


Multiplex Analysis of Serum Cytokine Profiles in Systemic Lupus Erythematosus and Multiple Sclerosis

 ,
,  , , , , , , , and
, , , , , , , and
Abstract
1. Introduction
2. Results
2.1. Clinical Characteristics of Analyzed Groups
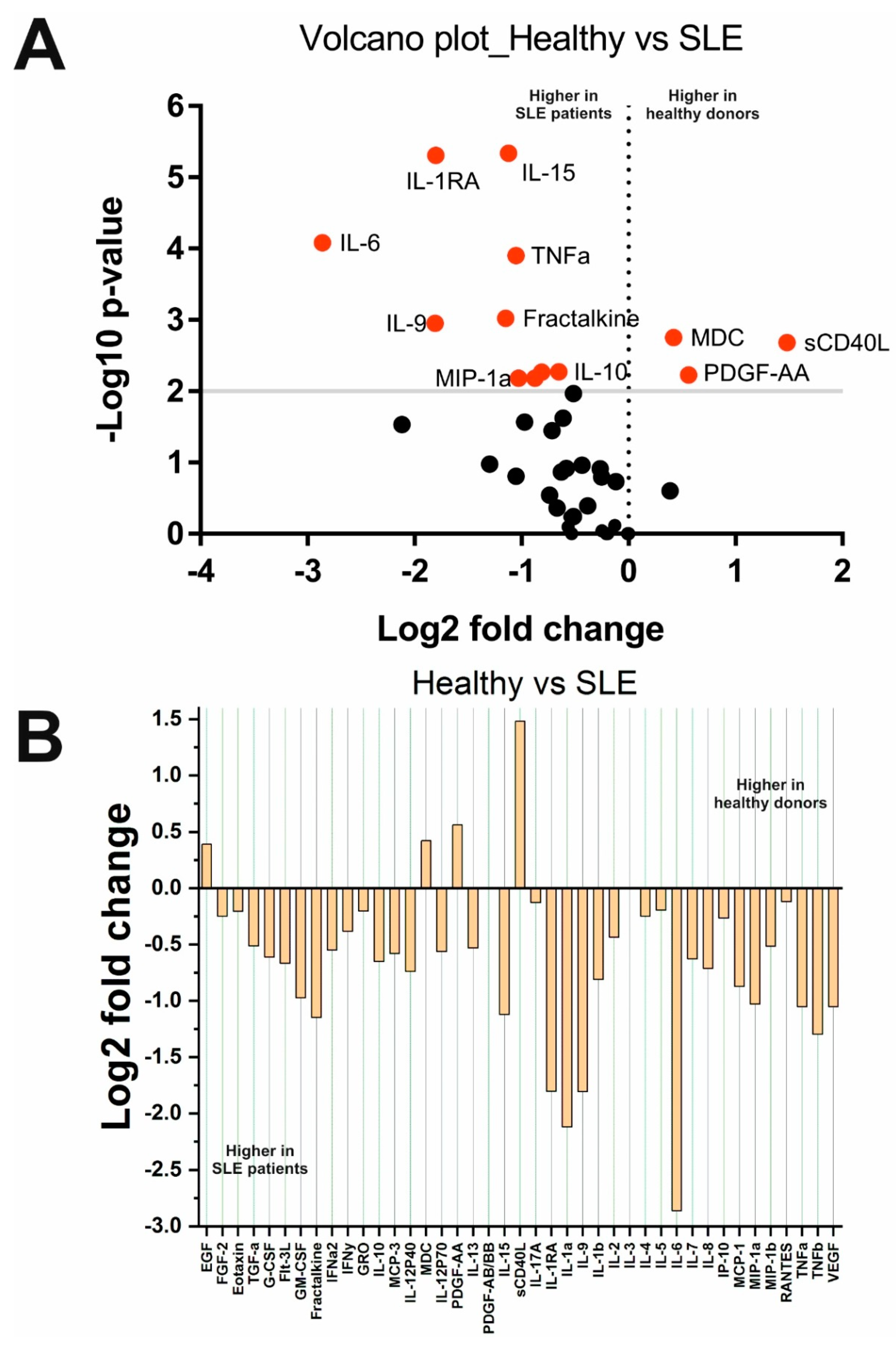
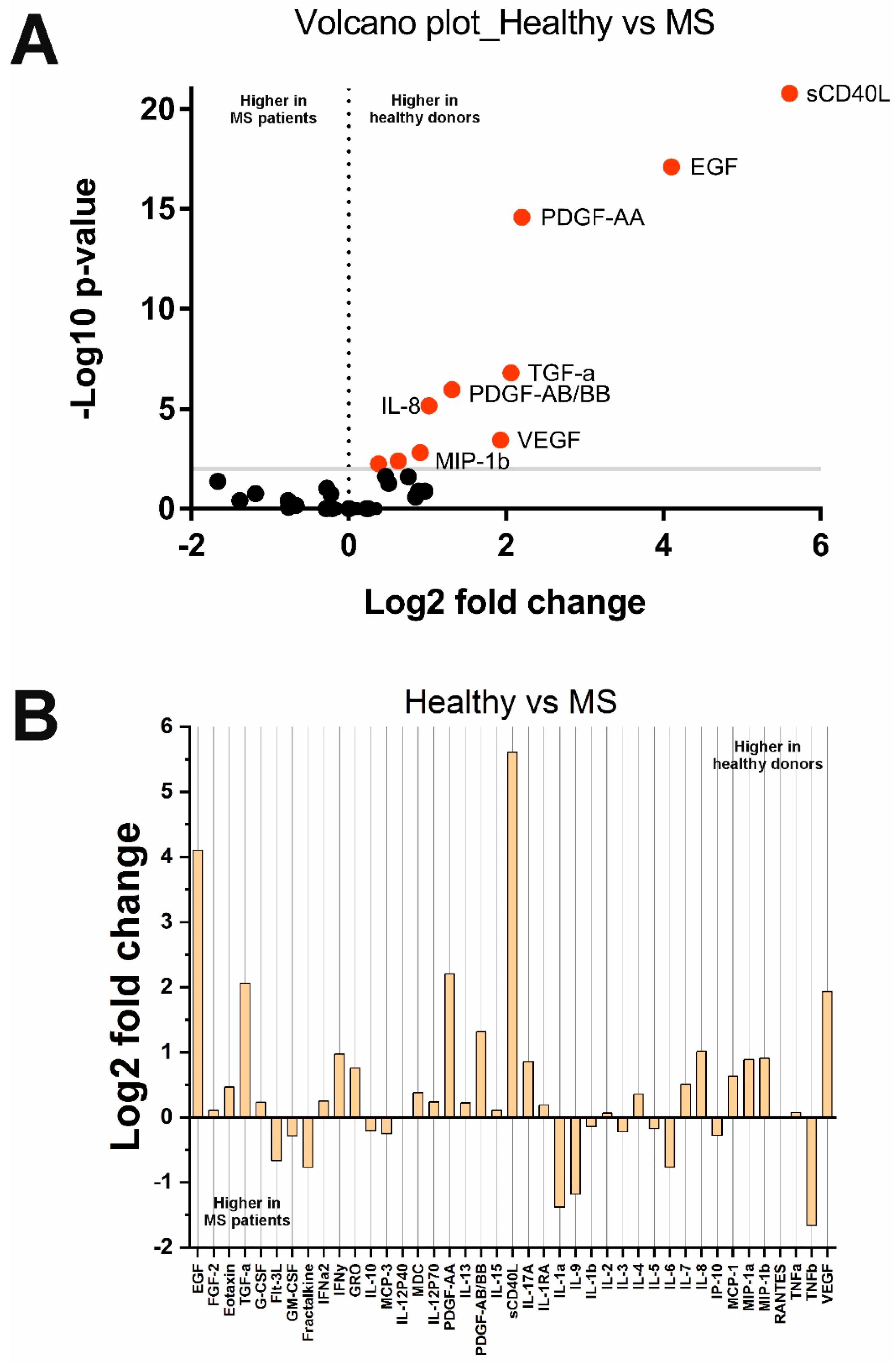
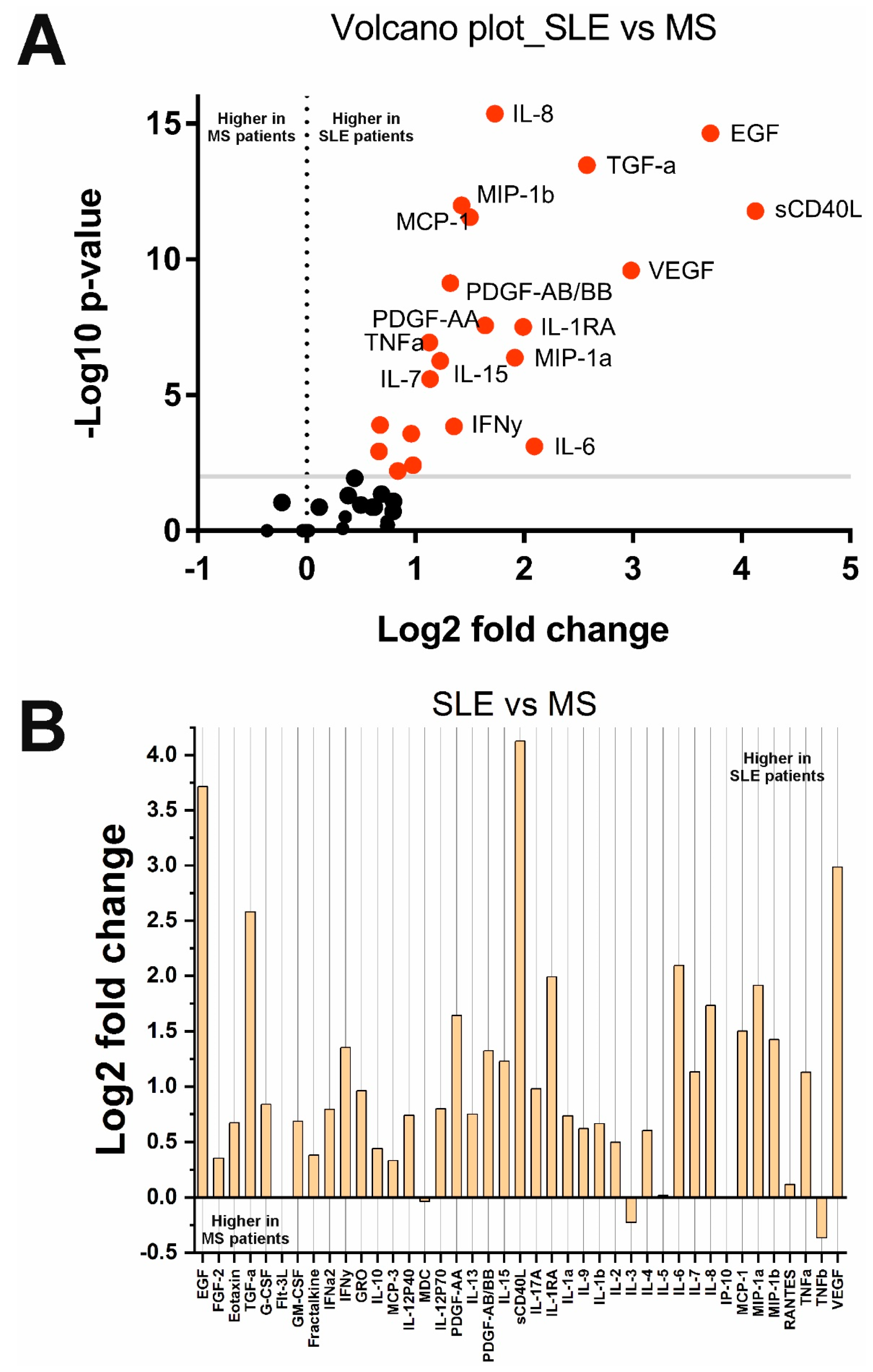
2.2. Changes in Cytokine Concentrations in the Analyzed Groups
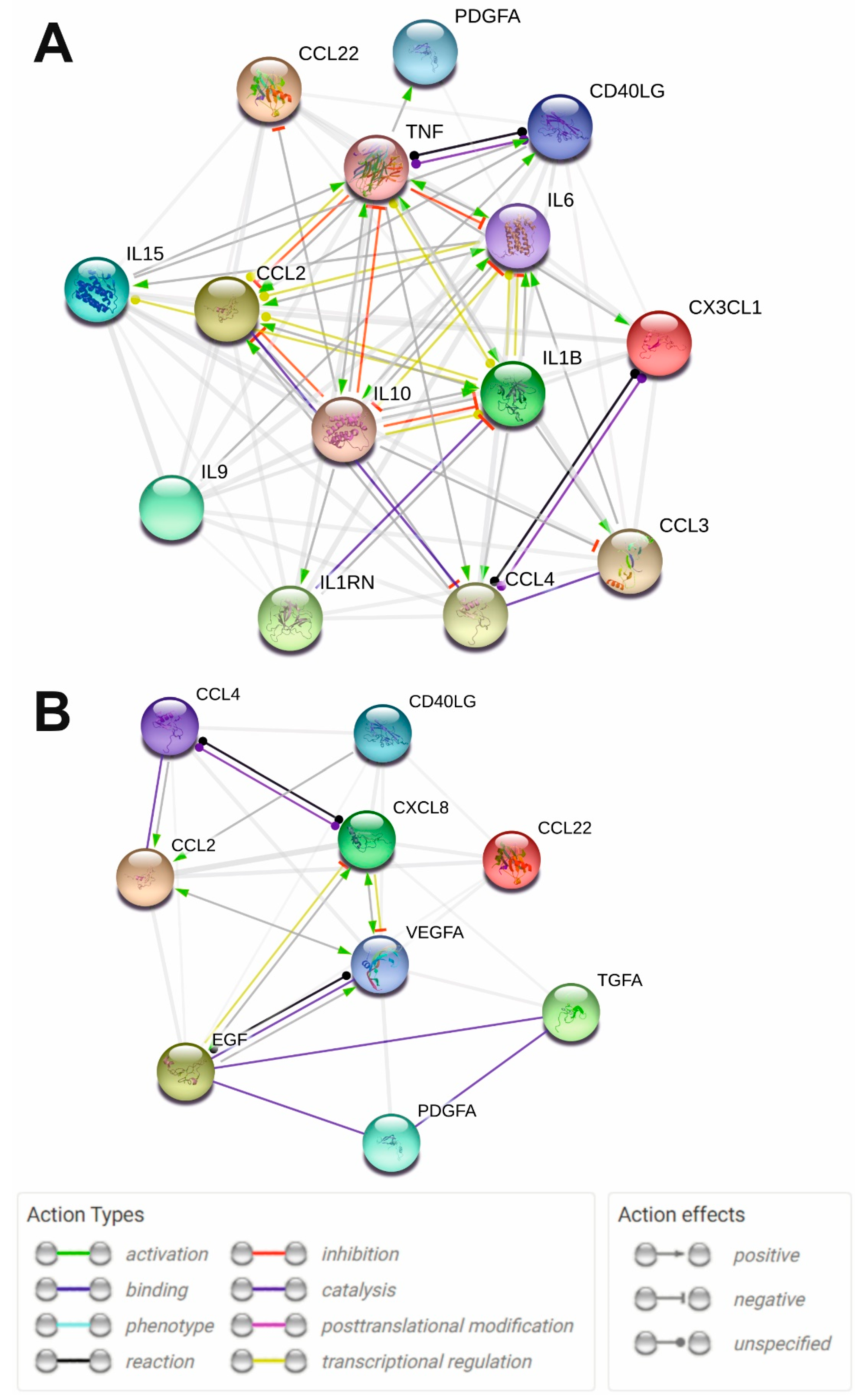
2.3. Protein Interaction Network Analysis of the Significantly Altered Cytokines in MS and SLE
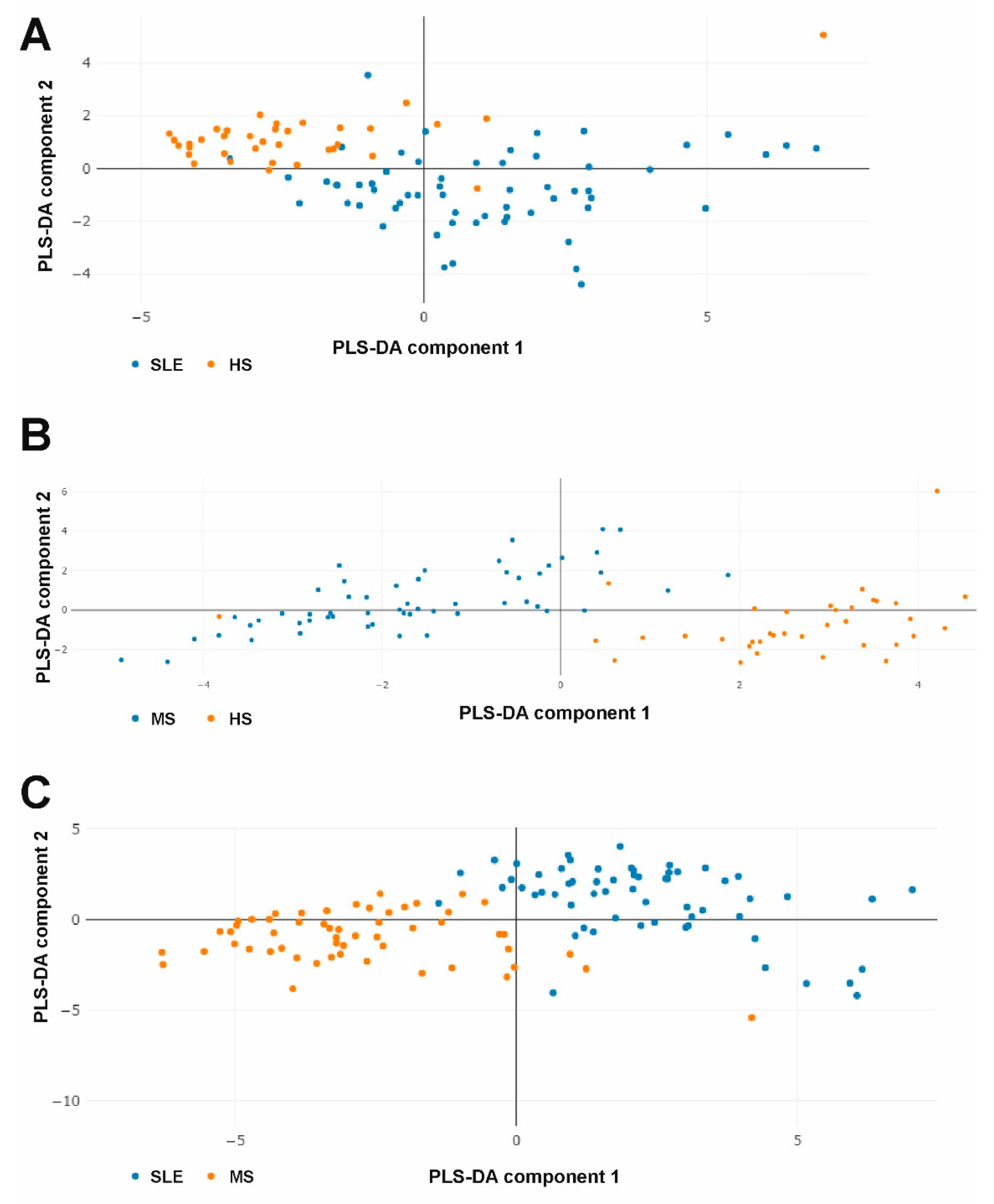
2.4. The Combined Multicytokine Profiles Are Different
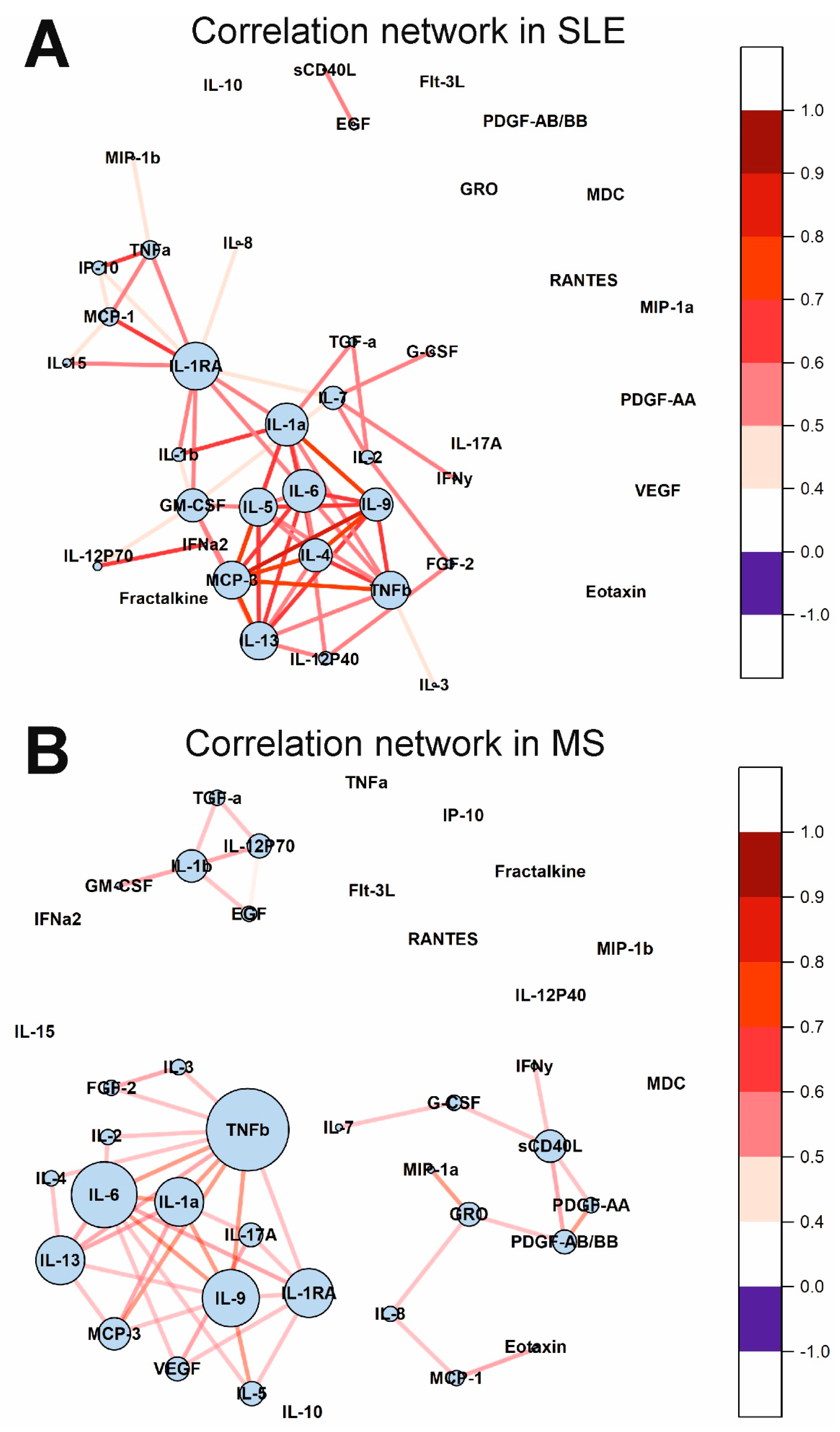
2.5. Cytokine Correlation Networks and Correlation Clusters
3. Discussion
3.1. Cytokine Profiles in SLE
3.2. Cytokine Profiles in MS
3.3. Cross-Disorder Analysis
3.4. Limitations
4. Materials and Methods
4.1. Design of the Study
4.2. Patients and Healthy Subjects
4.3. Collection of Serum Samples
4.4. Multiplex Serum Cytokine Immunoassay
4.5. Statistical Analysis
4.6. Protein-Protein Interaction Network Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviation | Description | Family |
|---|---|---|
| EGF | Epidermal growth factor | Growth factors |
| Eotaxin/CCL11 | C-C motif chemokine ligand 11, eosinophil chemotactic protein | Chemokines |
| FGF2 | Fibroblast growth factor 2 or Basic fibroblast growth factor | Growth factors |
| Flt-3L | Fms-related tyrosine kinase 3 ligand | Growth factors |
| Fractalkine/CX3CL1 | Fractalkine, C-X3-C motif chemokine ligand 1 | Chemokines |
| G-CSF | Granulocyte colony-stimulating factor | Growth factors |
| GM-CSF | Granulocyte-macrophage colony-stimulating factor | Growth factors |
| GRO/CXCL1 | Growth-regulated alpha protein, C-X-C motif chemokine ligand 1 | Chemokines |
| IFNa2 | Interferon α2 | Interferons |
| IFNy | Interferon γ | Interferons |
| IL-1a | Interleukin-1α | Interleukins |
| IL-1b | Interleukin-1β | Interleukins |
| IL-1RA | Interleukin-1 receptor antagonist | Interleukins |
| IL-2 | Interleukin-2 | Interleukins |
| IL-3 | Interleukin-3 | Interleukins |
| IL-4 | Interleukin-4 | Interleukins |
| IL-5 | Interleukin-5 | Interleukins |
| IL-6 | Interleukin-6 | Interleukins |
| IL-7 | Interleukin-7 | Interleukins |
| IL-8/CXCL8 | Interleukin-8, C-X-C motif chemokine ligand 8 | Interleukins/Chemokines |
| IL-9 | Interleukin-9 | Interleukins |
| IL-10 | Interleukin-10 | Interleukins |
| IL-12P40 | Subunit beta of interleukin 12, natural killer cell stimulatory factor 2, cytotoxic lymphocyte maturation factor p40, interleukin-12 subunit p40 | Interleukins |
| IL-12P70 | Interleukin-12 consisting of two covalently linked subunits p40 and p35 | Interleukins |
| IL-13 | Interleukin-13 | Interleukins |
| IL-15 | Interleukin-15 | Interleukins |
| IL-17A | Interleukin-17A | Interleukins |
| IP-10/CXCL10 | Interferon gamma-induced protein 10, C-X-C motif chemokine ligand 10 | Chemokines |
| MCP-1/CCL2 | Monocyte chemoattractant protein 1, C-C motif chemokine ligand 2 | Chemokines |
| MCP-3/CCL7 | Monocyte-chemotactic protein 3, C-C motif chemokine 7 | Chemokines |
| MDC/CCL22 | C-C motif chemokine 22, macrophage-derived chemokine | Chemokines |
| MIP-1a/CCL3 | Macrophage inflammatory protein-1α, C-C motif chemokine ligand 3 | Chemokines |
| MIP-1b/CCL4 | Macrophage inflammatory protein-1β, C-C motif chemokine ligand 4 | Chemokines |
