
Valorization of Biomasses from Energy Crops for the Discovery of Novel Thermophilic Glycoside Hydrolases through Metagenomic Analysis

 ,
,  ,
,  ,
,  and
and
Abstract
:
1. Introduction
2. Results
2.1. Enrichments
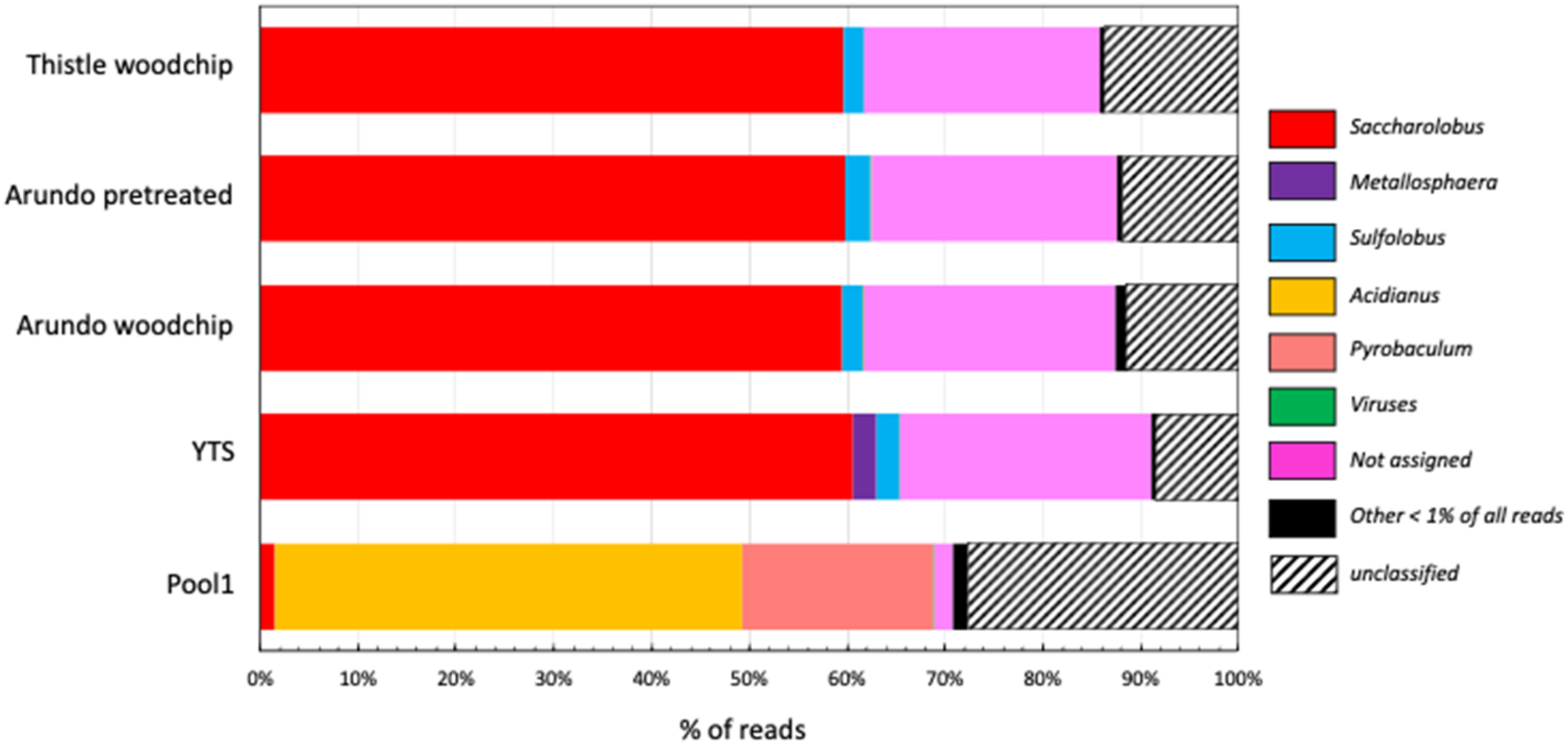
2.2. Sequence Analysis and Microbial Composition
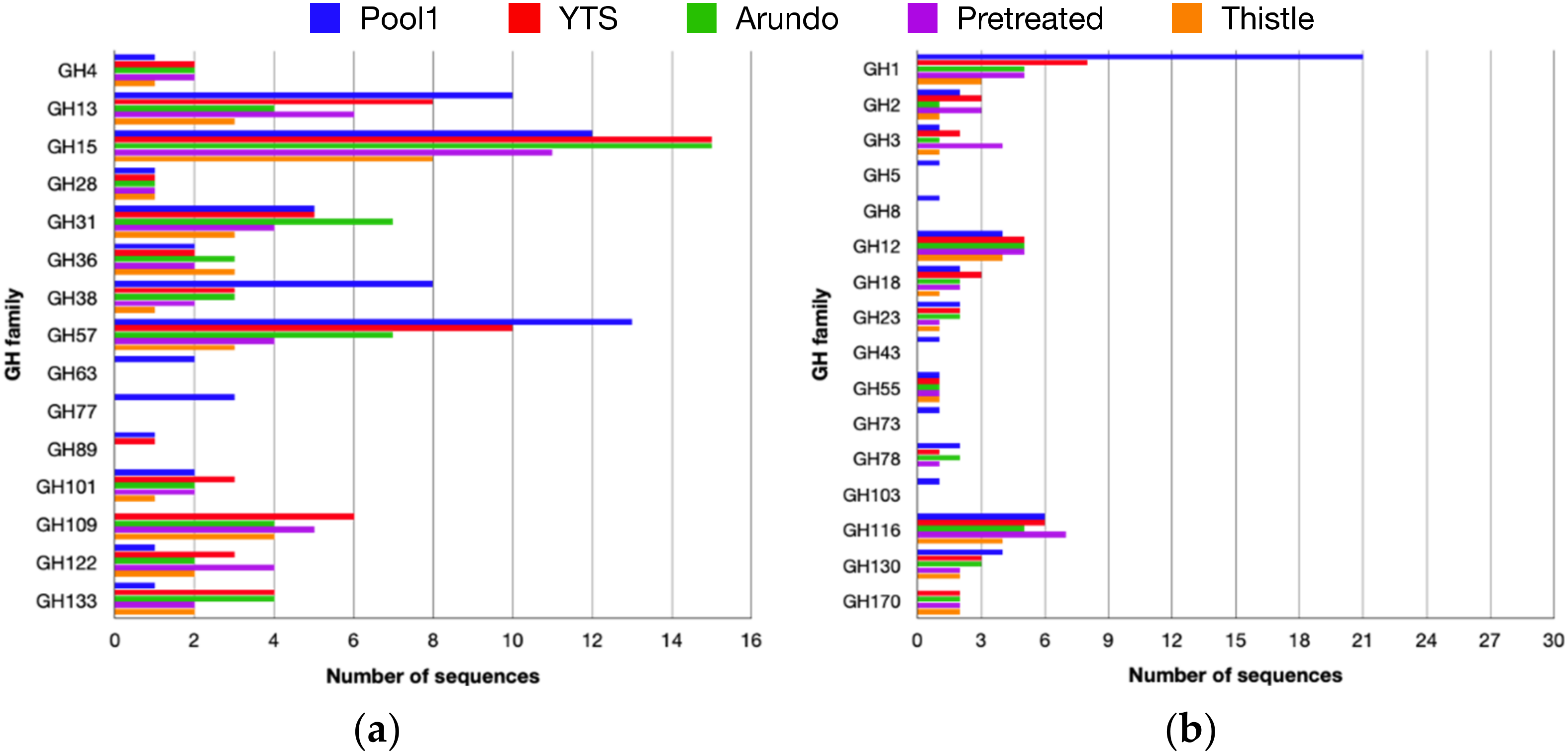
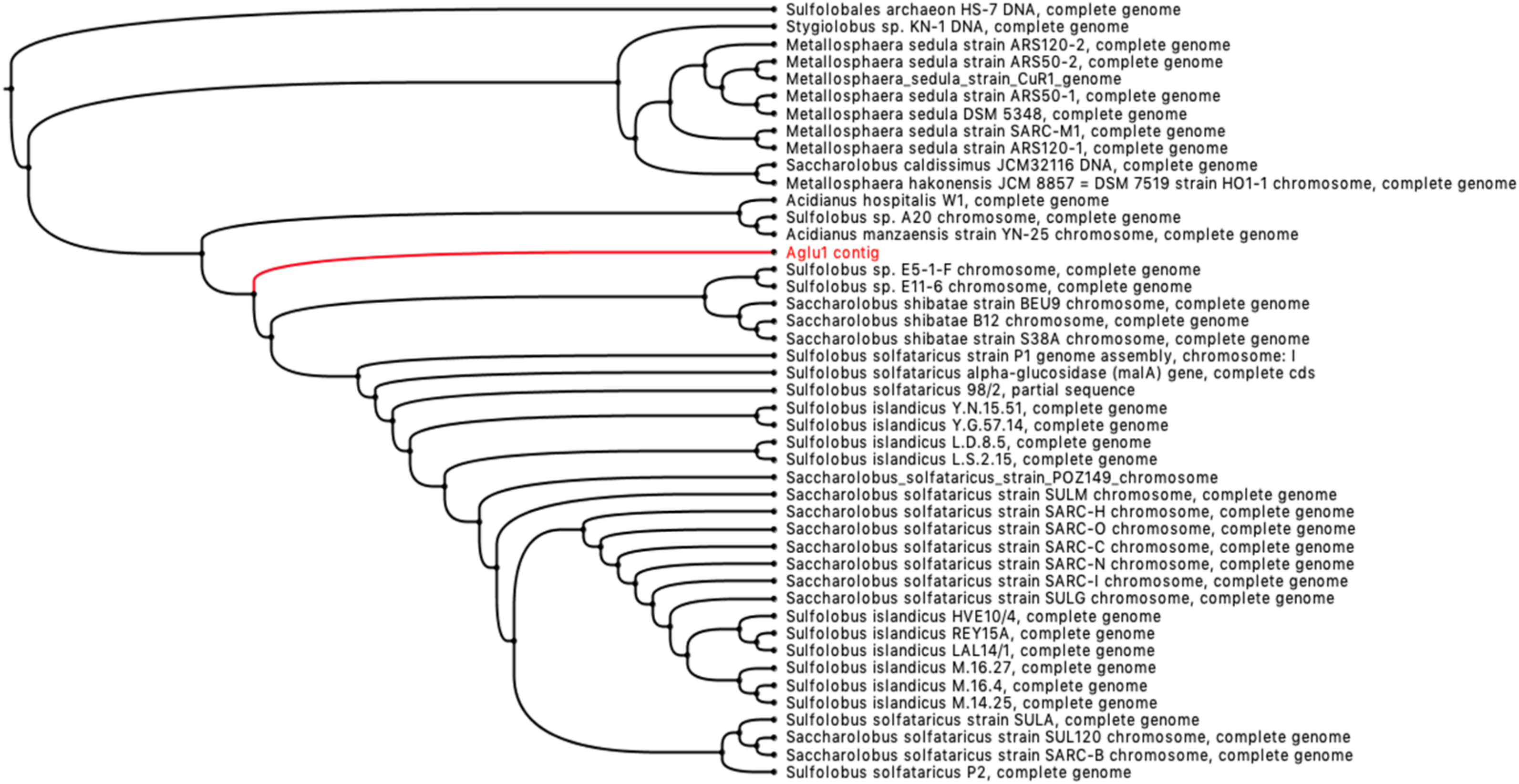
2.3. CAZome Analysis, Cloning, Expression, and Purification of Aglu1
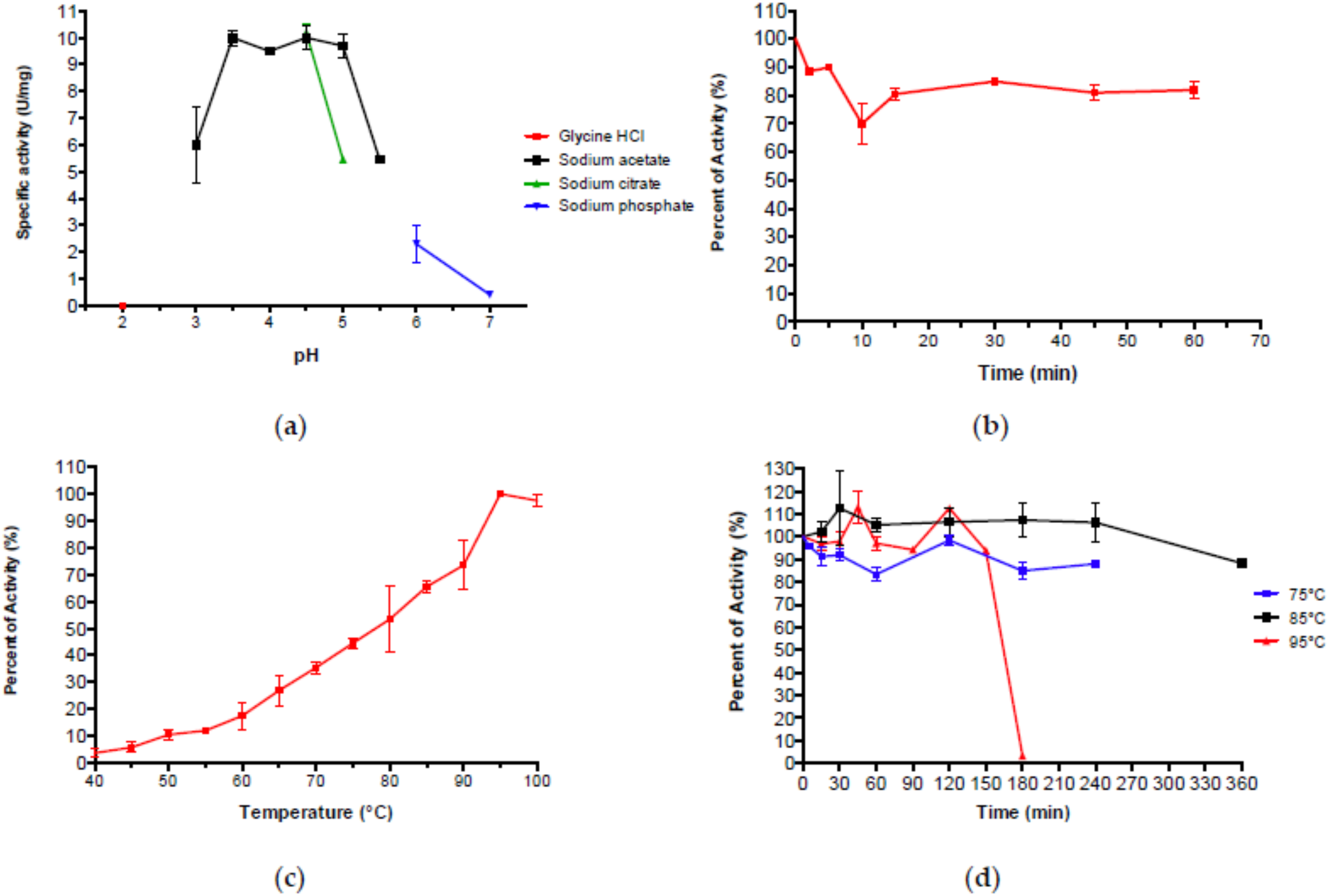
2.4. Molecular Mass Determination, pH and Temperature Dependence
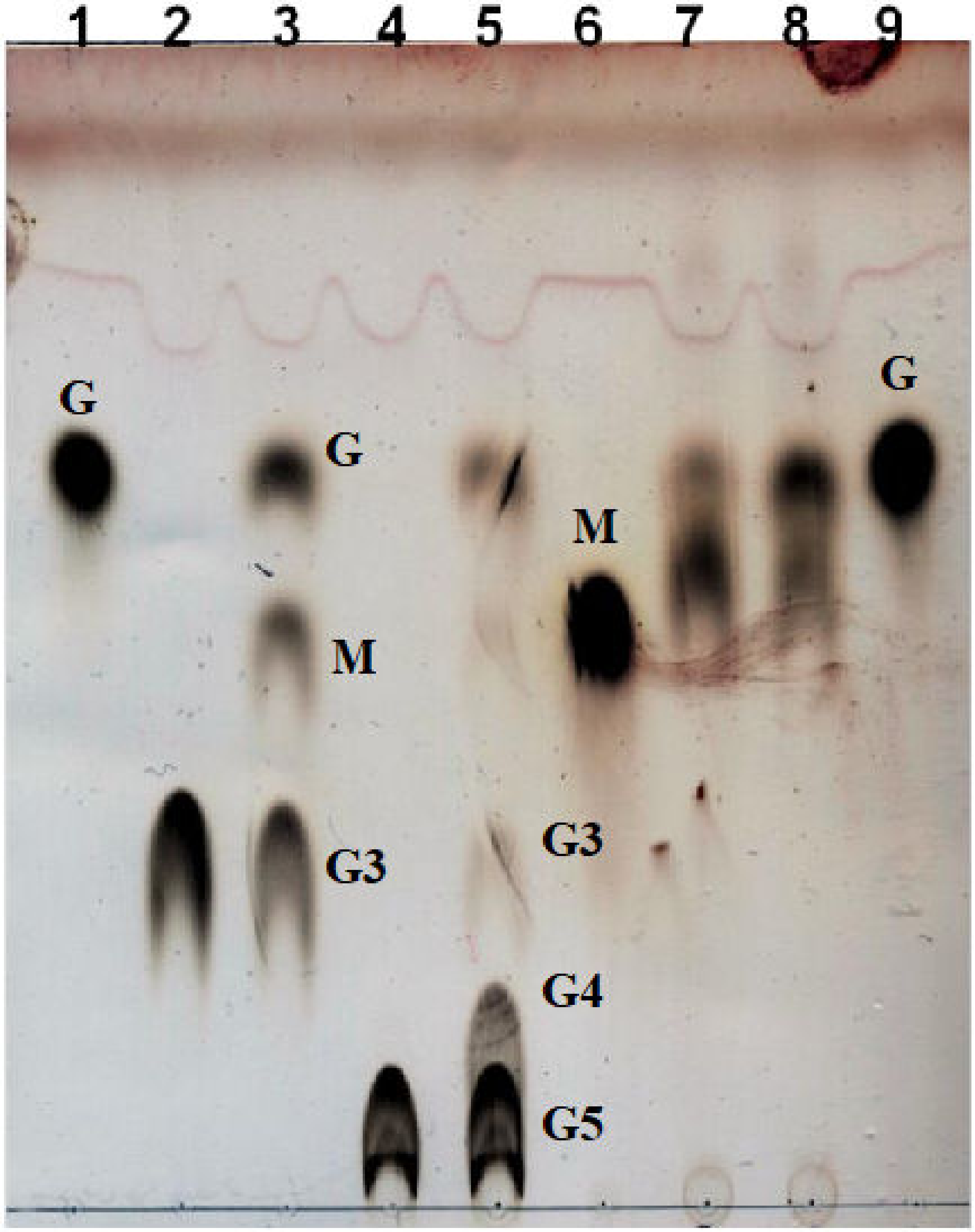
2.5. Substrate Specificity and Kinetics Parameters
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Environmental Sampling
4.3. Microbial Enrichment and Isolation of Metagenomic DNA
4.4. Taxonomic Analysis and Functional Annotation
4.5. Cloning, Expression, and Purification of Recombinant Aglu1
4.6. Molecular Mass Determination
4.7. Standard Assay
4.8. Temperature and pH Influence
4.9. Substrate Specificity and Steady-State Kinetic Constants
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cobucci-Ponzano, B.; Strazzulli, A.; Iacono, R.; Masturzo, G.; Giglio, R.; Rossi, M.; Moracci, M. Novel thermophilic hemicellulases for the conversion of lignocellulose for second generation biorefineries. Enzym. Microb. Technol. 2015, 78, 63–73. [Google Scholar] [CrossRef]
- Curci, N.; Strazzulli, A.; De Lise, F.; Iacono, R.; Maurelli, L.; Dal Piaz, F.; Cobucci-Ponzano, B.; Moracci, M. Identification of a novel esterase from the thermophilic bacterium Geobacillus thermodenitrificans NG80-2. Extremophiles 2019, 23, 407–419. [Google Scholar] [CrossRef]
- Dumorne, K.; Cordova, D.C.; Astorga-Elo, M.; Renganathan, P. Extremozymes: A Potential Source for Industrial Applications. J. Microbiol. Biotechnol. 2017, 27, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Adebisi, W.A.; Ahmad, F.; Sethupathy, S.; Danso, B.; Sun, J. Recent Development of Extremophilic Bacteria and Their Application in Biorefinery. Front. Bioeng. Biotechnol. 2020, 8, 483. [Google Scholar] [CrossRef] [PubMed]
- Saini, J.K.; Saini, R.; Tewari, L. Lignocellulosic agriculture wastes as biomass feedstocks for second-generation bioethanol production: Concepts and recent developments. 3 Biotech 2015, 5, 337–353. [Google Scholar] [CrossRef] [PubMed]
