
Telocytes’ Role in Modulating Gut Motility Function and Development: Medical Hypotheses and Literature Review


 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. TCs and Inflammatory Bowel Disease (IBD)
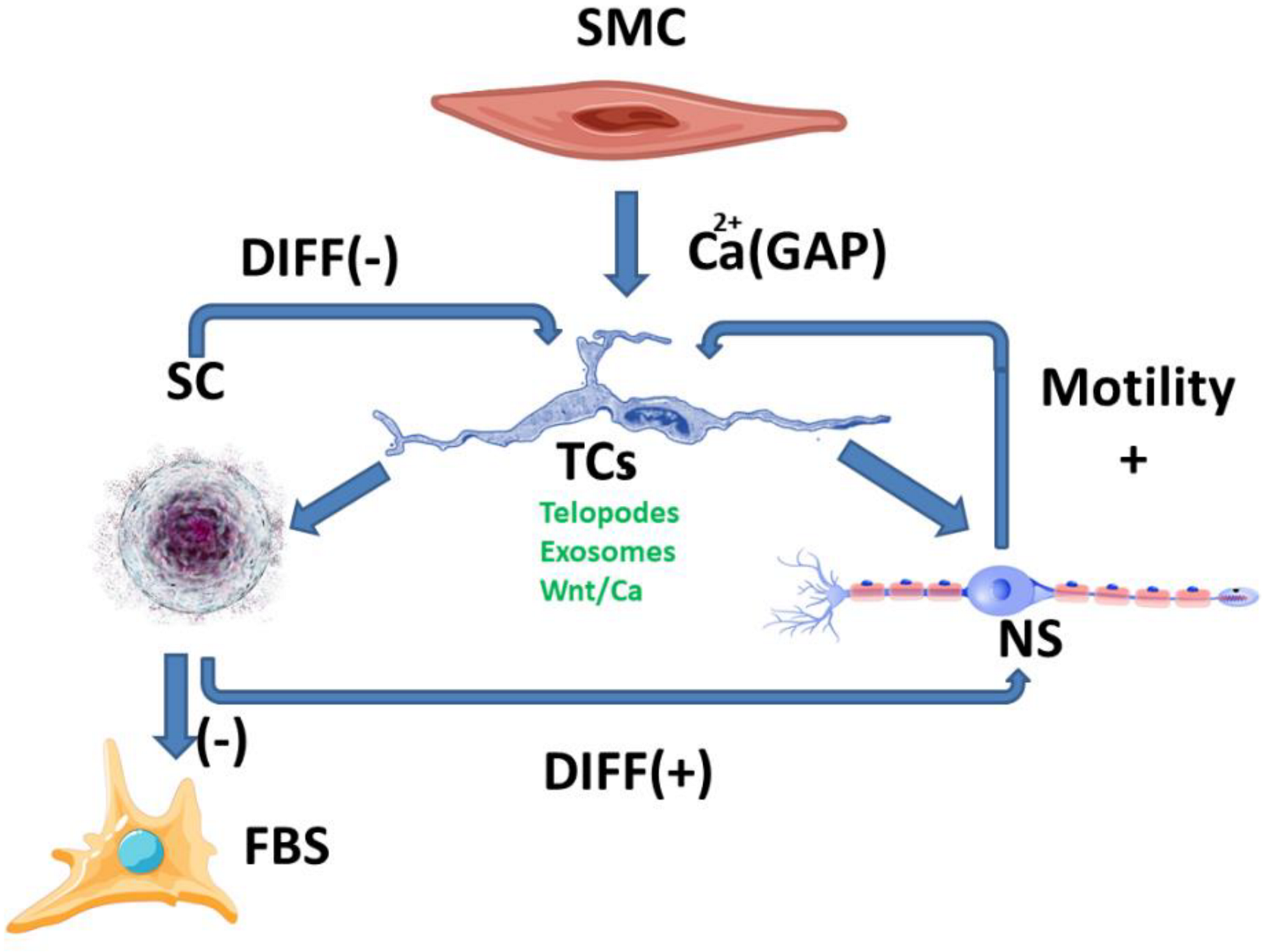
3. The Proposed Functional Model of TCs Signaling
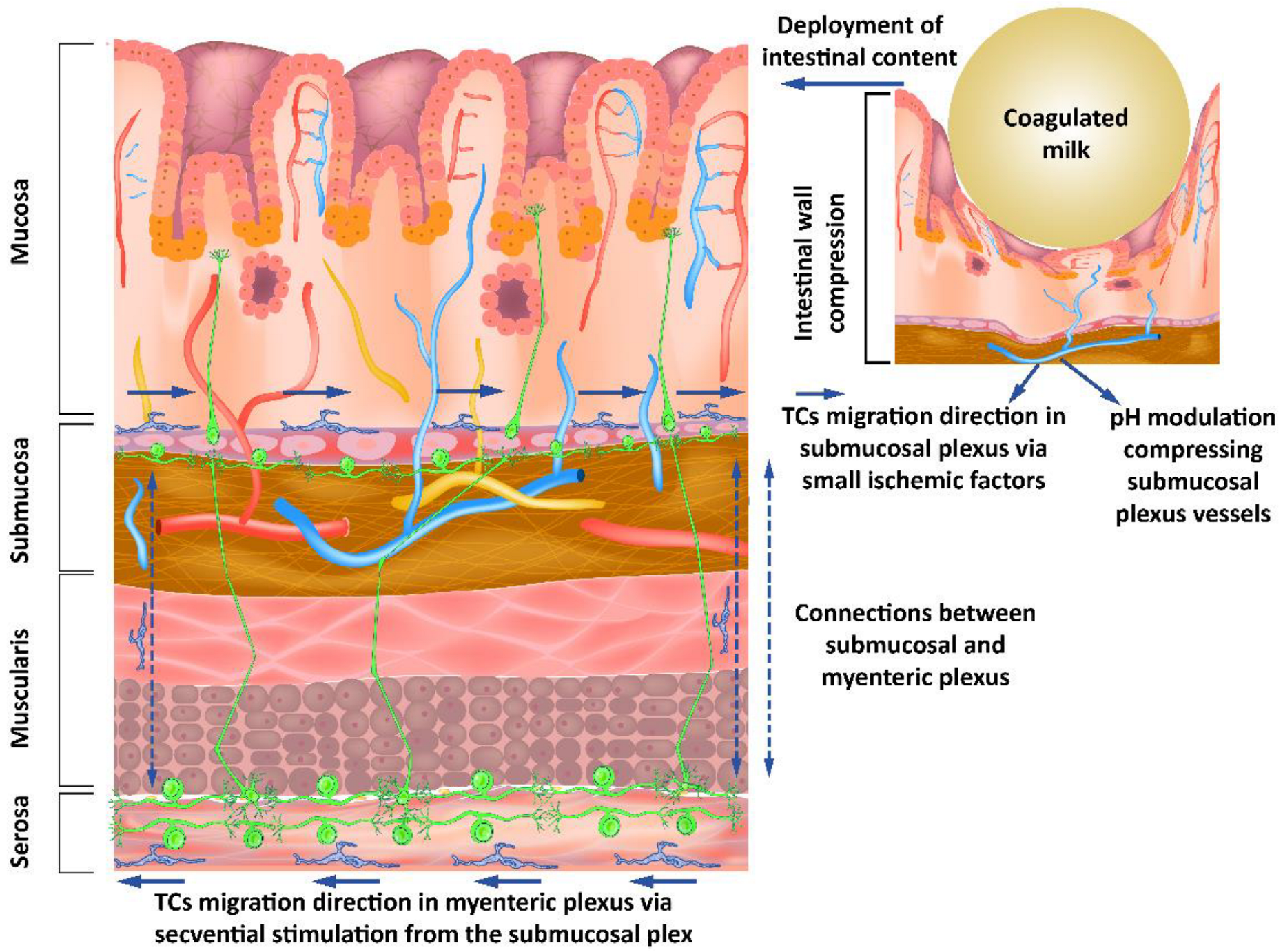
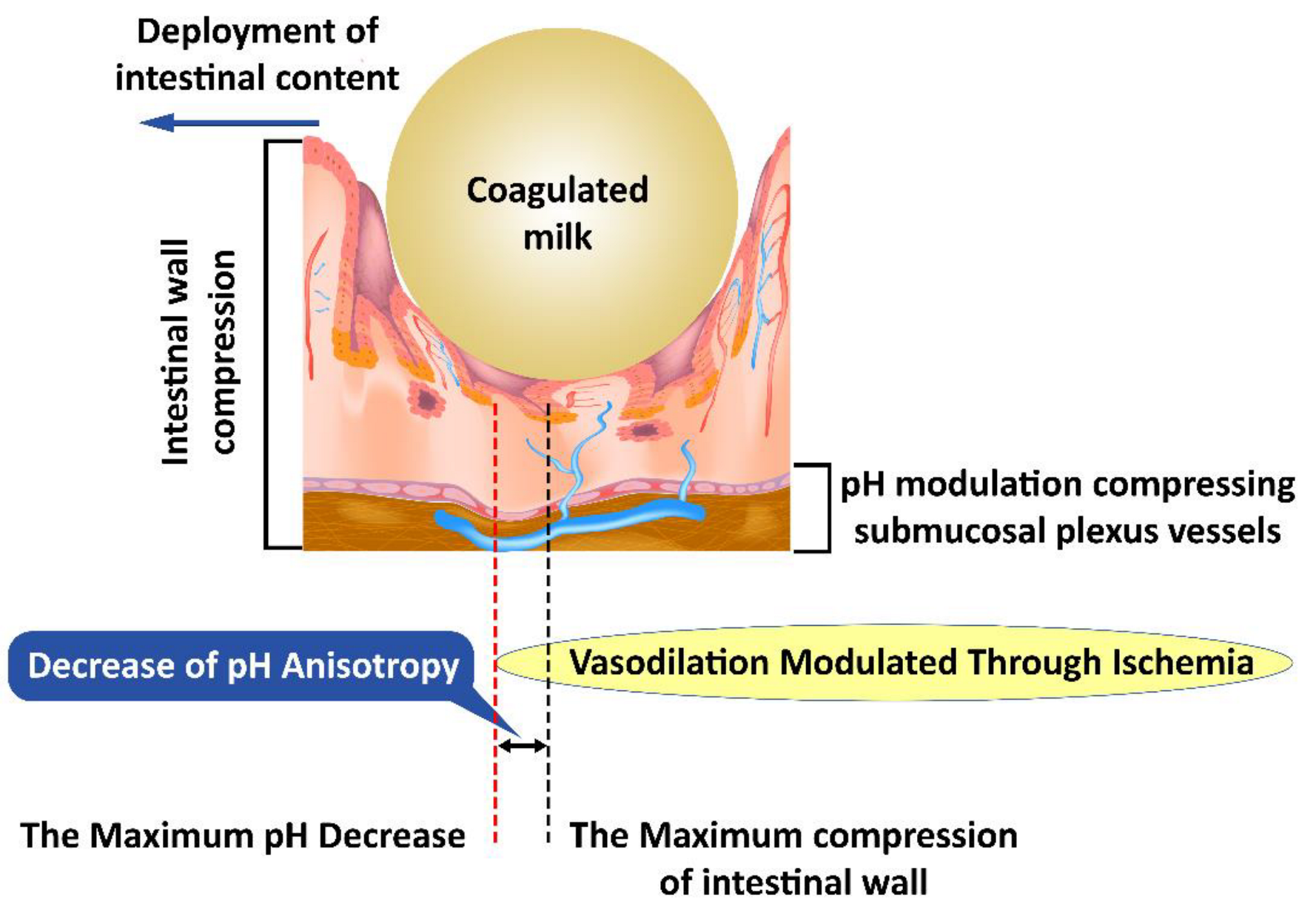
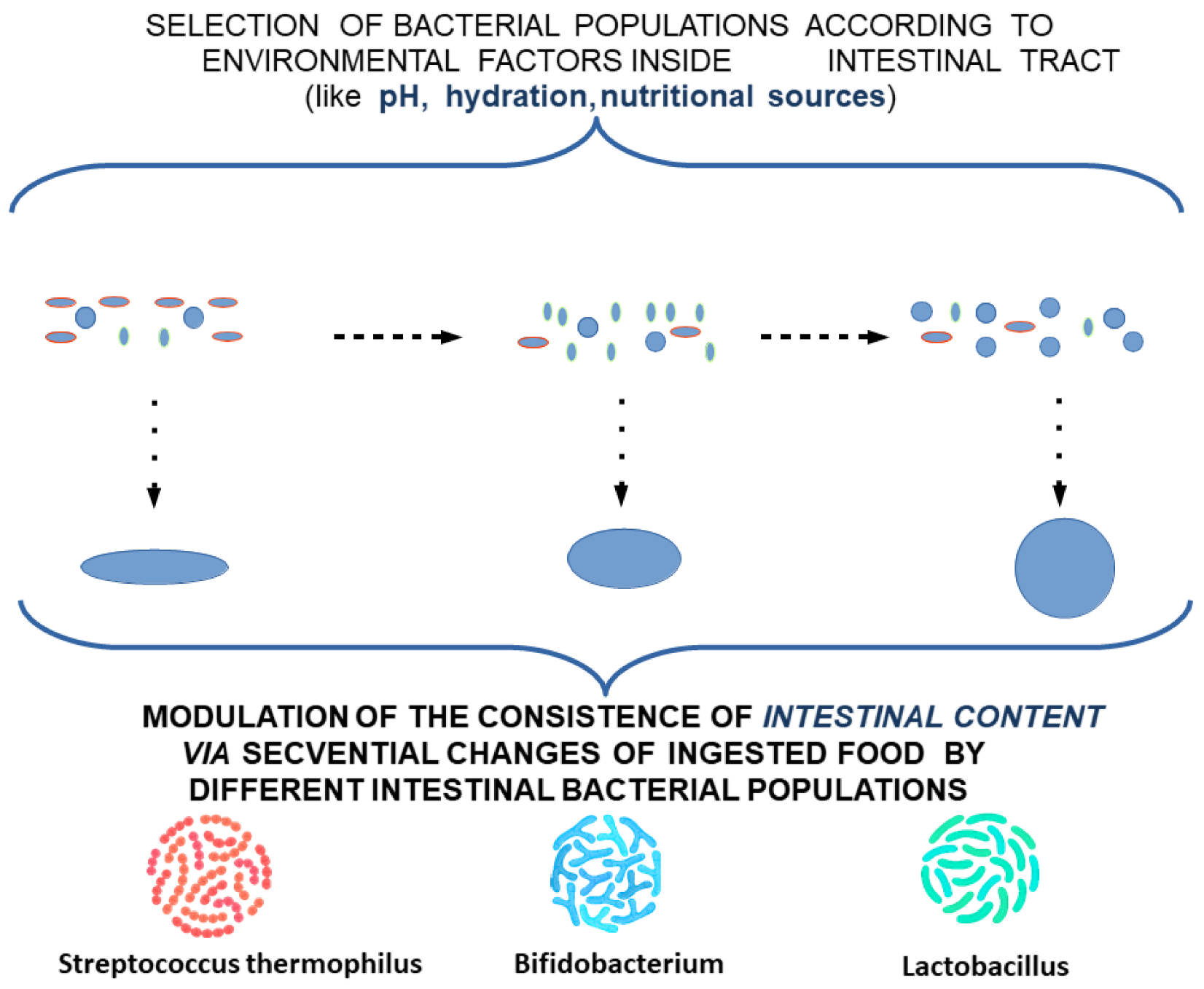
4. Intestinal Colics as a Simplified Model for Understanding the Role of TCs in Digestive Motility
5. CD, UC, and Irritable Colon as Models of Validation of the Proposed Theoretical Model
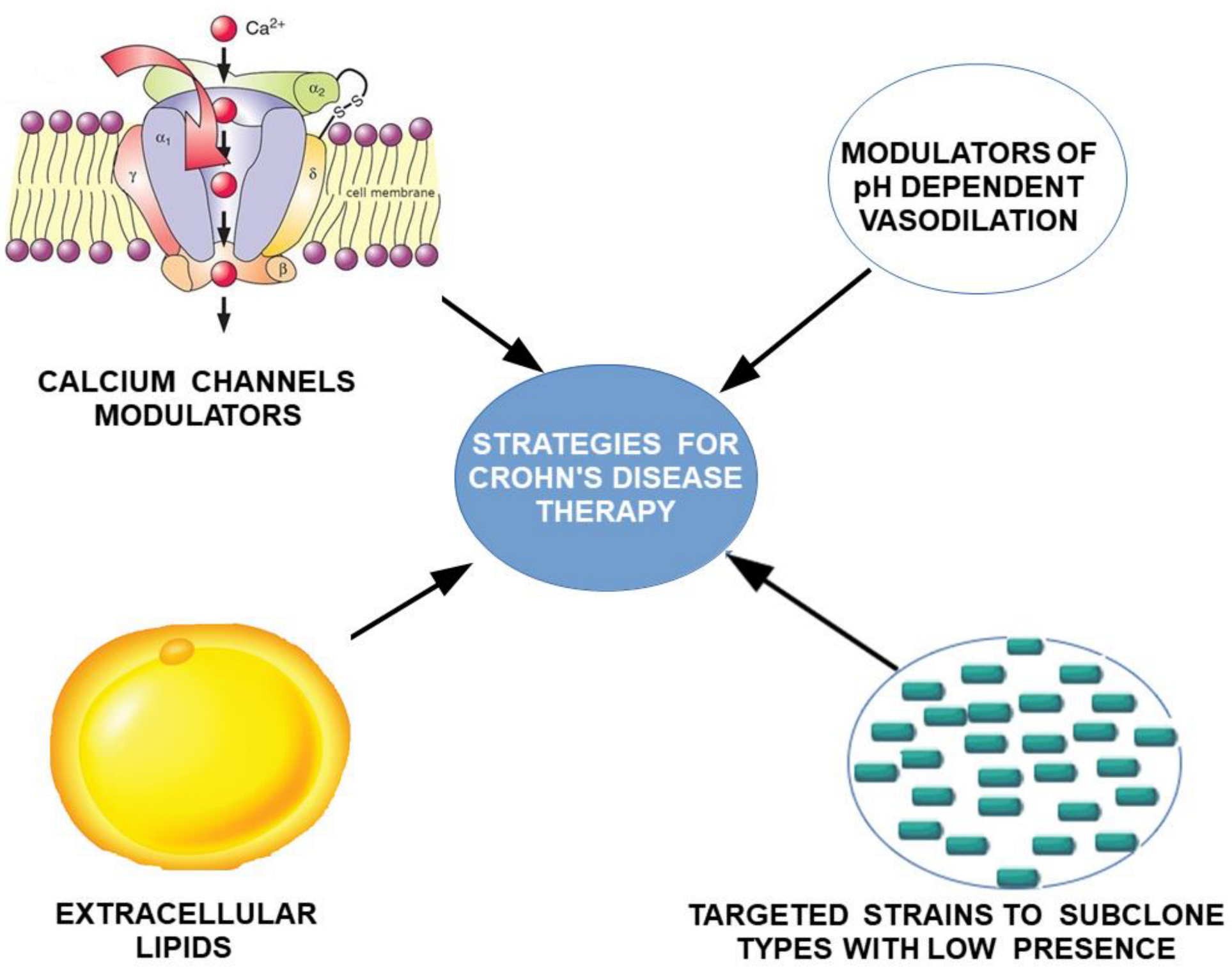
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Popescu, L.M.; Faussone-Pellegrini, M.S. TELOCYTES—A case of serendipity: The winding way from Interstitial Cells of Cajal (ICC), via Interstitial Cajal-Like Cells (ICLC) to TELOCYTES. J. Cell. Mol. Med. 2010, 14, 729–740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cretoiu, S.M.; Popescu, L.M. Telocytes revisited. Biomol. Concepts 2014, 5, 353–369. [Google Scholar] [CrossRef] [PubMed]
- Cretoiu, D.; Radu, B.M.; Banciu, A.; Banciu, D.D.; Cretoiu, S.M. Telocytes heterogeneity: From cellular morphology to functional evidence. Semin Cell Dev. Biol. 2017, 64, 26–39. [Google Scholar] [CrossRef] [PubMed]
- Campeanu, R.A.; Radu, B.M.; Cretoiu, S.M.; Banciu, D.D.; Banciu, A.; Cretoiu, D.; Popescu, L.M. Near-infrared low-level laser stimulation of telocytes from human myometrium. Lasers Med. Sci. 2014, 29, 1867–1874. [Google Scholar] [CrossRef] [Green Version]
- Popescu, L.M.; Fertig, E.T.; Gherghiceanu, M. Reaching out: Junctions between cardiac telocytes and cardiac stem cells in culture. J. Cell Mol. Med. 2016, 20, 370–380. [Google Scholar] [CrossRef] [Green Version]
