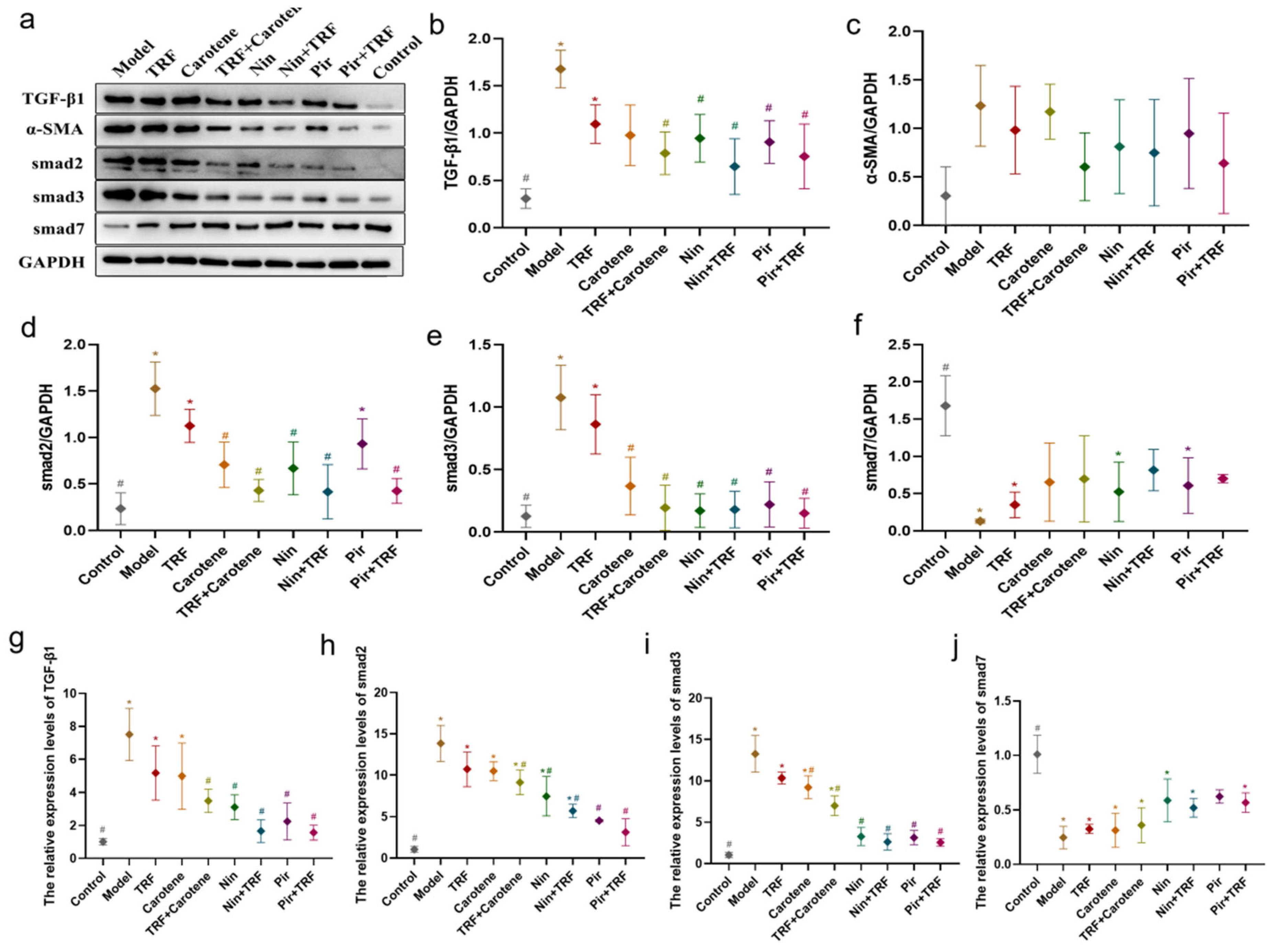
2.7. Effects on TGF-β/Smad Signaling Pathway of TRF
2.8. Effects on PI3K/Akt/mTOR Signaling Pathway of TRF
2.9. Effects on NF-κB Signaling Pathway of TRF
2.10. Discussion
3. Materials and Methods
3.1. Animals, Interventions and Experimental Design
3.2. Enzyme-Linked Immunosorbent Assay (ELISA)
3.3. Histopathological Analysis
3.4. Immunohistochemical (IHC) Determination of Collagen I and Collagen II
3.5. Western Blotting
3.6. Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR) Analysis
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| COVID-19 | Coronavirus disease |
| PF | Pulmonary fibrosis |
| FDA | U.S. Food and Drug Administration |
| Pir | Pirfenidone |
| Nin | Nintedanib |
| TRF | Tocotrienol-rich fractions |
| Carotene | Natural Mixed-Carotene Complex 20% Oil Concentrate |
| TGF-β1 | Transforming growth factor beta 1 |
| IL-6 | Interleukin-6 |
| MPO | Myeloperoxidase |
| IL-1β | Interleukin-1β |
| TNFα | Tumor necrosis factor-alpha |
| CAT | Catalase |
| MDA | Malondialdehyde |
| GSH | Glutathione |
| NO | Nitric oxide |
| SOD | Superoxide dismutase |
| HE | Hematoxylin-eosin |
| PBS | Phosphate-buffered saline |
| SA/HRP | Streptomyces ovalbumin working solution |
| DAB | Diaminobenzidine |
| WB | Western blot |
| ECL | Enhanced chemiluminescence |
| PI3K | Phosphatidylinositol 3-kinase |
| AKT | Protein kinase B, PKB |
| mTOR | Mammalian target of rapamycin |
| IFN-γ | Interferonγ |
| IL-13 | Interleukin-13 |
| IkBα | NF-kappa-B inhibitor alpha |
| Ikkβ | Inhibitor of nuclear factor kappa-B kinase |
| ECM | Excessive deposition of extracellular matrix |
References
- George, P.M.; Wells, A.U.; Jenkins, R.G. Pulmonary fibrosis and COVID-19: The potential role for antifibrotic therapy. Lancet Respir. Med. 2020, 8, 807–815. [Google Scholar] [CrossRef]
- Kayarat, B.; Khanna, P.; Sarkar, S. Pulmonary Fibrosis in COVID-19 Recovered Patients: Problem and Potential Management. Indian J. Crit. Care Med. 2021, 25, 242–244. [Google Scholar] [CrossRef] [PubMed]
- Thille, A.W.; Esteban, A.; Fernandez-Segoviano, P.; Rodriguez, J.M.; Aramburu, J.A.; Vargas-Errazuriz, P.; Martin-Pellicer, A.; Lorente, J.A.; Frutos-Vivar, F. Chronology of histological lesions in acute respiratory distress syndrome with diffuse alveolar damage: A prospective cohort study of clinical autopsies. Lancet Respir. Med. 2013, 1, 395–401. [Google Scholar] [CrossRef]
- Grillo, F.; Barisione, E.; Ball, L.; Mastracci, L.; Fiocca, R. Lung fibrosis: An undervalued finding in COVID-19 pathological series. Lancet Infect. Dis. 2020, 21, E72. [Google Scholar] [CrossRef]
