

Pathophysiology and Therapeutic Approaches for Spinal Cord Injury
 ,
,  and
and 
Abstract
1. Spinal Cord Anatomy and Physiology
2. Spinal Cord Injury
2.1. Primary Injury
2.2. Secondary Injury
2.2.1. Permeability and Vascular Alterations
2.2.2. Ionic Disruption and Glutamate Excitotoxicity
2.2.3. Metabolic Alterations
2.2.4. Inflammatory Response
2.2.5. Inhibitory Environment
2.2.6. Spinal Cord Scarring
2.3. Chronic Phase
3. Clinical Management
4. Innovative Therapeutic Approaches
4.1. Molecular Therapy

{kind=link}
{kind=link}
| Therapeutic Agent | Molecular Target | Mechanisms of Action | Reference |
|---|---|---|---|
| Diazoxide | K-ATP Channels | Aids spinal cord ischemia-reperfusion injury by decreasing apoptosis and preserving neuronal viability and motor function. The synergistic effect with erythropoietin contributes to higher therapeutic effects. | [109,110,111,112] |
| Glibenclamide | NCCa-ATP | Acts as an NCCa-ATP antagonist, improving behavioral outcomes, decreasing lesion volume, and significantly preserving white matter. | [113] |
| Levetiracetam | SV2A | Reduces glutamate excitotoxicity, lipid peroxidation, and apoptosis, and improves astrocitic function. Significant functional and histological improvements were observed. | [115,116] |
| Riluzole | Na+ Channels | Reduces glutamate excitotoxicity and inflammatory cytokines, and induces a less active state in microglia and macrophages. Decreases cavity size and neuropathic pain, and improves motor function recovery. | [118,119,121,122,123] |
| Progesterone | PR | Increases TGFβ1 positive astrocytes and microglia cells. Attenuates axonal dieback, neuronal death, and pro-inflammatory cytokines. Contradictory results were observed. | [134,135,136,137,250] |
| Estrogen | ER-α and ER-β receptors | Attenuates the expression of several inflammasome components, apoptosis, and ROS production. Reduces glial scar formation and demyelination, and improves axonal regeneration. | [138,144,145,146] |
| Atorvastatin | HMG-CoA reductase inhibitor | Reduces oxidative stress markers, pro-inflammatory cytokines, and apoptosis. Improve motor functions and increase spare axons. Synergic treatment with and L-carnitine enhances therapeutic outcomes. No improvements were observed when tested in humans aside from a decrease in neuropathic pain. | [148,150,151,152,153,154] |
| Resveratrol | Pleiotropic interactions | Activates autophagy and inhibits apoptosis and pro-inflammatory cytokines. Reduces astrocyte activation and glial scar formation. Improves motor function and survival of motor neurons. | [161,163,164,165,166,167,168,169] |
| Omega-3 fatty acids | Pleiotropic interactions | Reduces oxidative stress, apoptosis, and inflammatory markers, and increases autophagy. Inhibits microgliosis and demyelination and increases oligodendrocytes. Locomotor recovery was observed. | [16,176,177,178,179,180,251] |
| Minocycline | Pleiotropic interactions | Improves behavior outcomes. Reduction in free radicals, lipid peroxidation, glial fibrillary, and acidic protein expression, as well as an increase in brain-derived neurotrophic factor were observed. Synergic effects with olfactory ensheathing cells graft were observed. | [185,186,187,188] |
| Il-10 | IL-10R1 and IL-10R 2 | Improves motor function, reduces lesion volume, decreases TNF-α levels and modulation of macrophages. | [252,253] |
| IL-4 | IL-4Rα | Reduction in tissue damage and inflammatory markers were observed. Favors macrophage and microglia phenotype to a pro-regenerative phenotype. Improves locomotor recovery. | [205,206,254] |
| Maresin | RORα/LGR6 | Reduces pro-inflammatory cytokines, induces pro-regenerative macrophages phenotype, improves neutrophil clearance, and reduces macrophages in the lesion site. Improves locomotor recovery. | [210,255] |
| Hypothermia | - | Reduces inflammatory cell infiltration, cell death, MPO activity, and vasogenic edema, and increases BSCB stabilization. | [214] |
| Anti-Nogo A | Nogo-A receptor | Promotes axonal regrowth and sprouting. Improves functional recovery, and reduces urodynamic abnormalities. | [220,225,226,227,228] |
| ChABC | CSPGs | Promotes functional recovery, neural plasticity, and regeneration. Synergic treatment with cell therapies. | [232,236,237,245,246,247,248,249] |
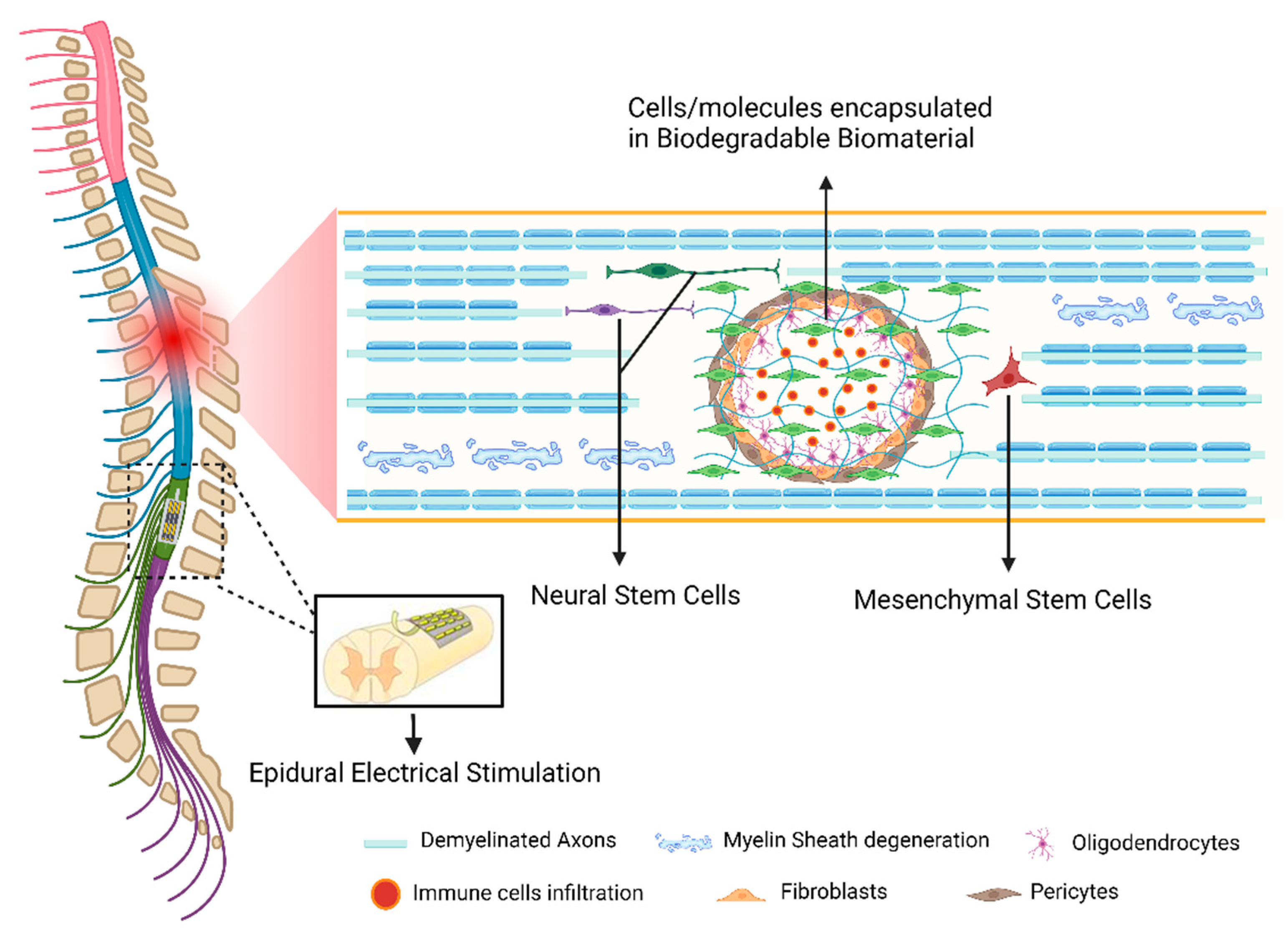
4.2. Cell-Based Strategies
4.3. Biomaterials
4.4. Novel Neurorehabilitation Protocols
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Silva, N.A.; Sousa, N.; Reis, R.L.; Salgado, A.J. From basics to clinical: A comprehensive review on spinal cord injury. Prog. Neurobiol. 2014, 114, 25–57. [Google Scholar] [CrossRef] [PubMed]
- Adigun, O.O.; Reddy, V.; Varacallo, M. Anatomy, Back, Spinal Cord; StatPearls: Treasure Island, Finland, 2022. [Google Scholar]
- Harrow-Mortelliti, M.; Reddy, V.; Jimsheleishvili, G. Physiology, Spinal Cord; StatPearls: Treasure Island, Finland, 2022. [Google Scholar]
- Mercadante, A.A.; Tadi, P. Neuroanatomy, Gray Matter; StatPearls: Treasure Island, Finland, 2022. [Google Scholar]
- Kaiser, J.T.; Lugo-Pico, J.G. Neuroanatomy, Spinal Nerves; StatPearls: Treasure Island, Finland, 2022. [Google Scholar]
- Injury, G.B.D.T.B.; Injury, C.S.C. Global, regional, and national burden of traumatic brain injury and spinal cord injury, 1990-2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Neurol. 2019, 18, 56–87. [Google Scholar]
- SCI Facts and Figures. J. Spinal Cord Med. 2017, 40, 126–127. [CrossRef] [PubMed][Green Version]
- Roberts, T.T.; Leonard, G.R.; Cepela, D.J. Classifications In Brief: American Spinal Injury Association (ASIA) Impairment Scale. Clin. Orthop. Relat. Res. 2017, 475, 1499–1504. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, A.; Dyck, S.M.; Karimi-Abdolrezaee, S. Traumatic Spinal Cord Injury: An Overview of Pathophysiology, Models and Acute Injury Mechanisms. Front. Neurol. 2019, 10, 282. [Google Scholar] [CrossRef]
- Muller-Jensen, L.; Ploner, C.J.; Kroneberg, D.; Schmidt, W.U. Clinical Presentation and Causes of Non-traumatic Spinal Cord Injury: An Observational Study in Emergency Patients. Front. Neurol. 2021, 12, 701927. [Google Scholar] [CrossRef]
- Fehlings, M. Essentials of Spinal Cord Injury: Basic Research to Clinical Practice; Thieme: New York, NY, USA, 2013. [Google Scholar]
- Hachem, L.D.; Fehlings, M.G. Pathophysiology of Spinal Cord Injury. Neurosurg. Clin. N. Am. 2021, 32, 305–313. [Google Scholar] [CrossRef]
- Hernandez-Gerez, E.; Fleming, I.N.; Parson, S.H. A role for spinal cord hypoxia in neurodegeneration. Cell Death Dis. 2019, 10, 861. [Google Scholar] [CrossRef]
- Quadri, S.A.; Farooqui, M.; Ikram, A.; Zafar, A.; Khan, M.A.; Suriya, S.S.; Claus, C.F.; Fiani, B.; Rahman, M.; Ramachandran, A.; et al. Recent update on basic mechanisms of spinal cord injury. Neurosurg. Rev. 2020, 43, 425–441. [Google Scholar] [CrossRef]
- Anwar, M.A.; Al Shehabi, T.S.; Eid, A.H. Inflammogenesis of Secondary Spinal Cord Injury. Front. Cell. Neurosci. 2016, 10, 98. [Google Scholar] [CrossRef]
- Campos, J.; Silva, N.A.; Salgado, A.J. Nutritional interventions for spinal cord injury: Preclinical efficacy and molecular mechanisms. Nutr. Rev. 2022, 80, 1206–1221. [Google Scholar] [CrossRef] [PubMed]
- Peplow, P.; Dambinova, S.A.; Gennarelli, T.A.; Martinez, B. Acute Brain Impairment: Scientific Discoveries and Translational Research; Royal Society of Chemistry: London, UK, 2018. [Google Scholar]
- Villegas, R.; Martinez, N.W.; Lillo, J.; Pihan, P.; Hernandez, D.; Twiss, J.L.; Court, F.A. Calcium release from intra-axonal endoplasmic reticulum leads to axon degeneration through mitochondrial dysfunction. J. Neurosci. 2014, 34, 7179–7189. [Google Scholar] [CrossRef] [PubMed]
- Anjum, A.; Yazid, M.D.; Daud, M.F.; Idris, J.; Ng, A.M.H.; Naicker, A.S.; Ismail, O.H.R.; Kumar, R.K.A.; Lokanathan, Y. Spinal Cord Injury: Pathophysiology, Multimolecular Interactions, and Underlying Recovery Mechanisms. Int. J. Mol. Sci. 2020, 21, 7533. [Google Scholar] [CrossRef]
- Liu, D.; Ling, X.; Wen, J.; Liu, J. The role of reactive nitrogen species in secondary spinal cord injury: Formation of nitric oxide, peroxynitrite, and nitrated protein. J. Neurochem. 2000, 75, 2144–2154. [Google Scholar] [CrossRef]
- Monteiro, S.; Pinho, A.G.; Macieira, M.; Serre-Miranda, C.; Cibrao, J.R.; Lima, R.; Soares-Cunha, C.; Vasconcelos, N.L.; Lentilhas-Graca, J.; Duarte-Silva, S.; et al. Splenic sympathetic signaling contributes to acute neutrophil infiltration of the injured spinal cord. J. Neuroinflamm. 2020, 17, 282. [Google Scholar] [CrossRef]
- Beck, K.D.; Nguyen, H.X.; Galvan, M.D.; Salazar, D.L.; Woodruff, T.M.; Anderson, A.J. Quantitative analysis of cellular inflammation after traumatic spinal cord injury: Evidence for a multiphasic inflammatory response in the acute to chronic environment. Brain 2010, 133 Pt 2, 433–447. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Xia, Y.; Li, B.; Li, Y.; Fu, C. Reverse Adverse Immune Microenvironments by Biomaterials Enhance the Repair of Spinal Cord Injury. Front. Bioeng. Biotechnol. 2022, 10, 812340. [Google Scholar] [CrossRef]
- Orr, M.B.; Gensel, J.C. Spinal Cord Injury Scarring and Inflammation: Therapies Targeting Glial and Inflammatory Responses. Neurotherapeutics 2018, 15, 541–553. [Google Scholar] [CrossRef]
- Wu, X.; Yan, Y.; Zhang, Q. Neuroinflammation and Modulation Role of Natural Products After Spinal Cord Injury. J. Inflamm. Res. 2021, 14, 5713–5737. [Google Scholar] [CrossRef]
- Kigerl, K.A.; de Rivero Vaccari, J.P.; Dietrich, W.D.; Popovich, P.G.; Keane, R.W. Pattern recognition receptors and central nervous system repair. Exp. Neurol. 2014, 258, 5–16. [Google Scholar] [CrossRef]
- Hellenbrand, D.J.; Quinn, C.M.; Piper, Z.J.; Morehouse, C.N.; Fixel, J.A.; Hanna, A.S. Inflammation after spinal cord injury: A review of the critical timeline of signaling cues and cellular infiltration. J. Neuroinflamm. 2021, 18, 284. [Google Scholar] [CrossRef] [PubMed]
- David, S.; Kroner, A. Repertoire of microglial and macrophage responses after spinal cord injury. Nat. Rev. Neurosci. 2011, 12, 388–399. [Google Scholar] [CrossRef] [PubMed]