- Sadh, P.K.; Duhan, S.; Duhan, J.S. Agro-industrial wastes and their utilization using solid state fermentation: A review. Bioresour. Bioprocess. 2018, 5, 1–15. [Google Scholar] [CrossRef]
- Sarsaiya, S.; Jain, A.; Kumar Awasthi, S.; Duan, Y.; Kumar Awasthi, M.; Shi, J. Microbial dynamics for lignocellulosic waste bioconversion and its importance with modern circular economy, challenges and future perspectives. Bioresour. Technol. 2019, 291, 121905. [Google Scholar] [CrossRef]
- Di Donato, P.; Fiorentino, G.; Anzelmo, G.; Tommonaro, G.; Nicolaus, B.; Poli, A. Re-Use of Vegetable Wastes as Cheap Substrates for Extremophile Biomass Production. Waste Biomass Valorization 2011, 2, 103–111. [Google Scholar] [CrossRef]
- Robak, K.; Balcerek, M. Current state-of-the-art in ethanol production from lignocellulosic feedstocks. Microbiol. Res. 2020, 240, 126534. [Google Scholar] [CrossRef]
- De Medeiros, T.D.M.; Dufossé, L.; Bicas, J.L. Lignocellulosic substrates as starting materials for the production of bioactive biopigments. Food Chem. 2022, 13, 100223. [Google Scholar] [CrossRef]
- Grewal, J.; Khare, S.K. One-pot bioprocess for lactic acid production from lignocellulosic agro-wastes by using ionic liquid stable Lactobacillus brevis. Bioresour. Technol. 2018, 251, 268–273. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Quero, A.; Pollet, E.; Avérous, L.; Phalip, V. Optimized Bioproduction of Itaconic and Fumaric Acids Based on Solid-State Fermentation of Lignocellulosic Biomass. Molecules 2020, 25, 1070. [Google Scholar] [CrossRef] [PubMed]
- Namnuch, N.T.A.; Thammasittirong, S.N.R. Lignocellulose hydrolytic enzymes production by Aspergillus flavus KUB2 using submerged fermentation of sugarcane bagasse waste. Mycology 2021, 12, 119–127. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira Rodrigues, P.G.L.; Pasquini, D.; Badotti, F.; Góes-Neto, A.; Baffi, M.A. Lignocellulose-degrading enzymes production by solid-state fermentation through fungal consortium among Ascomycetes and Basidiomycetes. Renew. Energy 2020, 145, 2683–2693. [Google Scholar] [CrossRef]
- Kawaguchi, H.; Ogino, C.; Kondo, A. Microbial conversion of biomass into bio-based polymers. Bioresour. Technol. 2017, 245, 1664–1673. [Google Scholar] [CrossRef]
