
Ciliary Neurotrophic Factor (CNTF) Inhibits In Vitro Cementoblast Mineralization and Induces Autophagy, in Part by STAT3/ERK Commitment


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
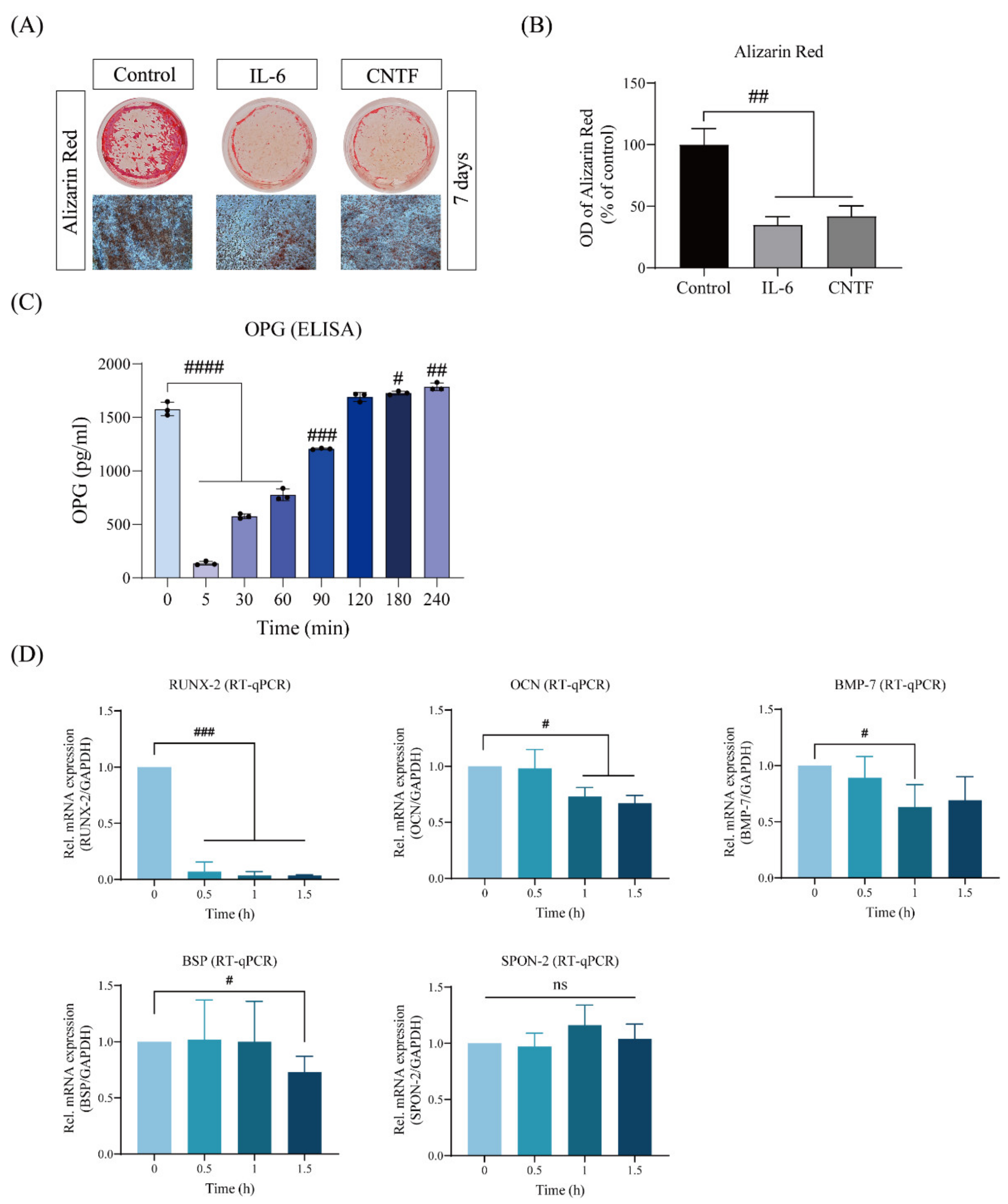
2.1. CNTF Impairs Cementoblast Mineralization and Impairs the Expression of Cementogenesis Hallmarkers
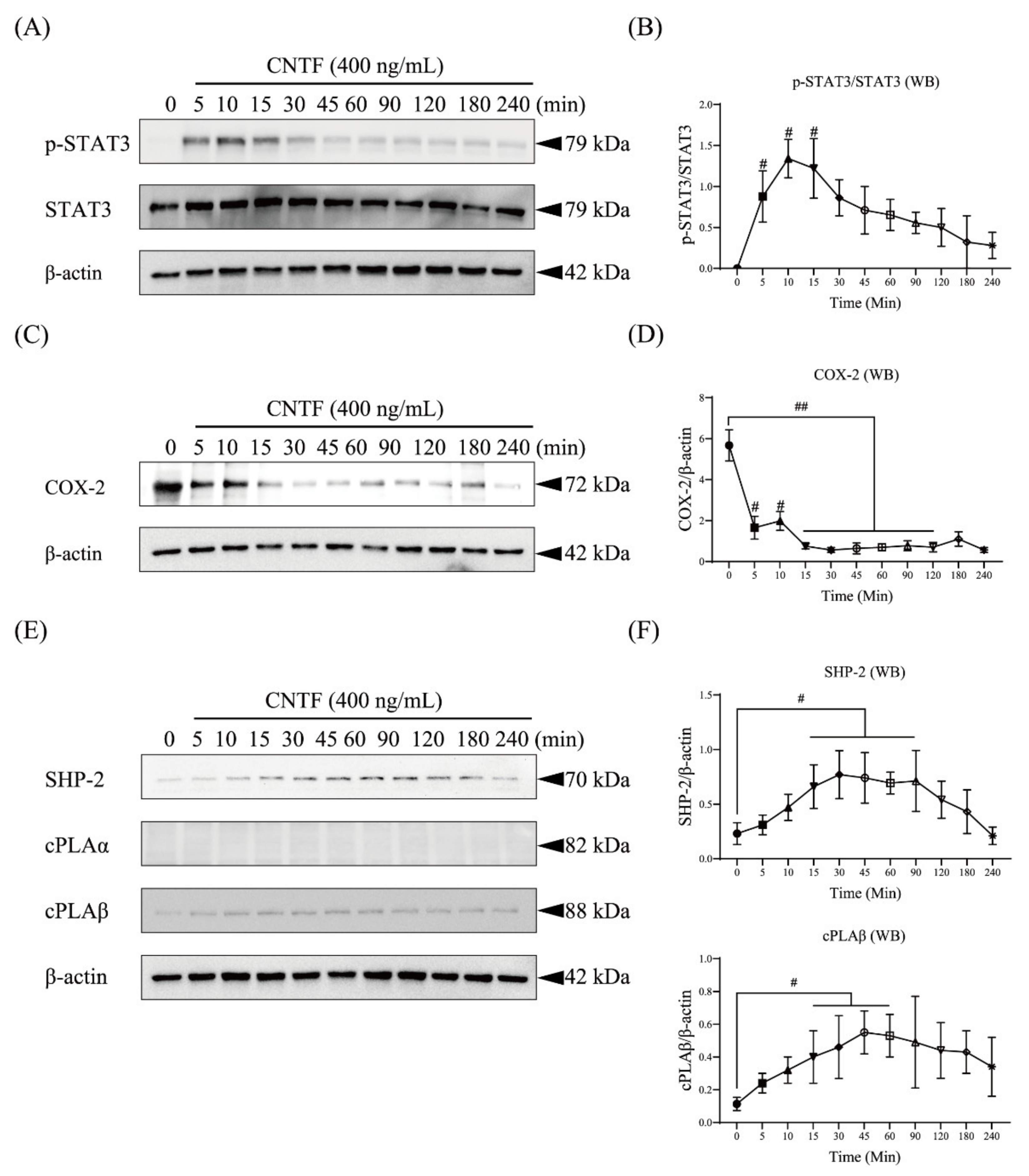
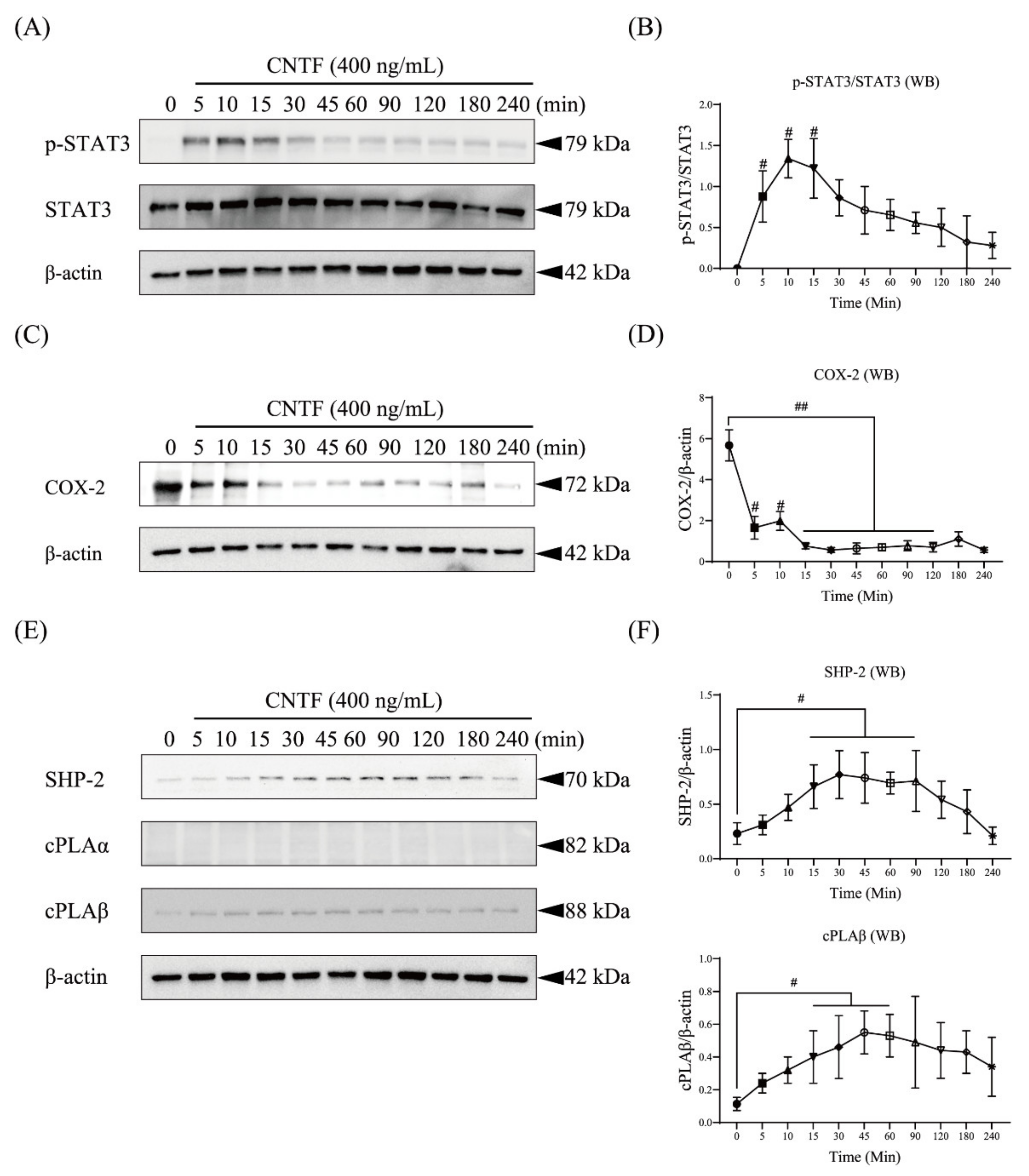
2.2. CNTF Regulates STAT3 as Well as COX-2, SHP-2, and cPLAβ Expression in OCCM-30 Cells
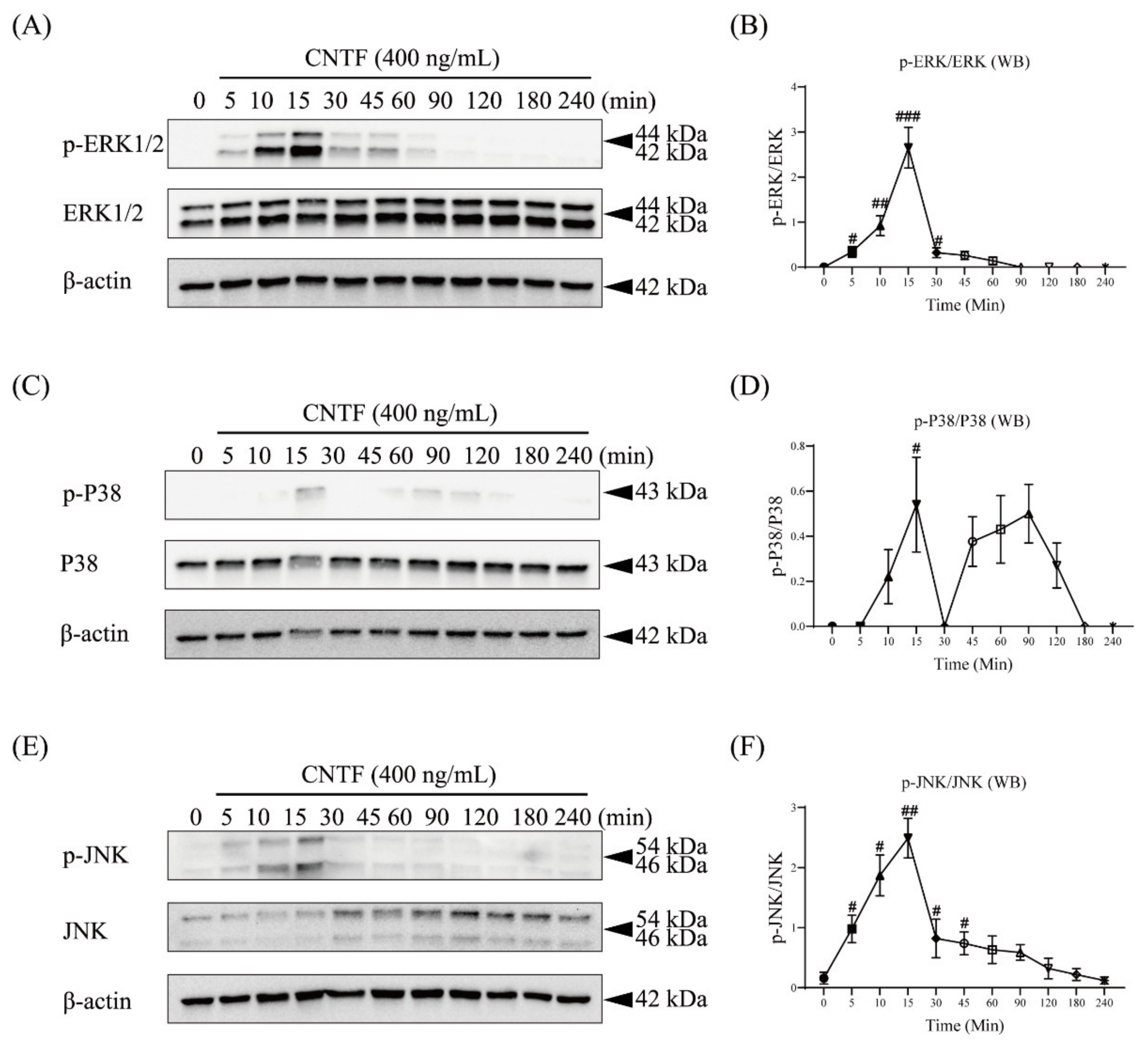
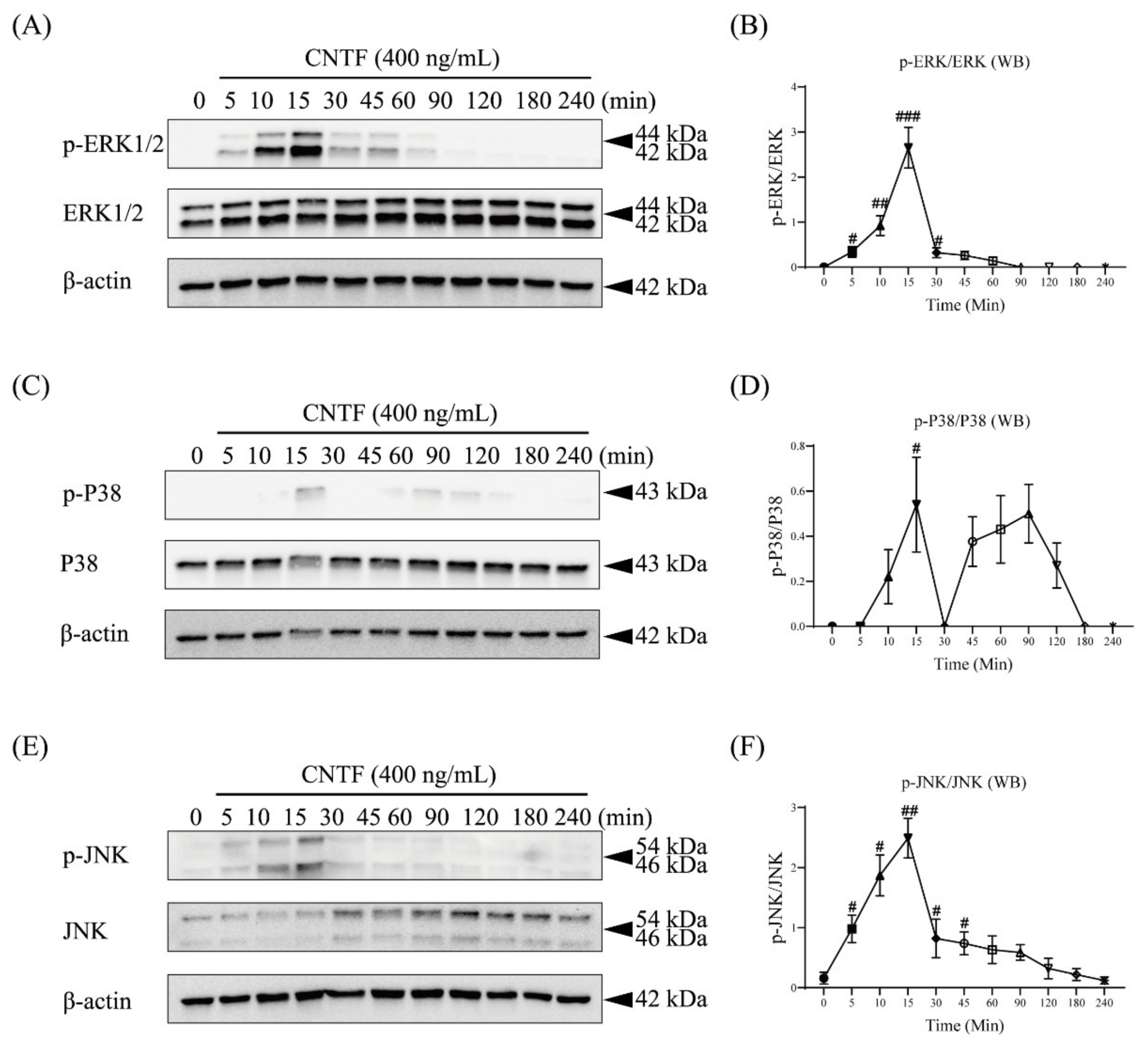
2.3. CNTF Triggers ERK1/2, P38, and JNK Phosphorylation in Cementoblasts
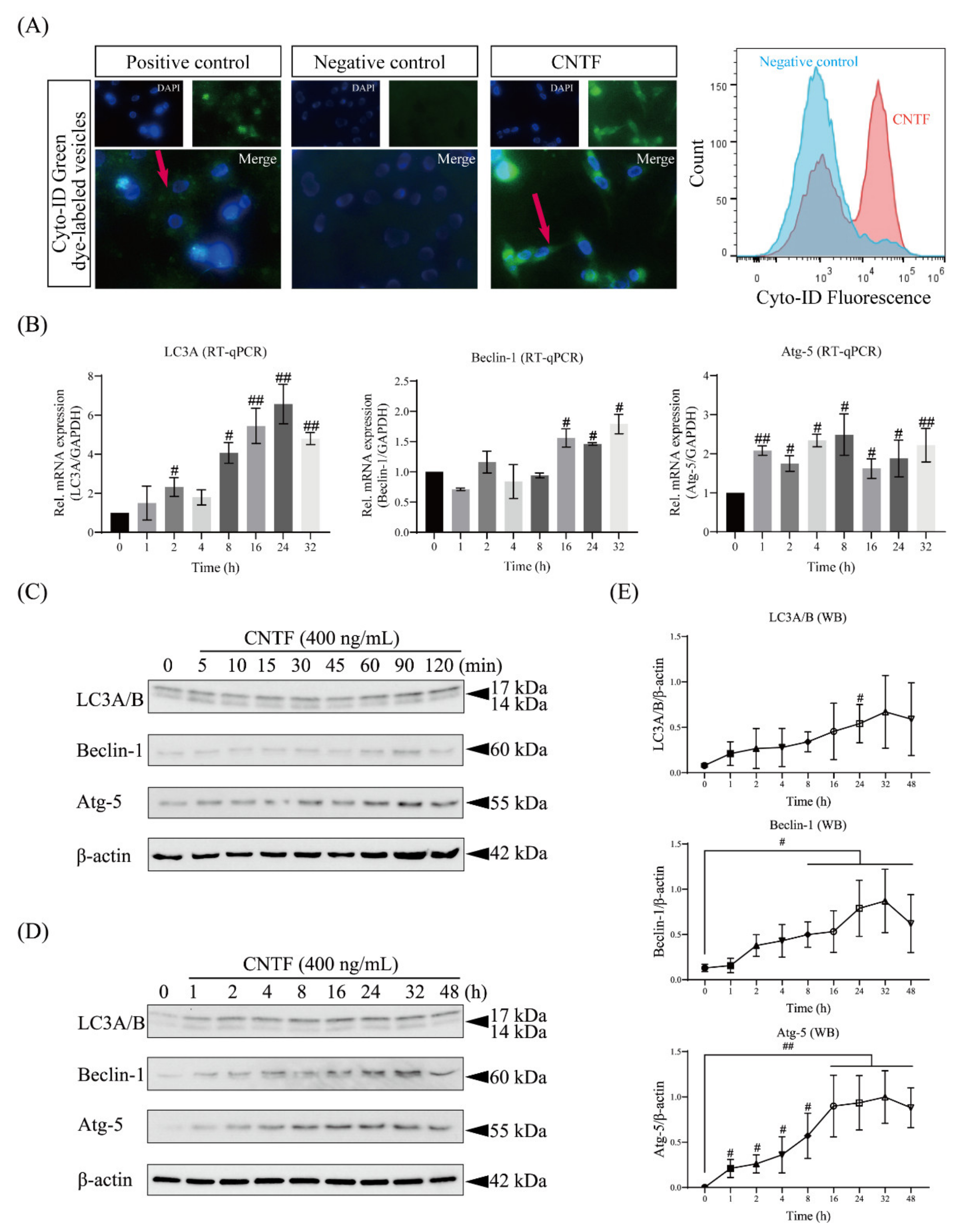
2.4. CNTF Induces the Autophagosome and Activates Autophagic Signaling
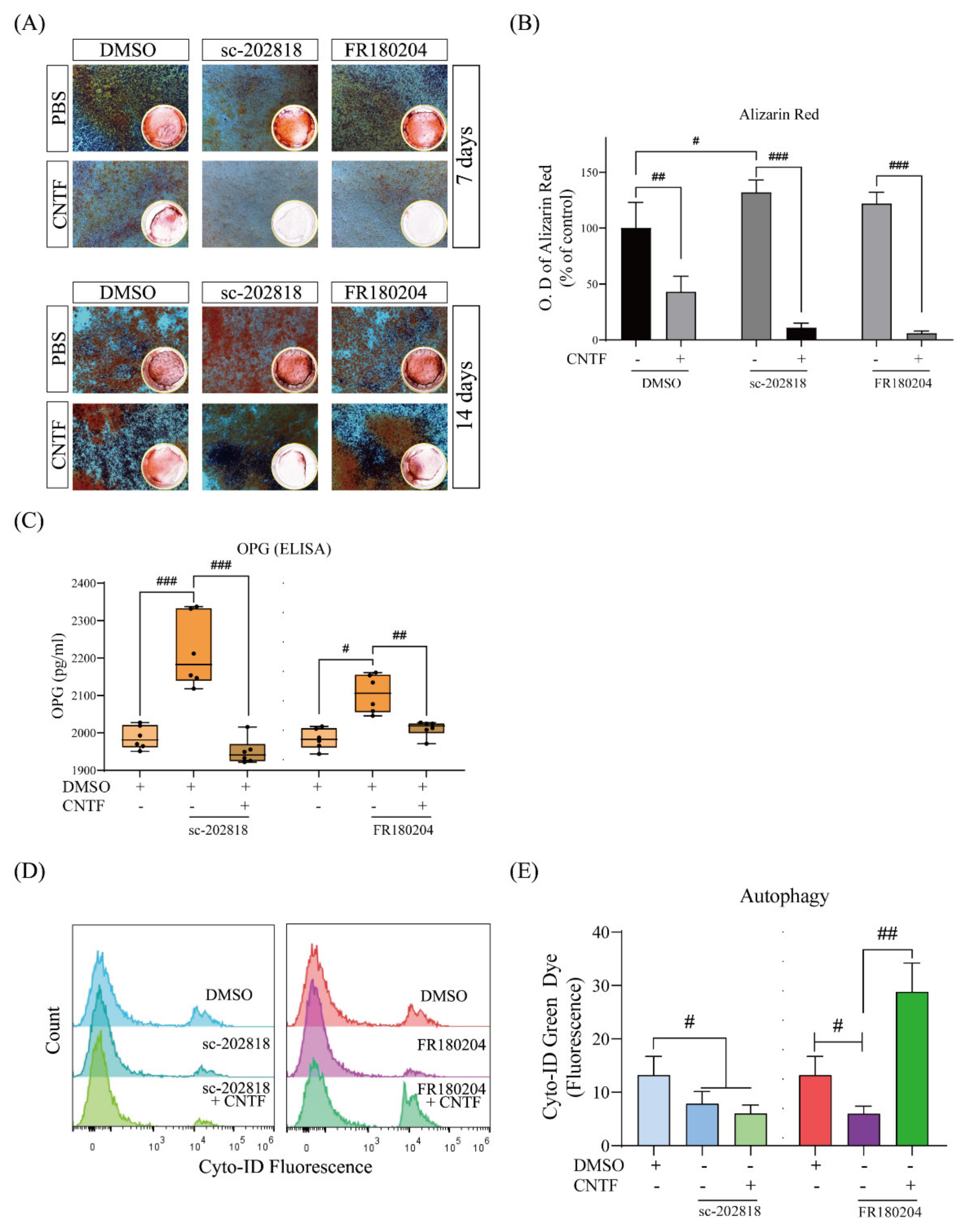
2.5. STAT3/ERK Are Involved in CNTF-Induced Inhibitory Cementoblast Mineralization Effects and Autophagy Induction
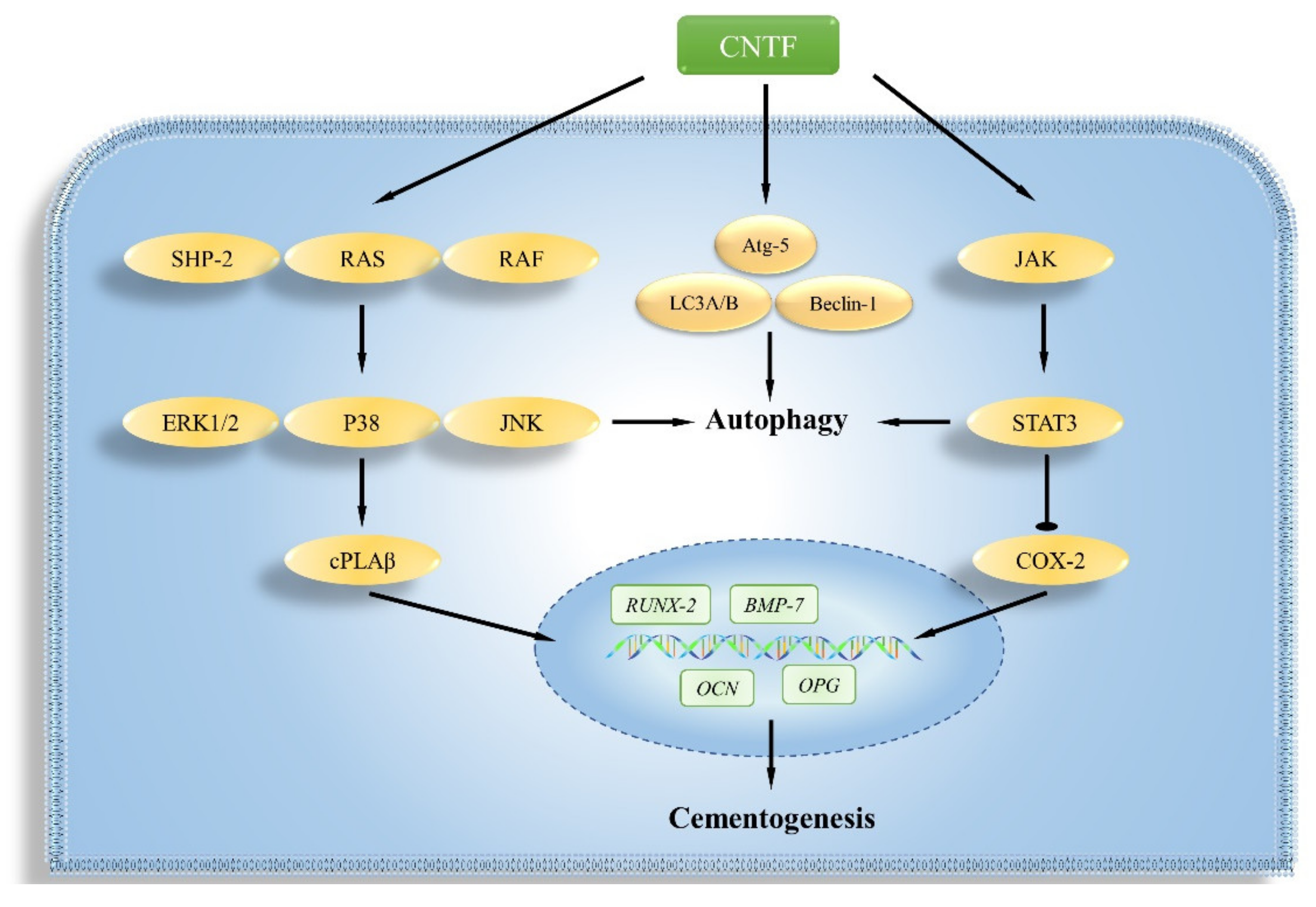
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Conditions
4.2. Differentiation (Mineralization) Induction
4.3. Reagents and Pharmacological Inhibitor
4.4. Alizarin Red Assay
4.5. Enzyme-Linked Immunosorbent Assay (ELISA)
4.6. Gene Expression Analysis
4.7. Protein Expression Analysis
4.8. Autophagy Detection (Autophagosome Quantification)