- Wynn, T.A.; Ramalingam, T.R. Mechanisms of fibrosis: Therapeutic translation for fibrotic disease. Nat. Med. 2012, 18, 1028–1040. [Google Scholar] [CrossRef] [PubMed]
- Morais, A.; Beltrao, M.; Sokhatska, O.; Costa, D.; Melo, N.; Mota, P.; Marques, A.; Delgado, L. Serum metalloproteinases 1 and 7 in the diagnosis of idiopathic pulmonary fibrosis and other interstitial pneumonias. Respir. Med. 2015, 109, 1063–1068. [Google Scholar] [CrossRef]
- Mapel, D.W.; Samet, J.M.; Coultas, D.B. Corticosteroids and the treatment of idiopathic pulmonary fibrosis—Past, present, and future. Chest 1996, 110, 1058–1067. [Google Scholar] [CrossRef]
- Raghu, G.; Remy-Jardin, M.; Myers, J.L.; Richeldi, L.; Ryerson, C.J.; Lederer, D.J.; Behr, J.; Cottin, V.; Danoff, S.K.; Morell, F.; et al. Diagnosis of Idiopathic Pulmonary Fibrosis An Official ATS/ERS/JRS/ALAT Clinical Practice Guideline. Am. J. Respir. Crit. Care Med. 2018, 198, E44–E68. [Google Scholar] [CrossRef]
- Spagnolo, P.; Maher, T.M.; Richeldi, L. Idiopathic pulmonary fibrosis: Recent advances on pharmacological therapy. Pharmacol. Ther. 2015, 152, 18–27. [Google Scholar] [CrossRef]
- Lehmann, S.; Uhlemann, M.; Leontyev, S.; Seeburger, J.; Garbade, J.; Merk, D.R.; Bittner, H.B.; Mohr, F.W. Bilateral Versus Single Lung Transplant for Idiopathic Pulmonary Fibrosis. Exp. Clin. Transplant. 2014, 12, 443–447. [Google Scholar] [CrossRef]
- Kumar, A.; Kapnadak, S.G.; Girgis, R.E.; Raghu, G. Lung transplantation in idiopathic pulmonary fibrosis. Expert Rev. Respir. Med. 2018, 12, 375–385. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Sundaram, C.; Prasad, S.; Kannappan, R. Tocotrienols, the vitamin E of the 21st century: Its potential against cancer and other chronic diseases. Biochem. Pharmacol. 2010, 80, 1613–1631. [Google Scholar] [CrossRef]
- Sailo, B.L.; Banik, K.; Padmavathi, G.; Javadi, M.; Bordoloi, D.; Kunnumakkara, A.B. Tocotrienols: The promising analogues of vitamin E for cancer therapeutics. Pharmacol. Res. 2018, 130, 259–272. [Google Scholar] [CrossRef]
- Kuhad, A.; Chopra, K. Attenuation of diabetic nephropathy by tocotrienol: Involvement of NFkB signaling pathway. Life Sci. 2009, 84, 296–301. [Google Scholar] [CrossRef]
- Ahmad, N.S.; Khalid, B.A.K.; Luke, D.A.; Nirwana, S.I. Tocotrienol offers better protection than tocopherol from free radical-induced damage of rat bone. Clin. Exp. Pharmacol. Physiol. 2005, 32, 761–770. [Google Scholar] [CrossRef]