- Hao, J.; Li, B.; Duan, H.Q.; Zhao, C.X.; Zhang, Y.; Sun, C.; Pan, B.; Liu, C.; Kong, X.H.; Yao, X.; et al. Mechanisms underlying the promotion of functional recovery by deferoxamine after spinal cord injury in rats. Neural. Regen. Res. 2017, 12, 959–968. [Google Scholar] [PubMed]
- Zivkovic, S.; Ayazi, M.; Hammel, G.; Ren, Y. For Better or for Worse: A Look Into Neutrophils in Traumatic Spinal Cord Injury. Front. Cell. Neurosci. 2021, 15, 648076. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, T.C.; Mathews, K.J.; Mao, Y.; Nguyen, T.; Gorrie, C.A. Differences in the Cellular Response to Acute Spinal Cord Injury between Developing and Mature Rats Highlights the Potential Significance of the Inflammatory Response. Front. Cell. Neurosci. 2016, 10, 310. [Google Scholar] [CrossRef]
- Soderblom, C.; Lee, D.H.; Dawood, A.; Carballosa, M.; Santamaria, A.J.; Benavides, F.D.; Jergova, S.; Grumbles, R.M.; Thomas, C.K.; Park, K.K.; et al. 3D Imaging of Axons in Transparent Spinal Cords from Rodents and Nonhuman Primates. eNeuro 2015, 2, ENEURO.0001-15.2015. [Google Scholar] [CrossRef]
- Fleming, J.C.; Norenberg, M.D.; Ramsay, D.A.; Dekaban, G.A.; Marcillo, A.E.; Saenz, A.D.; Pasquale-Styles, M.; Dietrich, W.D.; Weaver, L.C. The cellular inflammatory response in human spinal cords after injury. Brain 2006, 129 Pt 12, 3249–3269. [Google Scholar] [CrossRef]
- Donnelly, D.J.; Popovich, P.G. Inflammation and its role in neuroprotection, axonal regeneration and functional recovery after spinal cord injury. Exp. Neurol. 2008, 209, 378–388. [Google Scholar] [CrossRef]
- Gensel, J.C.; Zhang, B. Macrophage activation and its role in repair and pathology after spinal cord injury. Brain Res. 2015, 1619, 1–11. [Google Scholar] [CrossRef]
- Li, C.; Wu, Z.; Zhou, L.; Shao, J.; Hu, X.; Xu, W.; Ren, Y.; Zhu, X.; Ge, W.; Zhang, K.; et al. Temporal and spatial cellular and molecular pathological alterations with single-cell resolution in the adult spinal cord after injury. Signal. Transduct. Target Ther. 2022, 7, 65. [Google Scholar] [CrossRef]
- Bollaerts, I.; Van Houcke, J.; Andries, L.; De Groef, L.; Moons, L. Neuroinflammation as Fuel for Axonal Regeneration in the Injured Vertebrate Central Nervous System. Mediat. Inflamm. 2017, 2017, 9478542. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, S.; Salgado, A.J.; Silva, N.A. Immunomodulation as a neuroprotective strategy after spinal cord injury. Neural. Regen. Res. 2018, 13, 423–424. [Google Scholar] [PubMed]
- Hill, C.E. A view from the ending: Axonal dieback and regeneration following SCI. Neurosci. Lett. 2017, 652, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Chelyshev, Y.A.; Kabdesh, I.M.; Mukhamedshina, Y.O. Extracellular Matrix in Neural Plasticity and Regeneration. Cell. Mol. Neurobiol. 2022, 42, 647–664. [Google Scholar] [CrossRef]
- Wiese, S.; Faissner, A. The role of extracellular matrix in spinal cord development. Exp. Neurol. 2015, 274 Pt B, 90–99. [Google Scholar] [CrossRef]
- Lang, B.T.; Cregg, J.M.; DePaul, M.A.; Tran, A.P.; Xu, K.; Dyck, S.M.; Madalena, K.M.; Brown, B.P.; Weng, Y.L.; Li, S.; et al. Modulation of the proteoglycan receptor PTPsigma promotes recovery after spinal cord injury. Nature 2015, 518, 404–408. [Google Scholar] [CrossRef]
- Song, I.; Dityatev, A. Crosstalk between glia, extracellular matrix and neurons. Brain Res. Bull. 2018, 136, 101–108. [Google Scholar] [CrossRef]
- Kerschensteiner, M.; Schwab, M.E.; Lichtman, J.W.; Misgeld, T. In vivo imaging of axonal degeneration and regeneration in the injured spinal cord. Nat. Med. 2005, 11, 572–577. [Google Scholar] [CrossRef]
- Busch, S.A.; Horn, K.P.; Silver, D.J.; Silver, J. Overcoming macrophage-mediated axonal dieback following CNS injury. J. Neurosci. 2009, 29, 9967–9976. [Google Scholar] [CrossRef]
- Fan, B.; Wei, Z.; Yao, X.; Shi, G.; Cheng, X.; Zhou, X.; Zhou, H.; Ning, G.; Kong, X.; Feng, S. Microenvironment Imbalance of Spinal Cord Injury. Cell Transplant. 2018, 27, 853–866. [Google Scholar] [CrossRef]
- Boghdadi, A.G.; Teo, L.; Bourne, J.A. The Involvement of the Myelin-Associated Inhibitors and Their Receptors in CNS Plasticity and Injury. Mol. Neurobiol. 2018, 55, 1831–1846. [Google Scholar] [CrossRef] [PubMed]
- Caroni, P.; Savio, T.; Schwab, M.E. Central nervous system regeneration: Oligodendrocytes and myelin as non-permissive substrates for neurite growth. Prog. Brain Res. 1988, 78, 363–370. [Google Scholar] [PubMed]
- Loschinger, J.; Bandtlow, C.E.; Jung, J.; Klostermann, S.; Schwab, M.E.; Bonhoeffer, F.; Kater, S.B. Retinal axon growth cone responses to different environmental cues are mediated by different second-messenger systems. J. Neurobiol. 1997, 33, 825–834. [Google Scholar] [CrossRef]
- Bandtlow, C.E.; Schmidt, M.F.; Hassinger, T.D.; Schwab, M.E.; Kater, S.B. Role of intracellular calcium in NI-35-evoked collapse of neuronal growth cones. Science 1993, 259, 80–83. [Google Scholar] [CrossRef] [PubMed]
- Habib, A.A.; Marton, L.S.; Allwardt, B.; Gulcher, J.R.; Mikol, D.D.; Hognason, T.; Chattopadhyay, N.; Stefansson, K. Expression of the oligodendrocyte-myelin glycoprotein by neurons in the mouse central nervous system. J. Neurochem. 1998, 70, 1704–1711. [Google Scholar] [CrossRef]
- Kottis, V.; Thibault, P.; Mikol, D.; Xiao, Z.C.; Zhang, R.; Dergham, P.; Braun, P.E. Oligodendrocyte-myelin glycoprotein (OMgp) is an inhibitor of neurite outgrowth. J. Neurochem. 2002, 82, 1566–1569. [Google Scholar] [CrossRef]
- Huang, J.K.; Phillips, G.R.; Roth, A.D.; Pedraza, L.; Shan, W.; Belkaid, W.; Mi, S.; Fex-Svenningsen, A.; Florens, L.; Yates, J.R., 3rd; et al. Glial membranes at the node of Ranvier prevent neurite outgrowth. Science 2005, 310, 1813–1817. [Google Scholar] [CrossRef]
- Fernandez-Suarez, D.; Krapacher, F.A.; Andersson, A.; Ibanez, C.F.; Kisiswa, L. MAG induces apoptosis in cerebellar granule neurons through p75(NTR) demarcating granule layer/white matter boundary. Cell Death Dis. 2019, 10, 732. [Google Scholar] [CrossRef]
- Shibata, A.; Wright, M.V.; David, S.; McKerracher, L.; Braun, P.E.; Kater, S.B. Unique responses of differentiating neuronal growth cones to inhibitory cues presented by oligodendrocytes. J. Cell Biol. 1998, 142, 191–202. [Google Scholar] [CrossRef]
- McKerracher, L.; David, S.; Jackson, D.L.; Kottis, V.; Dunn, R.J.; Braun, P.E. Identification of myelin-associated glycoprotein as a major myelin-derived inhibitor of neurite growth. Neuron 1994, 13, 805–811. [Google Scholar] [CrossRef]
- Tang, S.; Qiu, J.; Nikulina, E.; Filbin, M.T. Soluble myelin-associated glycoprotein released from damaged white matter inhibits axonal regeneration. Mol. Cell. Neurosci. 2001, 18, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.; Woodhall, R.W.; Shen, Y.J.; deBellard, M.E.; Saffell, J.L.; Doherty, P.; Walsh, F.S.; Filbin, M.T. Soluble myelin-associated glycoprotein (MAG) found in vivo inhibits axonal regeneration. Mol. Cell. Neurosci. 1997, 9, 333–346. [Google Scholar] [CrossRef] [PubMed]
- Mueller-Buehl, C.; Reinhard, J.; Roll, L.; Bader, V.; Winklhofer, K.F.; Faissner, A. Brevican, Neurocan, Tenascin-C, and Tenascin-R Act as Important Regulators of the Interplay Between Perineuronal Nets, Synaptic Integrity, Inhibitory Interneurons, and Otx2. Front. Cell Dev. Biol. 2022, 10, 886527. [Google Scholar] [CrossRef] [PubMed]
- de Vivo, L.; Landi, S.; Panniello, M.; Baroncelli, L.; Chierzi, S.; Mariotti, L.; Spolidoro, M.; Pizzorusso, T.; Maffei, L.; Ratto, G.M. Extracellular matrix inhibits structural and functional plasticity of dendritic spines in the adult visual cortex. Nat. Commun. 2013, 4, 1484. [Google Scholar] [CrossRef]
- Deepa, S.S.; Umehara, Y.; Higashiyama, S.; Itoh, N.; Sugahara, K. Specific molecular interactions of oversulfated chondroitin sulfate E with various heparin-binding growth factors. Implications as a physiological binding partner in the brain and other tissues. J. Biol. Chem. 2002, 277, 43707–43716. [Google Scholar] [CrossRef]
- Frischknecht, R.; Heine, M.; Perrais, D.; Seidenbecher, C.I.; Choquet, D.; Gundelfinger, E.D. Brain extracellular matrix affects AMPA receptor lateral mobility and short-term synaptic plasticity. Nat. Neurosci. 2009, 12, 897–904. [Google Scholar] [CrossRef]
- Pan, D.; Li, Y.; Yang, F.; Lv, Z.; Zhu, S.; Shao, Y.; Huang, Y.; Ning, G.; Feng, S. Increasing toll-like receptor 2 on astrocytes induced by Schwann cell-derived exosomes promotes recovery by inhibiting CSPGs deposition after spinal cord injury. J. Neuroinflamm. 2021, 18, 172. [Google Scholar] [CrossRef]
- Asher, R.A.; Morgenstern, D.A.; Shearer, M.C.; Adcock, K.H.; Pesheva, P.; Fawcett, J.W. Versican is upregulated in CNS injury and is a product of oligodendrocyte lineage cells. J. Neurosci. 2002, 22, 2225–2236. [Google Scholar] [CrossRef]
- Uhlin-Hansen, L.; Wik, T.; Kjellen, L.; Berg, E.; Forsdahl, F.; Kolset, S.O. Proteoglycan metabolism in normal and inflammatory human macrophages. Blood 1993, 82, 2880–2889. [Google Scholar] [CrossRef]
- Jones, F.S.; Jones, P.L. The tenascin family of ECM glycoproteins: Structure, function, and regulation during embryonic development and tissue remodeling. Dev. Dyn. 2000, 218, 235–259. [Google Scholar] [CrossRef]
- Monnier, P.P.; Sierra, A.; Schwab, J.M.; Henke-Fahle, S.; Mueller, B.K. The Rho/ROCK pathway mediates neurite growth-inhibitory activity associated with the chondroitin sulfate proteoglycans of the CNS glial scar. Mol. Cell. Neurosci. 2003, 22, 319–330. [Google Scholar] [CrossRef]
- Shen, Y.; Tenney, A.P.; Busch, S.A.; Horn, K.P.; Cuascut, F.X.; Liu, K.; He, Z.; Silver, J.; Flanagan, J.G. PTPsigma is a receptor for chondroitin sulfate proteoglycan, an inhibitor of neural regeneration. Science 2009, 326, 592–596. [Google Scholar] [CrossRef]
- Kimura-Kuroda, J.; Teng, X.; Komuta, Y.; Yoshioka, N.; Sango, K.; Kawamura, K.; Raisman, G.; Kawano, H. An in vitro model of the inhibition of axon growth in the lesion scar formed after central nervous system injury. Mol. Cell. Neurosci. 2010, 43, 177–187. [Google Scholar] [CrossRef]
- Logan, A.; Berry, M.; Gonzalez, A.M.; Frautschy, S.A.; Sporn, M.B.; Baird, A. Effects of transforming growth factor beta 1 on scar production in the injured central nervous system of the rat. Eur. J. Neurosci. 1994, 6, 355–363. [Google Scholar] [CrossRef] [PubMed]