- Patel, S.K.S.; Gupta, R.K.; Das, D.; Lee, J.-K.; Kalia, V.C. Continuous biohydrogen production from poplar biomass hydrolysate by a defined bacterial mixture immobilized on lignocellulosic materials under non-sterile conditions. J. Clean. Prod. 2021, 287, 125037. [Google Scholar] [CrossRef]
- Fernando Sandun, A.S. Chandrapal Chauda, and Murali Naveen, Biorefineries: Current Status, Challenges, and Future Direction. Energy Fuels 2006, 20, 1727–1737. [Google Scholar] [CrossRef]
- Kamm, B.; Kamm, M. Principles of biorefineries. Appl. Microbiol. Biotechnol. 2004, 64, 137–145. [Google Scholar] [CrossRef]
- Ilari, A.; Duca, D.; Boakye-Yiadom, K.A.; Gasperini, T.; Toscano, G. Carbon Footprint and Feedstock Quality of a Real Biomass Power Plant Fed with Forestry and Agricultural Residues. Resources 2022, 11, 7. [Google Scholar] [CrossRef]
- Shatalov, A.A.; Pereira, H. Dissolving grade eco-clean cellulose pulps by integrated fractionation of cardoon (Cynara cardunculus L.) stalk biomass. Chem. Eng. Res. Des. 2014, 92, 2640–2648. [Google Scholar] [CrossRef]
- Branca, F.; Argento, S.; Paoletti, A.M.; Melilli, M.G. The Physiological Role of Inulin in Wild Cardoon (Cynara cardunculus L. var. sylvestris Lam.). Agronomy 2022, 12, 290. [Google Scholar] [CrossRef]
- Lemons e Silva, C.F.; Schirmer, M.A.; Maeda, R.N.; Barcelos, C.A.; Pereira, N. Potential of giant reed (Arundo donax L.) for second generation ethanol production. Electron. J. Biotechnol. 2015, 18, 10–15. [Google Scholar] [CrossRef]
- Proietti, S.; Moscatello, S.; Fagnano, M.; Fiorentino, N.; Impagliazzo, A.; Battistelli, A. Chemical composition and yield of rhizome biomass of Arundo donax L. grown for biorefinery in the Mediterranean environment. Biomass Bioenergy 2017, 107, 191–197. [Google Scholar] [CrossRef]
- Juerges, N.; Hansjürgens, B. Soil governance in the transition towards a sustainable bioeconomy—A review. J. Clean. Prod. 2018, 170, 1628–1639. [Google Scholar] [CrossRef]
- Strazzulli, A.; Iacono, R.; Giglio, R.; Moracci, M.; Cobucci-Ponzano, B. Hyperthermophilic Environments: Biodiversity and Biotechnology. In Microbial Ecology of Extreme Environments; Chénard, C.L.F., Lauro, F.M., Eds.; Springer: Cham, Switzerland, 2017. [Google Scholar]
- Iacono, R.; Cobucci-Ponzano, B.; De Lise, F.; Curci, N.; Maurelli, L.; Moracci, M.; Strazzulli, A. Spatial Metagenomics of Three Geothermal Sites in Pisciarelli Hot Spring Focusing on the Biochemical Resources of the Microbial Consortia. Molecules 2020, 25, 4023. [Google Scholar] [CrossRef]