- Albulescu, R.; Tanase, C.; Codrici, E.; Popescu, D.I.; Cretoiu, S.M.; Popescu, L.M. The secretome of myocardial telocytes modulates the activity of cardiac stem cells. J. Cell Mol. Med. 2015, 19, 1783–1794. [Google Scholar] [CrossRef]
- Cretoiu, D.; Xu, J.; Xiao, J.; Cretoiu, S.M. Telocytes and Their Extracellular Vesicles-Evidence and Hypotheses. Int. J. Mol. Sci. 2016, 17, 1322. [Google Scholar] [CrossRef] [Green Version]
- Ratajczak, M.Z.; Ratajczak, D.; Pedziwiatr, D. Extracellular Microvesicles (ExMVs) in Cell to Cell Communication: A Role of Telocytes. Adv. Exp. Med. Biol. 2016, 913, 41–49. [Google Scholar] [CrossRef]
- Vannucchi, M.G. The Telocytes: Ten Years after Their Introduction in the Scientific Literature. An Update on Their Morphology, Distribution, and Potential Roles in the Gut. Int. J. Mol. Sci. 2020, 21, 4478. [Google Scholar] [CrossRef]
- Milia, A.F.; Ruffo, M.; Manetti, M.; Rosa, I.; Conte, D.; Fazi, M.; Messerini, L.; Ibba-Manneschi, L. Telocytes in Crohn’s Disease. J. Cell. Mol. Med. 2013, 17, 1525–1536. [Google Scholar] [CrossRef]
- Zheng, Y.; Cretoiu, D.; Yan, G.; Cretoiu, S.M.; Popescu, L.M.; Fang, H.; Wang, X. Protein profiling of human lung telocytes and microvascular endothelial cells using iTRAQ quantitative proteomics. J. Cell. Mol. Med. 2014, 18, 1035–1059. [Google Scholar] [CrossRef] [PubMed]
- Foong, D.; Zhou, J.; Zarrouk, A.; Ho, V.; O’Connor, M.D. Understanding the Biology of Human Interstitial Cells of Cajal in Gastrointestinal Motility. Int. J. Mol. Sci. 2020, 21, 4540. [Google Scholar] [CrossRef] [PubMed]