- East, E.; Golding, J.P.; Phillips, J.B. A versatile 3D culture model facilitates monitoring of astrocytes undergoing reactive gliosis. J. Tissue Eng. Regen. Med. 2009, 3, 634–646. [Google Scholar] [CrossRef] [PubMed]
- Renault-Mihara, F.; Mukaino, M.; Shinozaki, M.; Kumamaru, H.; Kawase, S.; Baudoux, M.; Ishibashi, T.; Kawabata, S.; Nishiyama, Y.; Sugai, K.; et al. Regulation of RhoA by STAT3 coordinates glial scar formation. J. Cell Biol. 2017, 216, 2533–2550. [Google Scholar] [CrossRef]
- Bott, K.; Upton, Z.; Schrobback, K.; Ehrbar, M.; Hubbell, J.A.; Lutolf, M.P.; Rizzi, S.C. The effect of matrix characteristics on fibroblast proliferation in 3D gels. Biomaterials 2010, 31, 8454–8464. [Google Scholar] [CrossRef]
- Tran, A.P.; Warren, P.M.; Silver, J. The Biology of Regeneration Failure and Success After Spinal Cord Injury. Physiol. Rev. 2018, 98, 881–917. [Google Scholar] [CrossRef]
- Li, X.; Yang, B.; Xiao, Z.; Zhao, Y.; Han, S.; Yin, Y.; Chen, B.; Dai, J. Comparison of subacute and chronic scar tissues after complete spinal cord transection. Exp. Neurol. 2018, 306, 132–137. [Google Scholar] [CrossRef]
- Weidner, N.; Grill, R.J.; Tuszynski, M.H. Elimination of basal lamina and the collagen "scar" after spinal cord injury fails to augment corticospinal tract regeneration. Exp. Neurol. 1999, 160, 40–50. [Google Scholar] [CrossRef]
- Gonzalez-Perez, F.; Udina, E.; Navarro, X. Extracellular matrix components in peripheral nerve regeneration. Int. Rev. Neurobiol. 2013, 108, 257–275. [Google Scholar] [PubMed]
- Anderson, M.A.; Burda, J.E.; Ren, Y.; Ao, Y.; O’Shea, T.M.; Kawaguchi, R.; Coppola, G.; Khakh, B.S.; Deming, T.J.; Sofroniew, M.V. Astrocyte scar formation aids central nervous system axon regeneration. Nature 2016, 532, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Faulkner, J.R.; Herrmann, J.E.; Woo, M.J.; Tansey, K.E.; Doan, N.B.; Sofroniew, M.V. Reactive astrocytes protect tissue and preserve function after spinal cord injury. J. Neurosci. 2004, 24, 2143–2155. [Google Scholar] [CrossRef] [PubMed]
- Hemati-Gourabi, M.; Cao, T.; Romprey, M.K.; Chen, M. Capacity of astrocytes to promote axon growth in the injured mammalian central nervous system. Front. Neurosci. 2022, 16, 955598. [Google Scholar] [CrossRef]
- Bloom, O. Non-mammalian model systems for studying neuro-immune interactions after spinal cord injury. Exp. Neurol. 2014, 258, 130–140. [Google Scholar] [CrossRef]
- Zukor, K.A.; Kent, D.T.; Odelberg, S.J. Meningeal cells and glia establish a permissive environment for axon regeneration after spinal cord injury in newts. Neural. Dev. 2011, 6, 1. [Google Scholar] [CrossRef]
- Hara, M.; Kobayakawa, K.; Ohkawa, Y.; Kumamaru, H.; Yokota, K.; Saito, T.; Kijima, K.; Yoshizaki, S.; Harimaya, K.; Nakashima, Y.; et al. Interaction of reactive astrocytes with type I collagen induces astrocytic scar formation through the integrin-N-cadherin pathway after spinal cord injury. Nat. Med. 2017, 23, 818–828. [Google Scholar] [CrossRef]
- Rowland, J.W.; Hawryluk, G.W.; Kwon, B.; Fehlings, M.G. Current status of acute spinal cord injury pathophysiology and emerging therapies: Promise on the horizon. Neurosurg. Focus 2008, 25, E2. [Google Scholar] [CrossRef]
- Ahuja, C.S.; Wilson, J.R.; Nori, S.; Kotter, M.R.N.; Druschel, C.; Curt, A.; Fehlings, M.G. Traumatic spinal cord injury. Nat. Rev. Dis. Primers 2017, 3, 17018. [Google Scholar] [CrossRef]
- Ehlers, M.D. Deconstructing the axon: Wallerian degeneration and the ubiquitin-proteasome system. Trends Neurosci. 2004, 27, 3–6. [Google Scholar] [CrossRef]
- Sadek, A.R.; Nader-Sepahi, A. Spinal Arachnoid Cysts: Presentation, management and pathophysiology. Clin. Neurol. Neurosurg. 2019, 180, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Kwiecien, J.M.; Dabrowski, W.; Yaron, J.R.; Zhang, L.; Delaney, K.H.; Lucas, A.R. The Role of Astrogliosis in Formation of the Syrinx in Spinal Cord Injury. Curr. Neuropharmacol. 2021, 19, 294–303. [Google Scholar] [CrossRef] [PubMed]
- Scivoletto, G.; Masciullo, M.; Pichiorri, F.; Molinari, M. Silent post-traumatic syringomyelia and syringobulbia. Spinal Cord Ser. Cases 2020, 6, 15. [Google Scholar] [CrossRef]
- Lu, Z.; Fu, L.; Fan, X. Spinal cord stimulation for treatment of neuropathic pain associated with syringomyelia. Asian J. Surg. 2022. Online ahead of print. [Google Scholar] [CrossRef]
- Fehlings, M.G.; Vaccaro, A.; Wilson, J.R.; Singh, A.; D, W.C.; Harrop, J.S.; Aarabi, B.; Shaffrey, C.; Dvorak, M.; Fisher, C.; et al. Early versus delayed decompression for traumatic cervical spinal cord injury: Results of the Surgical Timing in Acute Spinal Cord Injury Study (STASCIS). PLoS ONE 2012, 7, e32037. [Google Scholar] [CrossRef]
- Wilson, J.R.; Tetreault, L.A.; Kwon, B.K.; Arnold, P.M.; Mroz, T.E.; Shaffrey, C.; Harrop, J.S.; Chapman, J.R.; Casha, S.; Skelly, A.C.; et al. Timing of Decompression in Patients With Acute Spinal Cord Injury: A Systematic Review. Glob. Spine J. 2017, 7 (Suppl. S3), 95S–115S. [Google Scholar] [CrossRef]
- Yousefifard, M.; Rahimi-Movaghar, V.; Baikpour, M.; Ghelichkhani, P.; Hosseini, M.; Jafari, A.; Aziznejad, H.; Tafakhori, A. Early versus late spinal decompression surgery in treatment of traumatic spinal cord injuries; a systematic review and meta-analysis. Emergency 2017, 5, e37. [Google Scholar] [PubMed]
- Fehlings, M.G.; Tetreault, L.A.; Wilson, J.R.; Aarabi, B.; Anderson, P.; Arnold, P.M.; Brodke, D.S.; Burns, A.S.; Chiba, K.; Dettori, J.R.; et al. A Clinical Practice Guideline for the Management of Patients With Acute Spinal Cord Injury and Central Cord Syndrome: Recommendations on the Timing (≤24 Hours Versus >24 Hours) of Decompressive Surgery. Glob. Spine J. 2017, 7 (Suppl. S3), 195S–202S. [Google Scholar] [CrossRef]
- Levi, L.; Wolf, A.; Belzberg, H. Hemodynamic parameters in patients with acute cervical cord trauma: Description, intervention, and prediction of outcome. Neurosurgery 1993, 33, 1007–1016, discussion 1016–7. [Google Scholar] [CrossRef]
- Yue, J.K.; Tsolinas, R.E.; Burke, J.F.; Deng, H.; Upadhyayula, P.S.; Robinson, C.K.; Lee, Y.M.; Chan, A.K.; Winkler, E.A.; Dhall, S.S. Vasopressor support in managing acute spinal cord injury: Current knowledge. J. Neurosurg. Sci. 2019, 63, 308–317. [Google Scholar] [CrossRef]
- Hadley, M.N.; Walters, B.C.; Grabb, P.A.; Oyesiku, N.M.; Przybylski, G.J.; Resnick, D.K.; Ryken, T.C. Blood pressure management after acute spinal cord injury. Neurosurgery 2002, 50 (Suppl. S3), S58–S62. [Google Scholar] [PubMed]
- Arnold, P.M.; Harrop, J.S.; Merli, G.; Tetreault, L.G.; Kwon, B.K.; Casha, S.; Palmieri, K.; Wilson, J.R.; Fehlings, M.G.; Holmer, H.K.; et al. Efficacy, Safety, and Timing of Anticoagulant Thromboprophylaxis for the Prevention of Venous Thromboembolism in Patients With Acute Spinal Cord Injury: A Systematic Review. Glob. Spine J. 2017, 7 (Suppl. S3), 138S–150S. [Google Scholar] [CrossRef] [PubMed]
- Bartholdi, D.; Schwab, M.E. Methylprednisolone inhibits early inflammatory processes but not ischemic cell death after experimental spinal cord lesion in the rat. Brain Res. 1995, 672, 177–186. [Google Scholar] [CrossRef]
- Ahuja, C.S.; Schroeder, G.D.; Vaccaro, A.R.; Fehlings, M.G. Spinal Cord Injury-What Are the Controversies? J. Orthop. Trauma 2017, 31 (Suppl. S4), S7–S13. [Google Scholar] [CrossRef]
- Bracken, M.B.; Shepard, M.J.; Collins, W.F.; Holford, T.R.; Young, W.; Baskin, D.S.; Eisenberg, H.M.; Flamm, E.; Leo-Summers, L.; Maroon, J.; et al. A randomized, controlled trial of methylprednisolone or naloxone in the treatment of acute spinal-cord injury. Results of the Second National Acute Spinal Cord Injury Study. N. Engl. J. Med. 1990, 322, 1405–1411. [Google Scholar] [CrossRef]
- Bracken, M.B.; Shepard, M.J.; Holford, T.R.; Leo-Summers, L.; Aldrich, E.F.; Fazl, M.; Fehlings, M.; Herr, D.L.; Hitchon, P.W.; Marshall, L.F.; et al. Administration of methylprednisolone for 24 or 48 hours or tirilazad mesylate for 48 hours in the treatment of acute spinal cord injury. Results of the Third National Acute Spinal Cord Injury Randomized Controlled Trial. National Acute Spinal Cord Injury Study. JAMA 1997, 277, 1597–1604. [Google Scholar]
- Bracken, M.B.; Shepard, M.J.; Hellenbrand, K.G.; Collins, W.F.; Leo, L.S.; Freeman, D.F.; Wagner, F.C.; Flamm, E.S.; Eisenberg, H.M.; Goodman, J.H.; et al. Methylprednisolone and neurological function 1 year after spinal cord injury. Results of the National Acute Spinal Cord Injury Study. J. Neurosurg. 1985, 63, 704–713. [Google Scholar] [CrossRef]
- Bracken, M.B.; Holford, T.R. Effects of timing of methylprednisolone or naloxone administration on recovery of segmental and long-tract neurological function in NASCIS 2. J. Neurosurg. 1993, 79, 500–507. [Google Scholar] [CrossRef]
- Ito, Y.; Sugimoto, Y.; Tomioka, M.; Kai, N.; Tanaka, M. Does high dose methylprednisolone sodium succinate really improve neurological status in patient with acute cervical cord injury?: A prospective study about neurological recovery and early complications. Spine (Phila Pa 1976) 2009, 34, 2121–2124. [Google Scholar] [CrossRef]
- Walters, B.C.; Hadley, M.N.; Hurlbert, R.J.; Aarabi, B.; Dhall, S.S.; Gelb, D.E.; Harrigan, M.R.; Rozelle, C.J.; Ryken, T.C.; Theodore, N.; et al. Guidelines for the management of acute cervical spine and spinal cord injuries: 2013 update. Neurosurgery 2013, 60, (CN_suppl_1). 82–91. [Google Scholar] [CrossRef]