- Strazzulli, A.; Cobucci-Ponzano, B.; Iacono, R.; Giglio, R.; Maurelli, L.; Curci, N.; Schiano-di-Cola, C.; Santangelo, A.; Contursi, P.; Lombard, V.; et al. Discovery of hyperstable carbohydrate-active enzymes through metagenomics of extreme environments. FEBS J. 2020, 287, 1116–1137. [Google Scholar] [CrossRef]
- Drula, E.; Garron, M.L.; Dogan, S.; Lombard, V.; Henrissat, B.; Terrapon, N. The carbohydrate-active enzyme database: Functions and literature. Nucleic Acids Res. 2022, 50, D571–D577. [Google Scholar] [CrossRef]
- Shiers, D.W.; Blight, K.R.; Ralph, D.E. Sodium sulphate and sodium chloride effects on batch culture of iron oxidising bacteria. Hydrometallurgy 2005, 80, 75–82. [Google Scholar] [CrossRef]
- Brock, T.D.; Brock, K.M.; Belly, R.T.; Weiss, R.L. Sulfolobus: A new genus of sulfur-oxidizing bacteria living at low pH and high temperature. Arch. Mikrobiol. 1972, 84, 54–68. [Google Scholar] [CrossRef]
- Amo, T.; Paje, M.L.; Inagaki, A.; Ezaki, S.; Atomi, H.; Imanaka, T. Pyrobaculum calidifontis sp. nov., a novel hyperthermophilic archaeon that grows in atmospheric air. Archaea 2002, 1, 113–121. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Yohe, T.; Huang, L.; Entwistle, S.; Wu, P.; Yang, Z.; Busk, P.K.; Xu, Y.; Yin, Y. dbCAN2: A meta server for automated carbohydrate-active enzyme annotation. Nucleic Acids Res. 2018, 46, W95–W101. [Google Scholar] [CrossRef] [PubMed]
- Rolfsmeier, M.; Blum, P. Purification and characterization of a maltase from the extremely thermophilic crenarchaeote Sulfolobus solfataricus. J. Bacteriol. 1995, 177, 482–485. [Google Scholar] [CrossRef] [PubMed]
- Moracci, M.; Cobucci Ponzano, B.; Trincone, A.; Fusco, S.; De Rosa, M.; van Der Oost, J.; Sensen, C.W.; Charlebois, R.L.; Rossi, M. Identification and molecular characterization of the first alpha-xylosidase from an archaeon. J. Biol. Chem. 2000, 275, 22082–22089. [Google Scholar] [CrossRef] [PubMed]
- Park, J.E.; Park, S.H.; Woo, J.Y.; Hwang, H.S.; Cha, J.; Lee, H. Enzymatic properties of a thermostable alpha-glucosidase from acidothermophilic crenarchaeon Sulfolobus tokodaii strain 7. J. Microbiol. Biotechnol. 2013, 23, 56–63. [Google Scholar] [CrossRef]