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Askmyr, M.; White, K.E.; Jovic, T.; King, H.A.; Quach, J.M.; Maluenda, A.C.; Baker, E.K.; Smeets, M.F.; Walkley, C.R.; Purton, L.E. Ciliary neurotrophic factor has intrinsic and extrinsic roles in regulating B cell differentiation and bone structure. Sci. Rep. 2015, 5, 15529. [Google Scholar] [CrossRef] [PubMed]
- Kruttgen, A.; Grotzinger, J.; Kurapkat, G.; Weis, J.; Simon, R.; Thier, M.; Schroder, M.; Heinrich, P.; Wollmer, A.; Comeau, M.; et al. Human ciliary neurotrophic factor: A structure-function analysis. Biochem. J. 1995, 309, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.F.; Mismer, D.; Lile, J.D.; Armes, L.G.; Butler, E.T., 3rd.; Vannice, J.L.; Collins, F. Purification, cloning, and expression of ciliary neurotrophic factor (CNTF). Science 1989, 246, 1023–1025. [Google Scholar] [CrossRef] [PubMed]
- Pasquin, S.; Sharma, M.; Gauchat, J.F. Ciliary neurotrophic factor (CNTF): New facets of an old molecule for treating neurodegenerative and metabolic syndrome pathologies. Cytokine Growth Factor Rev. 2015, 26, 507–515. [Google Scholar] [CrossRef]
- McGregor, N.E.; Poulton, I.J.; Walker, E.C.; Pompolo, S.; Quinn, J.M.; Martin, T.J.; Sims, N.A. Ciliary neurotrophic factor inhibits bone formation and plays a sex-specific role in bone growth and remodeling. Calcif. Tissue Int. 2010, 86, 261–270. [Google Scholar] [CrossRef]