| PDGF-AA | Platelet-derived growth factor AA | Growth factors |
| PDGF-AB/BB | Platelet-derived growth factor AB/BB | Growth factors |
| RANTES/CCL5 | Regulated on activation, normal T cell expressed and secreted (RANTES); C-C motif chemokine ligand 5 | Chemokines |
| sCD40L | Soluble CD-40 ligand, tumor necrosis factor ligand superfamily member 5 | TNF superfamily members |
| TGF-a | Transforming growth factor α | Growth factors |
| TNFα | Tumor necrosis factor α | TNF superfamily members |
| TNFb | Tumor necrosis factor β | TNF superfamily members |
| VEGF | Vascular endothelial growth factor | Growth factors |
References
- Kaul, A.; Gordon, C.; Crow, M.K.; Touma, Z.; Urowitz, M.B.; van Vollenhoven, R.; Ruiz-Irastorza, G.; Hughes, G. Systemic lupus erythematosus. Nat. Rev. Dis. Primers. 2016, 2, 16039. [Google Scholar] [CrossRef] [PubMed]
- Filippi, M.; Bar-Or, A.; Piehl, F.; Preziosa, P.; Solari, A.; Vukusic, S.; Rocca, M.A. Multiple sclerosis. Nat. Rev. Dis. Primers. 2018, 4, 43. [Google Scholar] [CrossRef] [PubMed]
- Barber, M.R.W.; Drenkard, C.; Falasinnu, T.; Hoi, A.; Mak, A.; Kow, N.Y.; Svenungsson, E.; Peterson, J.; Clarke, A.E.; Ramsey-Goldman, R. Global epidemiology of systemic lupus erythematosus. Nat. Rev. Rheumatol. 2021, 17, 515–532. [Google Scholar] [CrossRef] [PubMed]
- Alton, C.; King, R.; Rechtman, L.; Kaye, W.; Leray, E.; Marrie, R.A.; Robertson, N.; La Rocca, N.; Uitdehaag, B.; van der Mei, I.; et al. Rising prevalence of multiple sclerosis worldwide: Insights from the Atlas of MS. Mult. Scler. 2020, 26, 1816–1821. [Google Scholar] [CrossRef] [PubMed]
- Lessard, C.J.; Ice, J.A.; Adrianto, I.; Wiley, G.B.; Kelly, J.A.; Gaffney, P.M.; Montgomery, C.G.; Moser, K.L. The genomics of autoimmune disease in the era of genome-wide association studies and beyond. Autoimmun. Rev. 2012, 11, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Richard-Miceli, C.; Criswell, L.A. Emerging patterns of genetic overlap across autoimmune disorders. Genome Med. 2012, 4, 6. [Google Scholar] [CrossRef] [PubMed]
- Benedict, R.H.B.; Shucard, J.L.; Zivadinov, R.; Shucard, D.W. Neuropsychological impairment in systemic lupus erythematosus: A comparison with multiple sclerosis. Neuropsychol. Rev. 2008, 18, 149–166. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Covey, T.J.; Shucard, J.L.; Shucard, D.W.; Stegen, S.; Benedict, R.H. Comparison of neuropsychological impairment and vocational outcomes in systemic lupus erythematosus and multiple sclerosis patients. J. Int. Neuropsychol. Soc. 2012, 18, 530–540. [Google Scholar] [CrossRef]
- Baraczka, K.; Nékám, K.; Pozsonyi, T.; Jakab, L.; Szongoth, M.; Seszták, M. Concentration of soluble adhesion molecules (sVCAM-1, sICAM-1 and sL-selectin) in the cerebrospinal fluid and serum of patients with multiple sclerosis and systemic lupus erythematosus with central nervous involvement. Neuroimmunomodulation 2001, 9, 49–54. [Google Scholar] [CrossRef]
- Szymczak, F.; Colli, M.L.; Mamula, M.J.; Evans-Molina, C.; Eizirik, D.L. Gene expression signatures of target tissues in type 1 diabetes, lupus erythematosus, multiple sclerosis, and rheumatoid arthritis. Sci. Adv. 2021, 7, eabd7600. [Google Scholar] [CrossRef]
- Lossius, A.; Johansen, J.N.; Torkildsen, Ø.; Vartdal, F.; Holmøy, T. Epstein-Barr virus in systemic lupus erythematosus, rheumatoid arthritis and multiple sclerosis—association and causation. Viruses 2012, 4, 3701–3730. [Google Scholar] [CrossRef] [PubMed]