- Choi, K.H.; Hwang, S.; Cha, J. Identification and characterization of MalA in the maltose/maltodextrin operon of Sulfolobus acidocaldarius DSM639. J. Bacteriol. 2013, 195, 1789–1799. [Google Scholar] [CrossRef]
- Angelov, A.; Putyrski, M.; Liebl, W. Molecular and biochemical characterization of alpha-glucosidase and alpha-mannosidase and their clustered genes from the thermoacidophilic archaeon Picrophilus torridus. J. Bacteriol. 2006, 188, 7123–7131. [Google Scholar] [CrossRef]
- Seo, S.-H.; Choi, K.-H.; Hwang, S.; Kim, J.; Park, C.-S.; Rho, J.-R.; Cha, J. Characterization of the catalytic and kinetic properties of a thermostable Thermoplasma acidophilum α-glucosidase and its transglucosylation reaction with arbutin. J. Mol. Catal. B Enzym. 2011, 72, 305–312. [Google Scholar] [CrossRef]
- Ernst, H.A.; Lo Leggio, L.; Willemoes, M.; Leonard, G.; Blum, P.; Larsen, S. Structure of the Sulfolobus solfataricus alpha-glucosidase: Implications for domain conservation and substrate recognition in GH31. J. Mol. Biol. 2006, 358, 1106–1124. [Google Scholar] [CrossRef]
- Mirdita, M.; Schutze, K.; Moriwaki, Y.; Heo, L.; Ovchinnikov, S.; Steinegger, M. ColabFold: Making protein folding accessible to all. Nat. Methods 2022, 19, 679–682. [Google Scholar] [CrossRef]
- Nakai Hiroyuki, I.T.; Shigeki, T.; Kazuki, M.; Takeshi, Y.; Masayuki, O.; Haruhide, M.; Seiya, C.; Yoshio, S.; Atsuo, K. Plant α-Glucosidase: Molecular Analysis of Rice α-Glucosidase and Degradation Mechanism of Starch Granules in Germination Stage. J. Appl. Glycosci. 2006, 53, 137–142. [Google Scholar] [CrossRef]
- Cobucci-Ponzano, B.; Aurilia, V.; Riccio, G.; Henrissat, B.; Coutinho, P.M.; Strazzulli, A.; Padula, A.; Corsaro, M.M.; Pieretti, G.; Pocsfalvi, G.; et al. A new archaeal beta-glycosidase from Sulfolobus solfataricus: Seeding a novel retaining beta-glycan-specific glycoside hydrolase family along with the human non-lysosomal glucosylceramidase GBA2. J. Biol. Chem. 2010, 285, 20691–20703. [Google Scholar] [CrossRef]
- Entcheva, P.; Liebl, W.; Johann, A.; Hartsch, T.; Streit, W.R. Direct cloning from enrichment cultures, a reliable strategy for isolation of complete operons and genes from microbial consortia. Appl. Environ. Microbiol. 2001, 67, 89–99. [Google Scholar] [CrossRef]
