

Ontogenetic Variation in Macrocyclic and Hemicyclic Poplar Rust Fungi

Abstract
1. Introduction
2. Results
2.1. STE3.3 and STE3.4 Structure and Function Prediction
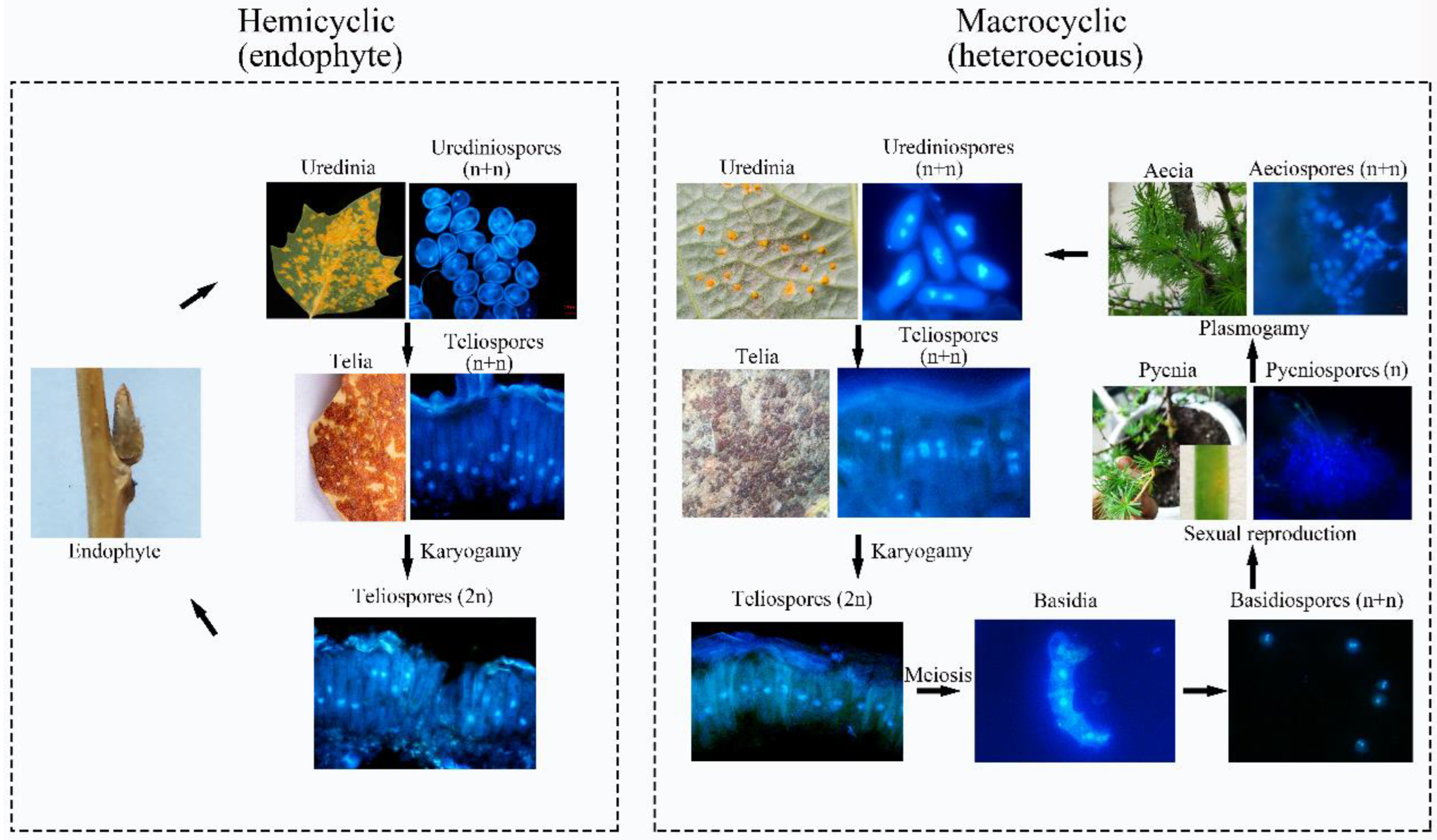
2.2. Divergence between Macrocyclic and Hemicyclic Rusts
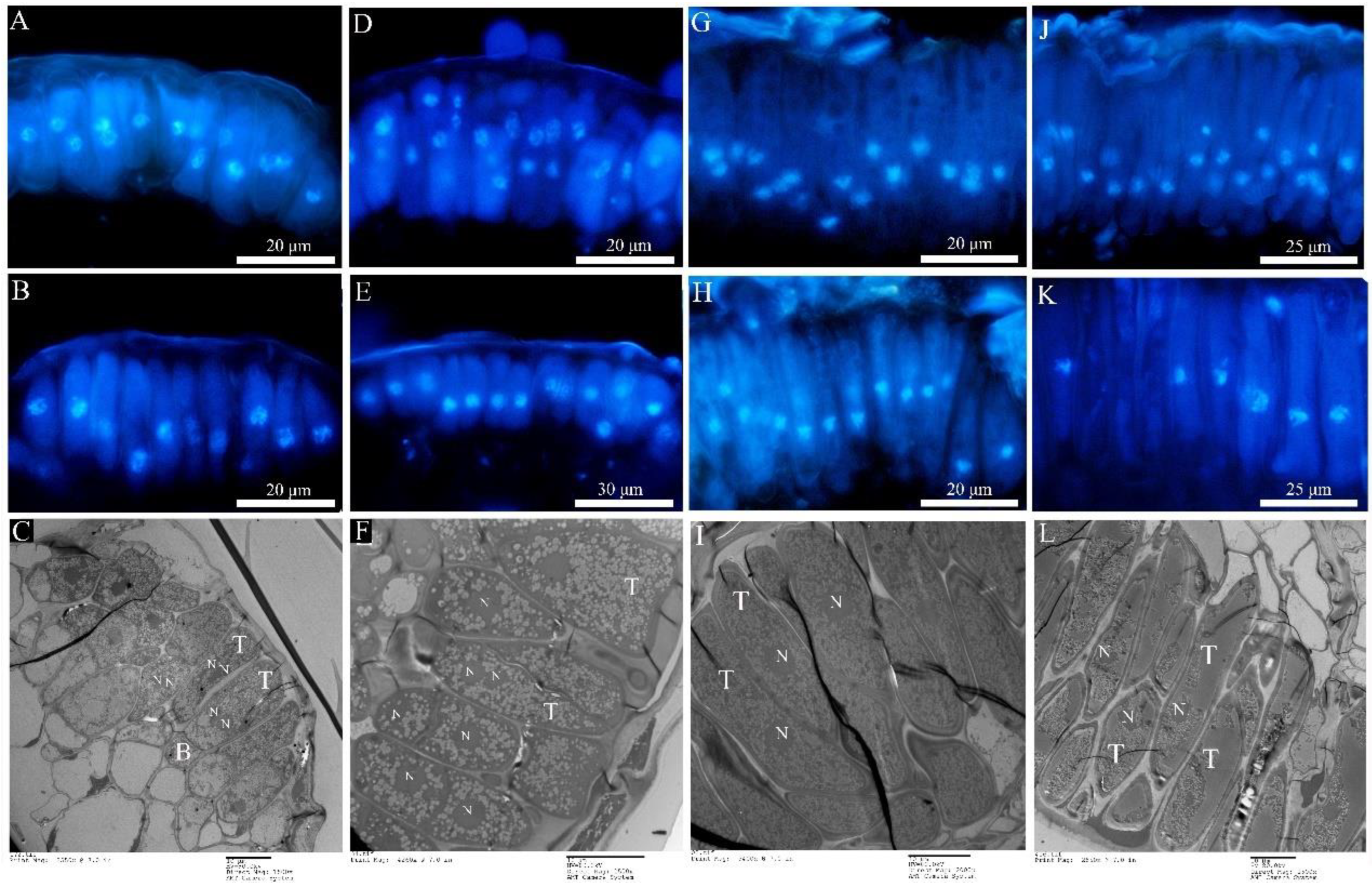
2.3. Ontogenetic Variation of Urediniospora Stage
2.4. Ontology in the Telia Stage
2.5. Ontology of Pycinospora and Aeciospora Stage
2.6. Detection of Rust in Winter Buds of Poplars
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. STE3.3 and STE3.4 Cloning
4.3. Predictions of STE3.3 and STE3.4 Structures and Functions
4.4. Molecular Clock Phylogenic Tree Construction
4.5. Observation of Ultrastructure at Uredia Stage
4.6. Ultrastructural Observation and qRT-PCR Analysis at the Telia Stage
4.7. Rust Detection and Verification by Artificial Inoculation of Poplar Winter Buds
4.8. Observation of the Ontogeny of the Basidiospores Stage
4.9. Ultrastructural Observation of Pycniospore Stage and Aeciospore Stage
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vialle, A.; Feau, N.; Frey, P.; Bernier, L.; Hamelin, R.C. Phylogenetic species recognition reveals host-specific lineages among poplar rust fungi. Mol. Phylogenet. Evol. 2013, 66, 628–644. [Google Scholar] [CrossRef] [PubMed]
- Vialle, A.; Frey, P.; Hambleton, S.; Bernier, L.; Hamelin, R.C. Poplar rust systematics and refinement of Melampsora species delineation. Fungal Divers. 2011, 50, 227–248. [Google Scholar] [CrossRef]
- Savile, D.B. Rusts that pass import inspection. Can. Plant Dis. Surv. 1973, 53, 105–106. [Google Scholar]
- Lorrain, C.; Gonçalves Dos Santos, K.C.; Germain, H.; Hecker, A.; Duplessis, S. Advances in understanding obligate biotrophy in rust fungi. New Phytol. 2019, 222, 1190–1206. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, H.A.; Kahmann, R.; Bölker, M. The pheromone response factor coordinates filamentous growth and pathogenicity in Ustilago maydis. EMBO J. 1996, 15, 1632–1642. [Google Scholar] [CrossRef] [PubMed]