- Hawryluk, G.W.; Rowland, J.; Kwon, B.K.; Fehlings, M.G. Protection and repair of the injured spinal cord: A review of completed, ongoing, and planned clinical trials for acute spinal cord injury. Neurosurg. Focus 2008, 25, E14. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Feng, L.; Yao, H.; Yang, L.; Qin, Y. Efficacy and safety of diazoxide for treating hyperinsulinemic hypoglycemia: A systematic review and meta-analysis. PLoS ONE 2021, 16, e0246463. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, K.; Eldeiry, M.; Aftab, M.; Ryan, T.J.; Roda, G.; Meng, X.; Weyant, M.J.; Cleveland, J.C., Jr.; Fullerton, D.A.; Reece, T.B. Pretreatment With Diazoxide Attenuates Spinal Cord Ischemia-Reperfusion Injury Through Signaling Transducer and Activator of Transcription 3 Pathway. Ann. Thorac. Surg. 2019, 107, 733–739. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, K.; Eldeiry, M.; Aftab, M.; Mares, J.; Ryan, T.J.; Meng, X.; Weyant, M.J.; Cleveland, J.C., Jr.; Fullerton, D.A.; Reece, T.B. Optimized induction of beta common receptor enhances the neuroprotective function of erythropoietin in spinal cord ischemic injury. J. Thorac. Cardiovasc. Surg. 2018, 155, 2505–2516. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, K.; Eldeiry, M.; Aftab, M.; Ryan, T.J.; Meng, X.; Weyant, M.J.; Fullerton, D.A.; Reece, T.B. Synergetic Induction of NGF with Diazoxide and Erythropoietin Attenuates Spinal Cord Ischemic Injury. J. Surg. Res. 2019, 233, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, K.; Eldeiry, M.; Aftab, M.; Ryan, T.J.; Mares, J.; Meng, X.; Weyant, M.J.; Cleveland, J.C., Jr.; Fullerton, D.A.; Reece, T.B. Synergistic Reduction of Apoptosis with Diazoxide and Erythropoietin in Spinal Cord Ischemic Injury. Ann. Thorac. Surg. 2018, 106, 1751–1758. [Google Scholar] [CrossRef] [PubMed]
- Simard, J.M.; Tsymbalyuk, O.; Ivanov, A.; Ivanova, S.; Bhatta, S.; Geng, Z.; Woo, S.K.; Gerzanich, V. Endothelial sulfonylurea receptor 1-regulated NC Ca-ATP channels mediate progressive hemorrhagic necrosis following spinal cord injury. J. Clin. Investig. 2007, 117, 2105–2113. [Google Scholar] [CrossRef]
- Minnema, A.J.; Mehta, A.; Boling, W.W.; Schwab, J.; Simard, J.M.; Farhadi, H.F. SCING-Spinal Cord Injury Neuroprotection with Glyburide: A pilot, open-label, multicentre, prospective evaluation of oral glyburide in patients with acute traumatic spinal cord injury in the USA. BMJ Open 2019, 9, e031329. [Google Scholar] [CrossRef]
- Lima, R.; Gomes, E.D.; Cibrao, J.R.; Rocha, L.A.; Assuncao-Silva, R.C.; Rodrigues, C.S.; Neves-Carvalho, A.; Monteiro, S.; Salgado, A.J.; Silva, N.A. Levetiracetam treatment leads to functional recovery after thoracic or cervical injuries of the spinal cord. NPJ Regen. Med. 2021, 6, 11. [Google Scholar] [CrossRef]
- Xia, H.; Huang, J.; Liu, J.; Zhang, H.; Chen, D.; Zhang, W.; Zhou, X.; Wei, Z.; Liu, Z. Levetiracetam Attenuates the Spinal Cord Injury Induced by Acute Trauma via Suppressing the Expression of Perforin. Comput. Math. Methods Med. 2022, 2022, 7218666. [Google Scholar] [CrossRef]
- Bellingham, M.C. A review of the neural mechanisms of action and clinical efficiency of riluzole in treating amyotrophic lateral sclerosis: What have we learned in the last decade? CNS Neurosci. Ther. 2011, 17, 4–31. [Google Scholar] [CrossRef] [PubMed]
- Tetreault, L.A.; Zhu, M.P.; Wilson, J.R.; Karadimas, S.K.; Fehlings, M.G. The Impact of Riluzole on Neurobehavioral Outcomes in Preclinical Models of Traumatic and Nontraumatic Spinal Cord Injury: Results From a Systematic Review of the Literature. Glob. Spine J. 2020, 10, 216–229. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Zhang, Y.; Zhang, Y.; Zhang, W.; Zhang, W.; Liu, Y.; Xu, S.; Guan, Y.; Chen, X. Riluzole improves functional recovery after acute spinal cord injury in rats and may be associated with changes in spinal microglia/macrophages polarization. Neurosci. Lett. 2020, 723, 134829. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Wu, Q.; Zhang, W.; Liu, T.; Zhang, Y.; Zhang, W.; Zhang, Y.; Chen, X. Riluzole Promotes Neurite Growth in Rats after Spinal Cord Injury through the GSK-3beta/CRMP-2 Pathway. Biol. Pharm. Bull. 2022, 45, 569–575. [Google Scholar] [CrossRef]
- Meshkini, A.; Salehpour, F.; Aghazadeh, J.; Mirzaei, F.; Naseri Alavi, S.A. Riluzole Can Improve Sensory and Motor Function in Patients with Acute Spinal Cord Injury. Asian J. Neurosurg. 2018, 13, 656–659. [Google Scholar] [CrossRef]
- Zhou, L.Y.; Tian, Z.R.; Yao, M.; Chen, X.Q.; Song, Y.J.; Ye, J.; Yi, N.X.; Cui, X.J.; Wang, Y.J. Riluzole promotes neurological function recovery and inhibits damage extension in rats following spinal cord injury: A meta-analysis and systematic review. J. Neurochem. 2019, 150, 6–27. [Google Scholar] [CrossRef]
- Srinivas, S.; Wali, A.R.; Pham, M.H. Efficacy of riluzole in the treatment of spinal cord injury: A systematic review of the literature. Neurosurg. Focus 2019, 46, E6. [Google Scholar] [CrossRef]
- Grossman, R.G.; Fehlings, M.G.; Frankowski, R.F.; Burau, K.D.; Chow, D.S.; Tator, C.; Teng, A.; Toups, E.G.; Harrop, J.S.; Aarabi, B.; et al. A prospective, multicenter, phase I matched-comparison group trial of safety, pharmacokinetics, and preliminary efficacy of riluzole in patients with traumatic spinal cord injury. J. Neurotrauma. 2014, 31, 239–255. [Google Scholar] [CrossRef]
- Fehlings, M.G.; Nakashima, H.; Nagoshi, N.; Chow, D.S.; Grossman, R.G.; Kopjar, B. Rationale, design and critical end points for the Riluzole in Acute Spinal Cord Injury Study (RISCIS): A randomized, double-blinded, placebo-controlled parallel multi-center trial. Spinal Cord 2016, 54, 8–15. [Google Scholar] [CrossRef]
- Nguyen, A.; Chow, D.S.; Wu, L.; Teng, Y.A.; Sarkar, M.; Toups, E.G.; Harrop, J.S.; Schmitt, K.M.; Johnson, M.M.; Guest, J.D.; et al. Longitudinal Impact of Acute Spinal Cord Injury on Clinical Pharmacokinetics of Riluzole, a Potential Neuroprotective Agent. J. Clin. Pharmacol. 2021, 61, 1232–1242. [Google Scholar] [CrossRef]
- Fehlings, M.G.; Kopjar, B.; Grossman, R.G. 100-Efficacy and Safety of Riluzole in Acute Spinal Cord Injury (SCI). Rationale and Design of AOSpine Phase III Multicenter Double-Blinded Randomized Controlled Trial (RISCIS). Spine J. 2017, 17 (Suppl. S10), S131. [Google Scholar] [CrossRef]
- Baulieu, E.E.; Schumacher, M. Progesterone as a neuroactive neurosteroid, with special reference to the effect of progesterone on myelination. Hum. Reprod. 2000, 15 (Suppl. S1), 1–13. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kapur, J.; Joshi, S. Progesterone modulates neuronal excitability bidirectionally. Neurosci. Lett. 2021, 744, 135619. [Google Scholar] [CrossRef] [PubMed]
- Nagy, B.; Szekeres-Bartho, J.; Kovacs, G.L.; Sulyok, E.; Farkas, B.; Varnagy, A.; Vertes, V.; Kovacs, K.; Bodis, J. Key to Life: Physiological Role and Clinical Implications of Progesterone. Int. J. Mol. Sci. 2021, 22, 11039. [Google Scholar] [CrossRef]
- Ghoumari, A.M.; Abi Ghanem, C.; Asbelaoui, N.; Schumacher, M.; Hussain, R. Roles of Progesterone, Testosterone and Their Nuclear Receptors in Central Nervous System Myelination and Remyelination. Int. J. Mol. Sci. 2020, 21, 3163. [Google Scholar] [CrossRef]
- Liu, C.; Gao, W.; Zhao, L.; Cao, Y. Progesterone attenuates neurological deficits and exerts a protective effect on damaged axons via the PI3K/AKT/mTOR-dependent pathway in a mouse model of intracerebral hemorrhage. Aging 2022, 14, 2574–2589. [Google Scholar] [CrossRef]
- Ludwig, P.E.; Patil, A.A.; Chamczuk, A.J.; Agrawal, D.K. Hormonal therapy in traumatic spinal cord injury. Am. J. Transl. Res. 2017, 9, 3881–3895. [Google Scholar]
- Coronel, M.F.; Villar, M.J.; Brumovsky, P.R.; Gonzalez, S.L. Spinal neuropeptide expression and neuropathic behavior in the acute and chronic phases after spinal cord injury: Effects of progesterone administration. Peptides 2017, 88, 189–195. [Google Scholar] [CrossRef]
- Jure, I.; De Nicola, A.F.; Labombarda, F. Progesterone effects on oligodendrocyte differentiation in injured spinal cord. Brain Res. 2019, 1708, 36–46. [Google Scholar] [CrossRef]
- Yang, Z.; Xie, W.; Ju, F.; Khan, A.; Zhang, S. In vivo two-photon imaging reveals a role of progesterone in reducing axonal dieback after spinal cord injury in mice. Neuropharmacology 2017, 116, 30–37. [Google Scholar] [CrossRef]
- Cavalcante, L.P.; Ferreira, S.G.; Pereira, D.R.; Moraes, S.R.; Simas, R.; Sannomiya, P.; Breithaupt-Faloppa, A.C.; Moreira, L.F.P. Acute administration of oestradiol or progesterone in a spinal cord ischaemia-reperfusion model in rats. Interact. Cardiovasc. Thorac. Surg. 2018, 26, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Johann, S.; Beyer, C. Neuroprotection by gonadal steroid hormones in acute brain damage requires cooperation with astroglia and microglia. J. Steroid. Biochem. Mol. Biol. 2013, 137, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Behl, C.; Manthey, D. Neuroprotective activities of estrogen: An update. J. Neurocytol. 2000, 29, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Woolley, C.S.; McEwen, B.S. Roles of estradiol and progesterone in regulation of hippocampal dendritic spine density during the estrous cycle in the rat. J. Comp. Neurol. 1993, 336, 293–306. [Google Scholar] [CrossRef]
- Cuzzocrea, S.; Genovese, T.; Mazzon, E.; Esposito, E.; Di Paola, R.; Muia, C.; Crisafulli, C.; Peli, A.; Bramanti, P.; Chaudry, I.H. Effect of 17beta-estradiol on signal transduction pathways and secondary damage in experimental spinal cord trauma. Shock 2008, 29, 362–371. [Google Scholar] [CrossRef]
- Ritz, M.F.; Hausmann, O.N. Effect of 17beta-estradiol on functional outcome, release of cytokines, astrocyte reactivity and inflammatory spreading after spinal cord injury in male rats. Brain Res. 2008, 1203, 177–188. [Google Scholar] [CrossRef]
- Yune, T.Y.; Kim, S.J.; Lee, S.M.; Lee, Y.K.; Oh, Y.J.; Kim, Y.C.; Markelonis, G.J.; Oh, T.H. Systemic administration of 17beta-estradiol reduces apoptotic cell death and improves functional recovery following traumatic spinal cord injury in rats. J. Neurotrauma. 2004, 21, 293–306. [Google Scholar] [CrossRef]
- Zendedel, A.; Monnink, F.; Hassanzadeh, G.; Zaminy, A.; Ansar, M.M.; Habib, P.; Slowik, A.; Kipp, M.; Beyer, C. Estrogen Attenuates Local Inflammasome Expression and Activation after Spinal Cord Injury. Mol. Neurobiol. 2018, 55, 1364–1375. [Google Scholar] [CrossRef]
- Cox, A.; Capone, M.; Matzelle, D.; Vertegel, A.; Bredikhin, M.; Varma, A.; Haque, A.; Shields, D.C.; Banik, N.L. Nanoparticle-Based Estrogen Delivery to Spinal Cord Injury Site Reduces Local Parenchymal Destruction and Improves Functional Recovery. J. Neurotrauma. 2021, 38, 342–352. [Google Scholar] [CrossRef]
- Haque, A.; Drasites, K.P.; Cox, A.; Capone, M.; Myatich, A.I.; Shams, R.; Matzelle, D.; Garner, D.P.; Bredikhin, M.; Shields, D.C.; et al. Protective Effects of Estrogen via Nanoparticle Delivery to Attenuate Myelin Loss and Neuronal Death after Spinal Cord Injury. Neurochem. Res. 2021, 46, 2979–2990. [Google Scholar] [CrossRef]
- Markwell, J. Molecules and medicine by E.J. Corey, B Czakó, and L. Kürti. Biochem. Mol. Biol. Educ. 2008, 36, 245–246. [Google Scholar] [CrossRef]