- Kaestner, K.H. The Intestinal Stem Cell Niche: A Central Role for Foxl1-Expressing Subepithelial Telocytes. Cell Mol. Gastroenterol. Hepatol. 2019, 8, 111–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shoshkes-Carmel, M.; Wang, Y.J.; Wangensteen, K.J.; Tóth, B.; Kondo, A.; Massasa, E.E.; Itzkovitz, S.; Kaestner, K.H. Subepithelial telo-cytes are an important source of Wnts that supports intestinal crypts. Nature 2018, 557, 242–246, Erratum in Nature 2018, 560, E29. [Google Scholar] [CrossRef]
- Wang, X.; Cretoiu, D. (Eds.) Telocytes; Springer: Singapore, 2016; Volume 913. [Google Scholar] [CrossRef]
- Bahar, H.K.; Massalha, H.; Zwick, R.K.; Moor, A.E.; Castillo-Azofeifa, D.; Rozenberg, M.; Farack, L.; Egozi, A.; Miller, D.R.; Aver-bukh, I.; et al. Lgr5+ telocytes are a signaling source at the intestinal villus tip. Nat. Commun 2020, 11, 1936. [Google Scholar] [CrossRef] [Green Version]
- Gersemann, M.; Wehkamp, J.; Fellermann, K.; Stange, E.F. Crohn’s disease-Defect in innate defence. World. J. Gastroenterol. 2008, 14, 5499–5503. [Google Scholar] [CrossRef]
- Koch, S.; Capaldo, C.T.; Hilgarth, R.S.; Fournier, B.; Parkos, C.A.; Nusrat, A. Protein kinase CK2 is a critical regulator of epithelial homeostasis in chronic intestinal inflammation. Mucosal Immunol. 2013, 6, 136–145. [Google Scholar] [CrossRef]
- Hashitani, H.; Lang, R.J. Functions of ICC-like cells in the urinary tract and male genital organs. J. Cell. Mol. Med. 2010, 14, 1199–1211. [Google Scholar] [CrossRef]
- Iatsyna, O.I.; Vernygorodskyi, S.V.; Kostyev, F.I. Morphological analysis of interstitial Cajal cells and mast cells in experimental hyperactivity bladder and stress incontinence under influence of pharmacocorrection. Rep. Morphol. 2018, 24, 5–13. [Google Scholar] [CrossRef]
- Banciu, D.D.; Cretoiu, D.; Cretoiu, S.; Banciu, A. Telocytes involvement in recovery after myocardial infarction. J. Shanghai Univ. 2017, 23, 155–160. [Google Scholar] [CrossRef]