- Ahn, K.S.; Sethi, G.; Krishnan, K.; Aggarwal, B.B. γ-Tocotrienol inhibits nuclear factor-κB signaling pathway through inhibition of receptor-interacting protein and TAK1 leading to suppression of antiapoptotic gene products and Potentiation of apoptosis. J. Biol. Chem. 2007, 282, 809–820. [Google Scholar] [CrossRef]
- Lee, S.P.; Mar, G.Y.; Ng, L.T. Effects of tocotrienol-rich fraction on exercise endurance capacity and oxidative stress in forced swimming rats. Eur. J. Appl. Physiol. 2009, 107, 587–595. [Google Scholar] [CrossRef]
- Palozza, P.; Verdecchia, S.; Avanzi, L.; Vertuani, S.; Serini, S.; Iannone, A.; Manfredini, S. Comparative antioxidant activity of tocotrienols and the novel chromanyl-polyisoprenyl molecule FeAox-6 in isolated membranes and intact cells. Mol. Cell. Biochem. 2006, 287, 21–32. [Google Scholar] [CrossRef]
- Osakada, F.; Hashino, A.; Kume, T.; Katsuki, H.; Kaneko, S.; Akaike, A. α-Tocotrienol provides the most potent neuroprotection among vitamin E analogs on cultured striatal neurons. Neuropharmacology 2004, 47, 904–915. [Google Scholar] [CrossRef]
- Palace, V.P.; Khaper, N.; Qin, Q.; Singal, P.K. Antioxidant potentials of vitamin A and carotenoids and their relevance to heart disease. Free Radic. Biol. Med. 1999, 26, 746–761. [Google Scholar] [CrossRef]
- de Bittencourt Pasquali, M.A.; Schnorr, C.E.; Hilbig Feistauer, L.B.; Gelain, D.P.; Fonseca Moreira, J.C. Vitamin A supplementation to pregnant and breastfeeding female rats induces oxidative stress in the neonatal lung. Reprod. Toxicol. 2010, 30, 452–456. [Google Scholar] [CrossRef] [PubMed]
- Cantin, A.M.; White, T.B.; Cross, C.E.; Forman, H.J.; Sokol, R.J.; Borowitz, D. Antioxidants in cystic fibrosis. Conclusions from the CF antioxidant workshop, Bethesda, Maryland, 11–12 November 2003. Free Radic. Biol. Med. 2007, 42, 15–31. [Google Scholar] [CrossRef] [PubMed]
- Lenz, A.G.; Costabel, U.; Maier, K.L. Oxidized BAL fluid proteins in patients with interstitial lung diseases. Eur. Respir. J. 1996, 9, 307–312. [Google Scholar] [CrossRef] [PubMed]
- de Bittencourt Pasquali, M.A.; Gelain, D.P.; de Oliveira, M.R.; Behr, G.A.; da Motta, L.L.; da Rocha, R.F.; Klamt, F.; Fonseca Moreira, J.C. Vitamin A Supplementation for Different Periods Alters Oxidative Parameters in Lungs of Rats. J. Med. Food 2009, 12, 1375–1380. [Google Scholar] [CrossRef] [PubMed]
- Phan, S.H.; Varani, J.; Smith, D. Rat Lung Fibroblast Collagen Metabolism in Bleomycin-induced Pulmonary Fibrosis. J. Clin. Invest. 1985, 76, 241–247. [Google Scholar] [CrossRef]
- Pardo, A.; Selman, M. Matrix metalloproteases in aberrant fibrotic tissue remodeling. Proc. Am. Thorac. Soc. 2006, 3, 383–388. [Google Scholar] [CrossRef]