- Giaveno, M.A.; Urbieta, M.S.; Ulloa, J.R.; Toril, E.G.; Donati, E.R. Physiologic versatility and growth flexibility as the main characteristics of a novel thermoacidophilic Acidianus strain isolated from Copahue geothermal area in Argentina. Microb. Ecol. 2013, 65, 336–346. [Google Scholar] [CrossRef]
- Huber, G.; Spinnler, C.; Gambacorta, A.; Stetter, K.O. Metallosphaera sedula gen, and sp. nov. Represents a New Genus of Aerobic, Metal-Mobilizing, Thermoacidophilic Archaebacteria. Syst. Appl. Microbiol. 1989, 12, 38–47. [Google Scholar] [CrossRef]
- Volkl, P.; Huber, R.; Drobner, E.; Rachel, R.; Burggraf, S.; Trincone, A.; Stetter, K.O. Pyrobaculum aerophilum sp. nov., a novel nitrate-reducing hyperthermophilic archaeum. Appl. Environ. Microbiol. 1993, 59, 2918–2926. [Google Scholar] [CrossRef]
- Zillig, W.; Stetter, K.O.; Wunderl, S.; Schulz, W.; Priess, H.; Scholz, I. The Sulfolobus-“Caldariella” group: Taxonomy on the basis of the structure of DNA-dependent RNA polymerases. Arch. Microbiol. 1980, 125, 259–269. [Google Scholar] [CrossRef]
- You, X.Y.; Liu, C.; Wang, S.Y.; Jiang, C.Y.; Shah, S.A.; Prangishvili, D.; She, Q.; Liu, S.J.; Garrett, R.A. Genomic analysis of Acidianus hospitalis W1 a host for studying crenarchaeal virus and plasmid life cycles. Extremophiles 2011, 15, 487–497. [Google Scholar] [CrossRef]
- Sakai, H.D.; Kurosawa, N. Saccharolobus caldissimus gen. nov., sp. nov., a facultatively anaerobic iron-reducing hyperthermophilic archaeon isolated from an acidic terrestrial hot spring, and reclassification of Sulfolobus solfataricus as Saccharolobus solfataricus comb. nov. and Sulfolobus shibatae as Saccharolobus shibatae comb. nov. Int. J. Syst. Evol. Microbiol. 2018, 68, 1271–1278. [Google Scholar]
- Rachel, R.; Bettstetter, M.; Hedlund, B.P.; Haring, M.; Kessler, A.; Stetter, K.O.; Prangishvili, D. Remarkable morphological diversity of viruses and virus-like particles in hot terrestrial environments. Arch. Virol. 2002, 147, 2419–2429. [Google Scholar] [CrossRef]
- Addou, S.; Rentzsch, R.; Lee, D.; Orengo, C.A. Domain-based and family-specific sequence identity thresholds increase the levels of reliable protein function transfer. J. Mol. Biol. 2009, 387, 416–430. [Google Scholar] [CrossRef]
- Rost, B.; Liu, J.; Nair, R.; Wrzeszczynski, K.O.; Ofran, Y. Automatic prediction of protein function. Cell. Mol. Life Sci. 2003, 60, 2637–2650. [Google Scholar]