- Sundar, K.; Jacques, S.; Gottlieb, P.; Villars, R.; Benito, M.E.; Taylor, D.K.; Spatz, L.A. Expression of the Epstein-Barr virus nuclear antigen-1 (EBNA-1) in the mouse can elicit the production of anti-dsDNA and anti-Sm antibodies. J. Autoimmun. 2004, 23, 127–140. [Google Scholar] [CrossRef] [PubMed]
- Lanz, T.V.; Brewer, R.C.; Ho, P.P.; Moon, J.S.; Jude, K.M.; Fernandez, D.; Fernandes, R.A.; Gomez, A.M.; Nadj, G.S.; Bartley, C.M.; et al. Clonally expanded B cells in multiple sclerosis bind EBV EBNA1 and GlialCAM. Nature 2022, 603, 321–327. [Google Scholar] [CrossRef]
- Liu, C.; Chu, D.; Kalantar-Zadeh, K.; George, J.; Young, H.A.; Liu, G. Cytokines: From Clinical Significance to Quantification. Adv. Sci. 2021, 8, e2004433. [Google Scholar] [CrossRef] [PubMed]
- Talaat, R.M.; Mohamed, S.F.; Bassyouni, I.H.; Raouf, A.A. Th1/Th2/Th17/Treg cytokine imbalance in systemic lupus erythematosus (SLE) patients: Correlation with disease activity. Cytokine 2015, 72, 146–153. [Google Scholar] [CrossRef]
- Svenungsson, E.; Fei, G.Z.; Jensen-Urstad, K.; de Faire, U.; Hamsten, A.; Frostegard, J. TNF-α: A link between hypertriglyceridaemia and inflammation in SLE patients with cardiovascular disease. Lupus 2003, 12, 454–461. [Google Scholar] [CrossRef]
- Melamud, M.M.; Ermakov, E.A.; Boiko, A.S.; Parshukova, D.A.; Sizikov, A.E.; Ivanova, S.A.; Nevinsky, G.A.; Buneva, V.N. Serum cytokine levels of systemic lupus erythematosus patients in the presence of concomitant cardiovascular diseases. Endocr. Metab. Immune Disord. Drug Targets 2022, 22, 852–861. [Google Scholar] [CrossRef]
- Bolouri, N.; Akhtari, M.; Farhadi, E.; Mansouri, R.; Faezi, S.T.; Jamshidi, A.; Mahmoudi, M. Role of the innate and adaptive immune responses in the pathogenesis of systemic lupus erythematosus. Inflammat. Res. 2022, 71, 537–554. [Google Scholar] [CrossRef]
- Tsokos, G.C.; Lo, M.S.; Reis, P.C.; Sullivan, K.E. New insights into the immunopathogenesis of systemic lupus erythematosus. Nat. Rev. Rheumatol. 2016, 12, 716–730. [Google Scholar] [CrossRef]
- Dong, C.; Fu, T.; Ji, J.; Li, Z.; Gu, Z. The role of interleukin-4 in rheumatic diseases. Clin. Exp. Pharmacol. Physiol. 2018, 45, 747–754. [Google Scholar] [CrossRef]
- Robinson, E.S.; Werth, V.P. The role of cytokines in the pathogenesis of cutaneous lupus erythematosus. Cytokine 2015, 73, 326–334. [Google Scholar] [CrossRef] [PubMed]
- Göbel, K.; Ruck, T.; Meuth, S.G. Cytokine signaling in multiple sclerosis: Lost in translation. Mult. Scler. J. 2018, 24, 432–439. [Google Scholar] [CrossRef] [PubMed]
- Wagner, C.A.; Roqué, P.J.; Goverman, J.M. Pathogenic T cell cytokines in multiple sclerosis. J. Exp. Med. 2020, 217, e20190460. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.Y.; Chu, S.F.; Chen, N.H. The role of chemokines and chemokine receptors in multiple sclerosis. Int. Immunopharmacol. 2020, 83, 106314. [Google Scholar] [CrossRef]
- Proescholdt, M.A.; Jacobson, S.; Tresser, N.; Oldfield, E.H.; Merrill, M.J. Vascular endothelial growth factor is expressed in multiple sclerosis plaques and can induce inflammatory lesions in experimental allergic encephalomyelitis rats. J. Neuropathol. Exp. Neurol. 2002, 61, 914–925. [Google Scholar] [CrossRef]
- Scalabrino, G. Epidermal growth factor in the CNS: A beguiling journey from integrated cell biology to multiple sclerosis. an extensive translational overview. Cell. Mol. Neurobiol. 2020, 42, 891–916. [Google Scholar] [CrossRef]