- Kronstad, J.W.; Staben, C. Mating type in filamentous fungi. Annu. Rev. Genet. 1997, 31, 245–276. [Google Scholar] [CrossRef]
- Kües, U.; Casselton, L.A. Homeodomains and regulation of sexual development in basidiomycetes. Trends Genet. 1992, 8, 154–155. [Google Scholar] [CrossRef]
- Kües, U.; Casselton, L.A. The origin of multiple mating types in mushrooms. J. Cell Sci. 1993, 104, 227–230. [Google Scholar] [CrossRef]
- Lin, X.; Jackson, J.C.; Feretzaki, M.; Xue, C.; Heitman, J. Transcription factors Mat2 and Znf2 operate cellular circuits orchestrating opposite- and same-sex mating in Cryptococcus neoformans. PLoS Genet. 2010, 6, e1000953. [Google Scholar] [CrossRef]
- Wahl, R.; Zahiri, A.; Kämper, J. The Ustilago maydis b mating type locus controls hyphal proliferation and expression of secreted virulence factors in planta. Mol. Microbiol. 2010, 75, 208–220. [Google Scholar] [CrossRef]
- Xue, C.; Hsueh, Y.P.; Heitman, J. Magnificent seven: Roles of G protein-coupled receptors in extracellular fungi. FEMS Microbiol. Rev. 2008, 32, 1010–1032. [Google Scholar] [CrossRef] [PubMed]
- Devier, B.; Aguileta, G.; Hood, M.; Giraud, T. Ancient trans-specifific polymorphism at pheromone receptor genes in basidiomycetes. Genetics 2009, 181, 209–223. [Google Scholar] [CrossRef] [PubMed]
- Duplessis, S.; Cuomo, C.A.; Lin, Y.C.; Aerts, A.; Tisserant, E.; Veneault-Fourrey, C.; Joly, D.L.; Hacquard, S.; Amselem, J.; Cantarel, B.L.; et al. Obligate biotrophy features unraveled by the genomic analysis of rust fungi. Proc. Natl. Acad. Sci. USA 2011, 108, 9166–9171. [Google Scholar] [CrossRef] [PubMed]
- Stajich, J.S.; Wilke, S.K.; Ahrén, D.; Au, C.H.; Birren, B.W.; Borodovsky, M.; Burns, C.; Canbäck, B.; Casselton, L.A.; Cheng, C.K.; et al. Insights into evolution of multicellular fungi from the assembled chromosomes of the mushroom Coprinopsis cinerea (Coprinus cinereus). Proc. Natl. Acad. Sci. USA 2010, 107, 11889–11894. [Google Scholar] [CrossRef]