- Pannu, R.; Barbosa, E.; Singh, A.K.; Singh, I. Attenuation of acute inflammatory response by atorvastatin after spinal cord injury in rats. J. Neurosci. Res. 2005, 79, 340–350. [Google Scholar] [CrossRef] [PubMed]
- Dery, M.A.; Rousseau, G.; Benderdour, M.; Beaumont, E. Atorvastatin prevents early apoptosis after thoracic spinal cord contusion injury and promotes locomotion recovery. Neurosci. Lett. 2009, 453, 73–76. [Google Scholar] [CrossRef] [PubMed]
- Kardes, O.; Civi, S.; Tufan, K.; Oyar, E.O.; Omeroglu, S.; Aykol, S. Effects of Atorvastatin on Experimental Spinal Cord Ischemia-Reperfusion Injury in Rabbits. Turk. Neurosurg. 2017, 27, 594–602. [Google Scholar] [CrossRef]
- Shen, Q.; Cao, Y.; Xia, Y. Atorvastatin attenuates spinal cord injury by chronic fluorosis in rats. Neuroreport 2019, 30, 1256–1260. [Google Scholar] [CrossRef]
- Hazzaa, S.M.; Abdou, A.G.; Ibraheim, E.O.; Salem, E.A.; Hassan, M.H.A.; Abdel-Razek, H.A.D. Effect of L-carnitine and atorvastatin on a rat model of ischemia-reperfusion injury of spinal cord. J. Immunoass. Immunochem. 2021, 42, 596–619. [Google Scholar] [CrossRef]
- Bimbova, K.; Bacova, M.; Kisucka, A.; Pavel, J.; Galik, J.; Zavacky, P.; Marsala, M.; Stropkovska, A.; Fedorova, J.; Papcunova, S.; et al. A Single Dose of Atorvastatin Applied Acutely after Spinal Cord Injury Suppresses Inflammation, Apoptosis, and Promotes Axon Outgrowth, Which Might Be Essential for Favorable Functional Outcome. Int. J. Mol. Sci. 2018, 19, 1106. [Google Scholar] [CrossRef]
- Aghazadeh, J.; Motlagh, P.S.; Salehpour, F.; Meshkini, A.; Fatehi, M.; Mirzaei, F.; Alavi, S.A.N. Effects of Atorvastatin in Patients with Acute Spinal Cord Injury. Asian Spine J. 2017, 11, 903–907. [Google Scholar] [CrossRef]
- Pannu, N.; Bhatnagar, A. Resveratrol: From enhanced biosynthesis and bioavailability to multitargeting chronic diseases. Biomed. Pharmacother. 2019, 109, 2237–2251. [Google Scholar] [CrossRef]
- Zhou, D.D.; Luo, M.; Huang, S.Y.; Saimaiti, A.; Shang, A.; Gan, R.Y.; Li, H.B. Effects and Mechanisms of Resveratrol on Aging and Age-Related Diseases. Oxid. Med. Cell Longev. 2021, 2021, 9932218. [Google Scholar] [CrossRef]
- Stockinger, J.; Maxwell, N.; Shapiro, D.; deCabo, R.; Valdez, G. Caloric Restriction Mimetics Slow Aging of Neuromuscular Synapses and Muscle Fibers. J. Gerontol. A Biol. Sci. Med. Sci. 2017, 73, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, C.X.; Liu, Y.M.; Chen, K.L.; Chen, G. A comparative study of anti-aging properties and mechanism: Resveratrol and caloric restriction. Oncotarget 2017, 8, 65717–65729. [Google Scholar] [CrossRef] [PubMed]
- Galiniak, S.; Aebisher, D.; Bartusik-Aebisher, D. Health benefits of resveratrol administration. Acta Biochim. Pol. 2019, 66, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Hecker, A.; Schellnegger, M.; Hofmann, E.; Luze, H.; Nischwitz, S.P.; Kamolz, L.P.; Kotzbeck, P. The impact of resveratrol on skin wound healing, scarring, and aging. Int. Wound J. 2022, 19, 9–28. [Google Scholar] [CrossRef]
- Meng, T.; Xiao, D.; Muhammed, A.; Deng, J.; Chen, L.; He, J. Anti-Inflammatory Action and Mechanisms of Resveratrol. Molecules 2021, 26, 229. [Google Scholar] [CrossRef]
- Pignet, A.L.; Schellnegger, M.; Hecker, A.; Kohlhauser, M.; Kotzbeck, P.; Kamolz, L.P. Resveratrol-Induced Signal Transduction in Wound Healing. Int. J. Mol. Sci. 2021, 22, 12614. [Google Scholar] [CrossRef]
- Liu, X.; Botchway, B.O.A.; Tan, X.; Zhang, Y.; Fang, M. Resveratrol treatment of spinal cord injury in rat model. Microsc. Res. Tech. 2019, 82, 296–303. [Google Scholar] [CrossRef]
- Miguel, C.A.; Noya-Riobo, M.V.; Mazzone, G.L.; Villar, M.J.; Coronel, M.F. Antioxidant, anti-inflammatory and neuroprotective actions of resveratrol after experimental nervous system insults. Special focus on the molecular mechanisms involved. Neurochem. Int. 2021, 150, 105188. [Google Scholar] [CrossRef]
- Fan, R.; Zhang, Y.; Botchway, B.O.A.; Liu, X. Resveratrol Can Attenuate Astrocyte Activation to Treat Spinal Cord Injury by Inhibiting Inflammatory Responses. Mol. Neurobiol. 2021, 58, 5799–5813. [Google Scholar] [CrossRef]
- Xu, L.; Botchway, B.O.A.; Zhang, S.; Zhou, J.; Liu, X. Inhibition of NF-kappaB Signaling Pathway by Resveratrol Improves Spinal Cord Injury. Front. Neurosci. 2018, 12, 690. [Google Scholar] [CrossRef]
- Zhou, J.; Huo, X.; Botchway, B.O.A.; Xu, L.; Meng, X.; Zhang, S.; Liu, X. Beneficial Effects of Resveratrol-Mediated Inhibition of the mTOR Pathway in Spinal Cord Injury. Neural. Plast. 2018, 2018, 7513748. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Chen, S.; Gao, K.; Zhou, Z.; Wang, C.; Shen, Z.; Guo, Y.; Li, Z.; Wan, Z.; Liu, C.; et al. Resveratrol protects against spinal cord injury by activating autophagy and inhibiting apoptosis mediated by the SIRT1/AMPK signaling pathway. Neuroscience 2017, 348, 241–251. [Google Scholar] [CrossRef]
- Meng, H.Y.; Shao, D.C.; Li, H.; Huang, X.D.; Yang, G.; Xu, B.; Niu, H.Y. Resveratrol improves neurological outcome and neuroinflammation following spinal cord injury through enhancing autophagy involving the AMPK/mTOR pathway. Mol. Med. Rep. 2018, 18, 2237–2244. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Ambigaipalan, P. Omega-3 Polyunsaturated Fatty Acids and Their Health Benefits. Annu. Rev. Food Sci. Technol. 2018, 9, 345–381. [Google Scholar] [CrossRef]
- Djuricic, I.; Calder, P.C. Beneficial Outcomes of Omega-6 and Omega-3 Polyunsaturated Fatty Acids on Human Health: An Update for 2021. Nutrients 2021, 13, 2421. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, T.; Yoshida, M.; Arita, M. Omega-3 fatty acid-derived mediators that control inflammation and tissue homeostasis. Int. Immunol. 2019, 31, 559–567. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, S.; Svahn, S.L.; Johansson, M.E. Effects of Omega-3 Fatty Acids on Immune Cells. Int. J. Mol. Sci. 2019, 20, 5028. [Google Scholar] [CrossRef] [PubMed]
- Larrieu, T.; Laye, S. Food for Mood: Relevance of Nutritional Omega-3 Fatty Acids for Depression and Anxiety. Front. Physiol. 2018, 9, 1047. [Google Scholar] [CrossRef]
- Sun, G.Y.; Simonyi, A.; Fritsche, K.L.; Chuang, D.Y.; Hannink, M.; Gu, Z.; Greenlief, C.M.; Yao, J.K.; Lee, J.C.; Beversdorf, D.Q. Docosahexaenoic acid (DHA): An essential nutrient and a nutraceutical for brain health and diseases. Prostaglandins Leukot. Essent. Fatty Acids 2018, 136, 3–13. [Google Scholar] [CrossRef]
- Allison, D.J.; Beaudry, K.M.; Thomas, A.M.; Josse, A.R.; Ditor, D.S. Changes in nutrient intake and inflammation following an anti-inflammatory diet in spinal cord injury. J. Spinal Cord Med. 2019, 42, 768–777. [Google Scholar] [CrossRef]
- Wojdasiewicz, P.; Poniatowski, L.A.; Turczyn, P.; Frasunska, J.; Paradowska-Gorycka, A.; Tarnacka, B. Significance of Omega-3 Fatty Acids in the Prophylaxis and Treatment after Spinal Cord Injury in Rodent Models. Mediat. Inflamm. 2020, 2020, 3164260. [Google Scholar] [CrossRef] [PubMed]
- Baazm, M.; Behrens, V.; Beyer, C.; Nikoubashman, O.; Zendedel, A. Regulation of Inflammasomes by Application of Omega-3 Polyunsaturated Fatty Acids in a Spinal Cord Injury Model. Cells 2021, 10, 3147. [Google Scholar] [CrossRef] [PubMed]
- Bi, J.; Chen, C.; Sun, P.; Tan, H.; Feng, F.; Shen, J. Neuroprotective effect of omega-3 fatty acids on spinal cord injury induced rats. Brain Behav. 2019, 9, e01339. [Google Scholar] [CrossRef] [PubMed]
- Nie, J.; Chen, J.; Yang, J.; Pei, Q.; Li, J.; Liu, J.; Xu, L.; Li, N.; Chen, Y.; Chen, X.; et al. Inhibition of mammalian target of rapamycin complex 1 signaling by n-3 polyunsaturated fatty acids promotes locomotor recovery after spinal cord injury. Mol. Med. Rep. 2018, 17, 5894–5902. [Google Scholar] [CrossRef] [PubMed]
- Naderi, Y.; Panahi, Y.; Barreto, G.E.; Sahebkar, A. Neuroprotective effects of minocycline on focal cerebral ischemia injury: A systematic review. Neural. Regen. Res. 2020, 15, 773–782. [Google Scholar]
- Liu, Y.; Li, Z.; Khan, S.; Zhang, R.; Wei, R.; Zhang, Y.; Xue, M.; Yong, V.W. Neuroprotection of minocycline by inhibition of extracellular matrix metalloproteinase inducer expression following intracerebral hemorrhage in mice. Neurosci. Lett. 2021, 764, 136297. [Google Scholar] [CrossRef]
- Bassett, B.; Subramaniyam, S.; Fan, Y.; Varney, S.; Pan, H.; Carneiro, A.M.D.; Chung, C.Y. Minocycline alleviates depression-like symptoms by rescuing decrease in neurogenesis in dorsal hippocampus via blocking microglia activation/phagocytosis. Brain Behav. Immun. 2021, 91, 519–530. [Google Scholar] [CrossRef]
- Singh, S.; Khanna, D.; Kalra, S. Minocycline and Doxycycline: More Than Antibiotics. Curr. Mol. Pharmacol. 2021, 14, 1046–1065. [Google Scholar] [CrossRef]
- Xu, J.; Ji, J.; Wang, Z.; Xu, T. Effects of minocycline on motor function recovery and expression of glial fibrillary acidic protein and brain-derived neurotrophic factor after spinal cord injury in rats. J. Pharm. Pharmacol. 2021, 73, 332–337. [Google Scholar] [CrossRef]
- Zhou, Y.Q.; Liu, D.Q.; Chen, S.P.; Sun, J.; Wang, X.M.; Tian, Y.K.; Wu, W.; Ye, D.W. Minocycline as a promising therapeutic strategy for chronic pain. Pharmacol. Res. 2018, 134, 305–310. [Google Scholar] [CrossRef]
- Pourkhodadad, S.; Oryan, S.H.; Kaka, G.; Sadraie, S.H. Neuroprotective Effects of Combined Treatment with Minocycline and Olfactory Ensheathing Cells Transplantation against Inflammation and Oxidative Stress after Spinal Cord Injury. Cell J. 2019, 21, 220–228. [Google Scholar] [PubMed]
- Squair, J.W.; Ruiz, I.; Phillips, A.A.; Zheng, M.M.Z.; Sarafis, Z.K.; Sachdeva, R.; Gopaul, R.; Liu, J.; Tetzlaff, W.; West, C.R.; et al. Minocycline Reduces the Severity of Autonomic Dysreflexia after Experimental Spinal Cord Injury. J. Neurotrauma. 2018, 35, 2861–2871. [Google Scholar] [CrossRef] [PubMed]
- Casha, S.; Zygun, D.; McGowan, M.D.; Bains, I.; Yong, V.W.; Hurlbert, R.J. Results of a phase II placebo-controlled randomized trial of minocycline in acute spinal cord injury. Brain 2012, 135 Pt 4, 1224–1236. [Google Scholar] [CrossRef]
- Casha, S. Minocycline in Acute Spinal Cord Injury (MASC). Available online: https://clinicaltrials.gov/ct2/show/NCT01828203 (accessed on 30 October 2014).