- Yan, C.; Yu, L.; Zhang, X.L.; Shang, J.J.; Ren, J.; Fan, J.; Feng, X.Q.; Zhang, R.W.; Xia, Z.B.; Duan, X.W. Cytokine profiling in Chinese SLE patients: Correlations with renal dysfunction. J. Immunol. Res. 2020, 2020, 8146502. [Google Scholar] [CrossRef]
- Taşdemir, N.; Karaca, E.E.; Ece, A.; Yücel, Y.; Dikici, S.; Taşdemir, M.S. Multiple sclerosis: Relationships between cytokines, MRI lesion burden, visual evoked potentials and disability scores. Eur. J. Gen. Med. 2010, 7, 167–173. [Google Scholar] [CrossRef]
- Ortiz, G.G.; Pacheco-Moisés, F.P.; Bitzer-Quintero, O.K.; Ramírez-Anguiano, A.C.; Flores-Alvarado, L.J.; Ramírez-Ramírez, V.; Macias-Islas, M.A.; Torres-Sánchez, E.D. Immunology and oxidative stress in multiple sclerosis: Clinical and basic approach. Clin. Dev. Immunol. 2013, 2013, 708659. [Google Scholar] [CrossRef]
- Park, J.; Jang, W.; Park, H.S.; Park, K.H.; Kwok, S.K.; Park, S.H.; Oh, E.J. Cytokine clusters as potential diagnostic markers of disease activity and renal involvement in systemic lupus erythematosus. J. Int. Med. Res. 2020, 48, 300060520926882. [Google Scholar] [CrossRef]
- Andrés-Rodríguez, L.; Borràs, X.; Feliu-Soler, A.; Pérez-Aranda, A.; Rozadilla-Sacanell, A.; Arranz, B.; Montero-Marin, J.; García-Campayo, J.; Angarita-Osorio, N.; Maes, M.; et al. Machine learning to understand the immune-inflammatory pathways in fibromyalgia. Int. J. Mol. Sci. 2019, 20, 4231. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Zhao, J.; Cai, T.; Stephens, A.; Su, S.H.; Sandford, E.; Flora, C.; Singer, B.H.; Ghosh, M.; Choi, S.W.; et al. Machine learning-based cytokine microarray digital immunoassay analysis. Biosens. Bioelectron. 2021, 180, 113088. [Google Scholar] [CrossRef] [PubMed]
- Goyal, M.; Khanna, D.; Rana, P.S.; Khaibullin, T.; Martynova, E.; Rizvanov, A.A.; Khaiboullina, S.F.; Baranwal, M. Computational intelligence technique for prediction of multiple sclerosis based on serum cytokines. Front. Neurol. 2019, 10, 781. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein–protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef]
- Kassem, O.; Al-Saleh, A.; Azizieh, F.; Dingle, K. CytokineExplore: An online tool for statistical analysis of cytokine concentration datasets. J. Inflamm. Res. 2020, 13, 401–410. [Google Scholar] [CrossRef]
- Fruchterman, T.M.; Reingold, E.M. Grapg drawing by force-directed placement. Softw. Pract. Exp. 1991, 21, 1129–1164. [Google Scholar] [CrossRef]
- Parodis, I.; Åkerström, E.; Sjöwall, C.; Sohrabian, A.; Jönsen, A.; Gomez, A.; Frodlund, M.; Zickert, A.; Bengtsson, A.A.; Rönnelid, J.; et al. Autoantibody and cytokine profiles during treatment with belimumab in patients with systemic lupus erythematosus. Int. J. Mol. Sci. 2020, 21, 3463. [Google Scholar] [CrossRef]
- Reske, D.; Thomas, A.V.; Petereit, H.F.; Fink, G.R.; Schroeter, M. Impact of immunomodulatory treatment on leukocyte cytokine production in multiple sclerosis patients and healthy donors. Neuroimmunomodulation 2009, 16, 385–391. [Google Scholar] [CrossRef]
- Idborg, H.; Eketjäll, S.; Pettersson, S.; Gustafsson, J.T.; Zickert, A.; Kvarnström, M.; Oke, V.; Jakobsson, P.J.; Gunnarsson, I.; Svenungsson, E. TNF-α and plasma albumin as biomarkers of disease activity in systemic lupus erythematosus. Lupus Sci. Med. 2018, 51, e000260. [Google Scholar] [CrossRef]