- Bakkeren, G.; Kronstad, J.W. Linkage of mating-type loci distinguishes bipolar from tetrapolar mating in basidiomycetous smut fungi. Proc. Natl. Acad. Sci. USA 1994, 91, 7085–7089. [Google Scholar] [CrossRef]
- Davidson, R.C.; Nichols, C.B.; Cox, G.M.; Perfect, J.R.; Heitman, J. A MAP kinase cascade composed of cell type specific and non-specific elements controls mating and differentiation of the fungal pathogen Cryptococcus neoformans. Mol. Microbiol. 2003, 49, 469–485. [Google Scholar] [CrossRef]
- Li, A.Z.; Xu, X.F.; Lin, F.X.; Cheng, S.M.; Lin, F.C. Cloning and identification of partial DNA fragment for the B mating-type factor in Lentinula edodes using degenerate PCR. World J. Microbiol. Biotechnol. 2007, 23, 411–415. [Google Scholar] [CrossRef]
- Hsueh, Y.P.; Xue, C.; Heitman, J. G protein signaling governing cell fate decisions involves opposing Ga subunits in Cryptococcus neoformans. Mol. Biol. Cell 2007, 18, 3237–3249. [Google Scholar] [CrossRef]
- Urban, M.; Kahmann, R.; Bölker, M. Identification of the pheromone response element in Ustilago maydis. Mol. Gen. Genet. 1996, 251, 31–37. [Google Scholar] [CrossRef]
- Lin, X.; Patel, S.; Litvintseva, A.P.; Floyd, A.; Mitchell, T.G.; Heitman, J. Diploids in the Cryptococcus neoformans serotype A population homozygous for a mating type originate via unisexual mating. PLoS Pathog. 2009, 5, e1000283. [Google Scholar] [CrossRef]
- Stanton, B.C.; Giles, S.S.; Kruzel, E.K.; Warren, C.L.; Ansari, A.Z.; Hull, C.M. Cognate site identififier analysis reveals novel binding properties of the sex inducer homeodomain proteins of Cryptococcus neoformans. Mol. Microbiol. 2009, 72, 1334–1347. [Google Scholar] [CrossRef] [PubMed]