- Casha, S.; Rice, T.; Stirling, D.P.; Silva, C.; Gnanapavan, S.; Giovannoni, G.; Hurlbert, R.J.; Yong, V.W. Cerebrospinal Fluid Biomarkers in Human Spinal Cord Injury from a Phase II Minocycline Trial. J. Neurotrauma. 2018, 35, 1918–1928. [Google Scholar] [CrossRef] [PubMed]
- Coll-Miro, M.; Francos-Quijorna, I.; Santos-Nogueira, E.; Torres-Espin, A.; Bufler, P.; Dinarello, C.A.; Lopez-Vales, R. Beneficial effects of IL-37 after spinal cord injury in mice. Proc. Natl. Acad. Sci. USA 2016, 113, 1411–1416. [Google Scholar] [CrossRef]
- Dooley, D.; Lemmens, E.; Vangansewinkel, T.; Le Blon, D.; Hoornaert, C.; Ponsaerts, P.; Hendrix, S. Cell-Based Delivery of Interleukin-13 Directs Alternative Activation of Macrophages Resulting in Improved Functional Outcome after Spinal Cord Injury. Stem Cell Rep. 2016, 7, 1099–1115. [Google Scholar] [CrossRef]
- Guo, Y.; Zhang, H.; Yang, J.; Liu, S.; Bing, L.; Gao, J.; Hao, A. Granulocyte colony-stimulating factor improves alternative activation of microglia under microenvironment of spinal cord injury. Neuroscience 2013, 238, 1–10. [Google Scholar] [CrossRef]
- Geng, Y.; Gulbins, E.; Altman, A.; Lotz, M. Monocyte deactivation by interleukin 10 via inhibition of tyrosine kinase activity and the Ras signaling pathway. Proc. Natl. Acad. Sci. USA 1994, 91, 8602–8606. [Google Scholar] [CrossRef]
- Mizuno, T.; Sawada, M.; Marunouchi, T.; Suzumura, A. Production of interleukin-10 by mouse glial cells in culture. Biochem. Biophys. Res. Commun. 1994, 205, 1907–1915. [Google Scholar] [CrossRef]
- Berkman, N.; John, M.; Roesems, G.; Jose, P.J.; Barnes, P.J.; Chung, K.F. Inhibition of macrophage inflammatory protein-1 alpha expression by IL-10. Differential sensitivities in human blood monocytes and alveolar macrophages. J. Immunol. 1995, 155, 4412–4418. [Google Scholar]
- Crisi, G.M.; Santambrogio, L.; Hochwald, G.M.; Smith, S.R.; Carlino, J.A.; Thorbecke, G.J. Staphylococcal enterotoxin B and tumor-necrosis factor-alpha-induced relapses of experimental allergic encephalomyelitis: Protection by transforming growth factor-beta and interleukin-10. Eur. J. Immunol. 1995, 25, 3035–3040. [Google Scholar] [CrossRef] [PubMed]
- Cunha, F.Q.; Moncada, S.; Liew, F.Y. Interleukin-10 (IL-10) inhibits the induction of nitric oxide synthase by interferon-gamma in murine macrophages. Biochem. Biophys. Res. Commun. 1992, 182, 1155–1159. [Google Scholar] [CrossRef]
- Frei, K.; Lins, H.; Schwerdel, C.; Fontana, A. Antigen presentation in the central nervous system. The inhibitory effect of IL-10 on MHC class II expression and production of cytokines depends on the inducing signals and the type of cell analyzed. J. Immunol. 1994, 152, 2720–2728. [Google Scholar]
- Issazadeh, S.; Lorentzen, J.C.; Mustafa, M.I.; Hojeberg, B.; Mussener, A.; Olsson, T. Cytokines in relapsing experimental autoimmune encephalomyelitis in DA rats: Persistent mRNA expression of proinflammatory cytokines and absent expression of interleukin-10 and transforming growth factor-beta. J. Neuroimmunol. 1996, 69, 103–115. [Google Scholar] [CrossRef]
- Rott, O.; Fleischer, B.; Cash, E. Interleukin-10 prevents experimental allergic encephalomyelitis in rats. Eur. J. Immunol. 1994, 24, 1434–1440. [Google Scholar] [CrossRef]
- Ho, I.C.; Miaw, S.C. Regulation of IL-4 Expression in Immunity and Diseases. Adv. Exp. Med. Biol. 2016, 941, 31–77. [Google Scholar]
- Chatterjee, P.; Chiasson, V.L.; Bounds, K.R.; Mitchell, B.M. Regulation of the Anti-Inflammatory Cytokines Interleukin-4 and Interleukin-10 during Pregnancy. Front. Immunol. 2014, 5, 253. [Google Scholar] [CrossRef]
- Francos-Quijorna, I.; Amo-Aparicio, J.; Martinez-Muriana, A.; Lopez-Vales, R. IL-4 drives microglia and macrophages toward a phenotype conducive for tissue repair and functional recovery after spinal cord injury. Glia 2016, 64, 2079–2092. [Google Scholar] [CrossRef]
- Lima, R.; Monteiro, S.; Lopes, J.P.; Barradas, P.; Vasconcelos, N.L.; Gomes, E.D.; Assuncao-Silva, R.C.; Teixeira, F.G.; Morais, M.; Sousa, N.; et al. Systemic Interleukin-4 Administration after Spinal Cord Injury Modulates Inflammation and Promotes Neuroprotection. Pharmaceuticals 2017, 10, 83. [Google Scholar] [CrossRef]
- Saito-Sasaki, N.; Sawada, Y.; Nakamura, M. Maresin-1 and Inflammatory Disease. Int. J. Mol. Sci. 2022, 23, 5522. [Google Scholar] [CrossRef]
- Chiang, N.; Libreros, S.; Norris, P.C.; de la Rosa, X.; Serhan, C.N. Maresin 1 activates LGR6 receptor promoting phagocyte immunoresolvent functions. J. Clin. Investig. 2019, 129, 5294–5311. [Google Scholar] [CrossRef] [PubMed]
- Im, D.S. Maresin-1 resolution with RORalpha and LGR6. Prog. Lipid Res. 2020, 78, 101034. [Google Scholar] [CrossRef] [PubMed]
- Francos-Quijorna, I.; Santos-Nogueira, E.; Gronert, K.; Sullivan, A.B.; Kopp, M.A.; Brommer, B.; David, S.; Schwab, J.M.; Lopez-Vales, R. Maresin 1 Promotes Inflammatory Resolution, Neuroprotection, and Functional Neurological Recovery After Spinal Cord Injury. J. Neurosci. 2017, 37, 11731–11743. [Google Scholar] [CrossRef]
- Jordan, J.D.; Carhuapoma, J.R. Hypothermia: Comparing technology. J. Neurol. Sci. 2007, 261, 35–38. [Google Scholar] [CrossRef] [PubMed]
- Green, R.S.; Howes, D.W. Stock your emergency department with ice packs: A practical guide to therapeutic hypothermia for survivors of cardiac arrest. CMAJ 2007, 176, 759–762. [Google Scholar] [CrossRef] [PubMed][Green Version]
- van der Worp, H.B.; Macleod, M.R.; Bath, P.M.; Demotes, J.; Durand-Zaleski, I.; Gebhardt, B.; Gluud, C.; Kollmar, R.; Krieger, D.W.; Lees, K.R.; et al. EuroHYP-1: European multicenter, randomized, phase III clinical trial of therapeutic hypothermia plus best medical treatment vs. best medical treatment alone for acute ischemic stroke. Int. J. Stroke 2014, 9, 642–645. [Google Scholar] [CrossRef] [PubMed]
- Martirosyan, N.L.; Patel, A.A.; Carotenuto, A.; Kalani, M.Y.; Bohl, M.A.; Preul, M.C.; Theodore, N. The role of therapeutic hypothermia in the management of acute spinal cord injury. Clin. Neurol. Neurosurg. 2017, 154, 79–88. [Google Scholar] [CrossRef]
- Hansebout, R.R.; Hansebout, C.R. Local cooling for traumatic spinal cord injury: Outcomes in 20 patients and review of the literature. J. Neurosurg. Spine 2014, 20, 550–561. [Google Scholar] [CrossRef]
- Levi, A.D.; Casella, G.; Green, B.A.; Dietrich, W.D.; Vanni, S.; Jagid, J.; Wang, M.Y. Clinical outcomes using modest intravascular hypothermia after acute cervical spinal cord injury. Neurosurgery 2010, 66, 670–677. [Google Scholar] [CrossRef]
- Vedantam, A.; Jimsheleishvili, G.; Harrop, J.S.; Alberga, L.R.; Ahmad, F.U.; Murphy, R.K.; Jackson, J.B., 3rd; Rodgers, R.B.; Levi, A.D. A prospective multi-center study comparing the complication profile of modest systemic hypothermia versus normothermia for acute cervical spinal cord injury. Spinal Cord 2022, 60, 510–515. [Google Scholar] [CrossRef]
- Mohammed, R.; Opara, K.; Lall, R.; Ojha, U.; Xiang, J. Evaluating the effectiveness of anti-Nogo treatment in spinal cord injuries. Neural. Dev. 2020, 15, 1. [Google Scholar] [CrossRef] [PubMed]
- Tsujioka, H.; Yamashita, T. Neural circuit repair after central nervous system injury. Int. Immunol. 2021, 33, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Bregman, B.S.; Kunkel-Bagden, E.; Schnell, L.; Dai, H.N.; Gao, D.; Schwab, M.E. Recovery from spinal cord injury mediated by antibodies to neurite growth inhibitors. Nature 1995, 378, 498–501. [Google Scholar] [CrossRef] [PubMed]
- Merkler, D.; Metz, G.A.; Raineteau, O.; Dietz, V.; Schwab, M.E.; Fouad, K. Locomotor recovery in spinal cord-injured rats treated with an antibody neutralizing the myelin-associated neurite growth inhibitor Nogo-A. J. Neurosci. 2001, 21, 3665–3673. [Google Scholar] [CrossRef] [PubMed]
- Schnell, L.; Schwab, M.E. Axonal regeneration in the rat spinal cord produced by an antibody against myelin-associated neurite growth inhibitors. Nature 1990, 343, 269–272. [Google Scholar] [CrossRef]
- Freund, P.; Schmidlin, E.; Wannier, T.; Bloch, J.; Mir, A.; Schwab, M.E.; Rouiller, E.M. Nogo-A-specific antibody treatment enhances sprouting and functional recovery after cervical lesion in adult primates. Nat. Med. 2006, 12, 790–792. [Google Scholar] [CrossRef]
- Freund, P.; Wannier, T.; Schmidlin, E.; Bloch, J.; Mir, A.; Schwab, M.E.; Rouiller, E.M. Anti-Nogo-A antibody treatment enhances sprouting of corticospinal axons rostral to a unilateral cervical spinal cord lesion in adult macaque monkey. J. Comp. Neurol. 2007, 502, 644–659. [Google Scholar] [CrossRef]
- Schwab, M.E. Nogo and axon regeneration. Curr. Opin. Neurobiol. 2004, 14, 118–124. [Google Scholar] [CrossRef]
- Liebscher, T.; Schnell, L.; Schnell, D.; Scholl, J.; Schneider, R.; Gullo, M.; Fouad, K.; Mir, A.; Rausch, M.; Kindler, D.; et al. Nogo-A antibody improves regeneration and locomotion of spinal cord-injured rats. Ann. Neurol. 2005, 58, 706–719. [Google Scholar] [CrossRef]
- Chen, K.; Marsh, B.C.; Cowan, M.; Al’Joboori, Y.D.; Gigout, S.; Smith, C.C.; Messenger, N.; Gamper, N.; Schwab, M.E.; Ichiyama, R.M. Sequential therapy of anti-Nogo-A antibody treatment and treadmill training leads to cumulative improvements after spinal cord injury in rats. Exp. Neurol. 2017, 292, 135–144. [Google Scholar] [CrossRef]
- Schneider, M.P.; Sartori, A.M.; Ineichen, B.V.; Moors, S.; Engmann, A.K.; Hofer, A.S.; Weinmann, O.; Kessler, T.M.; Schwab, M.E. Anti-Nogo-A Antibodies As a Potential Causal Therapy for Lower Urinary Tract Dysfunction after Spinal Cord Injury. J. Neurosci. 2019, 39, 4066–4076. [Google Scholar] [CrossRef] [PubMed]
- Kucher, K.; Johns, D.; Maier, D.; Abel, R.; Badke, A.; Baron, H.; Thietje, R.; Casha, S.; Meindl, R.; Gomez-Mancilla, B.; et al. First-in-Man Intrathecal Application of Neurite Growth-Promoting Anti-Nogo-A Antibodies in Acute Spinal Cord Injury. Neurorehabil. Neural. Repair. 2018, 32, 578–589. [Google Scholar] [CrossRef] [PubMed]
- Sartori, A.M.; Hofer, A.-S.; Schwab, M.E. Recovery after spinal cord injury is enhanced by anti-Nogo-A antibody therapy—from animal models to clinical trials. Curr. Opin. Physiol. 2020, 14, 1–6. [Google Scholar] [CrossRef]