- Yong, J.; von Bremen, J.; Ruiz-Heiland, G.; Ruf, S. Adiponectin as Well as Compressive Forces Regulate in vitro beta-Catenin Expression on Cementoblasts via Mitogen-Activated Protein Kinase Signaling Activation. Front. Cell Dev. Biol. 2021, 9, 645005. [Google Scholar] [CrossRef]
- Yong, J.; von Bremen, J.; Ruiz-Heiland, G.; Ruf, S. Adiponectin Interacts In-Vitro with Cementoblasts Influencing Cell Migration, Proliferation and Cementogenesis Partly Through the MAPK Signaling Pathway. Front. Pharmacol. 2020, 11, 585346. [Google Scholar] [CrossRef]
- Hakki, S.S.; Foster, B.L.; Nagatomo, K.J.; Bozkurt, S.B.; Hakki, E.E.; Somerman, M.J.; Nohutcu, R.M. Bone morphogenetic protein-7 enhances cementoblast function in vitro. J. Periodontol. 2010, 81, 1663–1674. [Google Scholar] [CrossRef]
- Yong, J.; Groeger, S.; von Bremen, J.; Ruf, S. Ciliary Neurotrophic Factor (CNTF) and Its Receptors Signal Regulate Cementoblasts Apoptosis through a Mechanism of ERK1/2 and Caspases Signaling. Int. J. Mol. Sci. 2022, 23, 8335. [Google Scholar] [CrossRef]
- Mayr, A.; Marciniak, J.; Eggers, B.; Blawat, K.; Wildenhof, J.; Bastos Craveiro, R.; Wolf, M.; Deschner, J.; Jager, A.; Beisel-Memmert, S. Autophagy Induces Expression of IL-6 in Human Periodontal Ligament Fibroblasts Under Mechanical Load and Overload and Effects Osteoclastogenesis in vitro. Front. Physiol. 2021, 12, 716441. [Google Scholar] [CrossRef]
- Mizushima, N. Autophagy. FEBS Lett. 2010, 584, 1279. [Google Scholar] [CrossRef] [PubMed]