- Brachmann, A.; Weinzierl, G.; Kämper, J.; Kahmann, R. Identification of genes of the bW/bE regulatory cascade in Ustilago maydis. Mol. Microbiol. 2001, 42, 1047–1063. [Google Scholar] [CrossRef] [PubMed]
- Zarnackz, K.; Eichhorn, H.; Kahmann, R.; Feldbrügge, M. Pheromone-regulated target genes respond differentially to MAPK phosphorylation of transcription factor Prf1. Mol. Microbiol. 2008, 69, 1041–1053. [Google Scholar] [CrossRef]
- Kües, U.; James, T.Y.; Heitman, J. Evolution of Fungi and Fungal-like Organisms; Pöggeler, S., Wöstemeyer, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; Volume 14, Chapter 6; pp. 97–160. [Google Scholar] [CrossRef]
- Lee, S.C.; Ni, M.; Li, W.; Shertz, C.; Heitman, J. The evolution of sex: A perspective from the fungal kingdom. Microbiol. Mol. Biol. Rev. 2010, 74, 298–340. [Google Scholar] [CrossRef] [PubMed]
- James, T.Y.; Lee, M.; van Diepen, L.T. A single mating type locus comprised of homeodomain genes promotes nuclear migration and heterokaryosis in the white-rot fungus Phanerochaete chrysosporium. Eukaryot. Cell 2011, 10, 249–261. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ono, Y. Life cycle and nuclear behavior in three rust fungi (Uredinales). Mycoscience 2002, 43, 37–45. [Google Scholar] [CrossRef]
- Casselton, L.A.; Olesnicky, N.S. Molecular genetics of mating type recognition in basidiomycete fungi. Microbiol. Mol. Biol. Rev. 1998, 62, 55–70. [Google Scholar] [CrossRef] [PubMed]
- Raudaskoski, M.; Kothe, E. Basidiomycete mating type genes and pheromone signaling. Eukaryot. Cell 2010, 9, 847–859. [Google Scholar] [CrossRef]
- Pöggeler, S.; Nowrousian, M.; Teichert, I.; Beier, A.; Kück, U. Fruiting Body Development in Ascomycetes. In Physiology and Genetics; Anke, T., Schüffler, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2018; Volume 15, Chapter 1; pp. 325–355. Available online: https://link.springer.com/chapter/10.1007/978-3-319-71740-1_1 (accessed on 2 July 2020).