- Park, K.S.; Cho, K.B.; Hwang, I.S.; Park, J.H.; Jang, B.I.; Kim, K.O.; Jeon, S.W.; Kim, E.S.; Park, C.S.; Kwon, J.G. Characterization of smooth muscle, enteric nerve, interstitial cells of Cajal, and fibroblast-like cells in the gastric musculature of patients with diabetes mellitus. World J. Gastroenterol. 2016, 22, 10131–10139. [Google Scholar] [CrossRef] [PubMed]
- Manetti, M.; Rosa, I.; Messerini, L.; Ibba-Manneschi, L. Telocytes are reduced during fibrotic remodelling of the colonic wall in ulcerative colitis. J. Cell. Mol. Med. 2015, 19, 62–73. [Google Scholar] [CrossRef] [PubMed]
- Barker, N.; van Es, J.H.; Kuipers, J.; Kujala, P.; van den Born, M.; Cozijnsen, M.; Haegebarth, A.; Korving, J.; Begthel, H.; Peters, P.J.; et al. Identification of stem cells in small intestine and colon by marker gene Lgr5. Nature 2007, 449, 1003–1007. [Google Scholar] [CrossRef] [PubMed]
- Smythies, J.; Edelstein, L. Telocytes, exosomes, gap junctions and the cytoskeleton: The makings of a primitive nervous system? Front. Cell. Neurosci. 2014, 7, 278. [Google Scholar] [CrossRef] [Green Version]
- Mischopoulou, M.; D’Ambrosio, M.; Bigagli, E.; Luceri, C.; Farrugia, G.; Cipriani, G. Role of Macrophages and Mast Cells as Key Players in the Maintenance of Gastrointestinal Smooth Muscle Homeostasis and Disease. Cell. Mol. Gastroenterol. Hepatol. 2022, 13, 1849–1862. [Google Scholar] [CrossRef]
- Xu, X.; Weksler-Zangen, S.; Pikarsky, A.; Pappo, O.; Wengrower, D.; Bischoff, S.C.; Pines, M.; Rivkind, A.; Goldin, E.; Levi-Schaffer, F. Mast Cells Involvement in the Inflammation and Fibrosis Development of the TNBS-Induced Rat Model of Colitis. Scand. J. Gastroenterol. 2002, 37, 330–337. [Google Scholar] [CrossRef]