- Hu, H.H.; Chen, D.Q.; Wang, Y.N.; Feng, Y.L.; Cao, G.; Vaziri, N.D.; Zhao, Y.Y. New insights into TGF-β/Smad signaling in tissue fibrosis. Chem. Biol. Interact. 2018, 292, 76–83. [Google Scholar] [CrossRef]
- Ciruelos Gil, E.M. Targeting the PI3K/AKT/mTOR pathway in estrogen receptor-positive breast cancer. Cancer Treat. Rev. 2014, 40, 862–871. [Google Scholar] [CrossRef]
- Roncolato, F.; Lindemann, K.; Wilson, M.L.; Martyn, J.; Mileshkin, L. PI3K/AKT/mTOR inhibitors for advanced or recurrent endometrial cancer. Cochrane Database Syst. Rev. 2019, 10, CD012160. [Google Scholar] [CrossRef]
- Zeng, J.; Zhao, H.; Chen, B. DJ-1/PARK7 inhibits high glucose-induced oxidative stress to prevent retinal pericyte apoptosis via the PI3K/AKT/mTOR signaling pathway. Exp. Eye Res. 2019, 189, 107830. [Google Scholar] [CrossRef]
- Yang, Y.; Li, L. Depleting microRNA-146a-3p attenuates lipopolysaccharide-induced acute lung injury via up-regulating SIRT1 and mediating NF-κB pathway. J. Drug Target. 2021, 29, 420–429. [Google Scholar] [CrossRef]
- Ali, H.; Khan, A.; Ali, J.; Ullah, H.; Khan, A.; Ali, H.; Irshad, N.; Khan, S. Attenuation of LPS-induced acute lung injury by continentalic acid in rodents through inhibition of inflammatory mediators correlates with increased Nrf2 protein expression. BMC Pharmacol. Toxicol. 2020, 21, 81. [Google Scholar] [CrossRef]
- Abidi, A.; Aissani, N.; Sebai, H.; Serairi, R.; Kourda, N.; Ben Khamsa, S. Protective Effect of Pistacia lentiscus Oil Against Bleomycin-Induced Lung Fibrosis and Oxidative Stress in Rat. Nutr. Cancer 2017, 69, 490–497. [Google Scholar] [CrossRef]
- Raish, M.; Ahmad, A.; Ansari, M.A.; Ahad, A.; Al-Jenoobi, F.I.; Al-Mohizea, A.M.; Khan, A.; Ali, N. Sinapic acid ameliorates bleomycin-induced lung fibrosis in rats. Biomed. Pharmacother. 2018, 108, 224–231. [Google Scholar] [CrossRef]
- Lu, Y.; Zhang, Y.; Pan, Z.; Yang, C.; Chen, L.; Wang, Y.; Xu, D.; Xia, H.; Wang, S.; Chen, S.; et al. Potential “Therapeutic” Effects of Tocotrienol-Rich Fraction (TRF) and Carotene “Against” Bleomycin-Induced Pulmonary Fibrosis in Rats via TGF-β/Smad, PI3K/Akt/mTOR and NF-κB Signaling Pathways. Nutrients 2022, 14, 1094. [Google Scholar] [CrossRef]
- Bahri, S.; Ben Ali, R.; Nahdi, A.; Mlika, M.; Abdennabi, R.; Jameleddine, S. Salvia officinalis attenuates bleomycin-induced oxidative stress and lung fibrosis in rats. Nutr. Cancer Int. J. 2020, 72, 1135–1145. [Google Scholar] [CrossRef]
- Mei, S.H.J.; McCarter, S.D.; Deng, Y.; Parker, C.H.; Liles, W.C.; Stewart, D.J. Prevention of LPS-induced acute lung injury in mice by mesenchymal stem cells overexpressing angiopoietin 1. PLoS Med. 2007, 4, 1525–1537. [Google Scholar] [CrossRef]