- Savile, D.B. Coevolution of the rust fungi and their hosts. Q. Rev. Biol. 1971, 46, 211–218. [Google Scholar] [CrossRef]
- Bagyanarayana, G. Proceedings of the First IUFRO Rust of Forest Trees Working Part Conference; Jalkannen, R., Crane, P.E., Walla, J., Aalto, T., Eds.; Finnish Forest Research Institute: Saariselka, Finland, 1998; pp. 37–53. [Google Scholar]
- Tian, C.M.; Kakishima, M. Rust Diseases of Willow and Poplar; Pei, M.H., McCracken, A.R., Eds.; CABI Publishing: London, UK, 2005; Chapter 8; pp. 99–112. [Google Scholar]
- Tai, F.L. Sylloge Fungorum Sinicorum; Science Press: Beijing, China, 1979; pp. 537–539. (In Chinese) [Google Scholar]
- Peng, Z.J.; Xiong, C.W.; Luo, Z.Y.; Hu, X.Y.; Yu, Z.D.; Chen, T.X.; Xu, Y.; Wang, B. First report of alternate hosts of willow rust disease caused by Melampsora ferrinii in China. Plant Dis. 2022, 1, 324. [Google Scholar] [CrossRef]
- Spiers, A.G.; Hopcroft, D.H. Penetration and infection of poplar leaves by urediniospores of Melampsora larici-populina and Melampsora medusae. N. Z. J. Bot. 1988, 26, 101–111. [Google Scholar] [CrossRef]
- Harder, D.E.; Chong, J. Electron Microscopy of Plant Pathogens; Mendgen, K., Lesemann, D.E., Eds.; Springer: Berlin/Heidelberg, Germany, 1991; Chapter 18; pp. 235–250. [Google Scholar] [CrossRef]
- Luna, E.; Pastor, V.; Robert, J.; Flors, V.; Mauch-Mani, B.; Ton, J. Callose deposition: A multifaceted plant defense response. Mol. Plant-Microbe Interact. 2011, 24, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Spiers, A.G.; Hopcroft, D.H. Ultrastructural studies of the spermatial and aecial stages of Melampsora larici-populina and Melampsora epilea on Larix decidua. N. Z. J. Bot. 1985, 23, 101–116. [Google Scholar] [CrossRef]
- Tian, C.M.; Liang, Y.M.; Kang, Z.S.; Li, Z.Q.; Zhao, Y.X. Cytological studies on the host-pathogen relationship in the course of poplar leaf infection by Melampsora larici-populina. Sci. Silvae Sin. 2002, 38, 87–93. (In Chinese) [Google Scholar] [CrossRef]
- Littlefield, L.J.; Bracker, C.E. Ultrastructural specialization at the host-pathogen interface in rust-infected flax. Protoplasma 1972, 74, 271–305. [Google Scholar] [CrossRef]
- Spiers, A.; Hopcroft, D.H. Ultrastructural studies of pathogenesis and uredinial devlopment of Melampsora larici-populina and M. medusae on poplar and M. coleosporioides and M. epitea on willow. N. Z. J. Bot. 1985, 23, 117–133. [Google Scholar] [CrossRef]
- Ono, Y. The diversity of nuclear cycle in microcyclic rust fungi (Uredinales) and its ecological and evolutionary implications. Mycoscience 2002, 43, 421–439. [Google Scholar] [CrossRef]