- Tian, W.; Skolnick, J. How well is enzyme function conserved as a function of pairwise sequence identity? J. Mol. Biol. 2003, 333, 863–882. [Google Scholar] [CrossRef] [Green Version]
- Todd, A.E.; Orengo, C.A.; Thornton, J.M. Evolution of function in protein superfamilies, from a structural perspective. J. Mol. Biol. 2001, 307, 1113–1143. [Google Scholar] [CrossRef]
- Dusan, V.; Nenad, M.; Dejan, B.; Filip, B.; Segal, A.M.; Dejan, S.; Jovana, T.; Aleksandra, D. The specificity of alpha-glucosidase from Saccharomyces cerevisiae differs depending on the type of reaction: Hydrolysis versus transglucosylation. Appl. Microbiol. Biotechnol. 2014, 98, 6317–6328. [Google Scholar] [CrossRef]
- Yamamoto, K.; Davis, B.G. Creation of an alpha-mannosynthase from a broad glycosidase scaffold. Angew. Chem. Int. Ed. 2012, 51, 7449–7453. [Google Scholar] [CrossRef]
- Tshililo, N.O.; Strazzulli, A.; Cobucci-Ponzano, B.; Maurelli, L.; Iacono, R.; Bedini, E.; Corsaro, M.M.; Strauss, E.; Moracci, M. The α-Thioglycoligase Derived from a GH89 α-N-Acetylglucosaminidase Synthesises α-N-Acetylglucosamine-Based Glycosides of Biomedical Interest. Adv. Synth. Catal. 2017, 359, 663–676. [Google Scholar] [CrossRef]
- Cobucci-Ponzano, B.; Perugino, G.; Strazzulli, A.; Rossi, M.; Moracci, M. Thermophilic glycosynthases for oligosaccharides synthesis. Methods Enzymol. 2012, 510, 273–300. [Google Scholar]
- Menzel, P.; Ng, K.L.; Krogh, A. Fast and sensitive taxonomic classification for metagenomics with Kaiju. Nat. Commun. 2016, 7, 11257. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Liu, C.M.; Luo, R.; Sadakane, K.; Lam, T.W. MEGAHIT: An ultra-fast single-node solution for large and complex metagenomics assembly via succinct de Bruijn graph. Bioinformatics 2015, 31, 1674–1676. [Google Scholar] [CrossRef] [PubMed]
- Hyatt, D.; Chen, G.L.; Locascio, P.F.; Land, M.L.; Larimer, F.W.; Hauser, L.J. Prodigal: Prokaryotic gene recognition and translation initiation site identification. BMC Bioinform. 2010, 11, 119. [Google Scholar] [CrossRef] [PubMed]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrates | VMAX (U mg−1) | KM (mM) | kcat (s−1) | kcat/KM (s−1 mM−1) |
|---|---|---|---|---|
| 4Np-α-Glc | 42.5 ± 1.1 | 3.59 ± 0.33 | 327.4 ± 8.4 | 91.14 |
| Maltose | 117.1 ± 6.9 | 3.16 ± 0.65 | 901.6 ± 53.6 | 284.97 |
| Maltotriose | 89.7 ± 3.2 | 1.03 ± 0.20 | 690.7 ± 24.4 | 670.64 |