- Rosa, I.; Taverna, C.; Novelli, L.; Marini, M.; Ibba-Manneschi, L.; Manetti, M. Telocytes Constitute a Widespread Interstitial Meshwork in the Lamina Propria and Underlying Striated Muscle of Human Tongue. Sci. Rep. 2019, 9, 5858. [Google Scholar] [CrossRef] [Green Version]
- He, X.; Wu, K.; Ogino, S.; Giovannucci, E.L.; Chan, A.T.; Song, M. Association between Risk Factors for Colorectal Cancer and Risk of Serrated Polyps and Conventional Adenomas. Gastroenterology 2018, 155, 355–373.e18. [Google Scholar] [CrossRef]
- Ibba-Manneschi, L.; Rosa, I.; Manetti, M. Telocytes in Chronic Inflammatory and Fibrotic Diseases. In Telocytes: Advances in Experimental Medicine and Biology; Wang, X., Cretoiu, D., Eds.; Springer: Singapore, 2016; Volume 913. [Google Scholar] [CrossRef]
- Moparthi, L.; Koch, S. Wnt signaling in intestinal inflammation. Differentiation 2019, 108, 24–32. [Google Scholar] [CrossRef]
- Kinchen, J.; Chen, H.H.; Parikh, K.; Antanaviciute, A.; Jagielowicz, M.; Fawkner-Corbett, D.; Ashley, N.; Cubitt, L.; Mellado-Gomez, E.; Attar, M.; et al. Structural Remodeling of the Human Colonic Mesenchyme in Inflammatory Bowel Disease. Cell 2018, 175, 372–386.e17. [Google Scholar] [CrossRef] [Green Version]
- Harhun, M.I.; Gordienko, D.V.; Povstyan, O.V.; Moss, R.F.; Bolton, T.B. Function of Interstitial Cells of Cajal in the Rabbit Portal Vein. Circ. Res. 2004, 95, 619–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Y.; Chen, X.; Qian, M.; Zhang, M.; Zhang, D.; Bai, C.; Wang, Q.; Wang, X. Human Lung Telocytes Could Promote the Proliferation and Angiogenesis of Human Pulmonary Microvascular Endothelial Cells in Vitro. Mol. Cell. Ther. 2014, 2, 3. [Google Scholar] [CrossRef] [Green Version]