- Hacquard, S.; Delaruelle, C.; Frey, P.; Tisserant, E.; Kohler, A.; Duplessis, S. Transcriptome analysis of poplar rust telia reveals overwintering adaptation and tightly coordinated karyogamy and meiosis processes. Front. Plant Sci. 2013, 4, 456. [Google Scholar] [CrossRef]
- Zheng, W.; Newcombe, G.; Hu, D.; Cao, Z.M.; Yu, Z.D.; Peng, Z.J. The first record of a north American poplar leaf rust fungus, Melampsora medusae, in China. Forests 2019, 10, 182. [Google Scholar] [CrossRef]
- Jiang, N.; Liang, Y.M.; Tian, C.M. Eight Melampsora species infected in poplars in China. J. Northwest For. Univ. 2021, 36, 149–157. (In Chinese) [Google Scholar] [CrossRef]
- Zhou, Y.; Dai, M.L.; Dai, X.G.; Li, S.X.; Yin, T.M.; Li, X.P. Isolation and Primary Identification of Pathogen of Leaf Rust on Black Cottonwood (Populus deltoides) in the South of China. Mol. Plant Breed. 2020, 18, 8240–8246. (In Chinese) [Google Scholar] [CrossRef]
- Kyte, J.; Doolittle, R.F. A simple method for displaying the hydropathic character of a problem. J. Mol. Biol. 1982, 157, 105–132. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Suleski, M.; Hedges, S.B. TimeTree: A Resource for Timelines, Timetrees, and Divergence Times. Mol. Biol. Evol. 2017, 34, 1812–1819. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z. PAML 4: Phylogenetic analysis by maximum likelihood. Mol. Biol. Evol. 2007, 24, 1586–1591. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.N.; Zhao, J.; Zhang, H.C.; Cheng, Y.L.; Kang, Z.S. Histological and cytological observations of pycnial and aecial formation of Puccinia recondite on Clematis intricate. Mycosystema 2015, 34, 1085–1091. [Google Scholar] [CrossRef]
- Ye, J.; Jin, C.F.; Li, N.; Liu, M.H.; Fei, Z.X.; Dong, L.Z.; Li, L.; Li, Z.Q. Selection of suitable reference genes for qRT-PCR normalisation under different experimental conditions in Eucommia ulmoides Oliv. Sci. Rep. 2018, 8, 15043. [Google Scholar] [CrossRef]
- Hacquard, S.; Petre, B.; Frey, P.; Hecker, A.; Rouhier, N.; Duplessis, S. The poplar-poplar rust interaction: Insights from genomics and transcriptomics. J. Pathog. 2011, 2011, 716041. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes--application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- Boyle, B.; Hamelin, R.C.; Séguin, A. In vivo monitoring of obligate biotrophic pathogen growth by kinetic PCR. Appl. Environ. Microbiol. 2005, 71, 1546–1552. [Google Scholar] [CrossRef]
- Pernaci, M.; De Mita, S.; Andrieux, A.; Pétrowski, J.; Halkett, F.; Duplessis, S.; Frey, P. Genome-wide patterns of segregation and linkage disequilibrium: The construction of a linkage genetic map of the poplar rust fungus Melampsora larici-populina. Front. Plant Sci. 2014, 5, 454. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Host Plants | Locality | Voucher Specimen No. | GenBank Accession | Species | |
