
Genetic Ablation of Nrf2 Exacerbates Neuroinflammation in Ocular Autoimmunity

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
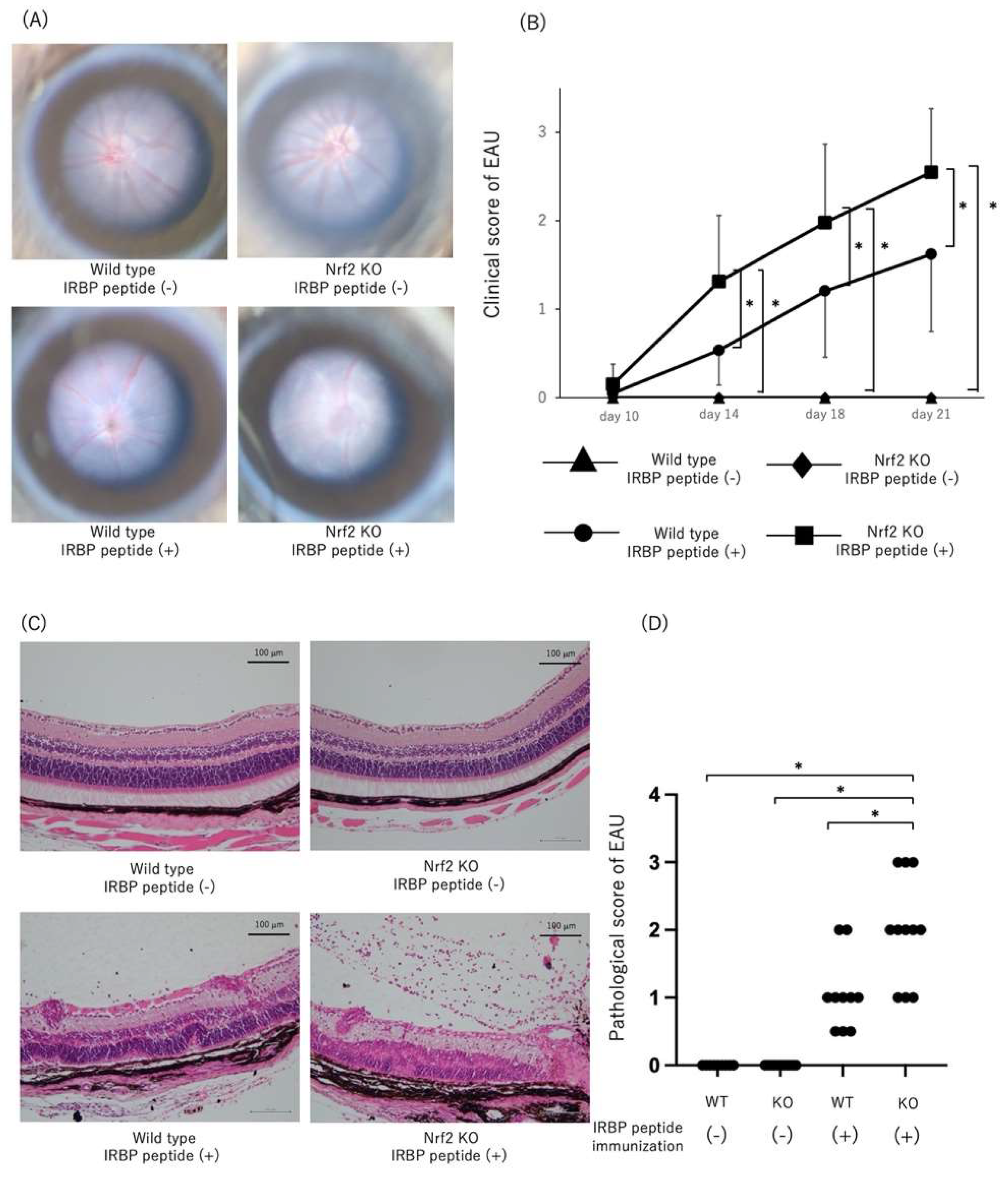
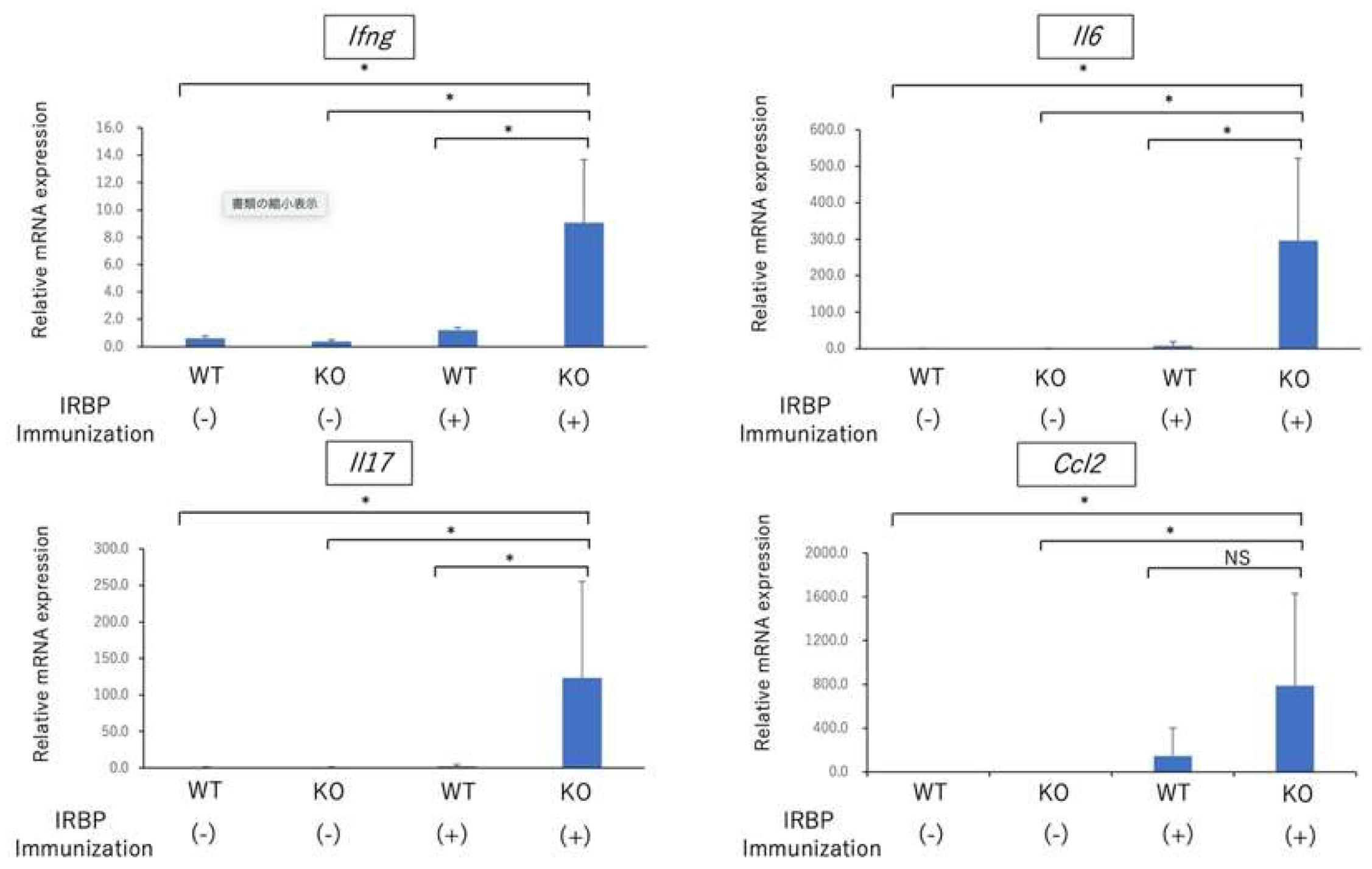
2.1. Exacerbation of EAU in Nrf2 Deficient Mice
2.2. GFAP and Iba-1 Expression in Retina of EAU-Induced Nrf2 Deficient Mice
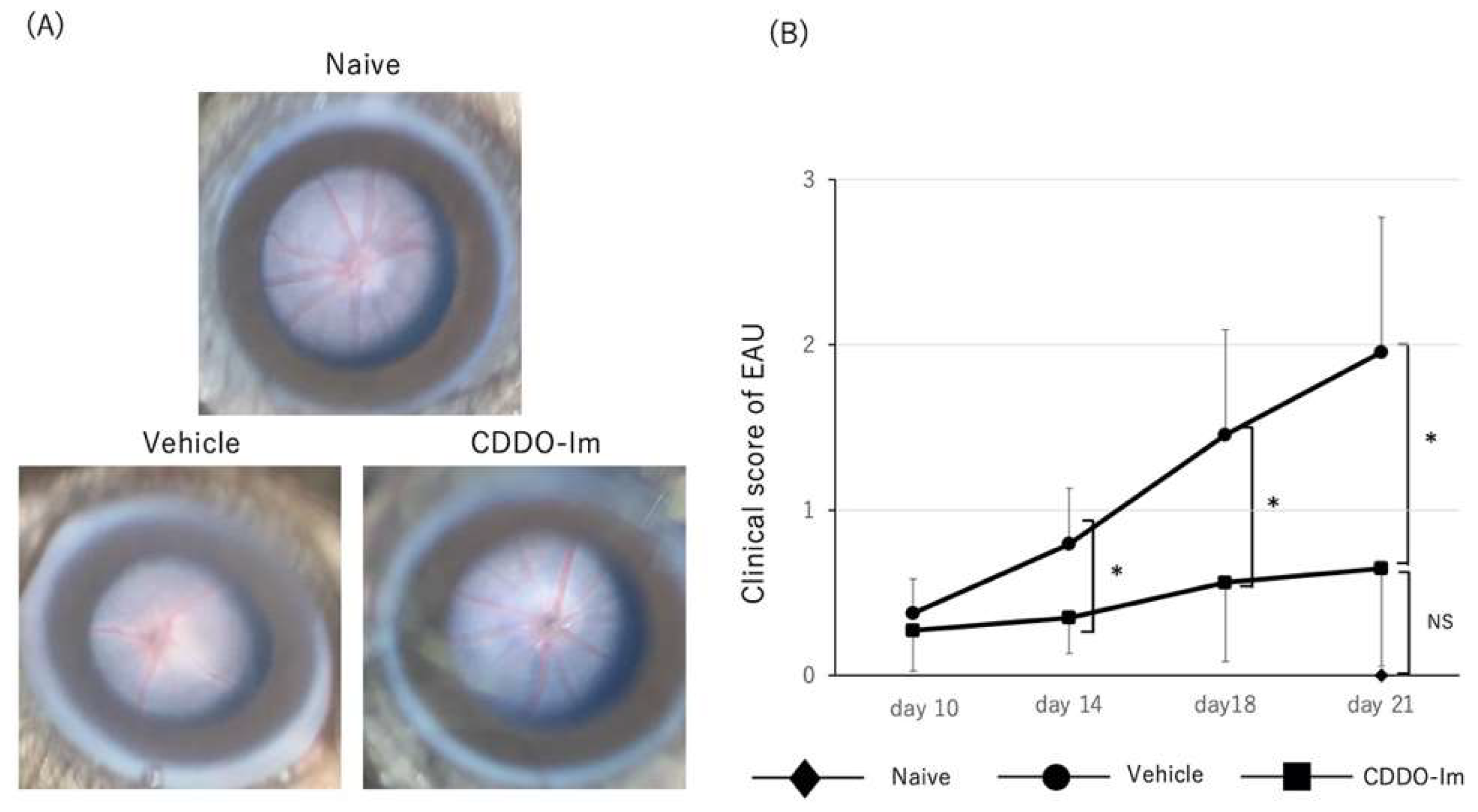
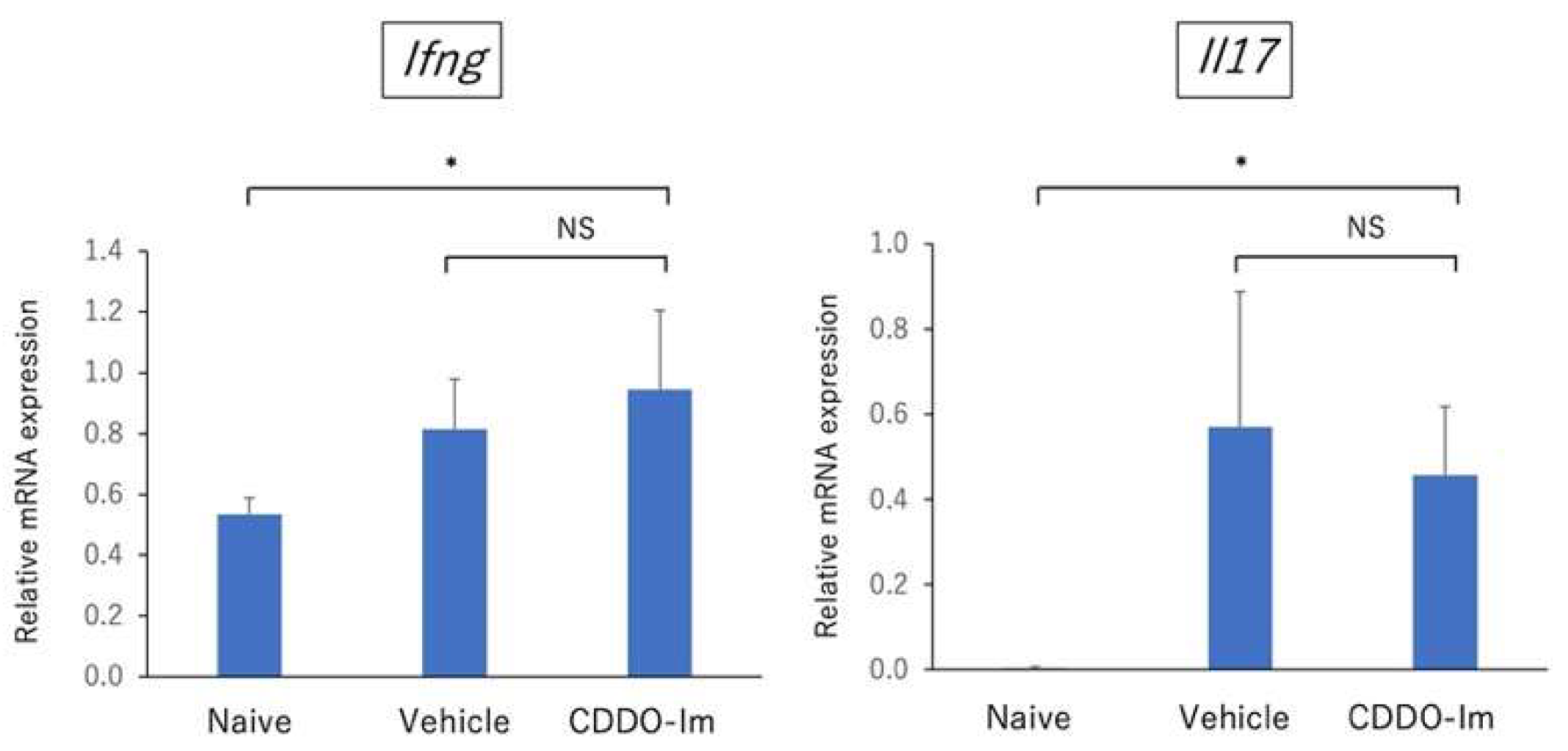
2.3. Amelioration of EAU by CDDO-Im Treatment
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Induction and Scoring of EAU
4.3. In Vitro Cytokine Assay Using EILSA
4.4. Reverse Transcription and Quantitative Polymerase Chain Reaction (PCR)
4.5. Immunohistochemistry
4.6. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rothova, A.; Schulten, M.S.S.-V.; Treffers, W.F.; Kijlstra, A. Causes and frequency of blindness in patients with intraocular inflammatory disease. Br. J. Ophthalmol. 1996, 80, 332–336. [Google Scholar] [CrossRef] [PubMed]
- Rosenbaum, J.T.; Bodaghi, B.; Couto, C.; Zierhut, M.; Acharya, N.; Pavesio, C.; Tay-Kearney, M.-L.; Neri, P.; Douglas, K.; Pathai, S.; et al. New observations and emerging ideas in diagnosis and management of non-infectious uveitis: A review. Semin. Arthritis Rheum. 2019, 49, 438–445. [Google Scholar] [CrossRef] [PubMed]