- Murrow, L.; Debnath, J. Autophagy as a stress-response and quality-control mechanism: Implications for cell injury and human disease. Annu. Rev. Pathol. 2013, 8, 105–137. [Google Scholar] [CrossRef] [PubMed]
- Kabeya, Y.; Mizushima, N.; Ueno, T.; Yamamoto, A.; Kirisako, T.; Noda, T.; Kominami, E.; Ohsumi, Y.; Yoshimori, T. LC3, a mammalian homologue of yeast Apg8p, is localized in autophagosome membranes after processing. EMBO J. 2000, 19, 5720–5728. [Google Scholar] [CrossRef] [PubMed]
- An, Y.; Liu, W.; Xue, P.; Zhang, Y.; Wang, Q.; Jin, Y. Increased autophagy is required to protect periodontal ligament stem cells from apoptosis in inflammatory microenvironment. J. Clin. Periodontol. 2016, 43, 618–625. [Google Scholar] [CrossRef]
- Pei, F.; Lin, H.; Liu, H.; Li, L.; Zhang, L.; Chen, Z. Dual role of autophagy in lipopolysaccharide-induced preodontoblastic cells. J. Dent. Res. 2015, 94, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Yong, J.; Groeger, S.; Meyle, J.; Ruf, S. MAPK and β-Catenin signaling: Implication and interplay in orthodontic tooth movement. Front. Biosci. Landmark 2022, 27, 54. [Google Scholar] [CrossRef] [PubMed]
- Huitema, L.F.; Vaandrager, A.B. What triggers cell-mediated mineralization? Front. Biosci. 2007, 12, 2631–2645. [Google Scholar] [CrossRef]