|---|---|---|---|---|---|
| ITS | D1/D2 | ||||
| P. purdomii | Taibai Shaanxi | HMAS 247968 | MK028576 | MK064523 | M. larici-populina |
| P. tomentosa | Tianshui Gansu | HMAS 247981 | MK028582 | MK064530 | M. magnusiana |
| P. euphratica | Inner Mongolia | HMAS 247982 | MK028585 | MK064533 | M. pruinosae |
| P. deltoides cv. “Zhonghua hongye” | Linyou Shaanxi | HMAS 247973 | MK028586 | MK064528 | M. medusae |
| Gene | Primer Name | Primer Sequence (5′–3′) | Annealing Temperature |
|---|---|---|---|
| STE3.3 | 5′RACE Outer1 | GATTACGCCAAGCTTGTGTTCTGAGGTTCGTGTGCTGGCG | 65 °C |
| 5′RACE Inner1 | GATTACGCCAAGCTTAGCCAATACAAC ACATACGCGGAGTA | 65 °C | |
| 3′RACE Outer1 | GATTACGCCAAGCTTGCTCAA TCTCGT GGGTCCAGTGGGTCTC | 69 °C | |
| 3′RACE Inner1 | GATTACGCCAAGCTTTGTATCAACTTC AAGCCCGCGACTGG | 60 °C | |
| STE3.4 | 5′RACE Outer2 | GATTACGCCAAGCTTGCAGCTAGGACG CACATTCGGACA | 66 °C |
| 5′RACE Inner2 | GATTACGCCAAGCTTAGCGGTGCAAAT TCAAAGGATTGGTCGA | 62 °C | |
| 3′RACE Outer2 | GATTACGCCAAGCTTGTCCCTCAATGT GTACCAGGCTC | 66 °C | |
| 3′RACE Inner2 | GATTACGCCAAGCTTCAATGAACTTGG AA TCCGGGAGTG | 58 °C |
| Gene Symbol | Primer Sequence (5′–3′) Forward/Reverse | Amplicon Length (bp) | Annealing Temperature |
|---|---|---|---|
| Kar5 | F-GCTAATATCTCTAGCTTCATGCC R-CTTGTAGACCCGGAAACCT | 166 | 53 |
| Kar9 | F-CATTGTCCCGTTAGCTGGTT R-CTGCTGAAGGTCCACCAAGT | 221 | 55 |
| Mnd1 | F-AGAACTTGATAGATGATGGATTAG R-GCTTTATTAATGTTCACCGATTG | 100 | 50 |
| Spo11 | F-AGTGGTTTGTTACTTGGTATTACT R-CATGCTGAGCCGGTAAGT | 139 | 53 |
| ITS | F-TGAGCGACTTTAATGTGACTC R-ATGTAAATCAAAGTTGCCTTTGCG | 123 | 55 |
| Alpha-tubulin (aTUB) | F-ATCTGTAACGAACCTCCTGCTA R-CCTCCTCCATACCTTCTCCAA | 168 | 55 |
| Elongation factor-1-alpha (ELF1a) | F-CGAGACTCCCAAATACTTCGTT R-GTTCACGAGTTTGACCATCCTT | 167 | 55 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, Z.; Peng, Z.; Qi, M.; Zheng, W. Ontogenetic Variation in Macrocyclic and Hemicyclic Poplar Rust Fungi. Int. J. Mol. Sci. 2022, 23, 13062. https://doi.org/10.3390/ijms232113062
Yu Z, Peng Z, Qi M, Zheng W. Ontogenetic Variation in Macrocyclic and Hemicyclic Poplar Rust Fungi. International Journal of Molecular Sciences. 2022; 23(21):13062. https://doi.org/10.3390/ijms232113062
Chicago/Turabian StyleYu, Zhongdong, Zijia Peng, Mei Qi, and Wei Zheng. 2022. "Ontogenetic Variation in Macrocyclic and Hemicyclic Poplar Rust Fungi" International Journal of Molecular Sciences 23, no. 21: 13062. https://doi.org/10.3390/ijms232113062
APA StyleYu, Z., Peng, Z., Qi, M., & Zheng, W. (2022). Ontogenetic Variation in Macrocyclic and Hemicyclic Poplar Rust Fungi. International Journal of Molecular Sciences, 23(21), 13062. https://doi.org/10.3390/ijms232113062