- Condrat, C.E.; Barbu, M.G.; Thompson, D.C.; Dănilă, C.A.; Boboc, A.E.; Suciu, N.; Crețoiu, D.; Voinea, S.C. Roles and Distribution of Telocytes in Tissue Organization in Health and Disease. In Tissue Barriers in Disease, Injury and Regeneration; Elsevier: Amsterdam, The Netherlands, 2021; pp. 1–41. [Google Scholar] [CrossRef]
- Vannucchi, M.G.; Traini, C.; Manetti, M.; Ibba-Manneschi, L.; Faussone-Pellegrini, M.S. Telocytes express PDGFRα in the human gastrointestinal tract. J. Cell. Mol. Med. 2013, 17, 1099–1108. [Google Scholar] [CrossRef] [PubMed]
- Barmpatsalou, V.; Dubbelboer, I.R.; Rodler, A.; Jacobson, M.; Karlsson, E.; Pedersen, B.L.; Bergström, C. Physiological properties, composition and structural profiling of porcine gastrointestinal mucus. Eur. J. Pharm. Biopharm. 2021, 169, 156–167. [Google Scholar] [CrossRef] [PubMed]
- Song, D.; Tang, L.; Huang, J.; Wang, L.; Zeng, T.; Wang, X. Roles of transforming growth factor-β and phosphatidylinositol 3-kinase isoforms in integrin β1-mediated bio-behaviors of mouse lung telocytes. J. Transl. Med. 2019, 17, 431. [Google Scholar] [CrossRef] [PubMed]
- Araujo, A.P.; Diniz, L.P.; Eller, C.M.; de Matos, B.G.; Martinez, R.; Gomes, F.C. Effects of Transforming Growth Factor Beta 1 in Cerebellar Development: Role in Synapse Formation. Front. Cell. Neurosci. 2016, 10, 104. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Zheng, H.; Zhu, H.Y.; Hu, S.; Wang, S.; Jiang, X.; Xu, G.Y. Acute Effects of Transforming Growth Factor-β1 on Neuronal Excitability and Involvement in the Pain of Rats with Chronic Pancreatitis. J. Neurogastroenterol. Motil. 2016, 22, 333–343. [Google Scholar] [CrossRef] [Green Version]
- Diniz, L.P.; Matias, I.; Siqueira, M.; Stipursky, J.; Gomes, F.C.A. Astrocytes and the TGF-Β1 Pathway in the Healthy and Diseased Brain: A Double-Edged Sword. Mol. Neurobiol. 2019, 56, 4653–4679. [Google Scholar] [CrossRef]