- Caspi, R.; Roberge, F.G.; Chan, C.C.; Wiggert, B.; Chader, G.J.; A Rozenszajn, L.; Lando, Z.; Nussenblatt, R.B. A new model of autoimmune disease. Experimental autoimmune uveoretinitis induced in mice with two different retinal antigens. J. Immunol. 1988, 140, 1490–1495. [Google Scholar]
- Gregerson, D.; Obritsch, W.F.; Fling, S.P.; Cameron, J.D. S-antigen-specific rat T cell lines recognize peptide fragments of S-antigen and mediate experimental autoimmune uveoretinitis and pinealitis. J. Immunol. 1986, 136, 2875–2882. [Google Scholar] [PubMed]
- Agarwal, R.K.; Silver, P.B.; Caspi, R.R. Rodent Models of Experimental Autoimmune Uveitis. Methods Mol. Biol. 2012, 900, 443–469. [Google Scholar] [CrossRef]
- Caspi, R.R. Th1 and Th2 Responses in Pathogenesis and Regulation of Experimental Autoimmune Uveoretinitis. Int. Rev. Immunol. 2002, 21, 197–208. [Google Scholar] [CrossRef] [PubMed]
- Luger, D.; Caspi, R.R. New perspectives on effector mechanisms in uveitis. Semin. Immunopathol. 2008, 30, 135–143. [Google Scholar] [CrossRef]
- Nussenblatt, R.B. Bench to Bedside: New Approaches to The Immunotherapy of Uveitic Disease. Int. Rev. Immunol. 2002, 21, 273–289. [Google Scholar] [CrossRef]
- Ung, L.; Pattamatta, U.; Carnt, N.; Wilkinson-Berka, J.L.; Liew, G.; White, A.J. Oxidative stress and reactive oxygen species: A review of their role in ocular disease. Clin. Sci. 2017, 131, 2865–2883. [Google Scholar] [CrossRef]
- Rao, N.; Wu, G.-S. Free radical mediated photoreceptor damage in uveitis. Prog. Retin. Eye Res. 1999, 19, 41–68. [Google Scholar] [CrossRef]