- Prabhakar, V.; Capila, I.; Bosques, C.J.; Pojasek, K.; Sasisekharan, R. Chondroitinase ABC I from Proteus vulgaris: Cloning, recombinant expression and active site identification. Biochem. J. 2005, 386 Pt 1, 103–112. [Google Scholar] [CrossRef]
- Bradbury, E.J.; Moon, L.D.; Popat, R.J.; King, V.R.; Bennett, G.S.; Patel, P.N.; Fawcett, J.W.; McMahon, S.B. Chondroitinase ABC promotes functional recovery after spinal cord injury. Nature 2002, 416, 636–640. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Alias, G.; Barkhuysen, S.; Buckle, M.; Fawcett, J.W. Chondroitinase ABC treatment opens a window of opportunity for task-specific rehabilitation. Nat. Neurosci. 2009, 12, 1145–1151. [Google Scholar] [CrossRef]
- Tester, N.J.; Howland, D.R. Chondroitinase ABC improves basic and skilled locomotion in spinal cord injured cats. Exp. Neurol. 2008, 209, 483–496. [Google Scholar] [CrossRef]
- Asher, R.A.; Morgenstern, D.A.; Moon, L.D.; Fawcett, J.W. Chondroitin sulphate proteoglycans: Inhibitory components of the glial scar. Prog. Brain Res. 2001, 132, 611–619. [Google Scholar]
- Takiguchi, M.; Miyashita, K.; Yamazaki, K.; Funakoshi, K. Chondroitinase ABC Administration Facilitates Serotonergic Innervation of Motoneurons in Rats With Complete Spinal Cord Transection. Front. Integr. Neurosci. 2022, 16, 881632. [Google Scholar] [CrossRef]
- Janzadeh, A.; Sarveazad, A.; Hamblin, M.R.; Teheripak, G.; Kookli, K.; Nasirinezhad, F. The effect of chondroitinase ABC and photobiomodulation therapy on neuropathic pain after spinal cord injury in adult male rats. Physiol. Behav. 2020, 227, 113141. [Google Scholar] [CrossRef]
- Lee, Y.S.; Lin, C.Y.; Jiang, H.H.; Depaul, M.; Lin, V.W.; Silver, J. Nerve regeneration restores supraspinal control of bladder function after complete spinal cord injury. J. Neurosci. 2013, 33, 10591–10606. [Google Scholar] [CrossRef] [PubMed]
- Alilain, W.J.; Horn, K.P.; Hu, H.; Dick, T.E.; Silver, J. Functional regeneration of respiratory pathways after spinal cord injury. Nature 2011, 475, 196–200. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Alias, G.; Lin, R.; Akrimi, S.F.; Story, D.; Bradbury, E.J.; Fawcett, J.W. Therapeutic time window for the application of chondroitinase ABC after spinal cord injury. Exp. Neurol. 2008, 210, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Alias, G.; Lopez-Vales, R.; Fores, J.; Navarro, X.; Verdu, E. Acute transplantation of olfactory ensheathing cells or Schwann cells promotes recovery after spinal cord injury in the rat. J. Neurosci. Res. 2004, 75, 632–641. [Google Scholar] [CrossRef]
- Muir, E.; De Winter, F.; Verhaagen, J.; Fawcett, J. Recent advances in the therapeutic uses of chondroitinase ABC. Exp. Neurol. 2019, 321, 113032. [Google Scholar] [CrossRef] [PubMed]
- Day, P.; Alves, N.; Daniell, E.; Dasgupta, D.; Ogborne, R.; Steeper, A.; Raza, M.; Ellis, C.; Fawcett, J.; Keynes, R.; et al. Targeting chondroitinase ABC to axons enhances the ability of chondroitinase to promote neurite outgrowth and sprouting. PLoS ONE 2020, 15, e0221851. [Google Scholar] [CrossRef]
- Burnside, E.R.; De Winter, F.; Didangelos, A.; James, N.D.; Andreica, E.C.; Layard-Horsfall, H.; Muir, E.M.; Verhaagen, J.; Bradbury, E.J. Immune-evasive gene switch enables regulated delivery of chondroitinase after spinal cord injury. Brain 2018, 141, 2362–2381. [Google Scholar] [CrossRef]
- Raspa, A.; Carminati, L.; Pugliese, R.; Fontana, F.; Gelain, F. Self-assembling peptide hydrogels for the stabilization and sustained release of active Chondroitinase ABC in vitro and in spinal cord injuries. J. Control Release 2021, 330, 1208–1219. [Google Scholar] [CrossRef]
- Prager, J.; Ito, D.; Carwardine, D.R.; Jiju, P.; Chari, D.M.; Granger, N.; Wong, L.F. Delivery of chondroitinase by canine mucosal olfactory ensheathing cells alongside rehabilitation enhances recovery after spinal cord injury. Exp. Neurol. 2021, 340, 113660. [Google Scholar] [CrossRef]
- Prager, J.; Fenn, J.; Plested, M.; Escauriaza, L.; Merwe, T.V.; King, B.; Chari, D.; Wong, L.F.; Granger, N. Transplantation of encapsulated autologous olfactory ensheathing cell populations expressing chondroitinase for spinal cord injury: A safety and feasibility study in companion dogs. J. Tissue Eng. Regen. Med. 2022, 16, 788–798. [Google Scholar] [CrossRef]
- Nori, S.; Khazaei, M.; Ahuja, C.S.; Yokota, K.; Ahlfors, J.E.; Liu, Y.; Wang, J.; Shibata, S.; Chio, J.; Hettiaratchi, M.H.; et al. Human Oligodendrogenic Neural Progenitor Cells Delivered with Chondroitinase ABC Facilitate Functional Repair of Chronic Spinal Cord Injury. Stem Cell Rep. 2018, 11, 1433–1448. [Google Scholar] [CrossRef] [PubMed]
- Fuhrmann, T.; Anandakumaran, P.N.; Payne, S.L.; Pakulska, M.M.; Varga, B.V.; Nagy, A.; Tator, C.; Shoichet, M.S. Combined delivery of chondroitinase ABC and human induced pluripotent stem cell-derived neuroepithelial cells promote tissue repair in an animal model of spinal cord injury. Biomed. Mater. 2018, 13, 024103. [Google Scholar] [CrossRef] [PubMed]
- Labombarda, F.; Deniselle, M.C.G.; De Nicola, A.F.; Gonzalez, S.L. Progesterone and the spinal cord: Good friends in bad times. Neuroimmunomodulation 2010, 17, 146–149. [Google Scholar] [CrossRef]
- Harayama, T.; Shimizu, T. Roles of polyunsaturated fatty acids, from mediators to membranes. J. Lipid Res. 2020, 61, 1150–1160. [Google Scholar] [CrossRef] [PubMed]
- Bethea, J.R.; Nagashima, H.; Acosta, M.C.; Briceno, C.; Gomez, F.; Marcillo, A.E.; Loor, K.; Green, J.; Dietrich, W.D. Systemically administered interleukin-10 reduces tumor necrosis factor-alpha production and significantly improves functional recovery following traumatic spinal cord injury in rats. J. Neurotrauma. 1999, 16, 851–863. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Fu, S.; Zamarin, D.; Bromberg, J. Interleukin-10. In Encyclopedia of Hormones; Henry, H.L., Norman, A.W., Eds.; Academic Press: New York, NY, USA, 2003; pp. 453–462. [Google Scholar]
- Hurdayal, R.; Brombacher, F. Interleukin-4 Receptor Alpha: From Innate to Adaptive Immunity in Murine Models of Cutaneous Leishmaniasis. Front. Immunol. 2017, 8, 1354. [Google Scholar] [CrossRef]
- Zhao, M.; Li, C.; Zhang, J.; Yin, Z.; Zheng, Z.; Wan, J.; Wang, M. Maresin-1 and its receptors RORalpha/LGR6 as potential therapeutic target for respiratory diseases. Pharmacol. Res. 2022, 182, 106337. [Google Scholar] [CrossRef]
- Gomes, E.D.; Rocha, L.A.; Assunção-Silva, R.C.; Lima, R.; Silva, N.A.; Salgado, A.J. Chapter 9-Cell therapies for spinal cord injury regeneration. In Spinal Cord Injury (SCI) Repair Strategies; Perale, G., Rossi, F., Eds.; Woodhead Publishing: Cambridge, UK, 2020; pp. 157–186. [Google Scholar]
- Willerth, S.M.; Sakiyama-Elbert, S.E. Cell therapy for spinal cord regeneration. Adv. Drug Deliv. Rev. 2008, 60, 263–276. [Google Scholar] [CrossRef]
- Rossi, S.L.; Keirstead, H.S. Stem cells and spinal cord regeneration. Curr. Opin. Biotechnol. 2009, 20, 552–562. [Google Scholar] [CrossRef]
- Pereira, I.M.; Marote, A.; Salgado, A.J.; Silva, N.A. Filling the Gap: Neural Stem Cells as A Promising Therapy for Spinal Cord Injury. Pharmaceuticals 2019, 12, 65. [Google Scholar] [CrossRef]
- Elisseeff, J.H. Embryonic stem cells: Potential for more impact. Trends Biotechnol. 2004, 22, 155–156. [Google Scholar] [CrossRef] [PubMed]
- Keirstead, H.S.; Nistor, G.; Bernal, G.; Totoiu, M.; Cloutier, F.; Sharp, K.; Steward, O. Human embryonic stem cell-derived oligodendrocyte progenitor cell transplants remyelinate and restore locomotion after spinal cord injury. J. Neurosci. 2005, 25, 4694–4705. [Google Scholar] [CrossRef] [PubMed]
- Kakinohana, O.; Juhasova, J.; Juhas, S.; Motlik, J.; Platoshyn, O.; Galik, J.; Hefferan, M.; Yuan, S.H.; Vidal, J.G.; Carson, C.T.; et al. Survival and differentiation of human embryonic stem cell-derived neural precursors grafted spinally in spinal ischemia-injured rats or in naive immunosuppressed minipigs: A qualitative and quantitative study. Cell Transplant. 2012, 21, 2603–2619. [Google Scholar] [CrossRef] [PubMed]
- Lukovic, D.; Stojkovic, M.; Moreno-Manzano, V.; Bhattacharya, S.S.; Erceg, S. Perspectives and future directions of human pluripotent stem cell-based therapies: Lessons from Geron’s clinical trial for spinal cord injury. Stem Cells Dev. 2014, 23, 1–4. [Google Scholar] [CrossRef]
- Pollack, A. Geron Is Shutting Down Its Stem Cell Clinical Trial. The New York Times, 14 November 2011. [Google Scholar]
- Shroff, G. Human Embryonic Stem Cell Therapy in Chronic Spinal Cord Injury: A Retrospective Study. Clin. Transl. Sci. 2016, 9, 168–175. [Google Scholar] [CrossRef]
- Shroff, G. 660. Bowel/Bladder Sensation and Control in Patients with Spinal Cord Injury Treated with Human Embryonic Stem Cell Therapy. Mol. Ther. 2015, 23, S262–S263. [Google Scholar] [CrossRef][Green Version]
- Sleeboom-Faulkner, M. Debates on Human Embryonic Stem Cell Research in Japan: Minority Voices and their Political Amplifiers. Sci. Cult. 2008, 17, 85–97. [Google Scholar] [CrossRef]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Okada, Y.; Itakura, G.; Iwai, H.; Nishimura, S.; Yasuda, A.; Nori, S.; Hikishima, K.; Konomi, T.; Fujiyoshi, K.; et al. Pre-evaluated safe human iPSC-derived neural stem cells promote functional recovery after spinal cord injury in common marmoset without tumorigenicity. PLoS ONE 2012, 7, e52787. [Google Scholar] [CrossRef]
- Tsuji, O.; Sugai, K.; Yamaguchi, R.; Tashiro, S.; Nagoshi, N.; Kohyama, J.; Iida, T.; Ohkubo, T.; Itakura, G.; Isoda, M.; et al. Concise Review: Laying the Groundwork for a First-In-Human Study of an Induced Pluripotent Stem Cell-Based Intervention for Spinal Cord Injury. Stem Cells 2019, 37, 6–13. [Google Scholar] [CrossRef]
- Santamaria, A.J.; Solano, J.P.; Benavides, F.D.; Guest, J.D. Intraspinal Delivery of Schwann Cells for Spinal Cord Injury. Methods Mol. Biol. 2018, 1739, 467–484. [Google Scholar] [PubMed]