- Matejuk, A.; Vandenbark, A.A.; Offner, H. Cross-Talk of the CNS with Immune Cells and Functions in Health and Disease. Front. Neurol. 2021, 12, 672455. [Google Scholar] [CrossRef]
- Sanders, K.M.; Ward, S.M.; Koh, S.D. Interstitial cells: Regulators of smooth muscle function. Physiol. Rev. 2014, 94, 859–907. [Google Scholar] [CrossRef]
- Citri, A.; Malenka, R.C. Synaptic plasticity: Multiple forms, functions, and mechanisms. Neuropsychopharmacology 2008, 33, 18–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chernikova, D.A.; Madan, J.C.; Housman, M.L.; Zain-Ul-Abideen, M.; Lundgren, S.N.; Morrison, H.G.; Sogin, M.L.; Williams, S.M.; Moore, J.H.; Karagas, M.R.; et al. The premature infant gut microbiome during the first 6 weeks of life differs based on gestational maturity at birth. Pediatr. Res. 2018, 84, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Skonieczna-Żydecka, K.; Janda, K.; Kaczmarczyk, M.; Marlicz, W.; Łoniewski, I.; Łoniewska, B. The Effect of Probiotics on Symptoms, Gut Microbiota and Inflammatory Markers in Infantile Colic: A Systematic Review, Meta-Analysis and Meta-Regression of Randomized Controlled Trials. J. Clin. Med. 2020, 9, 999. [Google Scholar] [CrossRef] [Green Version]
- Krulwich, T.A.; Sachs, G.; Padan, E. Molecular aspects of bacterial pH sensing and homeostasis. Nature reviews. Microbiology 2011, 9, 330–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goers Sweeney, E.; Henderson, J.N.; Goers, J.; Wreden, C.; Hicks, K.G.; Foster, J.K.; Parthasarathy, R.; Remington, S.J.; Guillemin, K. Structure and proposed mechanism for the pH-sensing Helicobacter pylori chemoreceptor TlpB. Structure 2012, 20, 1177–1188. [Google Scholar] [CrossRef] [Green Version]
- Soliman, S.A. Telocytes are major constituents of the angiogenic apparatus. Sci. Rep. 2021, 11, 5775. [Google Scholar] [CrossRef] [PubMed]