- Kalariya, N.M.; Ramana, K.V. Emerging Role of Antioxidants in the Protection of Uveitis Complications. Curr. Med. Chem. 2011, 18, 931–942. [Google Scholar] [CrossRef]
- Kaneko, Y.; Rao, N.A. Mitochondrial oxidative stress initiates visual loss in sympathetic ophthalmia. Jpn. J. Ophthalmol. 2012, 56, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Itoh, K.; Chiba, T.; Takahashi, S.; Ishii, T.; Igarashi, K.; Katoh, Y.; Oyake, T.; Hayashi, N.; Satoh, K.; Hatayama, I.; et al. An Nrf2/Small Maf Heterodimer Mediates the Induction of Phase II Detoxifying Enzyme Genes through Antioxidant Response Elements. Biochem. Biophys. Res. Commun. 1997, 236, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Cuadrado, A. Brain-Protective Mechanisms of Transcription Factor NRF2: Toward a Common Strategy for Neurodegenerative Diseases. Annu. Rev. Pharmacol. Toxicol. 2022, 62, 255–277. [Google Scholar] [CrossRef] [PubMed]
- Nagai, N.; Thimmulappa, R.K.; Cano, M.; Fujihara, M.; Izumi-Nagai, K.; Kong, X.; Sporn, M.B.; Kensler, T.W.; Biswal, S.; Handa, J.T. Nrf2 is a critical modulator of the innate immune response in a model of uveitis. Free Radic. Biol. Med. 2009, 47, 300–306. [Google Scholar] [CrossRef]
- Kobayashi, E.H.; Suzuki, T.; Funayama, R.; Nagashima, T.; Hayashi, M.; Sekine, H.; Tanaka, N.; Moriguchi, T.; Motohashi, H.; Nakayama, K.; et al. Nrf2 suppresses macrophage inflammatory response by blocking proinflammatory cytokine transcription. Nat. Commun. 2016, 7, 11624. [Google Scholar] [CrossRef]