- Godsell, J.; Rudloff, I.; Kandane-Rathnayake, R.; Hoi, A.; Nold, M.F.; Morand, E.F.; Harris, J. Clinical associations of IL-10 and IL-37 in systemic lupus erythematosus. Sci. Rep. 2016, 6, 34604. [Google Scholar] [CrossRef]
- Ding, J.; Su, S.; You, T.; Xia, T.; Lin, X.; Chen, Z.; Zhang, L. Serum interleukin-6 level is correlated with the disease activity of systemic lupus erythematosus: A meta-analysis. Clinics 2020, 75, e1801. [Google Scholar] [CrossRef] [PubMed]
- Leng, R.X.; Pan, H.F.; Ye, D.Q.; Xu, Y. Potential roles of IL-9 in the pathogenesis of systemic lupus erythematosus. Am. J. Clin. Exp. Immunol. 2012, 1, 28. [Google Scholar] [PubMed]
- Baranda, L.; de la Fuente, H.; Layseca-Espinosa, E.; Portales-Pérez, D.; Niño-Moreno, P.; Valencia-Pacheco, G.; Abud-Mendoza, C.; Alcocer-Varela, J.; González-Amaro, R. IL-15 and IL-15R in leucocytes from patients with systemic lupus erythematosus. Rheumatology 2005, 44, 1507–1513. [Google Scholar] [CrossRef] [PubMed]
- Vakkalanka, R.K.; Woo, C.; Kirou, K.A.; Koshy, M.; Berger, D.; Crow, M.K. Elevated levels and functional capacity of soluble CD40 ligand in systemic lupus erythematosus sera. Arthritis Rheum. 1999, 42, 871–881. [Google Scholar] [CrossRef]
- Menten, P.; Wuyts, A.; Van Damme, J. Macrophage inflammatory protein-1. Cytokine Growth Factor Rev. 2002, 13, 455–481. [Google Scholar] [CrossRef]
- Richter, J.R.; Sutton, J.M.; Belizaire, R.M.; Friend, L.A.; Schuster, R.M.; Johannigman, T.A.; Miller, S.G.; Lentsch, A.B.; Pritts, T.A. Macrophage-derived chemokine (MDC/CCL22) is a novel mediator of lung inflammation following hemorrhage and resuscitation. Shock 2014, 42, 525–531. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Shahir, M.; Taheri, M.; Salimi, A. A review on the role of chemokines in the pathogenesis of systemic lupus erythematosus. Cytokine 2021, 146, 155640. [Google Scholar] [CrossRef]
- Geneva-Popova, M.G.; Popova-Belova, S.D.; Gardzheva, P.N.; Kraev, K.I. A Study of IFN-α-Induced Chemokines CCL2, CXCL10 and CCL19 in Patients with Systemic Lupus Erythematosu. Life 2022, 12, 251. [Google Scholar] [CrossRef]
- Yajima, N.; Kasama, T.; Isozaki, T.; Odai, T.; Matsunawa, M.; Negishi, M.; Ide, H.; Kameoka, Y.; Hirohata, S.; Adachi, M. Elevated levels of soluble fractalkine in active systemic lupus erythematosus: Potential involvement in neuropsychiatric manifestations. Arthritis Rheum. 2005, 52, 1670–1675. [Google Scholar] [CrossRef]
- Rönnblom, L.; Leonard, D. Interferon pathway in SLE: One key to unlocking the mystery of the disease. Lupus Sci. Med. 2019, 6, e000270. [Google Scholar] [CrossRef]
- Lesiak, A.; Sysa-Jedrzejowska, A.; Narbutt, J.; Lukamowicz, J.; Robak, E.; Wozniacka, A. Proinflammatory cytokines in inactive lupus erythematosus patients. Przegl. Lek. 2005, 62, 838–842. [Google Scholar] [PubMed]
- Maczynska, I.; Millo, B.; Ratajczak-Stefańska, V.; Maleszka, R.; Szych, Z.; Kurpisz, M.; Giedrys-Kalemba, S. Proinflammatory cytokine (IL-1β, IL-6, IL-12, IL-18 and TNF-α) levels in sera of patients with subacute cutaneous lupus erythematosus (SCLE). Immunol. Lett. 2006, 102, 79–82. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, E.M.; Smith, S.; Lee, R.Z.; Cunnane, G.; Doran, M.F.; Donnelly, S.; Howard, D.; O’Connell, P.; Kearns, G.; Ní Gabhann, J.; et al. The association of cytokines with disease activity and damage scores in systemic lupus erythematosus patients. Rheumatology 2014, 53, 1586–1594. [Google Scholar] [CrossRef] [PubMed]