- Gelfand, A.A.; Goadsby, P.J.; Allen, I.E. The relationship between migraine and infant colic: A systematic review and meta-analysis. Cephalalgia Int. J. Headache 2015, 35, 63–72. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Banciu, D.D.; Crețoiu, D.; Crețoiu, S.M.; Banciu, A.; Popa, D.; David, R.; Berghea-Neamtu, C.S.; Cipaian, C.R.; Negrea, M.O.; Gheonea, M.; et al. Telocytes’ Role in Modulating Gut Motility Function and Development: Medical Hypotheses and Literature Review. Int. J. Mol. Sci. 2022, 23, 7017. https://doi.org/10.3390/ijms23137017
Banciu DD, Crețoiu D, Crețoiu SM, Banciu A, Popa D, David R, Berghea-Neamtu CS, Cipaian CR, Negrea MO, Gheonea M, et al. Telocytes’ Role in Modulating Gut Motility Function and Development: Medical Hypotheses and Literature Review. International Journal of Molecular Sciences. 2022; 23(13):7017. https://doi.org/10.3390/ijms23137017
Chicago/Turabian StyleBanciu, Daniel Dumitru, Dragoș Crețoiu, Sanda Maria Crețoiu, Adela Banciu, Daniel Popa, Rodica David, Cristian Stefan Berghea-Neamtu, Calin Remus Cipaian, Mihai Octavian Negrea, Mihaela Gheonea, and et al. 2022. "Telocytes’ Role in Modulating Gut Motility Function and Development: Medical Hypotheses and Literature Review" International Journal of Molecular Sciences 23, no. 13: 7017. https://doi.org/10.3390/ijms23137017
APA StyleBanciu, D. D., Crețoiu, D., Crețoiu, S. M., Banciu, A., Popa, D., David, R., Berghea-Neamtu, C. S., Cipaian, C. R., Negrea, M. O., Gheonea, M., & Neamtu, B. (2022). Telocytes’ Role in Modulating Gut Motility Function and Development: Medical Hypotheses and Literature Review. International Journal of Molecular Sciences, 23(13), 7017. https://doi.org/10.3390/ijms23137017