- Liu, M.; Reddy, N.M.; Higbee, E.M.; Potteti, H.R.; Noel, S.; Racusen, L.; Kensler, T.W.; Sporn, M.B.; Reddy, S.P.; Rabb, H. The Nrf2 triterpenoid activator, CDDO-imidazolide, protects kidneys from ischemia–reperfusion injury in mice. Kidney Int. 2014, 85, 134–141. [Google Scholar] [CrossRef]
- Mehta, J.; Rayalam, S.; Wang, X. Cytoprotective Effects of Natural Compounds against Oxidative Stress. Antioxidants 2018, 7, 147. [Google Scholar] [CrossRef]
- Xu, D.; Chen, L.; Chen, X.; Wen, Y.; Yu, C.; Yao, J.; Wu, H.; Wang, X.; Xia, Q.; Kong, X. The triterpenoid CDDO-imidazolide ameliorates mouse liver ischemia-reperfusion injury through activating the Nrf2/HO-1 pathway enhanced autophagy. Cell Death Dis. 2017, 8, e2983. [Google Scholar] [CrossRef]
- Saha, S.; Buttari, B.; Panieri, E.; Profumo, E.; Saso, L. An Overview of Nrf2 Signaling Pathway and Its Role in Inflammation. Molecules 2020, 25, 5474. [Google Scholar] [CrossRef]
- Tang, Y.; Le, W. Differential Roles of M1 and M2 Microglia in Neurodegenerative Diseases. Mol. Neurobiol. 2016, 53, 1181–1194. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Zhong, R.; Li, S.; Fu, Z.; Cheng, C.; Cai, H.; Le, W. Acute Hypoxia Induced an Imbalanced M1/M2 Activation of Microglia through NF-κB Signaling in Alzheimer’s Disease Mice and Wild-Type Littermates. Front. Aging Neurosci. 2017, 9, 282. [Google Scholar] [CrossRef] [PubMed]