- Rong, Y.; Cao, B.; Liu, B.; Li, W.; Chen, Y.; Chen, H.; Liu, Y.; Liu, T. A novel Gallic acid derivative attenuates BLM-induced pulmonary fibrosis in mice. Int. Immunopharmacol. 2018, 64, 183–191. [Google Scholar] [CrossRef]
- Martins, V.; Valenca, S.S.; Farias-Filho, F.A.; Molinaro, R.; Simoes, R.L.; Ferreira, T.P.T.; Silva, P.M.R.e.; Hogaboam, C.M.; Kunkel, S.L.; Fierro, I.M.; et al. ATLa, an Aspirin-Triggered Lipoxin A4 Synthetic Analog, Prevents the Inflammatory and Fibrotic Effects of Bleomycin-Induced Pulmonary Fibrosis. J. Immunol. 2009, 182, 5374–5381. [Google Scholar] [CrossRef]
- Hosseini, S.A.; Zahedipour, F.; Sathyapalan, T.; Jamialahmadi, T.; Sahebkar, A. Pulmonary fibrosis: Therapeutic and mechanistic insights into the role of phytochemicals. Biofactors 2021, 47, 250–269. [Google Scholar] [CrossRef]
- Wang, L.; Li, S.; Yao, Y.; Yin, W.; Ye, T. The role of natural products in the prevention and treatment of pulmonary fibrosis: A review. Food Funct. 2021, 12, 990–1007. [Google Scholar] [CrossRef] [PubMed]
- Landi, C.; Carleo, A.; Vantaggiato, L.; Bergantini, L.; d’Alessandro, M.; Cameli, P.; Sebastiani, G.; Dotta, F.; Bargagli, E. Common molecular pathways targeted by nintedanib in cancer and IPF: A bioinformatic study. Pulm. Pharmacol. Ther. 2020, 64, 101941. [Google Scholar] [CrossRef]
- Gokey, J.J.; Sridharan, A.; Xu, Y.; Green, J.; Carraro, G.; Stripp, B.R.; Perl, A.-K.T.; Whitsett, J.A. Active epithelial Hippo signaling in idiopathic pulmonary fibrosis. JCI Insight 2018, 3, e98738. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Nie, P.; Lou, Y.; Zhu, Y.; Jiang, S.; Li, B.; Luo, P. Pirfenidone is a renal protective drug: Mechanisms, signalling pathways, and preclinical evidence. Eur. J. Pharmacol. 2021, 911, 174503. [Google Scholar] [CrossRef]
- Gao, C.; Cao, X.; Huang, L.; Bao, Y.; Li, T.; Di, Y.; Wu, L.; Song, Y. Pirfenidone Alleviates Choroidal Neovascular Fibrosis through TGF-β/Smad Signaling Pathway. J. Ophthalmol. 2021, 2021, 8846708. [Google Scholar] [CrossRef]
- Kim, E.S.; Keating, G.M. Pirfenidone: A Review of Its Use in Idiopathic Pulmonary Fibrosis. Drugs 2015, 75, 219–230. [Google Scholar] [CrossRef]
- Ashcroft, T.; Simpson, J.M.; Timbrell, V. Simple method of estimating severity of pulmonary fibrosis on a numerical scale. J. Clin. Pathol. 1988, 41, 467–470. [Google Scholar] [CrossRef]