- Johnson, R.W.; White, J.D.; Walker, E.C.; Martin, T.J.; Sims, N.A. Myokines (muscle-derived cytokines and chemokines) including ciliary neurotrophic factor (CNTF) inhibit osteoblast differentiation. Bone 2014, 64, 47–56. [Google Scholar] [CrossRef]
- Trimarchi, T.; Pachuau, J.; Shepherd, A.; Dey, D.; Martin-Caraballo, M. CNTF-evoked activation of JAK and ERK mediates the functional expression of T-type Ca2+ channels in chicken nodose neurons. J. Neurochem. 2009, 108, 246–259. [Google Scholar] [CrossRef]
- Wang, L.; Wang, Y.; Du, M.; Liu, Z.; Cao, Z.; Hao, Y.; He, H. Inhibition of Stat3 signaling pathway decreases TNF-alpha-induced autophagy in cementoblasts. Cell Tissue Res. 2018, 374, 567–575. [Google Scholar] [CrossRef]
- Ma, L.; Liu, H.; Wang, X.; Jiang, C.; Yao, S.; Guo, Y.; Wang, H.; Cao, Z. CXXC5 orchestrates Stat3/Erk/Akt signaling networks to modulate P. gingivalis-elicited autophagy in cementoblasts. Biochim. Biophys. Acta Mol. Cell Res. 2021, 1868, 118923. [Google Scholar] [CrossRef] [PubMed]
- You, L.; Wang, Z.; Li, H.; Shou, J.; Jing, Z.; Xie, J.; Sui, X.; Pan, H.; Han, W. The role of STAT3 in autophagy. Autophagy 2015, 11, 729–739. [Google Scholar] [CrossRef] [PubMed]
- Vrahnas, C.; Blank, M.; Dite, T.A.; Tatarczuch, L.; Ansari, N.; Crimeen-Irwin, B.; Nguyen, H.; Forwood, M.R.; Hu, Y.; Ikegame, M.; et al. Increased autophagy in EphrinB2-deficient osteocytes is associated with elevated secondary mineralization and brittle bone. Nat. Commun. 2019, 10, 3436. [Google Scholar] [CrossRef] [PubMed]