- I Rojo, A.; Pajares, M.; García-Yagüe, A.J.; Buendia, I.; Van Leuven, F.; Yamamoto, M.; López, M.G.; Cuadrado, A. Deficiency in the transcription factor NRF2 worsens inflammatory parameters in a mouse model with combined tauopathy and amyloidopathy. Redox Biol. 2018, 18, 173–180. [Google Scholar] [CrossRef]
- Ren, P.; Chen, J.; Li, B.; Zhang, M.; Yang, B.; Guo, X.; Chen, Z.; Cheng, H.; Wang, P.; Wang, S.; et al. Nrf2 Ablation Promotes Alzheimer’s Disease-Like Pathology in APP/PS1 Transgenic Mice: The Role of Neuroinflammation and Oxidative Stress. Oxidative Med. Cell. Longev. 2020, 2020, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Okunuki, Y.; Mukai, R.; Nakao, T.; Tabor, S.J.; Butovsky, O.; Dana, R.; Ksander, B.R.; Connor, K.M. Retinal microglia initiate neuroinflammation in ocular autoimmunity. Proc. Natl. Acad. Sci. USA 2019, 116, 9989–9998. [Google Scholar] [CrossRef]
- Zhu, G.; Wang, X.; Chen, L.; Lenahan, C.; Fu, Z.; Fang, Y.; Yu, W. Crosstalk Between the Oxidative Stress and Glia Cells After Stroke: From Mechanism to Therapies. Front. Immunol. 2022, 13, 852416. [Google Scholar] [CrossRef] [PubMed]
- Liby, K.; Hock, T.; Yore, M.M.; Suh, N.; Place, A.E.; Risingsong, R.; Williams, C.R.; Royce, D.B.; Honda, T.; Honda, Y.; et al. The Synthetic Triterpenoids, CDDO and CDDO-Imidazolide, Are Potent Inducers of Heme Oxygenase-1 and Nrf2/ARE Signaling. Cancer Res. 2005, 65, 4789–4798. [Google Scholar] [CrossRef]