- Liu, M.-W.; Su, M.-X.; Tang, D.-Y.; Hao, L.; Xun, X.-H.; Huang, Y.-Q. Ligustrazin increases lung cell autophagy and ameliorates paraquat-induced pulmonary fibrosis by inhibiting PI3K/Akt/mTOR and hedgehog signalling via increasing miR-193a expression. BMC Pulm. Med. 2019, 19, 35. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Components | Values (mg/g) |
|---|---|
| α-Carotene | 65 |
| β-Carotene | 135 |
| γ-Carotene | 0.5 |
| Lycopene | 0.1 |
| Total mixed-carotene complex | 200.6 |
| Components | Values (wt/wt) |
|---|---|
| α-Tocopherol | 12.5 |
| α-Tocotrienol | 12.8 |
| β-Tocotrienol | 2.0 |
| γ-Tocotrienol | 19.5 |
| δ-Tocotrienol | 5.5 |
| Total mixed tocotrienols | 39.8 |
| Tocotrienol/Tocopherol complex | 52.3 |
| mRNA | Forward Primer | Reverse Primer |
|---|---|---|
| TGF-β1 | TCGCCCTTTCATTTCAGAT | TTTGCCGATGCTTTCTTG |
| Smad2 | AGGTGTCTCATCGGAAAG | CTCTGGTAGTGGTAAGGGT |
| Smad3 | AGCTTACAAGGCGGCACA | TGGGAGACTGGACGAAAA |
| Smad7 | CTTCCTCCGATGAAACCG | TCGAGTCTTCTCCTCCCAGTA |
| PI3K | GAAACCCAGTCACCTAGGGC | GGTGGGCAGTACGAACTCAA |
| AKT | GAGGAGCGGGAAGAGTG | GTGCCCTTGCCCAGTAG |
| mTOR | GGTGGACGAGCTCTTTGTC | AGGAGCCCTAACACTCGGAT |
| TNF-α | TGAGCACAGAAAGCATGATC | CATCTGCTGGTACCACCAGTT |
| IFN-γ | TTGCAGCTCTGCCTCAT | TTCGTGTTACCGTCCTT |
| IL-13 | CTCGCTTGCCTTGGTGG | TGATGTTGCTCAGCTCCTC |
| NF-κB | CTGTTTCCCCTCATCTTTCC | GTGCGTCTTAGTGGTATCTGTG |
| IkBα | CCAACTACAACGGCCACA | CAACAGGAGCGAGACCAG |
| Ikkβ | CATTGTTGTTAGCGAGGAC | CCCTTTGCCGAGGTTGC |
| GAPDH | AAGAAGG TGGTGAAGCAGGC | TCCACCACCCT GTTGCTGTA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, Y.; Zhang, Y.; Xu, D.; Wang, Y.; Pan, D.; Wang, P.; Xia, J.; Yin, S.; Liao, W.; Wang, S.; et al. Tocotrienol-Rich Fractions Offer Potential to Suppress Pulmonary Fibrosis Progression. Int. J. Mol. Sci. 2022, 23, 14331. https://doi.org/10.3390/ijms232214331
Lu Y, Zhang Y, Xu D, Wang Y, Pan D, Wang P, Xia J, Yin S, Liao W, Wang S, et al. Tocotrienol-Rich Fractions Offer Potential to Suppress Pulmonary Fibrosis Progression. International Journal of Molecular Sciences. 2022; 23(22):14331. https://doi.org/10.3390/ijms232214331
Chicago/Turabian StyleLu, Yifei, Yihan Zhang, Dengfeng Xu, Yuanyuan Wang, Da Pan, Pei Wang, Jiayue Xia, Shiyu Yin, Wang Liao, Shaokang Wang, and et al. 2022. "Tocotrienol-Rich Fractions Offer Potential to Suppress Pulmonary Fibrosis Progression" International Journal of Molecular Sciences 23, no. 22: 14331. https://doi.org/10.3390/ijms232214331
APA StyleLu, Y., Zhang, Y., Xu, D., Wang, Y., Pan, D., Wang, P., Xia, J., Yin, S., Liao, W., Wang, S., & Sun, G. (2022). Tocotrienol-Rich Fractions Offer Potential to Suppress Pulmonary Fibrosis Progression. International Journal of Molecular Sciences, 23(22), 14331. https://doi.org/10.3390/ijms232214331