- Nollet, M.; Santucci-Darmanin, S.; Breuil, V.; Al-Sahlanee, R.; Cros, C.; Topi, M.; Momier, D.; Samson, M.; Pagnotta, S.; Cailleteau, L.; et al. Autophagy in osteoblasts is involved in mineralization and bone homeostasis. Autophagy 2014, 10, 1965–1977. [Google Scholar] [CrossRef]
- Chen, L.; Hua, Y. Autophagy of periodontal ligament inhibits inflammation and reduces the decline of bone density during orthodontic tooth movement of mice. Arch. Oral Biol. 2021, 121, 104960. [Google Scholar] [CrossRef]
- Redmann, M.; Benavides, G.A.; Berryhill, T.F.; Wani, W.Y.; Ouyang, X.; Johnson, M.S.; Ravi, S.; Barnes, S.; Darley-Usmar, V.M.; Zhang, J. Inhibition of autophagy with bafilomycin and chloroquine decreases mitochondrial quality and bioenergetic function in primary neurons. Redox Biol. 2017, 11, 73–81. [Google Scholar] [CrossRef]
- Ha, S.W.; Weitzmann, M.N.; Beck, G.R., Jr. Bioactive silica nanoparticles promote osteoblast differentiation through stimulation of autophagy and direct association with LC3 and p62. ACS Nano 2014, 8, 5898–5910. [Google Scholar] [CrossRef] [PubMed]
- Yong, J.; von Bremen, J.; Groeger, S.; Ruiz-Heiland, G.; Ruf, S. Hypoxia-inducible factor 1-alpha acts as a bridge factor for crosstalk between ERK1/2 and caspases in hypoxia-induced apoptosis of cementoblasts. J. Cell. Mol. Med. 2021, 25, 9710–9723. [Google Scholar] [CrossRef]