- Jang, J.U.; Lee, S.H.; Choi, C.U.; Bahk, S.-C.; Chung, H.T.; Yang, Y.S. Effects of Heme Oxygenase-1 Inducer and Inhibitor on Experimental Autoimmune Uveoretinitis. Korean J. Ophthalmol. 2007, 21, 238–243. [Google Scholar] [CrossRef]
- Kirino, Y.; Takeno, M.; Watanabe, R.; Murakami, S.; Kobayashi, M.; Ideguchi, H.; Ihata, A.; Ohno, S.; Ueda, A.; Mizuki, N.; et al. Association of reduced heme oxygenase-1 with excessive Toll-like receptor 4 expression in peripheral blood mononuclear cells in Behçet’s disease. Arthritis Res. Ther. 2008, 10, R16. [Google Scholar] [CrossRef]
- Larabee, C.M.; Desai, S.; Agasing, A.; Georgescu, C.; Wren, J.D.; Axtell, R.C.; Plafker, S.M. Loss of Nrf2 exacerbates the visual deficits and optic neuritis elicited by experimental autoimmune encephalomyelitis. Mol. Vis. 2016, 22, 1503–1513. [Google Scholar]
- Avichezer, D.; Silver, P.B.; Chan, C.C.; Wiggert, B.; Caspi, R. Identification of a new epitope of human IRBP that induces autoimmune uveoretinitis in mice of the H-2b haplotype. Investig. Ophthalmol. Vis. Sci. 2000, 41, 127–131. [Google Scholar]
- Namba, K.; Kitaichi, N.; Nishida, T.; Taylor, A.W. Induction of regulatory T cells by the immunomodulating cytokines alpha-melanocyte-stimulating hormone and transforming growth factor-beta2. J. Leukoc. Biol. 2002, 72, 946–952. [Google Scholar] [CrossRef] [PubMed]