- Umare, V.; Pradhan, V.; Nadkar, M.; Rajadhyaksha, A.; Patwardhan, M.; Ghosh, K.K.; Nadkarni, A.H. Effect of proinflammatory cytokines (IL-6, TNF-α, and IL-1β) on clinical manifestations in Indian SLE patients. Mediat. Inflamm. 2014, 2014, 385297. [Google Scholar] [CrossRef] [PubMed]
- Scalabrino, G. New Epidermal-Growth-Factor-Related Insights Into the Pathogenesis of Multiple Sclerosis: Is It Also Epistemology? Front. Neurol. 2021, 12, 754270. [Google Scholar] [CrossRef]
- Girolamo, F.; Coppola, C.; Ribatti, D.; Trojano, M. Angiogenesis in multiple sclerosis and experimental autoimmune encephalomyelitis. Acta Neuropathol. Commun. 2014, 2, 84. [Google Scholar] [CrossRef]
- Bassi, M.S.; Iezzi, E.; Marfia, G.A.; Simonelli, I.; Musella, A.; Mandolesi, G.; Fresegna, D.; Pasqualetti, P.; Furlan, R.; Finardi, A.; et al. Platelet-derived growth factor predicts prolonged relapse-free period in multiple sclerosis. J. Neuroinflammation 2018, 15, 108. [Google Scholar] [CrossRef]
- Mori, F.; Rossi, S.; Piccinin, S.; Motta, C.; Mango, D.; Kusayanagi, H.; Bergami, A.; Studer, V.; Nicoletti, C.G.; Buttari, F.; et al. Synaptic plasticity and PDGF signaling defects underlie clinical progression in multiple sclerosis. J. Neurosci. 2013, 33, 19112–19119. [Google Scholar] [CrossRef]
- Woodruff, R.H.; Fruttiger, M.; Richardson, W.D.; Franklin, R.J. Platelet-derived growth factor regulates oligodendrocyte progenitor numbers in adult CNS and their response following CNS demyelination. Mol. Cell. Neurosci. 2004, 25, 252–262. [Google Scholar] [CrossRef]
- Reder, A.T.; Feng, X. How type I interferons work in multiple sclerosis and other diseases: Some unexpected mechanisms. J. Interferon. Cytokine Res. 2014, 34, 589–599. [Google Scholar] [CrossRef]
- Cirac, A.; Tsaktanis, T.; Beyer, T.; Linnerbauer, M.; Andlauer, T.; Grummel, V.; Nirschl, L.; Loesslein, L.; Quintana, F.J.; Hemmer, B.; et al. The Aryl Hydrocarbon Receptor–Dependent TGF-α/VEGF-B Ratio Correlates With Disease Subtype and Prognosis in Multiple Sclerosis. Neurol. Neuroimmunol. Neuroinflamm. 2021, 8, e1043. [Google Scholar] [CrossRef] [PubMed]
- Sindern, E.; Niederkinkhaus, Y.; Henschel, M.; Ossege, L.M.; Patzold, T.; Malin, J.P. Differential release of β-chemokines in serum and CSF of patients with relapsing–remitting multiple sclerosis. Acta Neurol. Scand. 2001, 104, 88–91. [Google Scholar] [CrossRef]
- Estevao, C.; Bowers, C.E.; Luo, D.; Sarker, M.; Hoeh, A.E.; Frudd, K.; Turowski, P.; Greenwood, J. CCL4 induces inflammatory signalling and barrier disruption in the neurovascular endothelium. Brain Behav. Immun. Health 2021, 18, 100370. [Google Scholar] [CrossRef]
- Khaibullin, T.; Ivanova, V.; Martynova, E.; Cherepnev, G.; Khabirov, F.; Granatov, E.; Rizvanov, A.; Khaiboullina, S. Elevated levels of proinflammatory cytokines in cerebrospinal fluid of multiple sclerosis patients. Front. Immunol. 2017, 8, 531. [Google Scholar] [CrossRef] [PubMed]
- Jafarzadeh, A.; Ebrahimi, H.A.; Bagherzadeh, S.; Zarkesh, F.; Iranmanesh, F.; Najafzadeh, A.; Khosravimashizi, A.; Nemati, M.; Sabahi, A.; Hajghani, H.; et al. Lower serum levels of Th2-related chemokine CCL22 in women patients with multiple sclerosis: A comparison between patients and healthy women. Inflammation 2014, 37, 604–610. [Google Scholar] [CrossRef]
- Du, L.; Chang, H.; Wei, Y.; Zhang, X.; Yin, L. Different roles of soluble CD40 ligand in central nervous system damage. Neurol. Res. 2020, 42, 372–378. [Google Scholar] [CrossRef]