- Anderson, K.D.; Guest, J.D.; Dietrich, W.D.; Bunge, M.B.; Curiel, R.; Dididze, M.; Green, B.A.; Khan, A.; Pearse, D.D.; Saraf-Lavi, E.; et al. Safety of Autologous Human Schwann Cell Transplantation in Subacute Thoracic Spinal Cord Injury. J. Neurotrauma. 2017, 34, 2950–2963. [Google Scholar] [CrossRef] [PubMed]
- Ursavas, S.; Darici, H.; Karaoz, E. Olfactory ensheathing cells: Unique glial cells promising for treatments of spinal cord injury. J. Neurosci. Res. 2021, 99, 1579–1597. [Google Scholar] [CrossRef] [PubMed]
- Gomes, E.D.; Mendes, S.S.; Assuncao-Silva, R.C.; Teixeira, F.G.; Pires, A.O.; Anjo, S.I.; Manadas, B.; Leite-Almeida, H.; Gimble, J.M.; Sousa, N.; et al. Co-Transplantation of Adipose Tissue-Derived Stromal Cells and Olfactory Ensheathing Cells for Spinal Cord Injury Repair. Stem Cells 2018, 36, 696–708. [Google Scholar] [CrossRef]
- Pinho, A.G.; Cibrao, J.R.; Lima, R.; Gomes, E.D.; Serra, S.C.; Lentilhas-Graca, J.; Ribeiro, C.; Lanceros-Mendez, S.; Teixeira, F.G.; Monteiro, S.; et al. Immunomodulatory and regenerative effects of the full and fractioned adipose tissue derived stem cells secretome in spinal cord injury. Exp. Neurol. 2022, 351, 113989. [Google Scholar] [CrossRef]
- Pinho, A.G.; Cibrao, J.R.; Silva, N.A.; Monteiro, S.; Salgado, A.J. Cell Secretome: Basic Insights and Therapeutic Opportunities for CNS Disorders. Pharmaceuticals 2020, 13, 31. [Google Scholar] [CrossRef]
- Serra, S.C.; Costa, J.C.; Assuncao-Silva, R.C.; Teixeira, F.G.; Silva, N.A.; Anjo, S.I.; Manadas, B.; Gimble, J.M.; Behie, L.A.; Salgado, A.J. Influence of passage number on the impact of the secretome of adipose tissue stem cells on neural survival, neurodifferentiation and axonal growth. Biochimie 2018, 155, 119–128. [Google Scholar] [CrossRef]
- Lammertse, D.P.; Jones, L.A.; Charlifue, S.B.; Kirshblum, S.C.; Apple, D.F.; Ragnarsson, K.T.; Falci, S.P.; Heary, R.F.; Choudhri, T.F.; Jenkins, A.L.; et al. Autologous incubated macrophage therapy in acute, complete spinal cord injury: Results of the phase 2 randomized controlled multicenter trial. Spinal Cord 2012, 50, 661–671. [Google Scholar] [CrossRef]
- Pang, Q.M.; Qian, N.N.; Zou, W.H.; Yang, Y.C.; Chen, H.; Zhang, M.; Zhang, Q.; Ao, J.; Zhang, T. PBMSCs transplantation facilitates functional recovery after spinal cord injury by regulating microglia/macrophages plasticity. Transpl. Immunol. 2022, 72, 101592. [Google Scholar] [CrossRef]
- Han, G.H.; Kim, S.J.; Ko, W.K.; Lee, D.; Han, I.B.; Sheen, S.H.; Hong, J.B.; Sohn, S. Transplantation of tauroursodeoxycholic acid-inducing M2-phenotype macrophages promotes an anti-neuroinflammatory effect and functional recovery after spinal cord injury in rats. Cell Prolif. 2021, 54, e13050. [Google Scholar] [CrossRef]
- Rocha, L.A.; Silva, D.; Barata-Antunes, S.; Cavaleiro, H.; Gomes, E.D.; Silva, N.A.; Salgado, A.J. Cell and Tissue Instructive Materials for Central Nervous System Repair. Adv. Funct. Mater. 2020, 30, 1909083. [Google Scholar] [CrossRef]
- Oliveira, E.; Assuncao-Silva, R.C.; Ziv-Polat, O.; Gomes, E.D.; Teixeira, F.G.; Silva, N.A.; Shahar, A.; Salgado, A.J. Influence of Different ECM-Like Hydrogels on Neurite Outgrowth Induced by Adipose Tissue-Derived Stem Cells. Stem Cells Int. 2017, 2017, 6319129. [Google Scholar] [CrossRef] [PubMed]
- Tabata, Y. Biomaterial technology for tissue engineering applications. J. R. Soc. Interface 2009, 6 (Suppl. S3), S311–S324. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.; Li, X.; Xiao, Z.; Zhao, Y.; Liang, H.; Wang, B.; Han, S.; Li, X.; Xu, B.; Wang, N.; et al. A modified collagen scaffold facilitates endogenous neurogenesis for acute spinal cord injury repair. Acta Biomater. 2017, 51, 304–316. [Google Scholar] [CrossRef] [PubMed]
- Zaviskova, K.; Tukmachev, D.; Dubisova, J.; Vackova, I.; Hejcl, A.; Bystronova, J.; Pravda, M.; Scigalkova, I.; Sulakova, R.; Velebny, V.; et al. Injectable hydroxyphenyl derivative of hyaluronic acid hydrogel modified with RGD as scaffold for spinal cord injury repair. J. Biomed. Mater. Res. A 2018, 106, 1129–1140. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Lee, J.Y.; Heo, E.Y.; Kwon, I.K.; Yune, T.Y.; Youn, I. Implantation of a Matrigel-loaded agarose scaffold promotes functional regeneration of axons after spinal cord injury in rat. Biochem. Biophys. Res. Commun. 2018, 496, 785–791. [Google Scholar] [CrossRef] [PubMed]
- Jarrah, R.; Sammak, S.E.; Onyedimma, C.; Ghaith, A.K.; Moinuddin, F.M.; Bhandarkar, A.R.; Siddiqui, A.; Madigan, N.; Bydon, M. The Role of Alginate Hydrogels as a Potential Treatment Modality for Spinal Cord Injury: A Comprehensive Review of the Literature. Neurospine 2022, 19, 272–280. [Google Scholar] [CrossRef]
- Gomes, E.D.; Ghosh, B.; Lima, R.; Goulao, M.; Moreira-Gomes, T.; Martins-Macedo, J.; Urban, M.W.; Wright, M.C.; Gimble, J.M.; Sousa, N.; et al. Combination of a Gellan Gum-Based Hydrogel With Cell Therapy for the Treatment of Cervical Spinal Cord Injury. Front. Bioeng. Biotechnol. 2020, 8, 984. [Google Scholar] [CrossRef]
- Rocha, L.A.; Gomes, E.D.; Afonso, J.L.; Granja, S.; Baltazar, F.; Silva, N.A.; Shoichet, M.S.; Sousa, R.A.; Learmonth, D.A.; Salgado, A.J. In vitro Evaluation of ASCs and HUVECs Co-cultures in 3D Biodegradable Hydrogels on Neurite Outgrowth and Vascular Organization. Front. Cell Dev. Biol. 2020, 8, 489. [Google Scholar] [CrossRef]
- Vacanti, C.A.; Vacanti, J.P. The science of tissue engineering. Orthop. Clin. N. Am. 2000, 31, 351–356. [Google Scholar] [CrossRef]
- Sun, X.; Bai, Y.; Zhai, H.; Liu, S.; Zhang, C.; Xu, Y.; Zou, J.; Wang, T.; Chen, S.; Zhu, Q.; et al. Devising micro/nano-architectures in multi-channel nerve conduits towards a pro-regenerative matrix for the repair of spinal cord injury. Acta Biomater. 2019, 86, 194–206. [Google Scholar] [CrossRef]
- Wong, D.Y.; Leveque, J.C.; Brumblay, H.; Krebsbach, P.H.; Hollister, S.J.; Lamarca, F. Macro-architectures in spinal cord scaffold implants influence regeneration. J. Neurotrauma. 2008, 25, 1027–1037. [Google Scholar] [CrossRef] [PubMed]
- Inada, Y.; Morimoto, S.; Moroi, K.; Endo, K.; Nakamura, T. Surgical relief of causalgia with an artificial nerve guide tube: Successful surgical treatment of causalgia (Complex Regional Pain Syndrome Type II) by in situ tissue engineering with a polyglycolic acid-collagen tube. Pain 2005, 117, 251–258. [Google Scholar] [CrossRef]
- Gomes, E.D.; Silva, N.A.; Salgado, A.J. Combinatorial therapies for spinal cord injury: Strategies to induce regeneration. Neural. Regen. Res. 2019, 14, 69–71. [Google Scholar] [PubMed]
- Philippson, M. L’autonomie et la Centralisation Dans le Système Nerveux des Animaux; Étude de Physiologie Expérimentale et Comparée; Falk: Bruxelles, Belgium, 1905. [Google Scholar]
- Sherrington, C.S. Flexion-reflex of the limb, crossed extension-reflex, and reflex stepping and standing. J. Physiol. 1910, 40, 28–121. [Google Scholar] [CrossRef] [PubMed]
- Brown, T.G.; Sherrington, C.S. The intrinsic factors in the act of progression in the mammal. Proc. R. Soc. Lond. Ser. B Contain. Pap. A Biol. Character 1911, 84, 308–319. [Google Scholar]
- Harkema, S.J. Plasticity of interneuronal networks of the functionally isolated human spinal cord. Brain Res. Rev. 2008, 57, 255–264. [Google Scholar] [CrossRef]
- Angeli, C.A.; Boakye, M.; Morton, R.A.; Vogt, J.; Benton, K.; Chen, Y.; Ferreira, C.K.; Harkema, S.J. Recovery of Over-Ground Walking after Chronic Motor Complete Spinal Cord Injury. N. Engl. J. Med. 2018, 379, 1244–1250. [Google Scholar] [CrossRef]
- Gill, M.L.; Grahn, P.J.; Calvert, J.S.; Linde, M.B.; Lavrov, I.A.; Strommen, J.A.; Beck, L.A.; Sayenko, D.G.; Van Straaten, M.G.; Drubach, D.I.; et al. Neuromodulation of lumbosacral spinal networks enables independent stepping after complete paraplegia. Nat. Med. 2018, 24, 1677–1682. [Google Scholar] [CrossRef]
- Wagner, F.B.; Mignardot, J.B.; Le Goff-Mignardot, C.G.; Demesmaeker, R.; Komi, S.; Capogrosso, M.; Rowald, A.; Seanez, I.; Caban, M.; Pirondini, E.; et al. Targeted neurotechnology restores walking in humans with spinal cord injury. Nature 2018, 563, 65–71. [Google Scholar] [CrossRef]
- Harkema, S.J.; Ditterline, B.L.; Wang, S.; Aslan, S.; Angeli, C.A.; Ovechkin, A.; Hirsch, G.A. Epidural Spinal Cord Stimulation Training and Sustained Recovery of Cardiovascular Function in Individuals With Chronic Cervical Spinal Cord Injury. JAMA Neurol. 2018, 75, 1569–1571. [Google Scholar] [CrossRef] [PubMed]
- Herrity, A.N.; Williams, C.S.; Angeli, C.A.; Harkema, S.J.; Hubscher, C.H. Lumbosacral spinal cord epidural stimulation improves voiding function after human spinal cord injury. Sci. Rep. 2018, 8, 8688. [Google Scholar] [CrossRef] [PubMed]
- Squair, J.W.; Gautier, M.; Mahe, L.; Soriano, J.E.; Rowald, A.; Bichat, A.; Cho, N.; Anderson, M.A.; James, N.D.; Gandar, J.; et al. Neuroprosthetic baroreflex controls haemodynamics after spinal cord injury. Nature 2021, 590, 308–314. [Google Scholar] [CrossRef] [PubMed]
- Wernig, A. No dawn yet of a new age in spinal cord rehabilitation. Brain 2015, 138 Pt 7, e362. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lima, R.; Monteiro, A.; Salgado, A.J.; Monteiro, S.; Silva, N.A. Pathophysiology and Therapeutic Approaches for Spinal Cord Injury. Int. J. Mol. Sci. 2022, 23, 13833. https://doi.org/10.3390/ijms232213833
Lima R, Monteiro A, Salgado AJ, Monteiro S, Silva NA. Pathophysiology and Therapeutic Approaches for Spinal Cord Injury. International Journal of Molecular Sciences. 2022; 23(22):13833. https://doi.org/10.3390/ijms232213833
Chicago/Turabian StyleLima, Rui, Andreia Monteiro, António J. Salgado, Susana Monteiro, and Nuno A. Silva. 2022. "Pathophysiology and Therapeutic Approaches for Spinal Cord Injury" International Journal of Molecular Sciences 23, no. 22: 13833. https://doi.org/10.3390/ijms232213833
APA StyleLima, R., Monteiro, A., Salgado, A. J., Monteiro, S., & Silva, N. A. (2022). Pathophysiology and Therapeutic Approaches for Spinal Cord Injury. International Journal of Molecular Sciences, 23(22), 13833. https://doi.org/10.3390/ijms232213833