- Keleku-Lukwete, N.; Suzuki, M.; Otsuki, A.; Tsuchida, K.; Katayama, S.; Hayashi, M.; Naganuma, E.; Moriguchi, T.; Tanabe, O.; Engel, J.D.; et al. Amelioration of inflammation and tissue damage in sickle cell model mice by Nrf2 activation. Proc. Natl. Acad. Sci.USA 2015, 112, 12169–12174. [Google Scholar] [CrossRef] [PubMed]
- Tsai, W.-H. Moment-preserving thresolding: A new approach. Comput. Vis. Graph. Image Process. 1985, 29, 377–393. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sato, Y.; Saito, S.; Nakayama, M.; Sugita, S.; Kudo, A.; Keino, H. Genetic Ablation of Nrf2 Exacerbates Neuroinflammation in Ocular Autoimmunity. Int. J. Mol. Sci. 2022, 23, 11715. https://doi.org/10.3390/ijms231911715
Sato Y, Saito S, Nakayama M, Sugita S, Kudo A, Keino H. Genetic Ablation of Nrf2 Exacerbates Neuroinflammation in Ocular Autoimmunity. International Journal of Molecular Sciences. 2022; 23(19):11715. https://doi.org/10.3390/ijms231911715
Chicago/Turabian StyleSato, Yasuhiko, Shoko Saito, Makiko Nakayama, Sunao Sugita, Akihiko Kudo, and Hiroshi Keino. 2022. "Genetic Ablation of Nrf2 Exacerbates Neuroinflammation in Ocular Autoimmunity" International Journal of Molecular Sciences 23, no. 19: 11715. https://doi.org/10.3390/ijms231911715
APA StyleSato, Y., Saito, S., Nakayama, M., Sugita, S., Kudo, A., & Keino, H. (2022). Genetic Ablation of Nrf2 Exacerbates Neuroinflammation in Ocular Autoimmunity. International Journal of Molecular Sciences, 23(19), 11715. https://doi.org/10.3390/ijms231911715