- Aloui, C.; Prigent, A.; Sut, C.; Tariket, S.; Hamzeh-Cognasse, H.; Pozzetto, B.; Richard, Y.; Cognasse, F.; Laradi, S.; Garraud, O. The signaling role of CD40 ligand in platelet biology and in platelet component transfusion. Int. J. Mol. Sci. 2014, 15, 22342–22364. [Google Scholar] [CrossRef] [PubMed]
- Orian, J.M.; D’Souza, C.S.; Kocovski, P.; Krippner, G.; Hale, M.W.; Wang, X.; Peter, K. Platelets in multiple sclerosis: Early and central mediators of inflammation and neurodegeneration and attractive targets for molecular imaging and site-directed therapy. Front. Immunol. 2021, 12, 620963. [Google Scholar] [CrossRef] [PubMed]
- Plant, S.R.; Arnett, H.A.; Ting, J.P. Astroglial-derived lymphotoxin-alpha exacerbates inflammation and demyelination, but not remyelination. Glia 2005, 49, 1–14. [Google Scholar] [CrossRef]
- World Medical Association. World Medical Association Declaration of Helsinki: Ethical principles for medical research involving human subjects. JAMA 2013, 310, 2191–2194. [Google Scholar] [CrossRef]
- Tamirou, F.; Arnaud, L.; Talarico, R.; Scirè, C.A.; Alexander, T.; Amoura, Z.; Avcin, T.; Bortoluzzi, A.; Cervera, R.; Conti, F.; et al. Systemic lupus erythematosus: State of the art on clinical practice guidelines. RMD Open. 2018, 4, e000793. [Google Scholar] [CrossRef] [PubMed]
- Ermakov, E.A.; Kabirova, E.M.; Sizikov, A.E.; Buneva, V.N.; Nevinsky, G.A. IgGs-Abzymes from the Sera of Patients with Systemic Lupus Erythematosus Hydrolyzed miRNAs. J. Inflamm. Res. 2020, 13, 681–699. [Google Scholar] [CrossRef] [PubMed]
- Petri, M.; Kim, M.Y.; Kalunian, K.C.; Grossman, J.; Hahn, B.H.; Sammaritano, L.R.; Lockshin, M.; Merrill, J.T.; Belmont, H.M.; Askanase, A.D.; et al. Combined oral contraceptives in women with systemic lupus erythematosus. N. Engl. J. Med. 2005, 353, 2550–2558. [Google Scholar] [CrossRef] [PubMed]
- Thompson, A.J.; Banwell, B.L.; Barkhof, F.; Carroll, W.M.; Coetzee, T.; Comi, G.; Correale, J.; Fazekas, F.; Filippi, M.; Freedman, M.S.; et al. Diagnosis of multiple sclerosis: 2017 revisions of the McDonald criteria. Lancet Neurol. 2018, 17, 162–173. [Google Scholar] [CrossRef]
- Kurtzke, J.F. Rating neurologic impairment in multiple sclerosis: An expanded disability status scale (EDSS). Neurology 1983, 33, 1444–1452. [Google Scholar] [CrossRef] [PubMed]







| Parameters | HS Group | SLE Group | MS Group | Statistics * |
|---|---|---|---|---|
| Number of participants | 36 | 60 | 56 | – |
| Age, years Me [Q1–Q3], (Min-Max) | 36.5 [28–58.25] (24–65) | 38.5 [28.8–59] (21–72) | 37 [26.8–43.3] (20–57) | N.S. |
| Sex, F/M, % | 50/50 | 83.3/16.7 | 52.7/47.3 | HS vs. SLE: p < 0.0001 HS vs. MS: N.S. SLE vs. MS: p < 0.0001 |
| Disease duration, years Me [Q1–Q3], (Min–Max) | – | 7 [4–16.3] (1–25) | 3 [1–7.8] (1–23) | SLE vs. MS: p = 0.0002 |
| SELENA-SLEDAI Index Score Me [Q1–Q3], (Min–Max) | – | 6.5 [4,5,6,7,8,9,10] (1–18) | – | – |
| Exacerbation: Yes/No, n | – | 51/9 | – | – |
| EDSS Index Score Me [Q1–Q3], (Min–Max) | – | – | 2.5 [1.5–3.5] (1–5.5) | – |
| MS type: PPMS/RRMS, n | – | – | 20/36 | – |
| MS phase: Stable/aggravation/ remission/N.D., n | – | – | 14 (PPMS)/16 (RRMS)/20 (RRMS)/6 (PPMS) | – |
| Patients received therapy, % | – | 100% | 0 | – |
| HS vs. SLE | HS vs. MS | SLE vs. MS | ||||||
|---|---|---|---|---|---|---|---|---|
| Cytokine * | p-Value | Comment | Cytokine * | p-Value | Comment | Cytokine * | p-Value | Comment |
| sCD40L | 0.002 | Decreased in SLE | sCD40L | <0.0001 | Decreased in MS | sCD40L | <0.0001 | Decreased in MS |