- Yong, J.; Gröger, S.; Meyle, J.; Ruf, S. Immunorthodontics: Role of HIF-1α in the Regulation of (Peptidoglycan-Induced) PD-L1 Expression in Cementoblasts under Compressive Force. Int. J. Mol. Sci. 2022, 23, 6977. [Google Scholar] [CrossRef]
- Yong, J.; Gröger, S.; von Bremen, J.; Meyle, J.; Ruf, S. Immunorthodontics: PD-L1, a Novel Immunomodulator in Cementoblasts, Is Regulated by HIF-1α under Hypoxia. Cells 2022, 11, 2350. [Google Scholar] [CrossRef]
- Yong, J.; Li, P.; Mizrahi, I.K.; Franzen, R.; Groeger, S.; Ruf, S.; Gutknecht, N.; Marques, M.M. Effect of Low-Level Er: YAG (2940 nm) laser irradiation on the photobiomodulation of mitogen-activated protein kinase cellular signaling pathway of rodent cementoblasts. Front. Biosci. (Landmark Ed.) 2022, 27, 62. [Google Scholar] [CrossRef]
- Yong, J.; Groeger, S.; Ruf, S.; Ruiz-Heiland, G. Influence of leptin and compression in GAS-6 mediated homeostasis of periodontal ligament cell. Oral Dis. 2021, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Yong, J.; Groeger, S.; Ruiz-Heiland, G.; Ruf, S. Selection and validation of reference gene for RT-qPCR studies in co-culture system of mouse cementoblasts and periodontal ligament cells. BMC Res. Notes 2022, 15, 57. [Google Scholar] [CrossRef] [PubMed]