| Kojibiose | 65.0 ± 1.9 | 1.41 ± 0.19 | 500.2 ± 14.6 | 352.49 |
| Substrate | Kinetic Parameter | Aglu1 a | MalA b S. solfataricus | AglA c P. torridus | MalA d S. acidocaldarius | ST2525 e S. tokodaii | AglA f T. acidophilum | |
|---|---|---|---|---|---|---|---|---|
| 1 | 4Np-α-Glc | KM (mM) | 3.59 ± 0.33 | 1.7 ± 0.2 | 0.94 ± 0.14 | 0.87 ± 0.18 | 3.18 | 1.77 |
| kcat (s−1) | 327.4 ± 8.3 | 38.5 ± 1.6 | 65.1 | 16.7 | 1.05 | 59.7 | ||
| kcat/KM (s−1 mM−1) | 91.14 | 22.1 | 69.25 | 19.2 | 0.33 | 33.7 | ||
| 2 | Maltose | KM (mM) | 3.16 ± 0.65 | 2.00 ± 0.12 a | 3.70 ± 0.32 | 0.80 ± 0.32 | 10.9 | 0.65 |
| kcat (s−1) | 901.6 ± 53.5 | 481.1 ± 6 10−5 a | 188.1 | 1277.8 | 6.5 | 143 | ||
| kcat/KM (s−1 mM−1) | 285.0 | 239.6 a | 50.83 | 1597.2 | 0.598 | 220 | ||
| 3 | Maltotriose | KM (mM) | 1.03 ± 0.20 | ND | ND | 0.30 ± 0.08 | 3.11 | 0.47 |
| kcat (s−1) | 690.7 ± 24.4 | 550 | 2.53 | 90.2 | ||||
| kcat/KM (s−1 mM−1) | 670.64 | 1833.3 | 0.814 | 194 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iacono, R.; Strazzulli, A.; Giglio, R.; Bitetti, F.; Cobucci-Ponzano, B.; Moracci, M. Valorization of Biomasses from Energy Crops for the Discovery of Novel Thermophilic Glycoside Hydrolases through Metagenomic Analysis. Int. J. Mol. Sci. 2022, 23, 10505. https://doi.org/10.3390/ijms231810505
Iacono R, Strazzulli A, Giglio R, Bitetti F, Cobucci-Ponzano B, Moracci M. Valorization of Biomasses from Energy Crops for the Discovery of Novel Thermophilic Glycoside Hydrolases through Metagenomic Analysis. International Journal of Molecular Sciences. 2022; 23(18):10505. https://doi.org/10.3390/ijms231810505
Chicago/Turabian StyleIacono, Roberta, Andrea Strazzulli, Rosa Giglio, Federica Bitetti, Beatrice Cobucci-Ponzano, and Marco Moracci. 2022. "Valorization of Biomasses from Energy Crops for the Discovery of Novel Thermophilic Glycoside Hydrolases through Metagenomic Analysis" International Journal of Molecular Sciences 23, no. 18: 10505. https://doi.org/10.3390/ijms231810505
APA StyleIacono, R., Strazzulli, A., Giglio, R., Bitetti, F., Cobucci-Ponzano, B., & Moracci, M. (2022). Valorization of Biomasses from Energy Crops for the Discovery of Novel Thermophilic Glycoside Hydrolases through Metagenomic Analysis. International Journal of Molecular Sciences, 23(18), 10505. https://doi.org/10.3390/ijms231810505