| MCP-1/CCL2 | 0.007 | Increased in SLE | MCP-1/CCL2 | 0.004 | Decreased in MS | MCP-1/CCL2 | <0.0001 | Decreased in MS |
| MIP-1b/CCL4 | 0.01 | Increased in SLE | MIP-1b/CCL4 | 0.001 | Decreased in MS | MIP-1b/CCL4 | <0.0001 | Decreased in MS |
| PDGF-AA | 0.006 | Decreased in SLE | PDGF-AA | <0.0001 | Decreased in MS | PDGF-AA | <0.0001 | Decreased in MS |
| MDC/CCL22 | 0.002 | Decreased in SLE | MDC/CCL22 | 0.005 | Decreased in MS | IL-1b | 0.001 | Decreased in MS |
| IL-1b | 0.005 | Increased in SLE | EGF | <0.0001 | Decreased in MS | IL-1RA | <0.0001 | Decreased in MS |
| IL-1RA | <0.0001 | Increased in SLE | IL-8/CXCL8 | <0.0001 | Decreased in MS | IL-6 | 0.0008 | Decreased in MS |
| IL-6 | <0.0001 | Increased in SLE | TGF-a | <0.0001 | Decreased in MS | IL-15 | <0.0001 | Decreased in MS |
| IL-15 | <0.0001 | Increased in SLE | PDGF-AB/BB | <0.0001 | Decreased in MS | EGF | <0.0001 | Decreased in MS |
| MIP-1b/CCL3 | 0.007 | Increased in SLE | VEGF | 0.0003 | Decreased in MS | MIP-1b/CCL4 | <0.0001 | Decreased in MS |
| IL-10 | 0.005 | Increased in SLE | – | – | – | IL-10 | 0.01 | Decreased in MS |
| TNFa | <0.001 | Increased in SLE | – | – | – | TNFa | <0.0001 | Decreased in MS |
| Fractalkine/CX3CL1 | <0.001 | Increased in SLE | – | – | – | IL-8/CXCL8 | <0.0001 | Decreased in MS |
| IL-9 | 0.001 | Increased in SLE | – | – | – | PDGF-AB/BB | <0.0001 | Decreased in MS |
| – | – | – | – | – | – | TGF-a | <0.0001 | Decreased in MS |
| – | – | – | – | – | – | VEGF | <0.0001 | Decreased in MS |
| – | – | – | – | – | – | Eotaxin/CCL11 | 0.0001 | Decreased in MS |
| – | – | – | – | – | – | G-CSF | 0.006 | Decreased in MS |
| – | – | – | – | – | – | GRO/CXCL1 | 0.0003 | Decreased in MS |
| – | – | – | – | – | – | IFNy | 0.0001 | Decreased in MS |
| – | – | – | – | – | – | IL-7 | <0.0001 | Decreased in MS |
| – | – | – | – | – | – | IL-17A | 0.004 | Decreased in MS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Melamud, M.M.; Ermakov, E.A.; Boiko, A.S.; Kamaeva, D.A.; Sizikov, A.E.; Ivanova, S.A.; Baulina, N.M.; Favorova, O.O.; Nevinsky, G.A.; Buneva, V.N. Multiplex Analysis of Serum Cytokine Profiles in Systemic Lupus Erythematosus and Multiple Sclerosis. Int. J. Mol. Sci. 2022, 23, 13829. https://doi.org/10.3390/ijms232213829
Melamud MM, Ermakov EA, Boiko AS, Kamaeva DA, Sizikov AE, Ivanova SA, Baulina NM, Favorova OO, Nevinsky GA, Buneva VN. Multiplex Analysis of Serum Cytokine Profiles in Systemic Lupus Erythematosus and Multiple Sclerosis. International Journal of Molecular Sciences. 2022; 23(22):13829. https://doi.org/10.3390/ijms232213829
Chicago/Turabian StyleMelamud, Mark M., Evgeny A. Ermakov, Anastasiia S. Boiko, Daria A. Kamaeva, Alexey E. Sizikov, Svetlana A. Ivanova, Natalia M. Baulina, Olga O. Favorova, Georgy A. Nevinsky, and Valentina N. Buneva. 2022. "Multiplex Analysis of Serum Cytokine Profiles in Systemic Lupus Erythematosus and Multiple Sclerosis" International Journal of Molecular Sciences 23, no. 22: 13829. https://doi.org/10.3390/ijms232213829
APA StyleMelamud, M. M., Ermakov, E. A., Boiko, A. S., Kamaeva, D. A., Sizikov, A. E., Ivanova, S. A., Baulina, N. M., Favorova, O. O., Nevinsky, G. A., & Buneva, V. N. (2022). Multiplex Analysis of Serum Cytokine Profiles in Systemic Lupus Erythematosus and Multiple Sclerosis. International Journal of Molecular Sciences, 23(22), 13829. https://doi.org/10.3390/ijms232213829