- Memmert, S.; Damanaki, A.; Weykopf, B.; Rath-Deschner, B.; Nokhbehsaim, M.; Gotz, W.; Golz, L.; Till, A.; Deschner, J.; Jager, A. Autophagy in periodontal ligament fibroblasts under biomechanical loading. Cell Tissue Res. 2019, 378, 499–511. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yong, J.; Gröger, S.; von Bremen, J.; Ruf, S. Ciliary Neurotrophic Factor (CNTF) Inhibits In Vitro Cementoblast Mineralization and Induces Autophagy, in Part by STAT3/ERK Commitment. Int. J. Mol. Sci. 2022, 23, 9311. https://doi.org/10.3390/ijms23169311
Yong J, Gröger S, von Bremen J, Ruf S. Ciliary Neurotrophic Factor (CNTF) Inhibits In Vitro Cementoblast Mineralization and Induces Autophagy, in Part by STAT3/ERK Commitment. International Journal of Molecular Sciences. 2022; 23(16):9311. https://doi.org/10.3390/ijms23169311
Chicago/Turabian StyleYong, Jiawen, Sabine Gröger, Julia von Bremen, and Sabine Ruf. 2022. "Ciliary Neurotrophic Factor (CNTF) Inhibits In Vitro Cementoblast Mineralization and Induces Autophagy, in Part by STAT3/ERK Commitment" International Journal of Molecular Sciences 23, no. 16: 9311. https://doi.org/10.3390/ijms23169311
APA StyleYong, J., Gröger, S., von Bremen, J., & Ruf, S. (2022). Ciliary Neurotrophic Factor (CNTF) Inhibits In Vitro Cementoblast Mineralization and Induces Autophagy, in Part by STAT3/ERK Commitment. International Journal of Molecular Sciences, 23(16), 9311. https://doi.org/10.3390/ijms23169311